Single-cell transcriptomics identifies senescence-associated secretory phenotype (SASP) features of testicular aging in human
Abstract
The male reproductive system experiences degradation with age, predominantly impacting the testes. Testicular aging can result in failure to produce physiological testosterone levels, normal sperm concentrations, or both. However, we cannot predict the onset of testicular aging in advance. Using single-cell RNA sequencing (scRNA-seq) from Gene Expression Omnibus (GEO) database, we conducted cell-cell communication network of human testis between older and young group, indicating Leydig cells’ potential role in spermatogenesis microenvironment of aging testis. And we depicted the senescence-Associated Secretory Phenotype (SASP) features of aging testis by identifying differentially expressed senescence-associated secretory phenotype (SASP)-related genes between two group. Notably, IGFBP7 mainly expressed in Leydig cells of those differentially expressed SASP-related genes in aging testis. Furthermore, IGFBP7 protein located in the interstitial compartment of older mice confirmed by immunofluorescence and highly expressed in both human seminal plasma and mouse testis in the older group confirmed through Western blot. Together, our findings suggest that IGFBP7 may be a new biomarker of testicular aging.
Introduction
Testicular aging, leading to a decrease in the levels of testosterone, decline in sperm quality, a decline in fertility, dysfunction body, mental and psychological disorders. To treat the decline of testosterone caused by testicular aging, testosterone replacement therapy (TRT) has been proposed, but it has great side effects including prostate cancers, uncontrolled congestive heart failure, severe lower-urinary-tract symptoms, and erythrocytosis [1]. In addition, male infertility is a growing concern due to sharp decline of sperm concentration and total sperm count worldwide especially in the aging male [2, 3]. More importantly, several research have reported that increasing male age is significantly associated with bad reproductive outcome [4–6]. Despite aging being a universal, multifactorial, progressive, and irreversible process, there have been numerous attempts to delay or prevent aging in general. In the modern societies where paternal age is on the rise, research on testicular aging is increasing. Moreover, the age-related decline in testicular function also has a broader impact on overall health [7–9]. However, there has been limited consensus on the process of testicular aging [10, 11]. So, it is an urgency to find out marker of testicular aging.
Both germ cells and the spermatogenesis microenvironment play an important role in spermatogenesis. It has been observed that the testis of older man is more likely to have decreased in germ cell and malfunction in spermatogenesis microenvironment [12, 13]. For the spermatogenesis microenvironment, more and more studies pay attention to the role of Sertoli and Leydig cells [14, 15]. Sertoli and Leydig cells show significant abnormalities with age, such as decreased number, morphological variations, organelle aging, abnormal hormone secretion, and blood–testicular barrier defects. Cellular senescence is a cellular stress response triggered by molecular damage, such as replication failure, activation of abnormal oncogenes, or chemotherapy treatment.
The senescence-associated secretory phenotype (SASP) is known as secreted by senescent cells which have the therapeutic potential to aging [16, 17]. Senolytic is a new approach to kill senescent cells selectively, SASP-centered approaches are emerging as alternatives to target senescence-associated diseases. For now, there are few studies on SASP in aging testis. Herein, we downloaded single-cell RNA sequencing (scRNA-seq) from Gene Expression Omnibus (GEO) database. According to age stratification, patients were divided into older and young groups. And then, we analyzed the differences in testicular cell-cell communication networks between older and young groups, recognized the differential expressing SASP-related genes and provided a foundation for further exploration of treating testicular aging.
Materials and Methods
Single-cell data acquisition and preprocessing
All raw data files were downloaded from Gene Expression Omnibus (GEO) database, including NCBI accession number GSE120508 (Donor_1, Donor_2, Donor_3), GSE215754 (Y1, HA1, HA2), GSE153947 (Normal_1, Normal_2, Normal_3) and GSE182786 (Young_1, Young_2, Young_3, Young_4, Older_1, Older_2, Older_3, Older_4, Older_5, Older_6, Older_7, Older_8). Participants were categorized into two groups: the older group and the young group. We employed an age-based classification, with individuals aged over 60 assigned to the older group, and those under 60 categorized as the young group. The Cell Ranger (v.7.0.1, 10x Genomics) was used to demultiplex the FASTQ reads align raw reads to the human reference genome (GRCh38, 10x Genomics), and generated the gene-cell unique molecular identifier (UMI) matrix for each sample. R package Seurat (v.4.3.0.1) was used to processes the count matrices. First, we filtered cells with high mitochondrial gene expression (>8%) by fitting the expression of mitochondrial genes to a normal distribution and applying a false discovery rate (FDR) threshold of <0.01. Second, we removed cells with a low number of genes detected, specifically those with fewer than 1000 genes. This step helped eliminate low-quality cells that may have been subject to technical artifacts. Third, we used the DoubleFinder (v. 2.0.3) to identify potential doublets and used a cutoff of the 92.5th percentile for the doublet score. Cells exceeding this threshold were considered potential doublets and were subsequently removed from the analysis. Following these quality control steps, we obtained a final dataset comprising 86, 626 single cells for the following analysis.
We used the harmony (v. 2.0.3) to integrate the individual samples and identify common sources of variation. The data were normalized by using “NormalizeData” function, ensuring that the number of UMIs in each cell was equal to the median UMI across the entire dataset. Additionally, a log-transformed was applied to the data. “FindVariableFeatures” function was used to identify the top 2,000 highly variable genes. Subsequently, the gene expression matrix was scale and center using the “ScaleData” function. For dimensionality reduction and visualization of the data, we performed principal component analysis (PCA) using the RunPCA function based on the highly variable genes. The resulting top 20 principal components were then used for uniform manifold approximation and projection (UMAP) analysis to visualize the clusters in 2D space. Finally, clustering was performed using the Leiden community detection algorithm, which allowed us to identify distinct cell populations based on their transcriptional profiles.
Annotation of cell clusters
To assign the major cell types for each cluster, we followed a two-step approach. First, we performed differential expression analysis using “FindAllMarkers” function with default parameters. By comparing each cluster against all other clusters, we identified genes that showed significant differential expression. In the next step, we aimed to assign cell types to the clusters by examining the presence of known cell-type-specific genes among the top rank of differentially expressed genes in each cluster. By referencing established knowledge about marker genes for specific cell types, we determined the most likely cell type for each cluster based on these differentially expressed genes.
Cell-cell communication analysis by CellCall
We conducted cell-cell communication analysis using CellCall [18] (v1.0.7). To ensure data integrity, we performed quality inspection and normalization on both the young and older datasets. Following the official workflow and default parameter settings, we loaded the young and older group datasets separately into CellCall. To identify potential ligand-receptor interactions between cell types in the young and older group datasets, developer of CellCall applied multiple databases including the NATMI [19], Cellinker [20], CellTalkDB [21], CellChat [22] and STRING v11 databases [23] to conduct human L–R interactions. By leveraging this R package, we screened for specific interactions and examined their potential significance in the context of young and older samples.
Differentially expressed gene analysis
To evaluate the differentially expressed SASP-related genes between young and older group, first, we applied utilized the “FindMarkers” function from the Seurst package (v4.3.0.1). SASP-related genes were downloaded from AgeAnno database [24]. In the next step, we intersected the differentially expressed genes and SASP related genes.
Animals
Testes of 6 mice (3 C57/BL6 mice of 3-month-old and 3 C57/BL6 mice of 20-month-old) were gathered in this experiment. All the mice were bought from the Laboratory Animal Center of Sun Yat-sen University. All experimental procedures involving animals were approved by the Institutional Animal Care and Use Committee of Sun Yat-sen University.
Western blot analysis
Total testis protein was extracted using RIPA lysis buffer (CW2333, CWBIO, China) containing proteinase and phosphatase inhibitors. Western blot analysis was conducted as previously described [25]. The primary antibodies included anti-IGFBP7 (#36930, 1:1000; SAB, USA) and anti-beta Actin Antibody (AF7018, 1:5000; Affinity, USA) antibodies. Beta-ACTIN was used as the control.
Semen collection
Human seminal plasmas were obtained from healthy donors that announced no reproductive system diseases history. All donors have signed an approval consent form.
ELISA
Human seminal plasmas were collected and diluted 100x and IGFBP7 level were detected through Human Insulin-like growth factor-binding protein 7 ELISA Kit (#EK4953, SAB, USA) in accordance with the manufacturer’s instructions.
Statistical analysis
Statistical analysis was performed using the R software (v.4.2.2). For all variables, we applied one-tailed Wilcoxon rank-sum test to assess the differences between groups. A significance threshold of p < 0.05 was used to determine statistical significance. Results that met this criterion were considered to have a statistically significant difference.
Data availability
The RNA-seq matrix data used in this study were available in NCBI with accession number GSE120508 (Donor_1, Donor_2, Donor_3), GSE215754 (Y1, HA1, HA2), GSE153947 (Normal_1, Normal_2, Normal_3) and GSE182786 (Young_1, Young_2, Young_3, Young_4, Older_1, Older_2, Older_3, Older_4, Older_5, Older_6, Older_7, Older_8).
Results
scRNA-seq analysis of human testis samples
We downloaded FASTQ files from GEO database. A total of 23 individuals scRNA-seq was obtained, including eight individuals in older group and 14 individuals in young group (Supplementary Table 1). After quality control, we obtained a total of 86, 626 high-quality single cells by scRNA-seq (Figure 1A). Based on the known markers (Figure 1A and Supplementary Figure 1A), we identified eight main clusters: Leydig cell, Sertoli cell, macrophage, peritubular myoid cells (PTM), endothelial cells, spermatogonia, spermatocyte, spermatid. Not surprisingly, germ cells significantly decreased in older samples, especially the spermatogonia. As for somatic cell, the number of PTM and endothelial cells increased remarkable in older group. No significant difference was observed in Macrophages, Leydig cell and Sertoli cell between older and young groups (Figure 1B and Supplementary Figure 1B, 1C). Together, these data indicated that we established a comprehensive single-cell transcriptomic of human testis, tissue-structure associated cell types inferred by scRNA-seq provided a proxy for dissecting molecular changes of tissue architecture of aging testis.
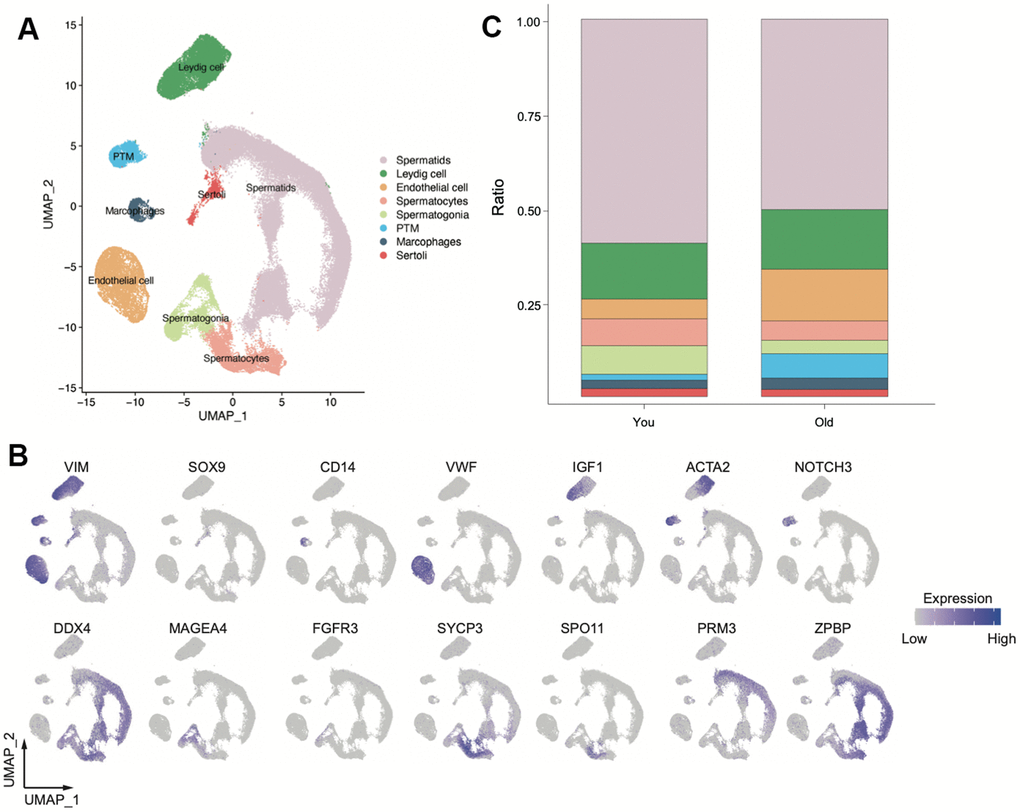
Figure 1. Overview of scRNA-seq transcriptomic profiles from young and old samples (You n=9, Old n=14). (A) Uniform manifold approximation and projection (UMAP) plot showing eight main clusters based on their expression of known markers (B). (C) Bar plot showing the percentage of major cell types for two group.
Leydig cells highly expressed differentially expressed SASP-related gene in aging testis
To further understand the characteristics of spermatogenesis microenvironment in older samples, we conducted cell-cell communication analysis using CellCall (v1.0.7). We found that the ligand-receptor pairs between Leydig cells and other germ cell were reduced in older group (Figure 2A), suggesting that the number of Leydig cells did not change significantly, but they did change functionally. To find out whether SASP plays a role in aging testis, we conducted differentially expressed gene (DEG) analysis between older and young group and labeled the SASP-related genes [24] in volcano plot (Figure 2B). We identified IGFBP7 expression enriched and JUN, FOS and UBB were downregulated in older samples (Supplementary Table 2). Intriguingly, IGFBP7 was highly expressed in Leydig cells, indicating that Leydig cells is a key SASP-spreading cell and may contribute to aging testis in the spermatogenesis microenvironment.
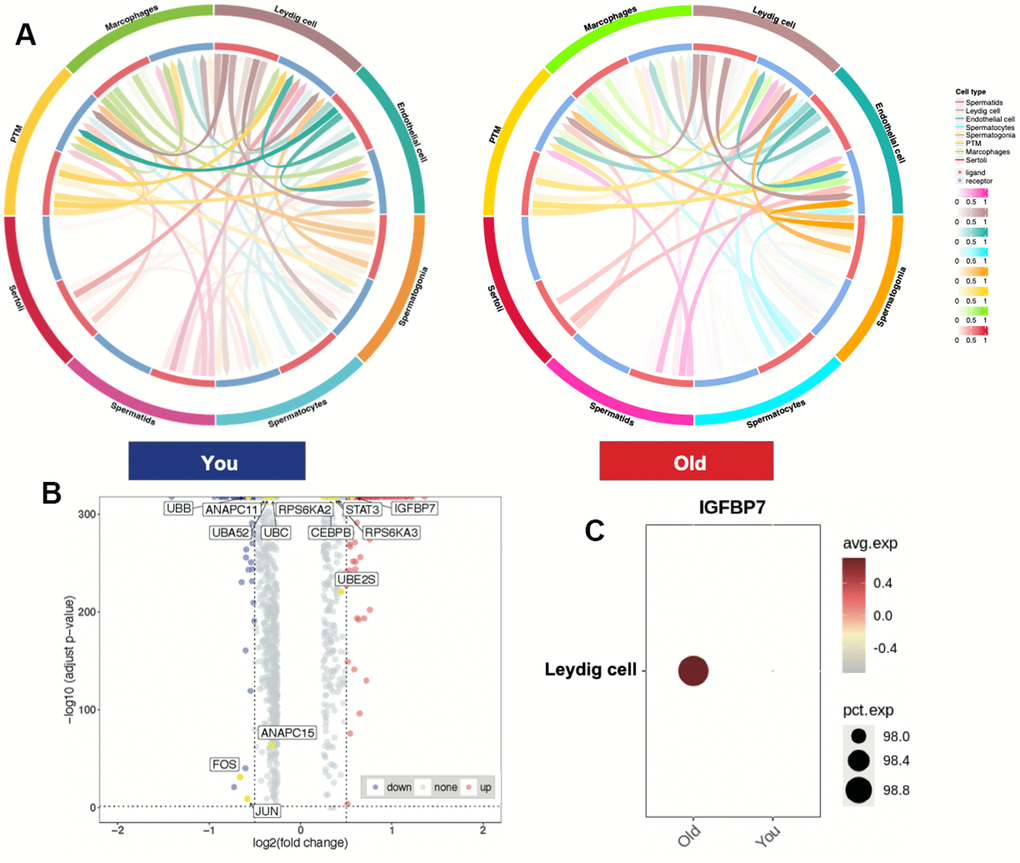
Figure 2. Leydig cells highly expressed differentially expressed SASP-related gene in aging testis. (A) Cell-cell communication between You and Old samples. (B) Volcano plot showing DEGs between You and Old samples, (Wilcoxon, adjusted p-values < 0.05, LogFC > 0.5) (SASP-related genes, yellow). (C) Dot plot showing IGFBP7 expression enriched in older samples.
IGFBP7 protein highly expressed in older samples
Senescent cells produce a series of profibrotic and proinflammatory factors, these factors are also known as SASP. IGFBP7 are important transcriptional inducers of the SASP. IGFBP7, derived from senescent cells, plays a crucial role in inducing senescence in young mesenchymal stem cells. To verify whether SASP such as IGFBP7 are secreted by Leydig cells and travel cross the seminal tubular basement membrane, we collected human seminal plasma from healthy donors and measured IGFBP7 protein levels. By using ELISA, we found that IGFBP7 protein could be detected in seminal plasma, and interestingly, seminal plasma IGFBP7 expression levels were higher in the older group than in the young group (Figure 3A). We also measured the location by immunofluorescence and expression level of IGFBP7 in the testes of C57/BL6 mice by Western blot. We found that IGFBP7 was mainly expressed in testicular interstitial compartment, and the level of IGFBP7 in the testes of mice in the older group was higher than that in the young group (Figure 3B, 3C), indicating that IGFBP7 may be a new biomarker of testicular aging.
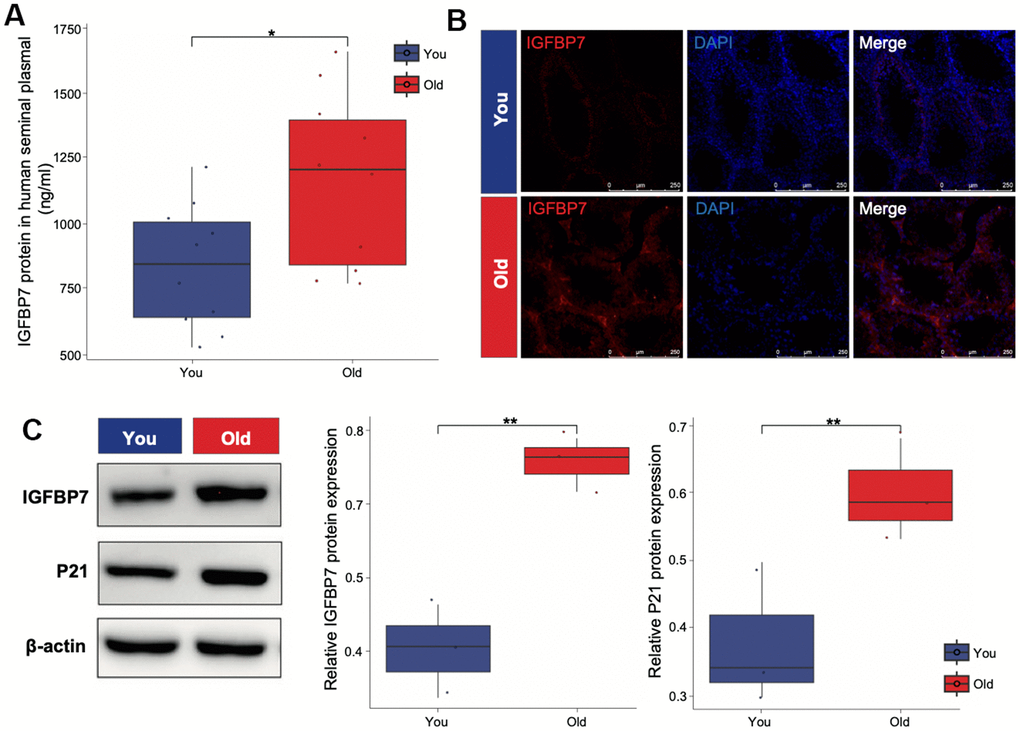
Figure 3. IGFBP7 protein expression of human seminal plasma and mice testis. (A) ELISA was applied to detect the expression of IGFBP7 in human seminal plasma. (B) Immunofluorescence showed the localization of IGFBP7 protein in the interstitial compartment of older mice. Scale bar: 250 μm (C) Western blot was applied to detect the expression of IGFBP7 and the senescence marker P21 in testis of mice. Data depict the mean ± SD; *P < 0.05. **P < 0.01.
Discussion
In this study, we established a comprehensive single-cell transcriptomic of aging testis in human. By conducting the cell-cell communication network with young and older samples, we were able to identify Leydig cells’ potential role in aging testis. The ligand-receptor pairs between Leydig cells and other germ cell were significantly reduced in older samples. In line with that, multiple studies also reported that Leydig cells from older men display malfunction, including lower production of testosterone [13, 26]. So, application of TRT to treat male infertility was mentioned [27]. However, the concern of promoting prostate cancer limits the use of TRT [28]. And the sole unequivocal indication for TRT is as replacement therapy for men with pathological hypogonadism. It does not apply to older infertile men with normal testosterone levels. There is an urging to find out other therapeutic strategies for aging men.
Nie recently generated single-cell transcriptomic sequencing and analysis in testes of young and old human exploring the correlation between testicular aging and elevated body mass index (BMI) and providing a comprehensive examination of functional changes in various cell types within aging testes [13]. However, they primarily focused on delineating functional changes in various cell types within the aging testes, with a notable absence of emphasis on senescence-associated secretory phenotype (SASP) factors in the context of testicular aging.
Cellular senescence promotes tissue remodeling through three sequential processes: a stable proliferative arrest; a secretory phenotype (SASP) that recruits immune cells and modifies the extracellular matrix; and the mobilization of nearby progenitors that repopulate the tissue [29, 30]. Senescent cells produce a series of profibrotic and proinflammatory factors, including interleukin 6 (IL-6), plasminogen activator inhibitor-1 (PAI-1), and transforming growth factor beta (TGF-β), these factors are also known as SASP [31]. SASP can have both beneficial effects and detrimental consequences. The SASP mediates the tumor suppressor functions of senescence [32]. On the other hand, aging cells can develop a characteristic pathogenic SASP that drives secondary senescence and disrupts tissue homeostasis, resulting in loss of tissue repair and regeneration [33, 34]. Identifying the key SASP factors in aging testis and developing a targeted drug to eliminate them may be a potential treatment for aging men [35, 36]. Our study identifies differentially expressed SASP-related genes between young and older human testis samples. Compared with young human testis, we identified upregulated SASP-related genes IGFBP7 and downregulated SASP-related gene including JUN, FOS and UBB. IGFBP7 are important transcriptional inducers of the SASP [37]. IGFBP7 derived from senescent cell has been proved to be key components that triggers senescence in young mesenchymal stem cells [38]. IGFBP7 mainly expressed by Leydig cells from older men. And we confirmed the IGFBP7 protein both in human seminal plasma and mice testis sample. Named by GFBP-related proteins, insulin-like growth factor-binding protein 7 (IGFBP7) is one of the proteins belonging to the IGFBP superfamily. IGFBP7 is a high-affinity insulin-binding protein, acting an inhibitory role by hindering proliferation and inducing apoptosis and senescence [39]. Wajapeyee and et al. reported melanocytes secreted IGFBP7, and it acted through an autocrine/paracrine pathway to induce senescence [40]. Previous studies have also identified dilp-binding protein ImpL2, a drosophila homolog of IGFBP7 blocked InR activation in stem cells of drosophila testis, inducing curb of somatic cell differentiation by downregulating of PI3K/Tor signaling [41]. Higher concentrations of IGFBP7 also indicate increasing risk of cardiovascular events [42]. These studies suggest that IGFBP7 has the potential to serve as a key factor in testicular aging and has significant drug development value.
Removal of senescent cells increases healthy life span in murine models [43, 44]. Due to the potential of reducing the burden of senescent cells to prolong healthy lifespan and delay the onset of age-related diseases, there is increasing interest in developing sensory therapies that integrate multidisciplinary technologies such as biology, chemistry, nanotechnology, and immunology [45–49]. Senolytic tending to selectively deliminate senescent cell is a new therapeutic strategy to manage testicular aging. Multiple basic research have laid the foundation for human clinical trials [50–54]. For now, many senolytic strategies have been investigated, such as inhibitors of the antiapoptotic BCL-2 family proteins, HSP90 inhibitors, USP7 inhibitors, p53 modulators, Na/K-ATPase inhibitors. In our previous study we discovered FOXO4-DRI, a specific FOXO4 blocker, selectively induced p53 nuclear exclusion and apoptosis in senescent Leydig cells, which improved the testicular microenvironment and alleviated age-related testosterone secretion insufficiency [25]. Gypenoside XLIX, a type of dammarane-type saponins that have diverse biological properties, including anti-inflammatory, antithrombotic, anticancer, hepatoprotection, and neuroprotective effects, markedly suppressed the levels of IGFBP7 and reduced the binding of IGFBP7 to IGF1 receptor in an acute kidney injury model, and showed great potential in releasing senescence. However, there were two limitations in our study. First, our conclusion was generated using retrospective data from public databases. Therefore, it should be validated in more prospective and multi-center aging cohorts in the future. Second, the underlying molecular mechanism of IGFBP7 still needs to be further explored.
Conclusions
In summary, here we provide new aspect concerning SASP factors in aging testis. Although there is limited understanding of the detailed molecular mechanisms so far, more investigation needs to be done. Collectively, our data implicate IGFBP7 as a promising aging suppressor protein in aging testis.
Author Contributions
G.L. and X.S. conceived the study and designed the experiments. J.H. performed the data analysis, wrote the methods, and generated the majority of the figures. J. L. and Y.L. performed experiments. J.H. and J.L. wrote the manuscript. X.Z., M.M. and L.L. collected human semen samples. All authors read and approved the manuscript.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Ethical Statement and Consent
This study was approved by the Ethics Review Board of the Sixth Affiliated Hospital of Sun Yat-sen University (2022ZSLYEC-564). All donors have signed an approval consent form. All mice were bought from the Laboratory Animal Center of Sun Yat-sen University. All experimental procedures involving animals were approved by the Institutional Animal Care and Use Committee of Sun Yat-sen University (protocl No. SYSU-IACUC-2022-001850).
Funding
This work is supported in part by the National Natural Science Foundation of China (81971759,82171604), Guangdong Basic and Applied Basic Research Foundation (2023B1515020108), Science and Technology Program of Guangzhou (202206010089), Excellent Talents Training Project of The Sixth Affiliated Hospital of Sun Yat-sen University (R20210217202601970).
References
-
1.
Basaria S. Male hypogonadism. Lancet. 2014; 383:1250–63. https://doi.org/10.1016/S0140-6736(13)61126-5 [PubMed]
-
2.
Levine H, Jørgensen N, Martino-Andrade A, Mendiola J, Weksler-Derri D, Jolles M, Pinotti R, Swan SH. Temporal trends in sperm count: a systematic review and meta-regression analysis of samples collected globally in the 20th and 21st centuries. Hum Reprod Update. 2023; 29:157–76. https://doi.org/10.1093/humupd/dmac035 [PubMed]
-
3.
Levine H, Jørgensen N, Martino-Andrade A, Mendiola J, Weksler-Derri D, Mindlis I, Pinotti R, Swan SH. Temporal trends in sperm count: a systematic review and meta-regression analysis. Hum Reprod Update. 2017; 23:646–59. https://doi.org/10.1093/humupd/dmx022 [PubMed]
-
4.
Humm KC, Sakkas D. Role of increased male age in IVF and egg donation: is sperm DNA fragmentation responsible? Fertil Steril. 2013; 99:30–6. https://doi.org/10.1016/j.fertnstert.2012.11.024 [PubMed]
-
5.
Kaufman JM, Lapauw B, Mahmoud A, T’Sjoen G, Huhtaniemi IT. Aging and the Male Reproductive System. Endocr Rev. 2019; 40:906–72. https://doi.org/10.1210/er.2018-00178 [PubMed]
-
6.
Sharma R, Agarwal A, Rohra VK, Assidi M, Abu-Elmagd M, Turki RF. Effects of increased paternal age on sperm quality, reproductive outcome and associated epigenetic risks to offspring. Reprod Biol Endocrinol. 2015; 13:35. https://doi.org/10.1186/s12958-015-0028-x [PubMed]
-
7.
Frungieri MB, Calandra RS, Bartke A, Matzkin ME. Male and female gonadal ageing: its impact on health span and life span. Mech Ageing Dev. 2021; 197:111519. https://doi.org/10.1016/j.mad.2021.111519 [PubMed]
-
8.
Pataky MW, Young WF, Nair KS. Hormonal and Metabolic Changes of Aging and the Influence of Lifestyle Modifications. Mayo Clin Proc. 2021; 96:788–814. https://doi.org/10.1016/j.mayocp.2020.07.033 [PubMed]
-
9.
Eisenberg ML, Li S, Behr B, Pera RR, Cullen MR. Relationship between semen production and medical comorbidity. Fertil Steril. 2015; 103:66–71. https://doi.org/10.1016/j.fertnstert.2014.10.017 [PubMed]
-
10.
Anawalt BD, Matsumoto AM. Aging and androgens: Physiology and clinical implications. Rev Endocr Metab Disord. 2022; 23:1123–37. https://doi.org/10.1007/s11154-022-09765-2 [PubMed]
-
11.
Dong S, Chen C, Zhang J, Gao Y, Zeng X, Zhang X. Testicular aging, male fertility and beyond. Front Endocrinol (Lausanne). 2022; 13:1012119. https://doi.org/10.3389/fendo.2022.1012119 [PubMed]
-
12.
Wolf KN, Wildt DE, Vargas A, Marinari PE, Kreeger JS, Ottinger MA, Howard JG. Age-dependent changes in sperm production, semen quality, and testicular volume in the black-footed ferret (Mustela nigripes). Biol Reprod. 2000; 63:179–87. https://doi.org/10.1095/biolreprod63.1.179 [PubMed]
-
13.
Nie X, Munyoki SK, Sukhwani M, Schmid N, Missel A, Emery BR, Do, Stukenborg JB, Mayerhofer A, Orwig KE, Aston KI, Hotaling JM, Cairns BR, Guo J. Single-cell analysis of human testis aging and correlation with elevated body mass index. Dev Cell. 2022; 57:1160–76.e5. https://doi.org/10.1016/j.devcel.2022.04.004 [PubMed]
-
14.
Mularoni V, Esposito V, Di Persio S, Vicini E, Spadetta G, Berloco P, Fanelli F, Mezzullo M, Pagotto U, Pelusi C, Nielsen JE, Rajpert-De Meyts E, Jorgensen N, et al. Age-related changes in human Leydig cell status. Hum Reprod. 2020; 35:2663–76. https://doi.org/10.1093/humrep/deaa271 [PubMed]
-
15.
Santiago J, Silva JV, Alves MG, Oliveira PF, Fardilha M. Testicular Aging: An Overview of Ultrastructural, Cellular, and Molecular Alterations. J Gerontol A Biol Sci Med Sci. 2019; 74:860–71. https://doi.org/10.1093/gerona/gly082 [PubMed]
-
16.
Birch J, Gil J. Senescence and the SASP: many therapeutic avenues. Genes Dev. 2020; 34:1565–76. https://doi.org/10.1101/gad.343129.120 [PubMed]
-
17.
Basisty N, Kale A, Jeon OH, Kuehnemann C, Payne T, Rao C, Holtz A, Shah S, Sharma V, Ferrucci L, Campisi J, Schilling B. A proteomic atlas of senescence-associated secretomes for aging biomarker development. PLoS Biol. 2020; 18:e3000599. https://doi.org/10.1371/journal.pbio.3000599 [PubMed]
-
18.
Zhang Y, Liu T, Hu X, Wang M, Wang J, Zou B, Tan P, Cui T, Dou Y, Ning L, Huang Y, Rao S, Wang D, Zhao X. CellCall: integrating paired ligand-receptor and transcription factor activities for cell-cell communication. Nucleic Acids Res. 2021; 49:8520–34. https://doi.org/10.1093/nar/gkab638 [PubMed]
-
19.
Hou R, Denisenko E, Ong HT, Ramilowski JA, Forrest ARR. Predicting cell-to-cell communication networks using NATMI. Nat Commun. 2020; 11:5011. https://doi.org/10.1038/s41467-020-18873-z [PubMed]
-
20.
Zhang Y, Liu T, Wang J, Zou B, Li L, Yao L, Chen K, Ning L, Wu B, Zhao X, Wang D. Cellinker: a platform of ligand-receptor interactions for intercellular communication analysis. Bioinformatics. 2021; btab036. [Epub ahead of print]. https://doi.org/10.1093/bioinformatics/btab036 [PubMed]
-
21.
Shao X, Liao J, Li C, Lu X, Cheng J, Fan X. CellTalkDB: a manually curated database of ligand-receptor interactions in humans and mice. Brief Bioinform. 2021; 22:bbaa269. https://doi.org/10.1093/bib/bbaa269 [PubMed]
-
22.
Jin S, Guerrero-Juarez CF, Zhang L, Chang I, Ramos R, Kuan CH, Myung P, Plikus MV, Nie Q. Inference and analysis of cell-cell communication using CellChat. Nat Commun. 2021; 12:1088. https://doi.org/10.1038/s41467-021-21246-9 [PubMed]
-
23.
Szklarczyk D, Gable AL, Lyon D, Junge A, Wyder S, Huerta-Cepas J, Simonovic M, Doncheva NT, Morris JH, Bork P, Jensen LJ, Mering CV. STRING v11: protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019; 47:D607–13. https://doi.org/10.1093/nar/gky1131 [PubMed]
-
24.
Huang K, Gong H, Guan J, Zhang L, Hu C, Zhao W, Huang L, Zhang W, Kim P, Zhou X. AgeAnno: a knowledgebase of single-cell annotation of aging in human. Nucleic Acids Res. 2023; 51:D805–15. https://doi.org/10.1093/nar/gkac847 [PubMed]
-
25.
Zhang C, Xie Y, Chen H, Lv L, Yao J, Zhang M, Xia K, Feng X, Li Y, Liang X, Sun X, Deng C, Liu G. FOXO4-DRI alleviates age-related testosterone secretion insufficiency by targeting senescent Leydig cells in aged mice. Aging (Albany NY). 2020; 12:1272–84. https://doi.org/10.18632/aging.102682 [PubMed]
-
26.
Luo D, Qi X, Xu X, Yang L, Yu C, Guan Q. Involvement of p38 MAPK in Leydig cell aging and age-related decline in testosterone. Front Endocrinol (Lausanne). 2023; 14:1088249. https://doi.org/10.3389/fendo.2023.1088249 [PubMed]
-
27.
Handelsman DJ. Testosterone and Male Aging: Faltering Hope for Rejuvenation. JAMA. 2017; 317:699–701. https://doi.org/10.1001/jama.2017.0129 [PubMed]
-
28.
Rodrigues Dos Santos M, Bhasin S. Benefits and Risks of Testosterone Treatment in Men with Age-Related Decline in Testosterone. Annu Rev Med. 2021; 72:75–91. https://doi.org/10.1146/annurev-med-050219-034711 [PubMed]
-
29.
Muñoz-Espín D, Serrano M. Cellular senescence: from physiology to pathology. Nat Rev Mol Cell Biol. 2014; 15:482–96. https://doi.org/10.1038/nrm3823 [PubMed]
-
30.
Storer M, Mas A, Robert-Moreno A, Pecoraro M, Ortells MC, Di Giacomo V, Yosef R, Pilpel N, Krizhanovsky V, Sharpe J, Keyes WM. Senescence is a developmental mechanism that contributes to embryonic growth and patterning. Cell. 2013; 155:1119–30. https://doi.org/10.1016/j.cell.2013.10.041 [PubMed]
-
31.
Valentijn FA, Falke LL, Nguyen TQ, Goldschmeding R. Cellular senescence in the aging and diseased kidney. J Cell Commun Signal. 2018; 12:69–82. https://doi.org/10.1007/s12079-017-0434-2 [PubMed]
-
32.
Acosta JC, O’Loghlen A, Banito A, Guijarro MV, Augert A, Raguz S, Fumagalli M, Da Costa M, Brown C, Popov N, Takatsu Y, Melamed J, d’Adda di Fagagna F, et al. Chemokine signaling via the CXCR2 receptor reinforces senescence. Cell. 2008; 133:1006–18. https://doi.org/10.1016/j.cell.2008.03.038 [PubMed]
-
33.
Zhang L, Pitcher LE, Yousefzadeh MJ, Niedernhofer LJ, Robbins PD, Zhu Y. Cellular senescence: a key therapeutic target in aging and diseases. J Clin Invest. 2022; 132:e158450. https://doi.org/10.1172/JCI158450 [PubMed]
-
34.
Cuollo L, Antonangeli F, Santoni A, Soriani A. The Senescence-Associated Secretory Phenotype (SASP) in the Challenging Future of Cancer Therapy and Age-Related Diseases. Biology (Basel). 2020; 9:485. https://doi.org/10.3390/biology9120485 [PubMed]
-
35.
Rodier F, Coppé JP, Patil CK, Hoeijmakers WA, Muñoz DP, Raza SR, Freund A, Campeau E, Davalos AR, Campisi J. Persistent DNA damage signalling triggers senescence-associated inflammatory cytokine secretion. Nat Cell Biol. 2009; 11:973–9. https://doi.org/10.1038/ncb1909 [PubMed]
-
36.
Takahashi A, Imai Y, Yamakoshi K, Kuninaka S, Ohtani N, Yoshimoto S, Hori S, Tachibana M, Anderton E, Takeuchi T, Shinkai Y, Peters G, Saya H, Hara E. DNA damage signaling triggers degradation of histone methyltransferases through APC/C(Cdh1) in senescent cells. Mol Cell. 2012; 45:123–31. https://doi.org/10.1016/j.molcel.2011.10.018 [PubMed]
-
37.
Jones TA, Jeyapalan JN, Forshew T, Tatevossian RG, Lawson AR, Patel SN, Doctor GT, Mumin MA, Picker SR, Phipps KP, Michalski A, Jacques TS, Sheer D. Molecular analysis of pediatric brain tumors identifies microRNAs in pilocytic astrocytomas that target the MAPK and NF-κB pathways. Acta Neuropathol Commun. 2015; 3:86. https://doi.org/10.1186/s40478-015-0266-3 [PubMed]
-
38.
Severino V, Alessio N, Farina A, Sandomenico A, Cipollaro M, Peluso G, Galderisi U, Chambery A. Insulin-like growth factor binding proteins 4 and 7 released by senescent cells promote premature senescence in mesenchymal stem cells. Cell Death Dis. 2013; 4:e911. https://doi.org/10.1038/cddis.2013.445 [PubMed]
-
39.
Jin L, Shen F, Weinfeld M, Sergi C. Insulin Growth Factor Binding Protein 7 (IGFBP7)-Related Cancer and IGFBP3 and IGFBP7 Crosstalk. Front Oncol. 2020; 10:727. https://doi.org/10.3389/fonc.2020.00727 [PubMed]
-
40.
Wajapeyee N, Serra RW, Zhu X, Mahalingam M, Green MR. Role for IGFBP7 in senescence induction by BRAF. Cell. 2010; 141:746–7. https://doi.org/10.1016/j.cell.2010.05.014 [PubMed]
-
41.
Amoyel M, Hillion KH, Margolis SR, Bach EA. Somatic stem cell differentiation is regulated by PI3K/Tor signaling in response to local cues. Development. 2016; 143:3914–25. https://doi.org/10.1242/dev.139782 [PubMed]
-
42.
Gandhi PU, Chow SL, Rector TS, Krum H, Gaggin HK, McMurray JJ, Zile MR, Komajda M, McKelvie RS, Carson PE, Januzzi JL Jr, Anand IS. Prognostic Value of Insulin-Like Growth Factor-Binding Protein 7 in Patients with Heart Failure and Preserved Ejection Fraction. J Card Fail. 2017; 23:20–8. https://doi.org/10.1016/j.cardfail.2016.06.006 [PubMed]
-
43.
Baker DJ, Wijshake T, Tchkonia T, LeBrasseur NK, Childs BG, van de Sluis B, Kirkland JL, van Deursen JM. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature. 2011; 479:232–6. https://doi.org/10.1038/nature10600 [PubMed]
-
44.
Baker DJ, Childs BG, Durik M, Wijers ME, Sieben CJ, Zhong J, Saltness RA, Jeganathan KB, Verzosa GC, Pezeshki A, Khazaie K, Miller JD, van Deursen JM. Naturally occurring p16(Ink4a)-positive cells shorten healthy lifespan. Nature. 2016; 530:184–9. https://doi.org/10.1038/nature16932 [PubMed]
-
45.
Paez-Ribes M, González-Gualda E, Doherty GJ, Muñoz-Espín D. Targeting senescent cells in translational medicine. EMBO Mol Med. 2019; 11:e10234. https://doi.org/10.15252/emmm.201810234 [PubMed]
-
46.
Zhang L, Pitcher LE, Prahalad V, Niedernhofer LJ, Robbins PD. Targeting cellular senescence with senotherapeutics: senolytics and senomorphics. FEBS J. 2023; 290:1362–83. https://doi.org/10.1111/febs.16350 [PubMed]
-
47.
Zhang L, Pitcher LE, Prahalad V, Niedernhofer LJ, Robbins PD. Recent advances in the discovery of senolytics. Mech Ageing Dev. 2021; 200:111587. https://doi.org/10.1016/j.mad.2021.111587 [PubMed]
-
48.
He S, Sharpless NE. Senescence in Health and Disease. Cell. 2017; 169:1000–11. https://doi.org/10.1016/j.cell.2017.05.015 [PubMed]
-
49.
Borghesan M, Hoogaars WMH, Varela-Eirin M, Talma N, Demaria M. A Senescence-Centric View of Aging: Implications for Longevity and Disease. Trends Cell Biol. 2020; 30:777–91. https://doi.org/10.1016/j.tcb.2020.07.002 [PubMed]
-
50.
Camell CD, Yousefzadeh MJ, Zhu Y, Prata LG, Huggins MA, Pierson M, Zhang L, O’Kelly RD, Pirtskhalava T, Xun P, Ejima K, Xue A, Tripathi U, et al. Senolytics reduce coronavirus-related mortality in old mice. Science. 2021; 373:eabe4832. https://doi.org/10.1126/science.abe4832 [PubMed]
-
51.
Xu M, Pirtskhalava T, Farr JN, Weigand BM, Palmer AK, Weivoda MM, Inman CL, Ogrodnik MB, Hachfeld CM, Fraser DG, Onken JL, Johnson KO, Verzosa GC, et al. Senolytics improve physical function and increase lifespan in old age. Nat Med. 2018; 24:1246–56. https://doi.org/10.1038/s41591-018-0092-9 [PubMed]
-
52.
Bussian TJ, Aziz A, Meyer CF, Swenson BL, van Deursen JM, Baker DJ. Clearance of senescent glial cells prevents tau-dependent pathology and cognitive decline. Nature. 2018; 562:578–82. https://doi.org/10.1038/s41586-018-0543-y [PubMed]
-
53.
Zhu Y, Tchkonia T, Pirtskhalava T, Gower AC, Ding H, Giorgadze N, Palmer AK, Ikeno Y, Hubbard GB, Lenburg M, O’Hara SP, LaRusso NF, Miller JD, et al. The Achilles’ heel of senescent cells: from transcriptome to senolytic drugs. Aging Cell. 2015; 14:644–58. https://doi.org/10.1111/acel.12344 [PubMed]
-
54.
Musi N, Valentine JM, Sickora KR, Baeuerle E, Thompson CS, Shen Q, Orr ME. Tau protein aggregation is associated with cellular senescence in the brain. Aging Cell. 2018; 17:e12840. https://doi.org/10.1111/acel.12840 [PubMed]