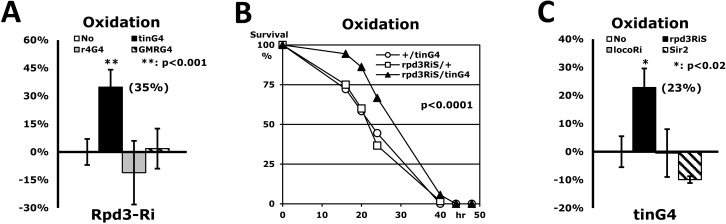
Figure 3.Only heart-specific Rpd3 downregulation enhances stress resistance(A) Rpd3 downregulation (rpd3Ri) limited to particular tissues (No: rpd3Ri/+ control; tinG4: heart; r4G4: fatbody; GMRG4: eye) on an oxidation stress assay. 2-day-old male flies were used for this experiment, with the exception of the rpd3Ri/r4G4 genotype, which used 5-day-old females due to the semi-lethality of the males. The percent change in median survival time of the flies is indicated as mean ± SEM, which was calculated from the stress response curves of at least 4 independent experiments. The data represent percent differences (%) normalized from the rpd3Ri/+ control (No), indicating that only heart-specific Rpd3 downregulation (rpd3Ri/tinG4) exhibits a statistically significant impact on stress tolerance (p <0.001). (B) The oxidation stress survival curve of heart-specific Rpd3 downregulation using a different rpd3RNAi (rpd3RiS/tinG4). The target region of rpd3Ri: 482bp in the middle of exon; rpd3RiS: 21bp in the 3′ UTR. (C) Heart-specific modulation of long-lived genes (No: +/tinG4 control; rpd3RiS: Rpd3 downregulation; locoRi: Loco downregulation; Sir2: Sir2 upregulation) on an oxidation stress assay. The percent change of median survival times was calculated from the 3 - 5 independent curves (B) of the oxidative stress test. The percentage data, normalized from the +/tinG4 control (No), shows that within the heart only Rpd3 downregulation (rpd3Ri/tinG4) significantly enhances stress resistance. The decrease of stress resistance in the rpd3Ri/r4G4 (A) and Sir2/tinG4 (C) genotypes were not statistically significant when compared to their respective single transgene controls (+/r4G4 and Sir2/+).