



Skeleton in vertebrates serves multiple mechanical, hematopoietic and endocrine functions [1,2]. In order to perform its functions properly, the skeleton continuously renews itself through a homeostatic process known as bone remodeling [1,3]. Bone remodeling occurs constantly and simultaneously in numerous parts of skeleton spread throughout the body and requires considerable inflow and utilization of energy [4]. Any perturbance in energy homeostasis of the body can therefore result in dramatic changes in skeletal metabolism [5,6]. For example, obesity or high body mass index often reduces fracture risk, whereas on the other hand anorexia enhances it [5,6]. These observations provide clinical evidence that bone and energy metabolism are balanced with each other and are likely co-regulated.
Serotonin (5-hydroxytryptamine) is a biogenic amine that functions both as a neurotransmitter in central nervous system and as a hormone in the periphery where most of it (95%) is produced [7,8]. Serotonin is generated through an enzymatic pathway in which L-tryptophan is converted into L-5OH-tryptophan by an enzyme called tryptophan hydroxylase (Tph); this inter mediate product is then converted to serotonin by an aromatic L-aminoacid decarboxylase [7,8]. There are two Tph genes: Tph1 and Tph2. Tph1 is expressed mostly in cells of the gut and is responsible for the production of peripheral serotonin [9]. Tph2 is expressed exclusively in neurons of the brainstem and is responsible for the production of serotonin in the brain [8]. Moreover, serotonin does not cross the blood brain barrier; therefore it should be viewed from a functional point of view as two distinct molecules [7]. Brain-derived serotonin (BDS) acts as a neurotransmitter, while gut-derived serotonin (GDS) acts as a hormone and regulates a wide variety of processes [10]. The importance of serotonin in the regulation of bone mass is underscored by two clinical observations. First, depressed patients, that allegedly have low serotonergic tone, also have low bone mass [11]; and second, serotonin reuptake inhibitors (SSRI's) when taken chronically can either increase or more often decrease bone mass [12].
Our studies with loss and gain of function mutations of low-density lipoprotein receptor-related protein 5 demonstrated that GDS is a powerful inhibitor of osteoblast proliferation and bone formation that does not affect bone resorption [13]. Although correlative in nature, these studies showed that an increase in extracellular concentration of blood serotonin in patients on SSRI's may explain their often observed low bone mass phenotype [12]. However, the influence of this gut-bone axis on bone mass could not explain the increase in bone mass observed in another study with SSRI's [14]. In our quest to understand the serotonin regulation of bone mass in vertebrates we inactivatedTph2, the gene that catalyzes the rate-limiting step in the biosynthesis of BDS. The absence of serotonin in the brain resulted in a severe low bone mass phenotype affecting the axial (vertebrae) and appendicular (long bones) skeleton [15]. This phenotype was secondary to a decrease in bone formation parameters (osteoblast numbers and bone formation rate) and to an increase in bone resorption parameters (osteoclast surface and circulating Dpd levels) [15]. Hence, BDS is a positive and powerful regulator of bone mass accrual acting on both arms of bone remodeling despite accounting for >5% of total serotonin pool in the body it overrides the GDS regulation of bone mass [15].
While we were doing these studies we noticed, upon opening the abdominal cavities, that Tph2-deficient animals had a dramatic decrease in their adipose mass [15]. This prompted us to analyze in great detail their energy metabolism phenotype. The decrease in their fat mass was due, in part, to the fact that these mice ate less and spent much more energy compared to their wild type littermates [15]. This observation was not entirely surprising since serotonin is known to play important roles in many other physiological processes. However what caught our attention was the fact that the three most notable phenotypes of adult Tph2-deficient animals i.e., decrease in bone mass and appetite, and an increase in the energy expenditure are a mirror image of what is observed in mice that lack leptin [16,17].
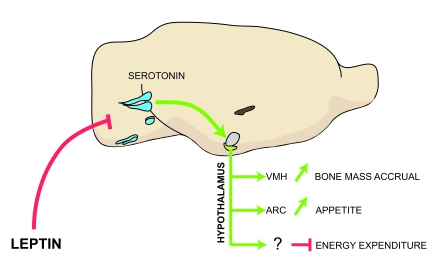
Figure 1. Model of the leptin-dependent central control of bone mass, appetite and energy expenditure.
Leptin inhibits release of brainstem-derived serotonin, which favors bone
mass accrual and appetite through its action on hypothalamic neurons.
Serotonergic neurons are in blue; VMH, ventromedial hypothalamus; ARC,
arcuate; VMH is in purple and arcuate is in green.
Three correlative experiments suggested that leptin might signal in the serotonin neurons to regulate some of its downstream functions. First, leptin receptor is expressed on serotonin neurons located in the raphe nuclei of brainstem and is functional [15]. Second, serotonin neurons project to the key hypothalamic nuclei responsible for the regulation of appetite, energy expenditure and bone mass [15]. Third, patients on SSRI's have been reported to have changes in their appetite and bone mass [12,18]. To explore that leptin may signal through the serotonin neurons to achieve these three functions, we inactivated leptin receptors in different nuclei of the hypothalamus or the serotonergic neurons of the brainstem [15]. Mice lacking ObRb either in Sf1-expressing neurons of the ventromedial hypothalamus (VMH) nuclei or in Pomc-expressing neurons of the arcuate (ARC) nuclei had normal sympathetic activity, bone remodeling parameters and bone mass; they also had normal appetite and energy expenditure, and when fed a normal diet, did not develop an obesity phenotype [19,20]. In contrast, mice that lack ObRb in serotonin neurons (ObRbSERT-/-) developed a high bone mass phenotype; they had a similar increase in appetite as ob/ob mice and had low energy expenditure. As a result, ObRbSERT-/- mice, when fed a normal diet, developed an obesity phenotype. These genetic studies demonstrated that leptin signals in the serotonin neurons of the brainstem to regulate, to the most part, bone mass, appetite and energy expenditure (Figure 1).
The demonstration that leptin-dependent central control of bone mass, appetite and energy expenditure occurs through its ability to inhibit serotonin production raised questions about the location and identity of serotonin receptors on hypothalamic neurons mediating these function. Double fluorescence in situ hybridization and nuclei-specific gene inactivation experiments revealed that serotonin promotes bone mass accrual through Htr2c receptors expressed on the VMH nuclei, while appetite through Htr2b and Htr1a receptors expressed on ARC nuclei of the hypothalamus. Further analysis revealed that Htr2c is upstream of the sympathetic center of the brain while Htr1a and Htr2b achieve their functions on appetite through modulation of melanocortin signaling (Figure 1).
In summary, these studies provide new insights into the central control of appetite and bone mass accrual and they identify serotonin as a focal point in the leptin-dependent common central control of bone and energy metabolisms.
Acknowledgement
This work was supported by NIH grants (VKY, GK) and a Rodan fellowship from IBMS (VKY).
Conflicts of Interest
The authors of this manuscript have no conflict of interest to declare.
References
- 1. Rodan GA and Martin TJ. Therapeutic approaches to bone diseases. Science. 2000; 289: 1508 -1514. [PubMed] .
- 2. Lee NK , Sowa H , Hinoi E , Ferron M , Ahn JD , Confavreux C , Dacquin R , Mee PJ , McKee MD , Jung DY , Zhang Z , Kim JK , Mauvais-Jarvis F , Ducy P and Karsenty G. Endocrine regulation of energy metabolism by the skeleton. Cell. 2007; 130: 456 -469. [PubMed] .
- 3. Zaidi M Skeletal remodeling in health and disease. Nat Med. 2007; 13: 791 -801. [PubMed] .
- 4. Karsenty G Convergence between bone and energy homeostases: Leptin regulation of bone mass. Cell Metab. 2006; 4: 341 -348. [PubMed] .
- 5. Felson DT , Zhang Y , Hannan MT and Anderson JJ. Effects of weight and body mass index on bone mineral density in men and women: the Framingham study. J Bone Miner Res. 1993; 8: 567 -573. [PubMed] .
- 6. Tremollieres FA , Pouilles JM and Ribot C. Vertebral postmenopausal bone loss is reduced in overweight women: a longitudinal study in 155 earlypostmenopausal women. J Clin Endocrinol Metab. 1993; 77: 683 -686. [PubMed] .
- 7. Mann JJ , McBride PA , Brown RP , Linnoila M , Leon AC , DeMeo M , Mieczkowski T , Myers JE and Stanley M. Relationship between central and peripheral serotonin indexes in depressed and suicidal psychiatric inpatients. Arch Gen Psychiatry. 1992; 49: 442 -446. [PubMed] .
- 8. Walther DJ , Peter JU , Bashammakh S , Hortnagl H , Voits M , Fink H and Bader M. Synthesis of serotonin by a second tryptophan hydroxylase isoform. Science. 2003; 299: 76 [PubMed] .
- 9. Gershon MD and Tack J. The serotonin signaling system: from basic understanding to drug development for functional GI disorders. Gastroenterology. 2007; 132: 397 -414. [PubMed] .
- 10. Heath MJ and Hen R. Serotonin receptors: Genetic insights into serotonin function. Curr Biol. 1995; 5: 997 -999. [PubMed] .
- 11. Eskandari F , Martinez PE , Torvik S , Phillips TM , Sternberg EM , Mistry S , Ronsaville D , Wesley R , Toomey C , Sebring NG , Reynolds JC , Blackman MR , Calis KA , Gold PW and Cizza G. Low bone mass in premenopausal women with depression. Arch Intern Med. 2007; 167: 2329 -2336. [PubMed] .
- 12. Richards JB , Papaioannou A , Adachi JD , Joseph L , Whitson HE , Prior JC and Goltzman D. Effect of selective serotonin reuptake inhibitors on the risk of fracture. Arch Intern Med. 2007; 167: 188 -194. [PubMed] .
- 13. Yadav VK , Ryu JH , Suda N , Tanaka KF , Gingrich JA , Schutz G , Glorieux FH , Chiang CY , Zajac JD , Insogna KL , Mann JJ , Hen R , Ducy P and Karsenty G. Lrp5 controls bone formation by inhibiting serotonin synthesis in the duodenum. Cell. 2008; 135: 825 -837. [PubMed] .
- 14. Battaglino R , Vokes M , Schulze-Spate U , Sharma A , Graves D , Kohler T , Muller R , Yoganathan S and Stashenko P. Fluoxetine treatment increases trabecular bone formation in mice. J Cell Biochem. 2007; 100: 1387 -1394. [PubMed] .
- 15. Yadav VK , Oury F , Suda N , Liu ZW , Gao XB , Confavreux C , Klemenhagen KC , Tanaka KF , Gingrich JA , Guo XE , Tecott LH , Mann JJ , Hen R , Horvath TL and Karsenty G. A serotonin-dependent mechanism explains the leptin regulation of bone mass, appetite, and energy expenditure. Cell. 2009; 138: 976 -989. [PubMed] .
- 16. Friedman JM and Halaas JL. Leptin and the regulation of body weight in mammals. Nature. 1998; 395: 763 -770. [PubMed] .
- 17. Spiegelman BM and Flier JS. Obesity and the regulation of energy balance. Cell. 2001; 104: 531 -543. [PubMed] .
- 18. Michelson D , Amsterdam JD , Quitkin FM , Reimherr FW , Rosenbaum JF , Zajecka J , Sundell KL , Kim Y and Beasley CM Jr. Changes in weight during a 1-year trial of fluoxetine. Am J Psychiatry. 1999; 156: 1170 -1176. [PubMed] .
- 19. Dhillon H , Zigman JM , Ye C , Lee CE , McGovern RA , Tang V , Kenny CD , Christiansen LM , White RD , Edelstein EA , Coppari R , Balthasar N , Cowley MA , Chua S Jr , Elmquist JK and Lowell BB. Leptin directly activates SF1 neurons in the VMH, and this action by leptin is required for normal body-weight homeostasis. Neuron. 2006; 49: 191 -203. [PubMed] .
- 20. Balthasar N , Coppari R , McMinn J , Liu SM , Lee CE , Tang V , Kenny CD , McGovern RA , Chua SC Jr , Elmquist JK and Lowell BB. Leptin receptor signaling in POMC neurons is required for normal body weight homeostasis. Neuron. 2004; 42: 983 -991. [PubMed] .