



Introduction
The link between aging and insulin/IGF-1 signaling has attracted substantial attention during last years. The potential connection was evidenced by an increase in incidence of insulin resistance and type 2 diabetes in accelerated aging syndromes, on the one side, as well as by life span extension due to caloric restriction (CR) in rodents, on the other[1]. Concomitant reduction in plasma insulin and plasma glucose levels, which implies increased sensitivity to insulin, emerges as a hallmark of increased longevity [2]. Hyperglycemia is an important aging factor involved in generation of advanced glycosylation endproducts (AGEs) [3,4]. There is evidence that hyperinsulinemia favors accumulation of oxidized protein by reducing its degradation as well as facilitates protein oxidation by increasing steady-state level of oxidative stress [3]. Untreated diabetics with elevated glucose levels suffer many manifestations of accelerated aging, such as impaired wound healing, obesity, cataracts, vascular and microvascular damage [5]. It is important to stress that hyperinsulinemia is an significant factor not only in aging but also in the development of cancer [5,13-18].
The concept of CR mimetics is now being intensively explored [9-12]. CR mimetics involve interventions that produce physiological and anti-aging effects similar to CR. It was suggested to use biguanide antidiabetics as a potential anti-aging treatment [5,13-18]. The anti-diabetic drugs, phenformin and buformin, were observed to reduce hyperglycemia and produce the following effects: improved glucose utilization; reduced free fatty acid utilization, gluconeogenesis, serum lipids, insulin and IGF-1, and reduced body weight both in humans and experimental animals [5,16-19]. The use of phenformin in humans has been limited the last two decades because of a potential association with lactic acidosis. Widely used antidiabetic biguanide, metformin (dimethylbiguanide) does not increase risk for lactic acidosis or for increased lactate levels in type 2 diabetes [20] but has some adverse effects, including renal insufficiency in some patients [21], vitamin B12 deficiency [22], and gastrointestinal disturbances [23]. In transgenic female HER-2/neu mice, it was shown that metformin slowed down aging and tumor development [24,25]. However, in female SHR mice metformin significantly increased life span but failed to inhibit spontaneous tumorigenesis [26]. Recently it was shown that treatment with metformin did not influence life span of male F344 rats [27]. To test strain and gender factors in present study we evaluated effects of metformin on some parameters of aging, life span and spontaneous tumorigenesis in inbred male and female 129/Sv mice.
Results
Age-related body weight dynamics
Male 129/Sv mice were heavier than females over the life span both in the control and treated with metformin groups (Fig. 1). The body weight of mice in both control and metformin-treated groups increased with age, exceeding at 18 months the body weight of 3-month-old animals by 27.7% in the control female group, and by 30.5 % in the female group treated with metformin. The corresponding indices for males were 14.8% and 12.2%. Thereafter the body weight decreased both in male and female mice. On the other hand, there was no difference in the mean body weight of mice exposed and non-exposed to the drug until the age of 22 months while afterwards a tendency to the decrease of body weight was observed in metformin treated group (Fig. 1).
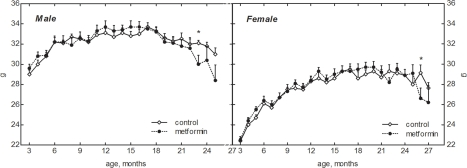
Figure 1. Dynamics of body weight in male and female 129/Sv mice treated or non-treated with metformin
Age-related dynamics of food and water consumption
Food consumption gradually increased after the age of 15 months both in female and male mice. The amount of food daily consumed by male mice was higher than in females throughout the whole period of observation (Fig. 2). It was approximately similar in the controls and in metformin-treated male and female groups until the age of 1 year. However, at the age of 15 to 21 months the food consumption was slightly but significantly (p<0.05) decreased in metformin-treated male group as compared with respective control (Fig. 2). The amount of water consumed by mice varied during the period of observation and was slightly increased by 10-20% in metformin-treated group practically at all ages (Fig. 3). Again (like in case of food consumption), males drank more water than females in both control and metformin groups.
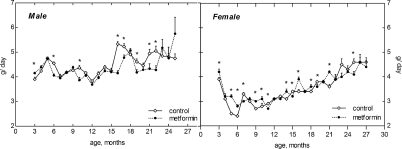
Figure 2. Dynamics of food consumption in male and female 129/Sv mice treated or non-treated with metformin
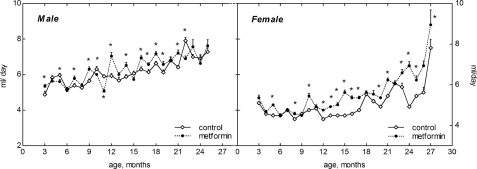
Figure 3. Dynamics of drinking water consumption in male and female 129/Sv mice treated or non-reated with metformin
Metabolic and hormonal parameters in mice treated and non-treated with metformin
No any difference was revealed in the levels of glucose, total cholesterol, triglycerides and insulin between 21-month-old male mice in control and metformin-treated group (Table 2).
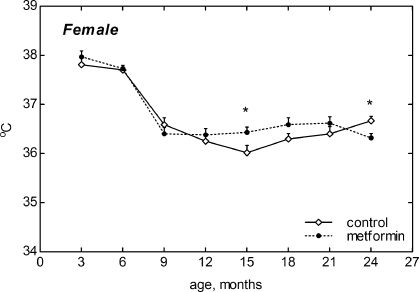
Figure 4. Dynamics of body temperature in female 129/Sv mice treated or non-treated with metformin
Table 3.
Effect of metformin on the age-related dynamics of the chromosome aberrations (ChA) incidence in bone marrow cells in male 129/Sv mice
| Age, months | Treatment | Total incidenceof ChA, % | Single bridges, % | Multiple bridges, % | Fragments, % | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 4 | Control | 15.4 ± 0.02 | 8.1 ± 0.01 | 5.3 ± 0.02 | 2.0 ± 0.01 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7 | Control | 19.1 ± 0.01 | 8.0 ± 0.01 | 11.1 ± 0.01 | 0 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Metformin | 27.3 ± 0.01* | 14.1± 0.02* | 12.1 ± 0.01 | 1.1 ± 0.01 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 18 | Control | 23.0 ± 0.06 | 7.2 ± 0.02 | 15.30 ± 0.04 | 0.5 ± 0.01 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Metformin | 28.0 ± 0.05* | 15.1±0.02 * | 12.50 ± 0.02 | 0.4 ± 0.01 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 22 | Control | 28.9 ± 0.02 | 8.5 ± 0.01 | 18.0 ± 0.02 | 2.4±0.01 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Metformin | 34.8± 0.03* | 20.3± 0.06* | 12.2 ± 0.03* | 2.3 ± 0.01 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| The difference with the control of the same age is significant: | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| - p<0,001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The total incidence of the chromosome aberrations in bone marrow of both control and treated mice increased with age. As compared with controls the chromosome aberrations value was increased in mice treated with metformin at the age 7, 18 and 22 months (p<0.001) (Table 3). Of note, this increase was related to the increase of incidence of single bridges whereas the incidence of multiple bridges was significantly decreased in 22-month-old metfromin-treated mice as compared with control mice of the same age.
Survival and longevity of 129/Sv mice
According to the log-rank test [28] the difference in survival of both male and female 129/Sv mice exposed to metformin treatment, compared to the control group, is non-significant (p-value is 0.359 for males and 0.652 for females). Survival dynamics in control and metformin-treated male and female mice are shown in Fig. 5, A and C. It is worthy of note that though there is no difference in life span distributions between male and female controls (p-value is 0.571, χ2 = 0.3 on 1 df) metformin treatment evidently produced somewhat different effect on survival of male and female mice, since the difference in life span distributions of males and females under metformin action became statistically significant (p-value is 0.0474, χ2 = 3.9 on 1 df).
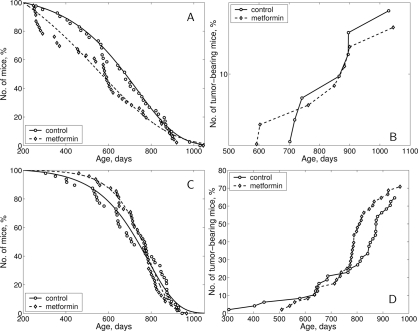
Figure 5. Survival curves and tumor yield curves of 129/Sv mice treated or non-treated with metformin (A) - survival curves, males; (B) - tumor yield curves, males; (C) - survival curves, females; D - tumor yield curves, females.
In males, metformin treatment significantly decreased the mean life span of all mice (−13.4%, p < 0.05) but not so in long-living individuals (last 10% of survivors) as well as it did not influence maximum life span (Table 4). In females, the mean life span and median of this parameter in metformin-treated animals were slightly increased (+4.4% and 7.8% respectively, p>0.05) in comparison to the controls, whereas mean life span of last 10% survivors and maximum life span were the same as in the control (Table 5). However, it deserves mentioning that until the age of 700 days survived 54.1% of mice in the control female group and 72.9% of metformin-treated females (p<0.03; Fischer's exact test).
Parameter α of the Gompertz model, which is interpreted as the rate of aging, was lower (by 1.67 times) in the male group subjected to metformin treatment than in controls males, whereas the mortality rate doubling time (MRDT) was increased in metformin-treated group of male mice (Table 4). In females, the parameter α was increased by 1.43 times and MRDT was decreased as compared to the relevant controls (Table 5).
According to the estimated parameters of the Cox's regression model being a female under metformin treatment decreases the relative risk of death compared to the male group kept under the same treatment. Cox's regression model parameters for all mice (males and females) was estimated as follows: β = −0.415; exp(β) = 0.66; se(β) = 0.211; p = 0.0504.
Table 4.
Effect of metformin on life span and tumorigenesis in male 129/Sv mice
| Parameters | Control | Metformin | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of mice | 41 | 46 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Effective number of mice | 25 | 22 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mean life span, days (M±S.E.M.) | 662±30.0 | 573±33.7* (−13.4%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mediana, days | 680 | 586 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mean life span of last 10% survivors, days | 951±32.5 | 931±28.7 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Maximum life span, days | 1029 | 1044 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| α×10−3, days−1 | 5.19 (4.37; 6.27) | 3.10 (2.42; 3.48)* | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MRDT, days | 134 (111; 159) | 224 (199; 287)* | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Number of tumor-bearing mice (%) | 7 (28.0%) | 7 (31.8%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Number of malignant tumor-bearing mice (%) | 5 (20.0%) | 5 (22.7%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mean life span of tumor-bearing mice, days | 835±45.2 | 804±62.1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Total number of tumors | 11 | 8 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Number of malignant tumors | 6 | 5 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Localization and type of tumors | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Skin: carcinoma | 1 | - | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Liver: haemangioma | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| hepatocellular carcinoma | 2 | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Lung: adenoma | - 3 | 2 2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| adenocarcinoma | 3 | 2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Spleen: leukemia | - | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Colon: polyp | 1 | - | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| adenocarcinoma | - | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Prepucial gland: cystadenoma | 2 | - | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Harderian gland: cystadenoma | 1 | - | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Notes: Difference with the controls is significant: | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| - p<0,05. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| α - aging rate; MRDT - mortality rate doubling time, days (in brackets - 95% confidential interval) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Spontaneous tumor development in 129/Sv mice
The dynamics of age-related increase in spontaneous tumor development is represented at the Fig. 5, B and D. The mean latent period of tumor development was similar in male and female mice of both groups (Tables 4 and 5). According to the long-rank test [28] there were no significant differences in age-related distributions of the total tumors occurrence in control and metformin-treated groups of both sexes.
The first tumor-bearing male mouse died at the age of 592 days in the metformin-treated group and at the 702 days in the control male group. Total tumor incidence in effective control male mice (survived by the time of the death from the first tumor in the experiment) was 28.0% and 31.8% in the metfromin-treated animals (Table 4). There were no significant differences in the incidence of malignant or benign tumors of any localization between the control and metformin-treated male mice.
In the female mice, the total tumor incidence was similar in the control and metformin-treated groups. However, the incidence of malignant tumors was significantly decreased (by 3.5 times) in the group given metformin in comparison to the control (Table 5). Benign vascular tumors of the uterus and ovary developed most frequently in female 129/Sv mice (Table 5), in line with oncological characteristics of this strain of mice [29], and were discovered more often in metformin-exposed animals. Thus, in toto uterine and ovarian haemangiomas and haemangioendoteliomas were revealed respectively in 54.2% of control and 68.8% of metformin-exposed female mice, p < 0,03 (Fisher's exact test). At the same time the differences in the incidence of angiogenic tumors only of ovaries or utery between treated and untreated with metformin was not statistically significant.
There was no significant difference in the incidence of any other tumors between mice treated or not with metformin.
Table 5.
Effect of metformin on life span and tumorigenesis in female 129/Sv mice
| Parameters | Control | Metformin | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of mice | 48 | 48 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Effective number of mice | 48 | 48 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mean life span, days (M±S.E.M.) | 711± 24.3 | 742 ± 15.8 (+4.4%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Median, days | 715 | 771 (+7.8%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mean life span of last 10% survivors, days | 910 ± 8.9 | 913 ± 19.2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Maximum life span, days | 945 | 966 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| α×10−3, days−1 | 7.27 (6.36; 8.20) | 10.40 (8.90;12.50)* | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MRDT, days | 95 (84;109) | 67 (56; 78)* | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Number of tumor-bearing mice (%) | 31 (64.6%) | 34 (70.8%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Number of malignant tumor-bearing mice (%) | 7 (15.6%) | 2 (4.2%)* | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mean life span of tumor-bearing mice, days | 766 ± 27.3 | 764 ± 18.4 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Total number of tumors | 46 | 50 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Number of malignant tumors | 9 | 2* | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Localization and type of tumors | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Uterus: haemangioma & haemangioendotelioma adenocarcinoma sarcoma | 23 2 1 | 30 1 - | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ovary: granulesa-cell tumor haemangioma & haemangioendothelioma cystadenoma | 3 7 2 | 1 11 5 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Lung: adenoma adenocarcinoma | 2 4 | 1 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Liver: hepatocellular carcinoma | 1 | - | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Haematopoietic tissue: leukemia | 1 | - | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Note: Difference with the control is significant: | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| - p<0,005. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| α - aging rate; MRDT - mortality rate doubling time, days (in brackets - 95% confidential interval). | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Discussion
In this paper for the first time were shown gender differences in effect of metformin on survival, life span and spontaneous carcinogenesis in 129/Sv mice. The reasons for these differences are at present unclear. There are several possibilities explaining these observations. One of them could be fundamental differences in mechanism of aging in males and females, another - gender peculiarities in the targets of drugs, including metformin. This question needs a special discussion.
Table 6.
Effect of antidiabetic drugs on life span and spontaneous carcinogenesis in rodents
| Species | Strain | Sex | Drug | Effect on mean life span | Effect on maximum life span | Effect on spontaneoustumor developmentc | References | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mice | C3H/Sn | F | Phenformin | +21.8% | +26.0% | ↓ | [13] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FVB/Na | F | Metformin | +8.0% | +16.2% | ↓ | [24] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FVB/Na | F | Metformin | +6.7% | -19.3% | ↓ | [25] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SHR | F | Metformin | +37.9% | +10.3% | = | [26] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NMRI | F | Diabenol | +6.7% | +1.4% | ↓ | [30] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FVB/Na | F | Diabenol | 0 | +6.0% | = | [30] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HDb | M | Metformin | +20.1% | +18.5% | N.D.d | [31] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HDb | F | Metformin | 0 | 0 | N.D.d | [31] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 129/Sv | F | Metformin | +4.4% | +7.8% | ↓ malignant. ↑ benign | Present paper | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 129/Sv | M | Metformin | -13.4% | +1.5% | = | Present paper | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Rat | LIO | F | Phenformin | 0 | 9.8% | ↓ | [15] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LIO | F | Buformin | +7.3% | +5.5% | ↓ | [15] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F344 | M | Metformin | +2.4% | 0 | N.D.d | [27] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Note: | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FVB/N mice transgenic HER-2/neu gene; | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HD, transgenic mouse model of Huntington's disease (R6/2 line with ∼ 150 glutamine repeats); | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ↓ Decrease in incidence and/or increase in tumor latency; = no effect; | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| No data. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Several years ago, it was originally suggested to use antidiabetic biguanides as mimetics of caloric restriction (CR) and a potential anti-aging treatment [5]. In a number of studies it was shown that treatment with biguanides (phenformin, buformin and metformin) increased life span and suppressed spontaneous tumorigenesis, however, in some experiments it failed to modify these parameters (Table 6). The causes of the observed contradictions are not completely clear. One of them could be differences in a dose of the drug, however, it is worthy of note that male and female 129/Sv mice demonstrated different reaction to the same dose of metformin. Also, it deserves mentioning that metformin failed to increase life span of male F344 rats [27].
Inhibitor of mammalian target of rapamycin (mTOR) increased life span of female in larger extend than in male hybrid mice [33]. It is worthy to note, the mean life span extension was observed in female (+20.4%) but it was not increased in male S6K1−/−mice [34]. Deletion of ribosomal S6 protein kinase 1 (S6K1), a component of the nutrient-responsive mTOR signaling pathway, led to increased life span in mice and to motor dysfunction and loss of insulin sensitivity [34]. Deletion of S6K1 induced gene expression patterns similar to those seen in CR or with pharmacological activation of adenosine monophosphate (AMP)-activated protein kinase (AMPK), a conserved regulator of the metabolic response to CR. The mean life span of oldest 10% survivors and maximum life span were also increased only in females. There was no difference in the incidence of macroscopic tumors in S6K1−/− and wild type mice.
Comparison of effects of metformin on various biomarkers of aging in different strains of mice shows rather notable similarity in observed patterns (Table 7). Thus, the body weight and food consumption were unchanged or slightly decreased at some periods of life in metformin-treated rodents. The body temperature was similar in control and metformin-treated mice. In females of 3 strains (129/Sv, HER-2/neu and SHR), the slow down of age-related disturbances in estrous function have been observed. It is worthy of note that in women with polycystic ovary syndrome metformin improves menstrual regularity, leading to spontaneous ovulation, and enhances the induction of ovulation with clomiphene citrate [35].
Table 7.
Effect of metformin on biomarkers of aging in rodents of various strains
| Strain, species: | 129/Sv mice | SHR mice | HER-2 mice | F344 rats | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Sex: | M | F | F | F | F | M | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| References: | Present paper | [26] | [24] | [25] | [27] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Body weight | ↓ | = | =↓ | = | ↓ | = | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Food consumption | ↓ | = | = | ↓ | = | = | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Body temperature | ND | ↑ | = | = | = | = | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Estrous function | ↑ | ↑ | ↑ | = | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tumor development | = | ↓ malignant ↑ benign | = | ↓ | ND | ND | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Glucose | = | ND | = | ↓ | = | ND | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Triglycerides | = | ND | ↓ | ↓ | ND | ND | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Cholesterol | = | ND | = | = | ND | ND | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Insulin | = | ND | = | = | = | ND | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Life span | ↓ | ↑ | ↑ | ↑ | = | = | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Notes: ND - not detected; ↓ - decreased; ↑ - increases; = no effect. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Treatment with metformin failed influence metabolic parameters in male 129/Sv mice and was followed by some improvements in female SHR and HER-2/neu mice. Metformin inhibited tumorigenesis in female 129/Sv and HER-2/neu mice and did not affect it in male 129/Sv and female SHR mice. In female 129/Sv mice treatment with metformin inhibited development of malignant tumors and increased the total incidence of benign angiogenic tumors. These data are in according with observations that metformin can to increase VEGF expression, intratumoral microvascular density and reduced necrosis thus promote the angiogenic phenotype and increased tumorigenic progression [36].
The incidence of chromosome aberrations in male 129/Sv mice was significantly higher at young, middle and old age than that in male CBA, SHR, FVB/N, C57BL/6J mice of the same age groups [37]. Treatment with metformin increased the incidence of chromosome aberrations in male 129/Sv mice that was in agreement with the observed reduction of life span in males exposed to the drug. The comparative study on the effect of metformin on female and male F344 rats would be important for understanding gender differences in response to metformin in rodents.
It is worthy to note, that experiments in yeast and C.elegans show that the life extension by CR is not a mechanical output of low calories and consequence of a reduction in ROS or AGE formation, but a process that is highly regulated, triggering metabolic shift toward respiration that activates the regulator SIR2 [38]. It was observed that phenformin inhibits proliferation and induced enhanced and transient expression of the cell cycle inhibitor p21 and apoptosis in human tumor cells lines [39].
Buformin was supplemented to nutrient medium in various concentrations (from 1.0 to 0.00001 mg/ml) during the larvae stage and over the life span of C. elegans. The drug given at the concentration of 0.1 mg/ml increased the mean life span of the worms by 23.4% (p < 0.05) and the maximum life span by 26.1% as compared to the controls [40]. Metformin supplementation (50 mM dose) was shown to increase the mean life span, but not maximum, of C. elegans, although 10 or 100 mM doses showed no significant life span benefit [41]. The authors have shown that metformin prolongs nematode healthspan, slowing lipofuscin accumulation, extending mean life span, and prolonging youthful locomotor ability in a dose-dependent manner. Genetic data suggest the metformin acts through a mechanism similar to that operative in eating-impaired CR mutants, but independent of insulin signaling pathway. Energy sensor AMPK and AMPK-activating kinase LKB1, which are activated in mammals by metformin treatment [42,43], are essential for health benefits in C. elegans, suggesting that metformin engages a metabolic loop conserved across phyla. It was also shown that metformin activated SKN-1/Nrf2, oxidative stress-responsive transcription factor [41].
Thus, biguanides could be potent geroprotectors and anticarcinogens. There are several suggestions on the mechanism of anti-carcinogenic and anti-tumor effects of metformin. Firstly, metformin is commonly considered to function as a sensitizer to insulin [5,43-46]. The antidiabetics biguanides inhibit fatty acid oxidation, suppress gluconeogenesis in the liver, increase the availability of insulin receptors, inhibit monoamine oxidase [44], increase sensitivity of hypothalamo-pituitary complex to negative feedback inhibition, reduce excretion of glucocorticoid metabolites and dehydroepiandrosterone-sulfate [5]. It was shown that metformin decreases platelet superoxide anion production in diabetic patients [47]. The capacity of metformin to activate AMPK (AMP-activated protein kinase) has been suggested to constitute an insulin-independent mechanism of inhibitory effect of metformin on proliferation of cancer cells, directly connected with promotion of the inhibition of the AMPK down-stream mammalian effector target of rapamycin (mTOR) [48-53].
mTOR controls cell growth and metabolism in response to nutrients (e.g., amino acids), insulin and growth factors such as IGF-1. Since TOR is activated by nutrients and insulin, calorie restriction deactivates TOR. Calorie restriction extends life span in a variety of species including primates [12,54]. Recently it was demonstrated that rapamycin like metformin extends life span in mice [33] and delays the development of mammary carcinomas in HER-2/neu transgenic mice [25]. Both rapamycin and biguanides inhibit carcinogenesis in rodents [25,46,55-59].
Our results alongside with recent findings of mTOR signaling pathway involvement in regulation of aging [34] and evidence of significant life span extension of mammals with rapamycin and calorie restriction [33,54] suggest that mimetic of calorie restriction, antidiabetic biguanide metformin may be rapidly contemplated for pharmacological intervention at a population level.
Materials and Methods
Animals.
Inbred 129/Sv mice were breed at the animal facility of N.N. Petrov Research Institute of Oncology. The mice were kept 5-7 in polypropylene cages (30 x 21 x 10 cm) under standard light/dark regimen (12 hours light :12 hours darkness) at 22 ± 2 °C, and received standard laboratory chow [60] and tap water ad libitum.
Experimental design.
Eighty seven male and 96 female 129/Sv mice at the age of 3 months were randomly divided into two groups. Mice of the first group were given metformin (1,1- Dimethylbiguanide hydrochloride, Biomedicals, France) with drinking water (100 mg/kg) daily, whereas the mice of the second group were given tap water without metformin and served as a control. This dose of metformin is similar to used in our earlier experiments with transgenic HER-2/neu and outbred SHR mice [24-26] and equal to 300 mg/m2 of the surface area. Recalculation for humans gives in average 510 mg/m2, that much less than commonly used in clinical practice (1.0 - 2.5 g per day). Once a week all mice were palpated for detection of tumor mass appearance. Once a month all mice were weighted and, simultaneously, the amount of consumed food and water was measured, and the rate of the consumed water (ml) and food (g) per mouse were calculated. Once in every 3 months, daily for 2 weeks vaginal smears of the animals were cytologically examined to estimate the estrous function. In the same period, rectal body temperatures of female mice were measured with an electronic thermometer, TPEM (KMIZ, Russia).
At the age of 21 month 10 male mice from the control group and the group treated with metformin were sacrificed by decapitation after overnight starvation. Samples of serum were obtained and stored at the −20°C for subsequent analyses. Other animals were observed until their natural deaths. The date of each death was registered, and the mean life span, the age at which 90% of the animals died, and the maximum life span were estimated.
Metabolic and hormonal assays.
The serum levels of glucose were estimated by enzymocolorimetric (glucose-oxidase) method with kits from “Impact” (Moscow, Russia); cholesterol and triglycerides - by enzymocolorimetric method with kits of “Olvex” (St.Petersburg, Russia); insulin - by immune enzyme assay (ELISA) with kits from Diagnostic Systems Laboratories, Inc. (U.S.A.).
Cytogenetic study.
Chromosomal aberrations in bone marrow cells was studied by modified Ford's method described in Rosenfeld et al.[37]. Mice were sacrificed with ether anaesthesia. Both femurs of each mouse were dissected and bone marrow cells flushed gently with 0.56% KCl solution into a centrifuge tube. Cells were treated for 20 min with hypotonic solution and fixed with ethanol: acetic acid mixture (3:1). Slides were stained with 4% acetoorseine. 20-30 well spread anaphases were analyzed for each animal and cells with chromosome breaks, acentric fragments, and other aberrations were evaluated on 1,000X magnification with a light microscope (Leitz, Germany).
Pathomorphological examination.
All animals were autopsied. Site, number and size of mammary tumors and their metastases in lungs were checked. All tumors, as well as the tissues and organs with suspected tumor development were excised and fixed in 10% neutral formalin. After the routine histological processing the tissues were embedded into paraffin. 5-7 μm thin histological sections were stained with haematoxylin and eosine and were microscopically examined. Tumors were classified according to International Agency for Research on Cancer recommendations [61].
Statistics.
Experimental results were statistically processed by the methods of variation statistics with the use of STATGRAPH statistic program kit. The significance of the discrepancies was defined according to the Student t-criterion, Fischer exact method, χ2, non-parametric Wilcoxon-Mann-Whitney and Friedman RM Anova on Ranks. Student-Newman-Keuls method was used for all pairwise multiple comparisons. Coefficient of correlation was estimated by Spearman method [62]. Differences in tumor incidence were evaluated by the Mantel-Haenszel log-rank test.
Parameters of Gompertz model were estimated using maximum likelihood method, non-linear optimization procedure [63] and self-written code in 'Matlab'; confidence intervals for the parameters were obtained using the bootstrap method [64].
For experimental group the Cox regression model [65] was used to estimate relative risk of death and tumor development under the treatment compared to the control group: h(t, z) = h0(t) exp(zβ), where h(t,z) and h0(t) denote the conditional hazard and baseline hazard rates, respectively, β is the unknown parameter for treatment group, and z takes values 0 and 1, being an indicator variable for two samples − the control and treatment group.
Acknowledgments
This article was supported in part by grant #08-04-00116-a from the Russian Foundation for Basic Research.
Conflicts of Interest
The authors of this paper declare no conflict of interests.
References
- 1. Fontana L, Partridge L, Longo VD. Extending healthy life span - from yeast to humans. Science. 2010; 328: 321 -326. [PubMed] .
- 2. Bartke A, Chandrashekar V, Dominici F, et al. Insulin-like growth factor 1 (IGF-1) and aging: controverses and new insights. Biogerontology. 2003; 4: 1 -8. [PubMed] .
- 3. Facchini FS, Hua NW, Reaven GM, Stoohs RA. Hyperinsulinemia: the missing link among oxidative stress and age-related diseases? Free Radicals Biol Med. 2000; 29: 1302 -1306. .
- 4. Elahi D, Muller DC, Egan JM, Andres R, Veldhuist J, Meneilly GS. Glucose tolerance, glucose utilization and insulin secretion in aging. Novartis Found. Symp. 2002; 242: 222 -242. [PubMed] .
- 5. Dilman VM. Development, Aging and Disease. A New Rationale for an Intervention Chur Harwood Academic Publishers 1994; .
- 6. Colangelo LA, Gapstur SM, Gann PH, Dyer AR, Liu K. Colorectal cancer mortality and factors related to the insulin resistance syndrome. Cancer Epidemiol. Biomarkers Prev. 2002; 11: 385 -391. .
- 7. Gupta K, Krishnaswamy G, Karnad A, Peiris AN. Insulin: a novel factor in carcinogenesis. Am J Med Sci. 2002; 323: 140 -145. [PubMed] .
- 8. Pollak MN, Schernhammer ES, Hankinson SE. Insulin-like growth factors and neoplasia. Nature Rev Cancer. 2004; 4: 505 -518. [PubMed] .
- 9. Hadley EC, Dutta C, Finkelstein J, Harris TB, Lane MA, Roth GS, Sherman SS, Starke-Reed PE. Human implications of caloric restriction's effect on laboratory animals: an overview of opportunities for research. J Gerontol Ser A. 2001; 56A (Special issue I): 5 -6. .
- 10. Ingram DK, Zhu M, Mamczarz J, Zou S, Lane MA, Roth GS, deCabo R. Calorie restriction mimetics: an emerging research field. Aging Cell. 2006; 5: 97 -108. [PubMed] .
- 11. Mattson MP, Duan W, Lee J, et al. Progress in the development of caloric restriction mimetic dietary supplements. J Anti-Aging Med. 2001; 4: 225 -232. .
- 12. Spindler SR. Caloric restriction: From soup to nuts. Ageing Res Rev. 2010; 9: 324 -353. [PubMed] .
- 13. Dilman VM and Anisimov VN. Effect of treatment with phenformin, dyphenylhydantoin or L-DOPA on life span and tumor incidence in C3H/Sn mice. Gerontology. 1980; 26: 241 -245. [PubMed] .
- 14. Anisimov VN. Carcinogenesis and Aging 2: Boca Raton CRC Press 1987; .
- 15. Anisimov VN, Semenchenko AV, Yashin AI. Insulin and longevity: antidiabetic biguanides as geroprotectors. Biogerontology. 2003; 4: 297 -307. [PubMed] .
- 16. Berstein LM. Clinical usage of hypolipidemic and antidiabetic drugs in the prevention and treatment of cancer. Cancer Lett. 2005; 224: 203 -212. [PubMed] .
- 17. Berstein LM. Modern approach to metabolic rehabilitation of cancer patients: biguanides (phenformin and metformin) and beyond. Future Oncol. 2010; 6: 1313 -1323. [PubMed] .
- 18. Berstein LM. Biguanides: An Expansion to Practical Oncology (past and present) St.Petersburg Aesculap 2010; 144 .
- 19. Blagosklonny MV. Validation of anti-aging drugs by treating age-related diseases. Aging. 2009; 1: 281 -288. [PubMed] .
- 20. Kruse JA. Review: Metformin does not increase risk for lactic acidosis or increase lactate levels in type 2 diabetes. ACP J Club. 2004; 141: 147 .
- 21. Nisbet JC, Strurtevant JM, Prins JB. Metformin and serious adverse effects. Med J Aust. 2004; 180: 53 -54. [PubMed] .
- 22. Vidal-Alaball J and Butler CC. Reduced serum vitamin B-12 in patients taking metformin. BMJ. 2010; 340: c2198 https://doi.org/10.1136/bmj.c2198 [PubMed] .
- 23. Krentz AJ, Ferner RE, Balley CJ. Comparative tolerability profiles of oral antidiabetic agents. Drug Safety. 1994; 11: 223 -241. [PubMed] .
- 24. Anisimov VN, Berstein LM, Egormin PA, Piskunova TS, Popovich IG, Zabezhinski MA, Kovalenko IG, Poroshina TE, Semenchenko AV, Provinciali M, Re F, Franceschi C. Effect of metformin on life span and on the development of spontaneous mammary tumors in HER-2/neu transgenic mice. Exp Gerontol. 2005; 40: 685 -693. [PubMed] .
- 25. Anisimov VN, Egormin PA, Piskunova TS, Popovich IG, Tyndyk ML, Yurova MN, Zabezhinski MA, Anikin IV, Karkach AS, Romanyukha AA. Metformin extends life span of HER-2/neu transgenic mice and in combination with melatonin inhibits growth of transplantable tumors in vivo. Cell Cycle. 2010; 9: 188 -197. [PubMed] .
- 26. Anisimov VN, Berstein LM, Egormin PA, Piskunova TS, Popovich IG, Zabezhinski MA, Tyndyk ML, Yurova MN, Kovalenko IG, Poroshina TE, Semenchenko AV. Metformin slows down aging and extends life span of female SHR mice. Cell Cycle. 2008; 7: 2769 -2773. [PubMed] .
- 27. Smith DL, Elam CF, Mattison JA, Lane MA, Roth GS, Ingram DK, Allison DB. Metformin supplementation and life span in Fischer-344 rats. J. Gerontol. Biol. Sci. 2010; 65A: 468 -474. .
- 28. Cox DR and Oakes D. Analysis of Survival Data London Chapman & Hall 1996; .
- 29. Piskunova TS, Yurova MN, Ovsjannikov AI, Semenchenko AV, Zabezhinski MA, Popovich IG, Wang Z-Q, Anisimov VN. Deficiency in poly(ADP-ribose) polymerase-1 (PARP-1) accelerates aging and spontaneous carcinogenesis in mice. Current Gerontol Geriatr Res. 2008: 2008; https://doi.org/10.1155/2008/754190 Article ID 754190, 11 pages .
- 30. Popovich IG, Zabezhinski MA, Egormin PA, Tyndyk ML, Anikin IV, Spasov AA, Semenchenko AV, Anisimov VN. Insulin in aging and cancer: new antidiabetic drug Diabenol as geroprotector and anticarcinogen. Int J Biochem Cell Biol. 2005; 37: 1117 -1129. [PubMed] .
- 31. Ma TC, Buescher JL, Oatis B, Funk JA, Nash AJ, Carrier RL, Hoyt KR. Metformin therapy in a transgenic mouse model of Hungtington's disease. Neurosci Lett. 2007; 411: 98 -103. [PubMed] .
- 32. Anisimov VN. Effect of buformin and diphenylhydantoin on life span, estrus function and spontaneous tumor incidence in female rats. Vopr Onkol. 1980; 26: 42 -48. [PubMed] .
- 33. Harrison DE, Strong R, Sharp ZD, Nelson JF, Astle CM, Flurkey K, Nadon NL, Wilkinson JE, Frenkel K, Carter CS, Pahor M, Javors MA, Fernandez E, Miller RA. Rapamycin fed late in life extends lifespan in genetically heterogenous mice. Nature. 2009; 460: 392 -396. [PubMed] .
- 34. Selman C, Tullet JMA, Wieser D, Irvine E, Lingard SJ, . Cloudhury AI, Claret M, Al-Quassab H, Carmignac D, Ramadani F, Woods A, Robisnon IC, Schuster E, Batterham RL, Kozma SC, Thomas G, Carling D, Okkenhasug K, Thornton JM, Patridge L, Gems D, Withers DJ. Ribosomal protein S6 kinase 1 signaling regulates mammalian life span. Science. 2009; 326: 140 -144. [PubMed] .
- 35. Awartani KA and Cheung AP. Metformin and polycystic ovary syndrome: a literature review. J. Obstet. Gynecol. Can. 2002; 24: 393 -401. [PubMed] .
- 36. Phoenix KN, Vumbaca F, Claffey KP. Therapeutic metformin/AMPK activation promotes the angiogenic phenotype in the ERα negative MDA-MB-435 breast cancer model. Breast Cancer Res Treat. 2009; 113: 101 -111. [PubMed] .
- 37. Rosenfeld SV, Togo EF, Mikheev VM, Popovich IG, Zabezhinski MA, Anisimov VN. Age-related dynamics of the chromosome lesion incidence in male mice of different strains. Bull Exp Biol Med. 2001; 131: 568 -570. [PubMed] .
- 38. Koubova J and Guarente L. How does calorie restriction work? Genes Dev. 2003; 17: 313 -321. [PubMed] .
- 39. Caraci F, Chisari M, Frasca G, et al. Effects of phenformin on the proliferation of human tumor cell lines. Life Sci. 2003; 74: 643 -650. [PubMed] .
- 40. Bakaev VV. Effect of 1-butylbiguanide hydrochloride on the longevity in the nematoda Caenorhabditis elegans. Biogerontology. 2002; 3: 1 23 -24. .
- 41. Onken B and Driscoll M. Metformin induces dietary restriction-like state and the oxidative stress response to extend C. elegans healthspan via AMPK, LKB1, and SKN-1. PloS ONE. 5: 1 e8758 https://doi.org/10.1371/journal.pone.0008758 [PubMed] .
- 42. Zhou G, Myers R, Li Y, Chen Y, Shen X, Fenyk-Melody J, et al. Role of AMP-activated protein kinase in mechaism of metformin action. J Clin Invest. 2001; 108: 1167 -1174. [PubMed] .
- 43. Shaw RJ, Lamia KA, Vasquez D, Koo S-H, Bardeesy N, DePinho RA, Montminy M. Cantley LC The kinase LKB1 mediates glucose homeostasis in liver and therapeutic effects of metformin. Science. 2005; 310: 1642 -1646. [PubMed] .
- 44. Muntoni S. Metformin and fatty acids. Diabetes Care. 1999; 22: 179 -180. [PubMed] .
- 45. Strack T. Metformin: a review. Drugs Today. 2008; 44: 303 -314. [PubMed] .
- 46. Anisimov VN. Metformin for aging and cancer prevention. Aging. 2010; 2: 760 -774. [PubMed] .
- 47. Gargiulo P, Caccese D, Pignatelli P, et al. Metformin decreases platelet superoxide anion production in diabetic patients. Diabetes Metab Res Rev. 2002; 18: 156 -159. [PubMed] .
- 48. Huang X, Wullschleger S, Shpiro N, McGuire VA, Sakaoto K, Woods YL, McBurnie W, Fleming S, Alessi DR. Important role of the LKB1-AMPK pathway in suppressing tumorigenesis in PTEN-deficient mice. Biochem J. 2008; 412: 211 -221. [PubMed] .
- 49. Zakikhani M, Dowling R, Fantus IG, Sonenberg N, Pollak M. Metformin is an AMP kinase-dependent growth inhibitor for breast cancer cells. Cancer Res. 2006; 66: 10269 -10273. [PubMed] .
- 50. Alimova IN, Liu B, Fan Z, Edgerton SM, Dillon T, Lind SE, Thor AD. Metformin inhibits breast cancer cell growth, colony formation and induces cell cycle arrest in vitro. Cell Cycle. 2009; 15: 8:909 -915. .
- 51. Blagosklonny MV. Aging and immortality. Quasi-programmed senescence and its phamacological inhibition. Cell Cycle. 2006; 5: 2087 -2102. [PubMed] .
- 52. Blagosklonny MV. An anti-aging drug today: from senescence-promoting genes to anti-aging pill. Drug Discovery Today. 2007; 12: 218 -224. [PubMed] .
- 53. Blagosklonny MV. Prevention of cancer by inhibiting aging. Cancer Biol Ther. 2008; 7: 1520 -1524. [PubMed] .
- 54. Colman RJ, Anderson RM, Johnson SC, Kastman EK, Kosmatka KJ, Beasley TM, Allison DB, Cruzen C, Simmons HA, Kemnitz JW, Weindruch R. Caloric restriction delays disease onset and mortality in rhesus monkeys. Science. 2009; 325: 201 -204. [PubMed] .
- 55. Schneider MB, Matsuzaki H, Harorah J, et al. Prevention of pancreatic cancer induction in hamsters by metformin. Gastroenterology. 2001; 120: 163 -170. .
- 56. Mabuchi S, Altomare DA, Connolly DC, Klein-Szanto A, Litwin S, Hoelzle MK, et al. RAD001 (Everolimus) delays tumor onset and progression in a transgenic mouse model of ovarian cancer. Cancer Res. 2007; 67: 2408 -2413. [PubMed] .
- 57. Robinson J, Lai C, Martin A, Nye E, Tomlinson I, Silver A. Oral rapamycin reduces tumour burden and vascularization in Lkb1(+/−) mice. J Pathol. 2009; 219: 35 -40. [PubMed] .
- 58. Deriabina ON, Plotnikova NA, Anisimov VN. Melatonin and metformin inhibits skin carcinogenesis induced by benzo(a)pyrene in mice. Vopr. Onkol. 2010; 56: 583 -587. .
- 59. Anisimov VN, Zabezhinski MA, Popovich IG, Piskunova TS, Semenchenko AV, Tyndyk ML, Yurova MN, Antoch MP, Blagosklonny MV. Rapamycin extends maximal life span in cancer-prone mice. Am J Pathol. 2010; 176: 1092 -1096. .
- 60. Anisimov VN, Popovich IG, Zabezhinski MA. Methods of evaluating the effect of pharmacological drugs on aging and life span in mice. Methods Mol Biol. 2007; 371: 227 -236. [PubMed] .
- 61. Turusov VS and Mohr U. Pathology of Tumours in Laboratory Animals. I: Tumours of the Mouse IARC Sci Publ 111, Lyon IARC 1994; .
- 62. Goubler EV. Computing Methods of Pathology Analysis and Recognition Leningrad Meditsina 1978; .
- 63. Fletcher R. Practical Methods of Optimization New York Wiley 1987; .
- 64. Davison AC and Hinkley DV. Bootstrap Methods and Their Application Cambridge Cambridge University Press 1997; .
- 65. Cox D. Regression models and life-tables (with discussion). J. Royal Statistical Soc. Series B (Methodological). 1972; 34: 187 -220. .