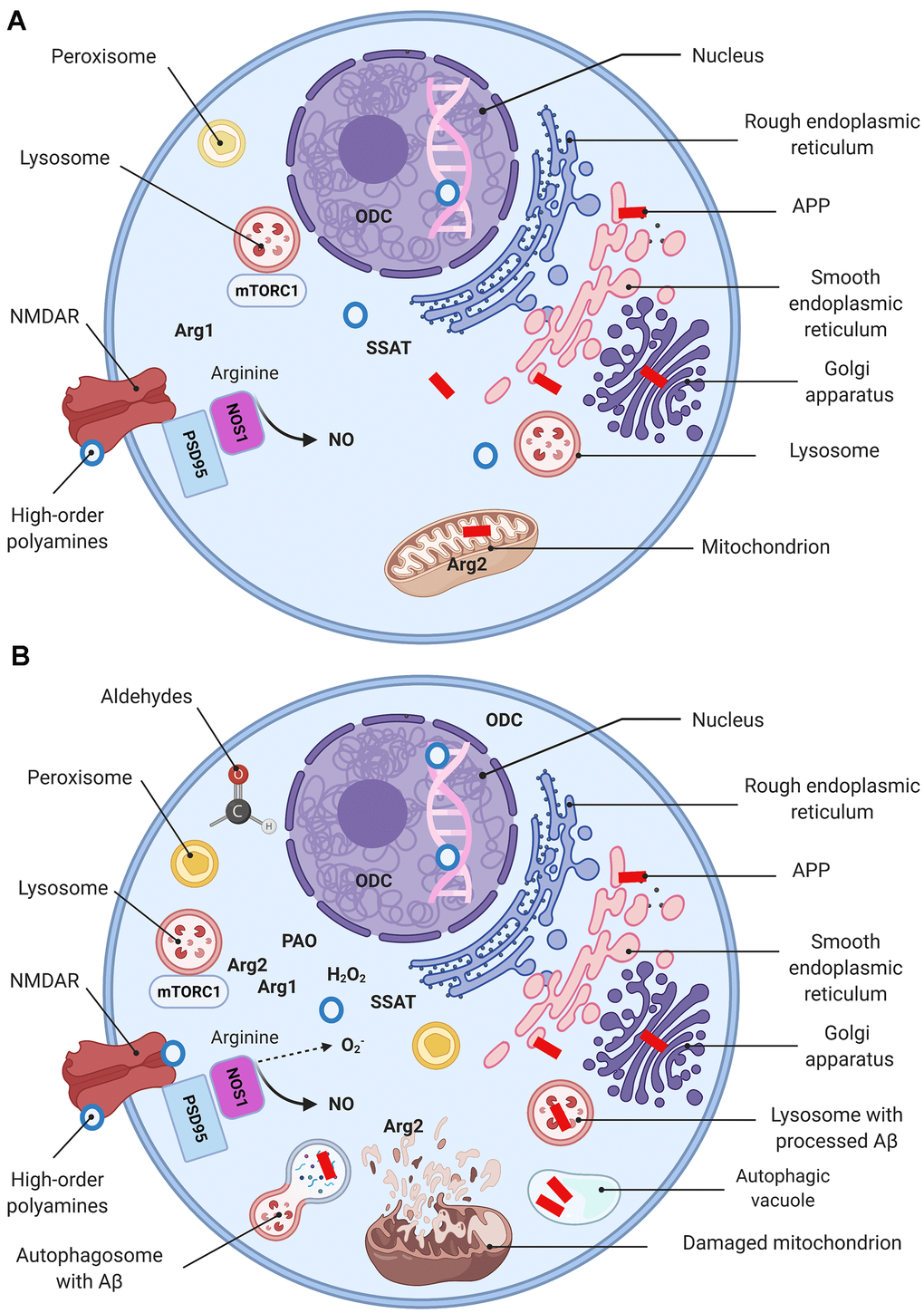
Figure 4.Molecular basis of AD (a simplified model). Schematic representation of a “normal” (A) and a dysfunctional (B) neuron with main organelles involved in APP processing and PSR. Following synthesis, APP undergoes posttranslational modifications in the ER and Golgi where it generates Aβ. In the healthy brain Aβ is localized in perikarya, trans-Golgi network, Golgi-derived vesicles (A), while oxidative stress upsurges intralysosomal Aβ content (B) (149). Mitochondria are essential for maintaining neuronal integrity and function. Arg2 is a typical mitochondria-associated enzyme (A). Mitochondria are targets for Aβ/ROS-mediated damage, which leads to swelling, outer membrane rupture, and followed by Arg2 appearance in the cytoplasm. Arg2, in turn, induces re-distribution of lysosome and mTOR from perinuclear area to cell periphery (B), activates mTORC1-S6K1 signaling and contributes to cell senescence phenotype characterized by impaired autophagy and apoptosis. This process eventuates in impediment of the autophagolysosomes maturation and lysosome-associated Aβ degradation, and leads to Aβ accumulation in autophagic vacuoles (B). Peroxisomes are not competent to cope with growing oxidative stress and become target for ROS. Oxidative stress leads to Arg1 overexpression, ODC translocation to the cytoplasm, and eventuates by polyamine overproduction (PSR). High-order polyamines stabilize the DNA conformation and modulate the chromatin structure and gene transcription via ionic interactions (A). Their elevated levels shut down some vital genes (B). Extensive polyamine catabolism is followed by the generation of hydrogen peroxide and cytotoxic aldehydes. NOS1 deprived of arginine undergoes uncoupling and switches to the production of superoxide anion. NMDA receptor function is modulated by polyamines via a special recognition site.