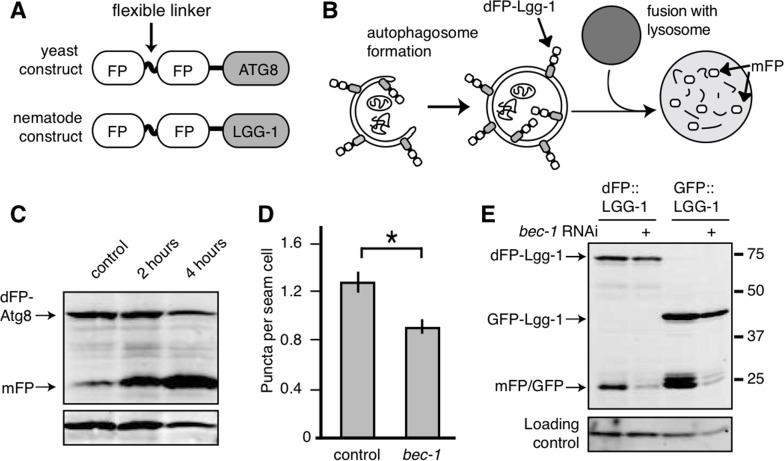
Figure 1.Monomeric FP released from dFP::LGG-1 correlates with autophagic flux(A) Yeast and nematode constructs were made by fusing a double fluorescent protein module, dFP, to ATG8 (yeast) or LGG-1 (nematode). The dFP contained two fluorescent proteins (labeled FP) separated by a protease-sensitive flexible linker. (B) The dFP Atg8/LGG-1 constructs are recruited to the growing autophagosome. Upon fusion of the autophagosome with the lysosome, the dFP is cleaved by lysosomal proteases, releasing soluble FP monomers. These monomers are fairly stable, but will eventually be further degraded by proteaolysis in the lysosome. (C) S. cerevisiae expressing dFP::ATG8 showed a time-dependent increase in mFP abundance with 2 or 4 hours of nitrogen starvation. (D) The number of dFP punctae in seam cells of L3 nematodes expressing dFP::LGG-1 from extrachromosomal arrays was reduced when the nematodes were treated with bec-1 RNAi to impair autophagy, as has been previously observed for GFP::LGG-1 [14]. Error bars represent standard error, asterisk for p<0.05. (E) C. elegans with extrachromosomal arrays expressing either eft-3p::dFP::LGG-1 or lgg-1p::GFP::LGG-1 show mFP release. Full-length dFP::LGG-1 is present at 73kDa, GFP-LGG-1 at 40kDa, and the monomers of GFP or mFP are present at 25kDa. The accumulation of mFP was inhibited by treatment with bec-1 RNAi (lanes 2 and 4). The accumulation of mFP in animals grown on control RNAi food is shown in lanes 1 and 3.