p53, ROS and senescence in the control of aging
Abstract
Abstract: In addition to its function as a tumour suppressor, p53 is also involved in an increasing number of pathology associated with aging. Several activities of p53 appear contribute to its role in aging; one function that might be particularly relevant in this context is the regulation of senescence. The control of ROS and senescence by p53 may help to explain how p53 can function to both restrain and promote aging.
p53
functions as a longevity assurance gene (by virtue of its strong tumor
suppressor activity) and a regulator of aging. In several mouse models,
persistent low-level activation of p53, either through deregulated expression
of p53 itself or in response to constitutive stress like DNA damage/telomere
erosion, leads to premature aging [1,2]. However,
mice with normal basal p53 levels that have been engineered to a show a
heightened ability to mount a p53 response show a very strong resistance to
tumourigenesis without evidence of premature aging [3]. Indeed, in
several of these models a decreased level of aging related damage is observed,
indicating that p53 may also help to promote longevity. The control of aging
reflects numerous activities of p53, including the modulation of the IGFR
pathway through interplay between full-length p53 and N-terminally truncated
splice variants of p53 [4] and the
ability of p53 to restrict stem cell function [5]. p53 is also
a key regulator of senescence, a central stress response that plays an
important role in tumour suppression, but may also help to promote cancer
development by inducing an inflammatory response [6]. The ability
to control senescence is consistent with p53's function in restraining cancer
development, but can the mechanisms through which p53 regulates senescence also
contribute to the control of aging? Induction of senescence by p53 is
associated with the regulation of p53-dependent genes that can participate in
cell cycle arrest. While depletion of these components can impact senescence
induction - supporting their role in mediating this response - the inhibition
of cell cycle progression alone does not explain how this arrest can be turned
into the definitive and permanent proliferation block that is characteristic of
senescence. Furthermore, despite the clear documentation of p53's ability to
induce senescence, more recent evidence shows that p53 can also function to
inhibit senescence while promoting cell cycle arrest [7]. So how can
p53 both suppress and promote senescence? An important component of this may be the ability
of p53 to control cell growth and metabolic stressthrough
different pathways, including the regulation of ROS levels and the activity of
mTOR (Figure 1). The ability of p53 to promote ROS production has been shown to
participate in the induction of apoptosis by p53 [8]. But ROS are also known to be critical for senescence
[9] and the p53 target genes that increase ROS may
also play an important role in senescence induction. However, p53 also promotes
the expression of a number of antioxidant genes, accounting for p53's ability
to control oxidative stress in cells and mice [10]. So p53's ability to decrease and increase
oxidative stress likely contributes to its dual effect on senescence. Another
factor that influences the outcome to p53 activation is mTOR. While mTOR is
normally associated with cell growth, activation of mTOR can contribute to and
be essential for certain types of senescence [11,12], and the maintenance of mTOR
signalling under conditions of cell cycle arrest leads to senescence in
cultured cells [13]. p53 inhibits the mTOR pathway at several levels [14], contributing to the anti-senescence activity of
p53 [15]. Furthermore, mTOR can be activated by ROS [16], so p53's antioxidant activities may reinforce the
dampening of mTOR and senescence (Figure 1).
One of the main
responses to mTOR inhibition is the induction of autophagy, a response that
allows survival under conditions of nutrient deprivation. There are several
possible links between autophagy and senescence. Inhibition of autophagy
results in the accumulation of protein aggregates, ER stress and mitochondrial
dysfunction, each of which could promote senescence. However, other studies
suggest that autophagy may be required for an efficient senescence response [17]. In either
case, the ability of p53 to both enhance and inhibit autophagy [18] provides a
further mechanism for the modulation of senescence.
The
activity of p53 is regulated through many mechanisms, but of particular
interest with respect to the control of senescence and aging is a role for the
histone deacetylase Sirt1, whose expression is strongly down regulated in
senescent cells [19]. In
contrast nutrient deprivation, which inhibits mTOR and can impede cellular
senescence [13], has been
shown to increase Sirt1 levels [20]. One way in
which Sirt1 functions is to deacetylate p53, modulating p53 activity and
impeding decreasing senesence [21]. Deactylation inhibits p53's ability to
transcriptionally activate some, but not all,
target genes - including those involved in apoptosis induction, ROS production [22,23], and
presumably also senescence (Figure 1). The presence of a chronic DNA damage
response (as may be seen in cancer cells), which is linked to the induction of
senescence [24], can directly
increase p53 acetylation by inducing the phosphorylation of the N-terminus of
p53 and so promoting the interaction with the acetyl transferases CBP/p300. Mouse
models have shown that expression of phosphorylation resistant p53 inhibits the
induction of senescence [25], while
cells harbouring p53 with acetyl-mimicking mutations of the last seven lysine
residues have an accelerated entry into senescence and are very resistant to senescence bypass [26], although the cell cycle arrest
response in these cells remains normal. Phosphorylation and acetylation of p53
is also seen to be important during Ras-induced or replicative senescence [27,28]. Under these circumstances, it
would seem that deacetylation of p53 by Sirt1 impedes the induction of
senescence, as well as apoptosis. Taken together there is good evidence that
acetylation of p53 promotes sense-cence and apoptosis, so inhibitors of the
deacetylation enzymes might be useful drugs for the reactivation of these p53
responses for cancer therapy [29].
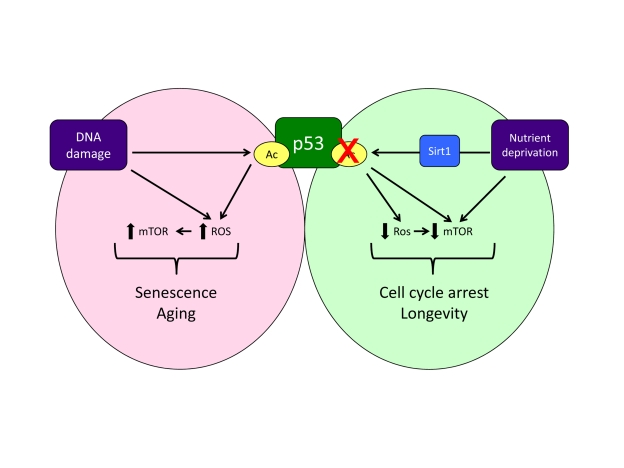
Figure 1. A model of how acetylation, oxidative stress and mTOR activity might influence the response to p53. Note that this model does not account
for all published observations (e.g. reduction of the initial burst of mTOR
activity during oncogene induced senescence [17]) and represents
an oversimplification of these signalling pathways.
Several
of the mechanisms implicated in the induction of senescence by p53 have also
been linked to the regulation of longevity. Induction of mTOR and oxidative
stress - and the complex interplay between them - is associated with aging [16] and Sirt1
is emerging as a key supporter of longevity in many organisms [30]. The
induction of cellular senescence itself may result in loss of tissue renewal
and architecture, organ dysfunction and organismal aging [31], while
autophagy can protect from aging [32]. So it seems
reasonable to propose that p53's ability to influence aging is reflected - at
least in part - by the mechanisms through which p53 controls senescence.But as we have discussed, p53 can promote and impede
both senescence and aging - so which output prevails? The answer is not yet
clear, but one determining factor may be the type or extent of the p53-inducing
stress. Current models suggest that mild or constitutive stress induced by
normal growth and proliferation lead to p53-induced antioxidant and repair
functions, while strong or persistent p53 activity may tip the balance towards
the induction of apoptosis or senescence, thereby favoring aging. The mouse
models also clearly suggest that inappropriate p53 activity promotes aging
while a robust but normally regulated p53 response protects from the aging
process. One prediction of this model is that the persistent stress encountered
in tumors would favor p53-induced senescence over a more transient cell cycle
arrest - and indeed the activation of p53 in established tumors has been shown
to promote senescence in some tissue types [33].
p53
is emerging as an important, but complex, player in the regulation of
senescence and longevity. The ability of p53 to both activate and inhibit
senescence is reflected in the ability to promote and inhibit oxidative stress
and autophagy, and the ultimate establishment of senescence or quiescence is
highly dependent on collaborating factors such as mTOR activity or oxidative
stress. Ultimately, these bipolar activities of p53 become manifest in the
contradictory effects on longevity and aging. p53 based cancer therapies may be
rendered more effective by an increased propensity of transformed cells to
undergo senescence, compared to normal cells. However, the idea that p53 can
both promote and prevent aging adds even more spice to the consideration of how
to use drugs that can induce or inhibit p53 activity.
Acknowledgments
We
are very grateful to Peter Adams for his helpful comments and advice.
References
-
1.
Tyner
SD
, Venkatachalam
S
, Choi
J
, Ghebranious
N
and Igelmann
H.
p53 mutant mice that display early ageing-associated phenotypes.
Nature.
2002;
415:
45
-53.
[PubMed]
.
-
2.
Maier
B
, Gluba
W
, Bernier
B
, Turner
T
and Mohammad
K.
Modulation of mammalian life span by the short isoform of p53.
Genes Dev.
2004;
18:
306
-319.
[PubMed]
.
-
3.
Matheu
A
, Maraver
A
, Klatt
P
, Flores
I
and Garcia-Cao
I.
Delayed ageing through damage protection by the Arf/p53 pathway.
Nature.
2007;
448:
375
-379.
[PubMed]
.
-
4.
Pehar
M
, O'Riordan
KJ
, Burns-Cusato
M
, Andrzejewski
ME
and del
Alcazar CG.
Altered longevity-assurance activity of p53:p44 in the mouse causes memory loss, neurodegeneration and premature death.
Aging Cell.
2010;
9:
174
-190.
[PubMed]
.
-
5.
Zhao
T
and Xu
Y.
p53 and stem cells: new developments and new concerns.
Trends Cell Biol.
2010;
20:
170
-175.
[PubMed]
.
-
6.
Coppe
JP
, Desprez
PY
, Krtolica
A
and Campisi
J.
The senescence-associated secretory phenotype: the dark side of tumor suppression.
Annu Rev Pathol.
2010;
5:
99
-118.
[PubMed]
.
-
7.
Demidenko
ZN
, Korotchkina
LG
, Gudkov
AV
and Blagosklonny
MV.
Paradoxical suppression of cellular senescence by p53.
Proc Natl Acad Sci USA.
2010;
107:
9660
-9664.
[PubMed]
.
-
8.
Johnson
TM
, Yu
Z-X
, Ferrans
VJ
, Lowenstein
RA
and Finkel
T.
Reactive oxygen species are downstream mediators of p53-depen-dent apoptosis.
Proc Natl Acad Sci USA.
1996;
93:
11848
-11852.
[PubMed]
.
-
9.
Lu
T
and Finkel
T.
Free radicals and senescence.
Exp Cell Res.
2008;
314:
1918
-1922.
[PubMed]
.
-
10.
Olovnikov
IA
, Kravchenko
JE
and Chumakov
PM.
Homeostatic functions of the p53 tumor suppressor: regulation of energy metabolism and antioxidant defense.
Semin Cancer Biol.
2009;
19:
32
-41.
[PubMed]
.
-
11.
Nardella
C
, Chen
Z
, Salmena
L
, Carracedo
A
and Alimonti
A.
Aberrant Rheb-mediated mTORC1 activation and Pten haploinsufficiency are cooperative oncogenic events.
Genes Dev.
2008;
22:
2172
-2177.
[PubMed]
.
-
12.
Alimonti
A
, Nardella
C
, Chen
Z
, Clohessy
JG
and Carracedo
A.
A novel type of cellular senescence that can be enhanced in mouse models and human tumor xenografts to suppress prostate tumorigenesis.
J Clin Invest.
2010;
120:
681
-693.
[PubMed]
.
-
13.
Demidenko
ZN
and Blagosklonny
MV.
Growth stimulation leads to cellular senescence when the cell cycle is blocked.
Cell Cycle.
2008;
7:
3355
-3361.
[PubMed]
.
-
14.
Feng
Z
and Levine
AJ.
The regulation of energy metabolism and the IGF-1/mTOR pathways by the p53 protein.
Trends Cell Biol.
2010;
20:
427
-434.
[PubMed]
.
-
15.
Korotchkina
LG
, Leontieva
OV
, Bukreeva
EI
, Demidenko
ZN
and Gudkov
AV.
The choice between p53-induced senescence and quiescence is determined in part by the mTOR pathway.
Aging.
2010;
2:
344
-352.
[PubMed]
.
-
16.
Blagosklonny
MV
Aging: ROS or TOR.
Cell Cycle.
2008;
7:
3344
-3354.
[PubMed]
.
-
17.
Young
AR
, Narita
M
, Ferreira
M
, Kirschner
K
and Sadaie
M.
Autophagy mediates the mitotic senescence transition.
Genes Dev.
2009;
23:
798
-803.
[PubMed]
.
-
18.
Maiuri
MC
, Galluzzi
L
, Morselli
E
, Kepp
O
and Malik
SA.
Autophagy regulation by p53.
Curr Opin Cell Biol.
2010;
22:
181
-185.
[PubMed]
.
-
19.
Sasaki
T
, Maier
B
, Bartke
A
and Scrable
H.
Progressive loss of SIRT1 with cell cycle withdrawal.
Aging Cell.
2006;
5:
413
-422.
[PubMed]
.
-
20.
Kanfi
Y
, Peshti
V
, Gozlan
YM
, Rathaus
M
and Gil
R.
Regulation of SIRT1 protein levels by nutrient availability.
FEBS Lett.
2008;
582:
2417
-2423.
[PubMed]
.
-
21.
Langley
E
, Pearson
M
, Faretta
M
, Bauer
UM
and Frye
RA.
Human SIR2 deacetylates p53 and antagonizes PML/p53-induced cellular senescence.
Embo J.
2002;
21:
2383
-2396.
[PubMed]
.
-
22.
Luo
J
, Nikolaev
AY
, Imai
S
, Chen
D
and Su
F.
Negative control of p53 by Sir2alpha promotes cell survival under stress.
Cell.
2001;
107:
137
-148.
[PubMed]
.
-
23.
Vaziri
H
, Dessain
SK
, Ng
Eaton E
, Imai
SI
and Frye
RA.
hSIR2(SIRT1) functions as an NAD-dependent p53 deacetylase.
Cell.
2001;
107:
149
-159.
[PubMed]
.
-
24.
Pospelova
TV
, Demidenko
ZN
, Bukreeva
EI
, Pospelov
VA
and Gudkov
AV.
Pseudo-DNA damage response in senescent cells.
Cell Cycle.
2009;
8:
4112
-4118.
[PubMed]
.
-
25.
Armata
HL
, Garlick
DS
and Sluss
HK.
The ataxia telangiectasia-mutated target site Ser18 is required for p53-mediated tumor suppression.
Cancer Res.
2007;
67:
11696
-11703.
[PubMed]
.
-
26.
Krummel
KA
, Lee
CJ
, Toledo
F
and Wahl
GM.
The C-terminal lysines fine-tune p53 stress response in a mouse model but are not required for stability control or transactivation.
Proc Natl Acad Sci USA.
2005;
102:
10188
-10193.
[PubMed]
.
-
27.
Pedeux
R
, Sengupta
S
, Shen
JC
, Demidov
ON
and Saito
S.
ING2 regulates the onset of replicative senescence by induction of p300-dependent p53 acetylation.
Mol Cell Biol.
2005;
25:
6639
-6648.
[PubMed]
.
-
28.
Pearson
M
, Carbone
R
, Sebastiani
C
, Cioce
M
and Fagioli
M.
PML regulates p53 acetylation and premature senescence induced by oncogenic ras.
Nature.
2000;
406:
207
-210.
[PubMed]
.
-
29.
Lain
S
, Hollick
JJ
, Campbell
J
, Staples
OD
and Higgins
M.
Discovery, in vivo activity, and mechanism of action of a small-molecule p53 activator.
Cancer Cell.
2008;
13:
454
-463.
[PubMed]
.
-
30.
Finkel
T
, Deng
CX
and Mostoslavsky
R.
Recent progress in the biology and physiology of sirtuins.
Nature.
2009;
460:
587
-591.
[PubMed]
.
-
31.
Campisi
J
Senescent cells, tumor suppression, and organismal aging: good citizens, bad neighbors.
Cell.
2005;
120:
513
-522.
[PubMed]
.
-
32.
Salminen
A
and Kaarniranta
K.
Regulation of the aging process by autophagy.
Trends Mol Med.
2009;
15:
217
-224.
[PubMed]
.
-
33.
Xue
W
, Zender
L
, Miething
C
, Dickins
RA
and Hernando
E.
Senescence and tumour clearance is triggered by p53 restoration in murine liver carcinomas.
Nature.
2007;
445:
656
-660.
[PubMed]
.