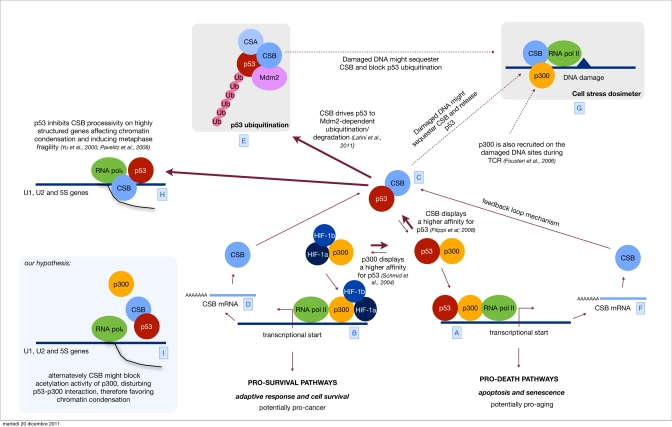
Figure 1.Orchestrating p53 activityHypoxic stress is known to up regulate both the HIF-1 controlled response and the antagonistic p53 response [A]. Sustained p53 activity results in cell demise by promoting the transcription of cell cycle arrest genes, such as p21 and apoptosis-inducing genes, such as Bax [B]. Alternatively, HIF-1 can prevent cell death by promoting the transcription of genes that determine adaptive responses and cell survival [C]. We have previously showed that CSB, which is also induced by hypoxia [D], interacts with p53 and therefore releases the limiting co-factor p300 from p53. By doing so, CSB negatively modulates the transactivation activity of p53 and up regulates the transactivation of other transcription factor related pathways (in the specific the HIF-1 dependent pathway) therefore re-equilibrating the physiological response toward cell proliferation and survival instead of cell cycle arrest and cell death. [E] Recently we have discovered that CSB drives p53 to ubiquitination/degradation, in an Mdm2 dependent fashion, therefore down regulating the cellular levels of p53. CSB is part of a feedback loop where its expression is induced by p53 itself [F]. This mechanism helps the cell to find a reasonable equilibrium between the transient up regulation of the p53 response, aimed to temporarily arrest the cell cycle and potentiate the DNA repair mechanism, and its down modulation in order not to reach the point of no-return that would trigger apoptosis. Others have established that after genotoxic stress (UV, oxidative stress) CSB is also recruited to DNA damaged sites [G], we propose a model in which CSB, besides acting in DNA repair, may act as a sort of dosimeter in order to modulate either the transcriptional activity of p53 (p53-p300 interaction) or its degradation. Indeed, high levels of DNA damage would sequester CSB at the damaged sites thus blocking its function in the down modulation of p53 activity and degradation. Parallel recruitment of p300 by CSB favors the accessibility of the damaged site to the repairosome and potentially inhibits the transcription (at least the p300 related) contributing to the transient shut down of the transcription in response to DNA damage. [H] Finally, the interaction CSB/p53 has been suggested also to play a role in metaphase fragility. p53 inhibits CSB-processivity affecting chromatin condensation of highly structured genes, such as U1, U2 and 5S, resulting in metaphase fragility [I]. We propose an alternative hypothesis, where CSB might favor chromatin condensation on these sites by disturbing p300/p53 interaction and chromatin acetylation.