



Introduction
Stem cells are characterized by their ability to divide to generate additional stem cells and differentiated cell types. Proliferative division can be symmetric or asymmetric, thereby one stem cell generates either two daughter stem cells or one stem cell plus one differentiated cell type, thus, triggering the expansion or self-renewal of the stem cell pool, respectively. Due to these unique features, stem cells are essential during embryonic development for tissue formation and during adulthood to ensure tissue homeostasis, repair, and regeneration [1-5].
The balance between cell loss and cell replacement is brilliantly maintained in most tissues by resident stem cells up to a point when such equilibrium starts to be progressively lost and newborn cells are not able to compensate for the ones that died, resulting in decline in tissue integrity and function and a diminished capacity of regeneration upon damage [2, 6]. The physiological loss of tissue homeostasis during life is typically concomitant to a progressive and extensive decline in the physical and cognitive performance of the whole organism that is commonly referred to as aging. In addition to loss of stem cell turnover, aging is also caused by a decrease in the overall function of differentiated cells that is due to a number of cell-intrinsic and environmental factors including DNA damage, reduction in telomere length, oxidative stress, induction of stress response pathways, and production and accumulation of misfolded proteins, which may result in a number of disorders including cancer, heart failure, neurological diseases, and many others [7-10].
For humans, aging is one of the most evident biological processes whose molecular and cellular mechanisms are still poorly understood. Since recent studies in the field of stem cell biology have brought new light into the regenerative potential of this cell population, several laboratories have been prompted to investigate the use of stem cells for attenuating the effects of aging. It has been suggested that exhaustion of stem cells may be a primary cause of aging [11, 12] and, in fact, forcing regeneration leads to stem cell exhaustion and premature aging [13]. Certainly, stem cells can contribute to aging in different tissues as a reflection of their relative contribution to cell turnover and regeneration implying that organs with a low turnover should be minimally affected by aging of stem cells [3]. However, while comparing different species, the lack of an obvious correlation between the abundance of stem cells, regenerative capacity, aging, and lifespan hardly allows to make any link [3]. Moreover, recent evidences indicate that the relative abundance of stem cells in certain organs does not necessarily correlate with their impact on organ function. Specifically, the mammalian brain is perhaps the organ with the lowest regenerative potential but the one in which the signs of aging are more manifested. Using the words of the renaissance writer Michel de Montaigne, “age imprints more wrinkles on the mind than it does on the face” indicating that age-related cognitive decline has the highest impact on the quality of life. To which extent this decline is dependent on neural stem and progenitor cells (together referred to as NSCs) is hard to tell but growing evidences indicate that, despite their negligible numbers, the few resident NSCs that are located in specific brain regions, most notably the subgranular zone of the hippocampus, seem to play a major role in cognitive functions such as learning, memory, and emotional behavior by generating, through intermediate progenitors, neurons that are constantly added to the brain circuitry throughout life [14-18].
Perhaps not coincidentally, aging constitutes one of the major factors reducing the proliferation of NSCs [19-21] while cognitive decline, including a reduced learning and memory performance, is commonly observed in aged individuals [22-24]. Certainly, NSCs are not the only cause of brain aging because non-neurogenic areas, most prominently the prefrontal cortex, are known to be responsible for many phenotypes connected to senescence including forgetfulness and distractibility [25-27]. A positive correlation between NSC activity, hippocampal function, and cognitive performance during aging has been proposed for groups of animals and single individuals [28-31] but other studies have challenged this view [32-35]. Nevertheless, manipulations that decrease neurogenesis typically worsen cognitive performance in senescent animals while, conversely, an increase in neurogenesis tends to improve learning and memory [29, 36-38]. These findings in basic stem cell research during aging have led to the hope that stem cell-based approaches might be useful to compensate, at least in part, the age-related cognitive decline that is occurring in human. In this context, it is important to notice that the final amount of newborn neurons integrating in the brain circuitry during life is controlled at different levels including NSCs proliferation, cell fate change, neuronal survival, and maturation. Given the importance of understanding the biology of NSCs during aging, in this review we discuss the present knowledge about the age-related changes in neurogenesis and its underlying mechanisms at the systemic, cellular, and molecular level. Particular attention is given to the manipulations that allowed the increase of neurogenesis as a potential means to improve cognitive performance during aging.
NSCs proliferation and fate
Since the original observation of Altman and Das [19], several studies have consistently confirmed a major reduction in the abundance of proliferating NSCs during aging from rodents to primates [20, 21, 39]. In most studies, BrdU labeling, eventually in combination with markers of cycling cells, e.g.: Ki67 or PCNA, was commonly used to quantify the amount of cells undergoing cell cycle progression. Probably as a result of decreased proliferation, aging was shown to correlate with a decrease in newly generated neurons [20, 21, 39].Regarding NSCs, an important question remains as to whether the decrease in the number of proliferating cells reflects an increase in quiescence as opposed to their depletion. Moreover, an increase in the proportion of quiescent NSCs could be due to a higher number of cells leaving the cell cycle to become quiescent and/or a lower number of quiescent cells entering the cell cycle. Similarly, depletion of NSCs may be due to an increase in differentiation with concomitant loss of self-renewal and/or increased cell death (Figure 1). Discriminating between these possibilities becomes particularly important for studies aimed at compensating the age-related decline in neurogenesis though manipulations of endogenous NSCs either by controlling their cell cycle entry versus exit or, alternatively, by manipulating their cell fate change versus survival.
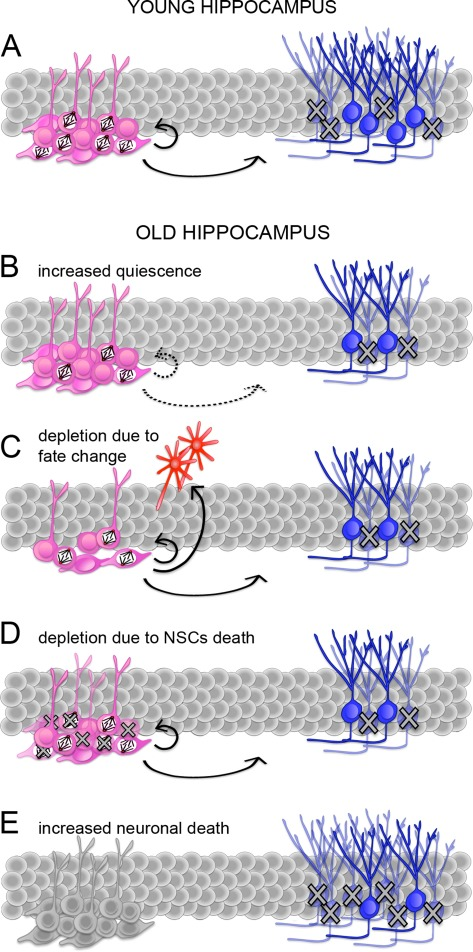
Figure 1. Potential mechanisms responsible for the age-related decline in neurogenesis (A-E) Schematic representation of adult neurogenesis in the young (A) or senescent (B-E) hippocampus. Proliferating and quiescent neural stem and progenitors cells (NSCs; pink) are indicated by the presence or lack of mitotic spindles, respectively. Proliferating NSCs divide to generate (arrows) additional NSCs or immature neurons (blue). During maturation, many neurons undergo apoptosis (crosses). A reduced number of neurons in the senescent hippocampus (B-E) can be explained by an increase in quiescence i.e. a lower proportion of proliferating NSCs (B), a change in NSCs fate i.e. an increase in differentiative at the expense of proliferative and/or gliogenic at the expense of neurogenic division (C), depletion of NSCs by cell death (D), or a higher proportion of newborn neurons undergoing apoptosis (E). Continuous or dashed arrows (A-D) indicate a constant or decreases proportion of proliferating NSCs and neurons generated from the total pool of NSCs, respectively. Note that NSCs in E are not colored because neuronal death could potentially occur concomitantly to any of the previous conditions (A-D).
Certain studies based on the expression of NSCs markers that are independent from their proliferative state indicated a strong reduction in cell numbers in the aging hippocampus of rodents [40-42]. Conversely, other studies reported a change in their proliferative state but not in their absolute numbers [43, 44]. These discrepancies can partly be explained by considering that diverse strains, transgenic models, or even species were used and, in fact, it has been reported that senescent rodents and primates are characterized by a constant or decreased number of NSC, respectively [45]. In addition, it should be considered that the different markers used to characterize stem and progenitor cell types in different studies might identify slightly different cell populations. Assuming that the effects of aging may vary among progenitor subtypes, the discrepancies observed could, thus, reflect a diverse proportion of cell subpopulations identified using any specific marker within a given species.
The dichotomy between depletion of NSCs versus their entry in quiescence during life also emerged in two recent works, each supporting one of two alternative scenarios. According to the disposable stem cell model [46], quiescent NSCs enter the cell cycle and undergo a limited number of asymmetric cell divisions after which they terminally differentiate into mature astrocytes, thus, leading to depletion of the stem cell pool during life. Clonal analyses by a second group reported a diverse behavior of NSCs with a component of depletion but, in addition, a significant proportion of quiescent as well as cycling NSCs being detected over long periods of time [47].
The contradicting results concerning depletion versus quiescence of NSCs during aging are still debated and even more confusion is caused by failing to appreciate the differences between a reversible cell cycle arrest and a permanent loss of proliferative potential, in which only the former can properly be defined to as cellular senescence [48]. Nevertheless, a number of studies have consistently shown that proliferation of NSCs in old animals can be induced under certain conditions, including physical activity [38, 49-51] and disease [44, 52-55] indicating that, despite a partial depletion and, possibly, cellular senescence, a pool of NSCs is maintained that can be efficiently induced to re-enter the cell cycle and potentially contribute to improvement of brain function.
In addition to proliferation, NSCs activity can also be regulated at the level of a change in their fate (e.g.: by changing the proportion of cells undergoing proliferative versus differentiative, symmetric versus asymmetric, and/or gliogenic versus neurogenic divisions). Unfortunately, “decision making” in term of cell fate change as a factor accounting for the reduction in neurogenesis during aging is also debated. In fact, a decrease in the number of cells acquiring a neuronal phenotype has been proposed [36, 38] but other studies reported that even if the number of proliferating NSCs and newborn neurons decreases during aging, the proportion of differentiated cell types generated from the pool of cycling NSCs is similar or only slightly changed [35, 56-58].
Nevertheless, it should be considered that developing approaches to compensate for the age-related decline in NSC activity could be achieved at any level, including those that are not necessarily changed physiologically. Thus, it becomes important not only to identify the physiological alterations occurring in the NSC niche but also to consider the systemic, cellular, and molecular manipulations that could be used to control this process.
Systemic effects of aging on NSCs
Among the systemic changes influencing NSCs activity, age-related alteration in the vasculature seems to assume a critical role. The tight relationship and relative dependency between adult neurogenesis and the vascular niche has been consistently described [59-62]. However, it is not without a certain sense of frustration that one realizes that, similarly to NSCs quiescence, depletion, and fate, the effects of aging on the brain vasculature are also highly controversial. While no change in the volume of capillaries in the dentate gyrus has been reported [38], the opposite result has also been observed with a decrease in capillary volume and proportion of NSCs clustering in their proximity [43] and even without considering specifically the neurogenic niches conflicting reports have been alternatively published [63].
Perhaps more conclusively, aging was shown to correlate with changes in hemorheological parameters resulting in a diminished cerebral blood flow and a consequent reduced availability of oxygen, metabolites, and neurotrophic factors [64-66] that are important for NSCs and whose alterations were associated with impairment in spatial memory [67].
Availability of oxygen may constitute an important factor responsible for the reduced neurogenesis of the aged hippocampus. As it had emerged from a number of in vitro studies using embryonic-derived NSCs, reducing the atmospheric oxygen tension to physiological levels i.e. from ca. 20% to 3%, increases NSCs proliferation [68-71]. A similar effect is now being revealed in the adult brain by studies linking the activity of hypoxia-inducible transcription factors (HIF) to the induction of genes known to be involved in stem cell differentiation including Wnt, Notch, and BMP [72-75]. Interestingly, aging mice display impaired hypoxic response including a deficiency in the activation of HIF-1 downstream targets [76] and an initial delay in the angiogenic response in hypoxic conditions [77].
In addition to oxygen, systemically released factors have a prominent role in NSCs activity. A recent study showed that during heterochronic parabiosis the old mouse displays higher level of NSCs proliferation and higher number of newborn neurons due to a change in soluble factors, including chemokines, present in the plasma of the young animal [78] but no molecular link between these chemoattractants and NSCs proliferation has been established. Other blood-released factors with a role on adult neurogenesis and aging deserving a special mention include steroid hormones and growth factors such as corticosterone (CORT), insulin-like growth factor-1 (IGF-1), fibroblast growth factor-2 (FGF-2), and vascular endothelial growth factor (VEGF).
Glucocorticoids are steroid hormones involved in age-related changes in the hippocampus, whose production and secretion by the adrenal gland are regulated by the hypothalamic-pituitary-adrenal axis, mainly in response to stress [79]. The mineralcorticoid and glucocorticoid receptors, strongly expressed in granule cells of the hippocampus [80, 81], and the latter also in NSCs [82], regulate gene expression upon activation by DNA binding of the glucocorticoid response element [79]. CORT is the primary glucocorticoid in rodents that downregulates cell proliferation in the hippocampus, as shown by CORT administration or adrenalectomy experiments in both young and senescent animals [42, 83-86] in which a decrease in CORT levels was shown to correlate with improved learning and memory [29]. In old rodents, CORT basal level is enhanced but stress-induced CORT increases only in the blood stream but not in the hippocampus [87]. In addition, expression of CORT receptors in NSCs and immature neurons of the hippocampus increases during aging[82].
Contrary to CORT, the three growth factors IGF-1, FGF-2 and VEGF were shown to have very similar effects in increasing NSCs proliferation. Expression and/or concentration of these factors and their receptors is known to decrease in the hippocampus during aging [88-91] and, in particular, levels of IGF-1 were shown to be higher, and cognitive performance to be improved, in long-living mutants [92]. A substantial number of experiments were performed to study the effects of the three factors on neurogenesis during adulthood or aging indicating a strong positive effect on NSCs proliferation by IGF-1 [93-95], FGF-2 [96-100], and VEGF [101-104] signaling, with the latter also resulting in improved spatial memory functions [104]. To which extent these or other factors released in the blood can directly influence cognitive function during aging is currently debated [105, 106].
The previous findings describing the role of the vasculature, oxygen tension, and blood-released factors deserve a special emphasis in the context of the physiological stimulus known to have the strongest positive effect on NSCs proliferation: physical exercise. Physical exercise, like voluntary running, is a physiological activity known to enhance NSCs proliferation and neurogenesis in the adult hippocampus and, concomitantly, improve cognitive functions in both young [107, 108] and old [38, 49-51] animals. The increase in cell proliferation induced by running was shown to depend on the release of IGF-1 [109, 110] and on the complex VEGF-dependent effects induced at the level of the vasculature [111].
Another physiological stimulus triggering systemic changes that correlate with a decrease in stem cell exhaustion and cellular senescence while, conversely, increasing lifespan is dietary restriction [112, 113]. Effects of dietary restriction are in part mediated by an inhibition of the mammalian target of rapamycin (mTOR) leading to a decreased senescence of stem and progenitor cells [48, 113-115] and several evidences indicate that these observations can be extended also to NSC of the adult brain [116-120].
Paradoxically, the two strongest physiological stimuli known to date to increase adult neurogenesis and lifespan, physical exercise [38, 49-51]and dietary restriction [112], respectively, are the ones that current societies of the western world more consistently neglect while asking biomedical research to fill the gap. To achieve this, manipulation of the factors released by the vasculature may provide a means to better understand and intervene in the patho-physiology of the aging brain [23, 121, 122].
Cellular and molecular effects of aging on NSCs
In addition to systemic effects, NSCs proliferation can also be influenced by cellular and molecular changes taking place within the neurogenic niche (Figure 2). In fact, not only some of the factors mentioned above but also a long list of other signaling molecules and transcription factors known to regulate neurogenesis are produced locally by NSCs, astrocytes, or endothelial cells such as Notch, Wnt, BMP, Shh and several others [123-125]. Unfortunately, in the aging brain most of these factors were not investigated but it is interesting to notice that most, if not all, have been shown to inhibit differentiation and, in addition, shorten the cell cycle of NSCs [126-129]. Since the age-related reduction in NSCs proliferation is known to correlate with reduced levels of growth factors [89, 90], it would not come as a surprise if expression of genes regulating cell cycle length (or quiescence) should also be altered in the aging brain. In fact, expression of the inhibitors of G1 progression p16INK4 and p19Arf, that were previously linked to cellular senescence [130], were found to increase in the subventricular zone of old animals [131, 132] and deletion of p16INK4 was shown to promote NSC proliferation and neurogenesis in the aged subventricular zone but, apparently, not in the hippocampus [131]. Conversely, positive regulators of the cell cycle, most notably cyclinD1, were found to decrease during aging [133]. While not considering the differences that could exist within progenitors subtypes [40], measurements of cell cycle parameters in the dentate gyrus of young and 10 months old (middle-aged) rats did not detect changes in cell cycle length but only in the proportion of cycling cells [134] and similar measurements in the subventricular zone of 18 months old (senescent) mice corroborated this conclusion [135]. Interestingly, growing aged NSCs in culture was reported to shorten their cell cycle and induce proliferative, as opposed to neurogenic, divisions [135].
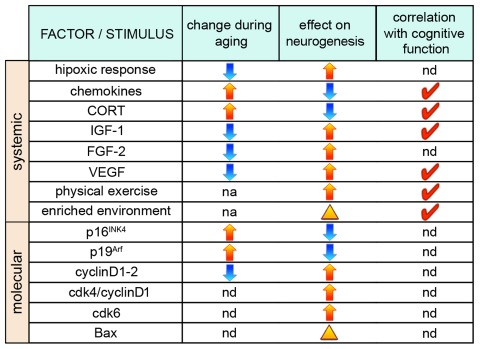
Figure 2. Factors influencing neurogenesis during aging (From left to right) Factors and stimuli influencing neurogenesis, their physiological increase (yellow arrows) or decrease (blue arrows) during aging, as well as effects on neurogenesis through, primarily, a change in NSCs proliferation (arrows) or neuronal survival (arrowhead) are indicated. Red ticks indicate factors whose manipulation has been shown to correlate with cognitive function. nd=not determined; na=not applicable.
In addition to the cell fate determinants and growth factors mentioned above, also cell cycle regulators can control the fate of NSCs during embryonic development and adulthood [126-129] and even if not physiologically responsible for the aging phenotype, manipulation of cell cycle length could still be used as a strategy to overcome the reduction in neurogenesis occurring in aged animals. An impressive number of studies on cell cycle regulators and neurogenesis were already thoroughly reviewed [126-129]. Focusing exclusively on acute manipulations that increased neurogenesis, our group has recently found that expansion of NSCs can be achieved during embryonic development [136] and adulthood[137] through overexpression of the positive regulators of G1 progression cdk4/cyclinD1. This finding is consistent with the opposite effect reported in the adult brain after deletion of similar G1 regulators such as cdk6 [138] and cyclinD2 [139] and corroborate the notion (the cell cycle length hypothesis[140]) that manipulation of G1 length may influence the fate of somatic stem cells [128, 141]. Additional studies are needed to extend the cell cycle length hypothesis from the adult [137-139] to the senescent brain but findings using aged NSCs in vitro [135] makes this possibility likely.
Neuronal maturation and survival
In addition to cell proliferation and fate, neuronal survival strongly influences the number of neurons produced in the adult hippocampus. It is known that the vast majority of newborn neurons are eliminated by apoptosis in the following 6-8 weeks and that only a small fraction is selected for long-term survival [142]. However, neuronal survival does not seem to be altered in aged animals since birthdating experiments based on the comparison of BrdU positive neurons at different times upon labeling in old and young animals showed that the proportion of neurons dying after birth is unchanged [56-58]. Migration, dendritogenesis, and expression of mature markers of newborn neurons were delayed in old animals [56] but this effect was compensated at later stages [56, 143].
Even if aging does not influence neuronal death, physiological stimuli can still be used to increase survival in senescent animals. In particular, living in an enriched environment that provides access to social and inanimate stimuli, such as toys, proved to be the strongest physiological condition increasing the survival of neurons in young and aged mice [36, 144] suggesting that the aged brain retains a certain level of neuroplasticity. Similar results were obtained after long-term exposure to enriched environment indicating an overall increase in the neuronal survival baseline rather than an acute response to new stimuli [37]. Importantly, increased neuronal survival in old animals positively correlated with a better performance in spatial memory tests [36, 37] end even if the molecular mechanism underlying this correlation is largely unknown, recent studies suggest a role of steroid hormone receptors [82].
In young mice, neuronal survival and, consequently, integration can be genetically enhanced, as recently shown after ablation of the proapoptotic gene Bax, leading to improvement in certain cognitive functions such as pattern separation [145]. As proposed by the authors of this work, promoting neuronal survival could constitute a new way to increase the number of neurons in old animals and, possibly, compensate age-related learning and memory deficits.
Concluding remarks
A substantial number of evidences indicate that age-related decrease in adult neurogenesis is an important factor influencing cognitive performance. While several mechanisms may influence the number of mature neurons functionally integrated into the brain circuitry over time, the available data strongly suggests that aging almost exclusively acts at the level of NSC proliferation. Yet, the many contradicting results and uncertainties on identifying the exact causes of this “decreased proliferation” (i.e. quiescence, cellular senescence, cell cycle lengthening, and/or depletion via cell death or fate change) need to be fully acknowledged in order to give a rigorous and meaningful direction to this relatively new field. Nevertheless, in the context of therapy, also NSC fate, neuronal survival, and integration could potentially become the focus of interventions aimed at compensating for the decline in neurogenesis occurring during aging.
In this perspective, significant resources are invested in stem cell research in the hope that basic knowledge could one day be used for developing treatments of age-related cognitive decline and therapy of neurodegenerative diseases [146-148]. In this frame, it is interesting to notice that regulation of NSCs proliferation, although not necessarily associated with an increase in the number of integrated neurons, constitute a physiological response to certain diseases and that this response is maintained in aged rodents upon seizure [44, 52] and ischemia [53-55]. At least for stroke, a proliferative response has been observed in non-neurogenic areas also in human [149-151].
The fact that NSCs can efficiently respond to physiological and pathological stimuli to increase neurogenesis indicates that stimulation of endogenous NSCs offers a promising alternative to transplantation approaches that until now were intensely investigated but with very limited success [146-148, 152]. Proof of principle that increased neurogenesis in the adult hippocampus by acute manipulation of NSCs proliferation and fate [137] or neuronal survival [145] has been provided. Certainly, these reports fell short from demonstrating that a similar strategy can be applied in senescent mice to improve cognitive performance. Moreover, the studies discussed here indicate that a diminished neurogenesis during aging is primarily due to an increased quiescence and/or depletion of NSCs and since cdk4/cyclinD1 apparently changes the fate of NSCs but not their quiescence [137], while Bax knock-out inhibits neuronal death without increasing NSCs proliferation [145], it becomes unlikely that either of the two approaches would alone be sufficient to significantly improve cognitive function in senescent mice.
The strong positive correlation between increased endogenous neurogenesis and improved cognitive performance in senescent mice has already been provided upon life-long manipulation of blood-released factors [29] or acute exposure to physiological stimuli [38, 49-51]. It remains to be shown whether these effects can be reproduced by more specific manipulations triggering the appropriate combination of changes at the level of NSCs quiescence, fate, neuronal survival, and/or integration to improve cognitive performance in senescent individuals; even despite a lack of dietary moderation and physical exercise.
Acknowledgments
FC and BA are supported by the DFG-funded Center for Regenerative Therapies, the Medical Faculty of the Technical University Dresden, the Max Planck Institute of Molecular Cell Biology and Genetics and the DFG-Collaborative Research Center SFB655 (subproject A20).
Conflicts of Interest
The authors of this manuscript have no conflict of interest to declare.
References
- 1. Weissman IL. Stem cells: units of development, units of regeneration, and units in evolution. Cell. 2000; 100: 157 -168. [PubMed] .
- 2. Rossi DJ, Jamieson CH, Weissman IL. Stems cells and the pathways to aging and cancer. Cell. 2008; 132: 681 -696. [PubMed] .
- 3. Rando TA. Stem cells, ageing and the quest for immortality. Nature. 2006; 441: 1080 -1086. [PubMed] .
- 4. Fuchs E. The tortoise and the hair: slow-cycling cells in the stem cell race. Cell. 2009; 137: 811 -819. [PubMed] .
- 5. Morrison SJ and Kimble J. Asymmetric and symmetric stem-cell divisions in development and cancer. Nature. 2006; 441: 1068 -1074. [PubMed] .
- 6. Liu L and Rando TA. Manifestations and mechanisms of stem cell aging. J Cell Biol. 2011; 193: 257 -266. [PubMed] .
- 7. Hoeijmakers JH. DNA damage, aging, and cancer. N Engl J Med. 2009; 361: 1475 -1485. [PubMed] .
- 8. Haigis MC and Yankner BA. The aging stress response. Mol Cell. 2010; 40: 333 -344. [PubMed] .
- 9. Kirkwood TB. Understanding the odd science of aging. Cell. 2005; 120: 437 -447. [PubMed] .
- 10. Balaban RS, Nemoto S, Finkel T. Mitochondria, oxidants, and aging. Cell. 2005; 120: 483 -495. [PubMed] .
- 11. Sharpless NE and DePinho RA. How stem cells age and why this makes us grow old. Nat Rev Mol Cell Biol. 2007; 8: 703 -713. [PubMed] .
- 12. Sahin E and Depinho RA. Linking functional decline of telomeres, mitochondria and stem cells during ageing. Nature. 2010; 464: 520 -528. [PubMed] .
- 13. Ruzankina Y, Pinzon-Guzman C, Asare A, Ong T, Pontano L, Cotsarelis G, Zediak VP, Velez M, Bhandoola A, Brown EJ. Deletion of the developmentally essential gene ATR in adult mice leads to age-related phenotypes and stem cell loss. Cell Stem Cell. 2007; 1: 113 -126. [PubMed] .
- 14. Yassa MA and Stark CE. Pattern separation in the hippocampus. Trends Neurosci. 2011; 34: 515 -525. [PubMed] .
- 15. Deng W, Aimone JB, Gage FH. New neurons and new memories: how does adult hippocampal neurogenesis affect learning and memory? Nat Rev Neurosci. 2010; 11: 339 -350. [PubMed] .
- 16. Kempermann G. The neurogenic reserve hypothesis: what is adult hippocampal neurogenesis good for? Trends Neurosci. 2008; 31: 163 -169. [PubMed] .
- 17. Ming GL and Song H. Adult neurogenesis in the mammalian brain: significant answers and significant questions. Neuron. 2011; 70: 687 -702. [PubMed] .
- 18. Marin-Burgin A and Schinder AF. Requirement of adult-born neurons for hippocampus-dependent learning. Behav Brain Res. 2012; 227: 391 -399. [PubMed] .
- 19. Altman J and Das GD. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J Comp Neurol. 1965; 124: 319 -335. [PubMed] .
- 20. Kuhn HG, Dickinson-Anson H, Gage FH. Neurogenesis in the dentate gyrus of the adult rat: age-related decrease of neuronal progenitor proliferation. J Neurosci. 1996; 16: 2027 -2033. [PubMed] .
- 21. Seki T and Arai Y. Age-related production of new granule cells in the adult dentate gyrus. Neuroreport. 1995; 6: 2479 -2482. [PubMed] .
- 22. Gage FH, Kelly PA, Bjorklund A. Regional changes in brain glucose metabolism reflect cognitive impairments in aged rats. J Neurosci. 1984; 4: 2856 -2865. [PubMed] .
- 23. Lazarov O, Mattson MP, Peterson DA, Pimplikar SW, van Praag H. When neurogenesis encounters aging and disease. Trends Neurosci. 2010; 33: 569 -579. [PubMed] .
- 24. Bishop NA, Lu T, Yankner BA. Neural mechanisms of ageing and cognitive decline. Nature. 2010; 464: 529 -535. [PubMed] .
- 25. Gazzaley A, Cooney JW, Rissman J, D'Esposito M. Top-down suppression deficit underlies working memory impairment in normal aging. Nat Neurosci. 2005; 8: 1298 -1300. [PubMed] .
- 26. Prakash RS, Erickson KI, Colcombe SJ, Kim JS, Voss MW, Kramer AF. Age-related differences in the involvement of the prefrontal cortex in attentional control. Brain Cogn. 2009; 71: 328 -335. [PubMed] .
- 27. Wang M, Gamo NJ, Yang Y, Jin LE, Wang XJ, Laubach M, Mazer JA, Lee D, Arnsten AF. Neuronal basis of age-related working memory decline. Nature. 2011; 476: 210 -213. [PubMed] .
- 28. Drapeau E, Mayo W, Aurousseau C, Le Moal M, Piazza PV, Abrous DN. Spatial memory performances of aged rats in the water maze predict levels of hippocampal neurogenesis. Proc Natl Acad Sci U S A. 2003; 100: 14385 -14390. [PubMed] .
- 29. Montaron MF, Drapeau E, Dupret D, Kitchener P, Aurousseau C, Le Moal M, Piazza PV, Abrous DN. Lifelong corticosterone level determines age-related decline in neurogenesis and memory. Neurobiol Aging. 2006; 27: 645 -654. [PubMed] .
- 30. Driscoll I, Howard SR, Stone JC, Monfils MH, Tomanek B, Brooks WM, Sutherland RJ. The aging hippocampus: a multi-level analysis in the rat. Neuroscience. 2006; 139: 1173 -1185. [PubMed] .
- 31. Wimmer ME, Hernandez PJ, Blackwell J, Abel T. Aging impairs hippocampus-dependent long-term memory for object location in mice. Neurobiol Aging. 2011; http://dx.doi.org/10.1016/j.neurobiolaging.2011.07.007 .
- 32. Aizawa K, Ageyama N, Yokoyama C, Hisatsune T. Age-dependent alteration in hippocampal neurogenesis correlates with learning performance of macaque monkeys. Exp Anim. 2009; 58: 403 -407. [PubMed] .
- 33. Bizon JL and Gallagher M. Production of new cells in the rat dentate gyrus over the lifespan: relation to cognitive decline. Eur J Neurosci. 2003; 18: 215 -219. [PubMed] .
- 34. Merrill DA, Karim R, Darraq M, Chiba AA, Tuszynski MH. Hippocampal cell genesis does not correlate with spatial learning ability in aged rats. J Comp Neurol. 2003; 459: 201 -207. [PubMed] .
- 35. Bizon JL, Lee HJ, Gallagher M. Neurogenesis in a rat model of age-related cognitive decline. Aging Cell. 2004; 3: 227 -234. [PubMed] .
- 36. Kempermann G, Kuhn HG, Gage FH. Experience-induced neurogenesis in the senescent dentate gyrus. J Neurosci. 1998; 18: 3206 -3212. [PubMed] .
- 37. Kempermann G, Gast D, Gage FH. Neuroplasticity in old age: sustained fivefold induction of hippocampal neurogenesis by long-term environmental enrichment. Ann Neurol. 2002; 52: 135 -143. [PubMed] .
- 38. van Praag H, Shubert T, Zhao C, Gage FH. Exercise enhances learning and hippocampal neurogenesis in aged mice. J Neurosci. 2005; 25: 8680 -8685. [PubMed] .
- 39. Leuner B, Kozorovitskiy Y, Gross CG, Gould E. Diminished adult neurogenesis in the marmoset brain precedes old age. Proc Natl Acad Sci U S A. 2007; 104: 17169 -17173. [PubMed] .
- 40. Walter J, Keiner S, Witte OW, Redecker C. Age-related effects on hippocampal precursor cell subpopulations and neurogenesis. Neurobiol Aging. 2011; 32: 1906 -1914. [PubMed] .
- 41. Jinno S. Decline in adult neurogenesis during aging follows a topographic pattern in the mouse hippocampus. J Comp Neurol. 2011; 519: 451 -466. [PubMed] .
- 42. Alonso G. Proliferation of progenitor cells in the adult rat brain correlates with the presence of vimentin-expressing astrocytes. Glia. 2001; 34: 253 -266. [PubMed] .
- 43. Hattiangady B and Shetty AK. Aging does not alter the number or phenotype of putative stem/progenitor cells in the neurogenic region of the hippocampus. Neurobiol Aging. 2008; 29: 129 -147. [PubMed] .
- 44. Lugert S, Basak O, Knuckles P, Haussler U, Fabel K, Gotz M, Haas CA, Kempermann G, Taylor V, Giachino C. Quiescent and active hippocampal neural stem cells with distinct morphologies respond selectively to physiological and pathological stimuli and aging. Cell Stem Cell. 2010; 6: 445 -456. [PubMed] .
- 45. Aizawa K, Ageyama N, Terao K, Hisatsune T. Primate-specific alterations in neural stem/progenitor cells in the aged hippocampus. Neurobiol Aging. 2011; 32: 140 -150. [PubMed] .
- 46. Encinas JM, Michurina TV, Peunova N, Park JH, Tordo J, Peterson DA, Fishell G, Koulakov A, Enikolopov G. Division-coupled astrocytic differentiation and age-related depletion of neural stem cells in the adult hippocampus. Cell Stem Cell. 2011; 8: 566 -579. [PubMed] .
- 47. Bonaguidi MA, Wheeler MA, Shapiro JS, Stadel RP, Sun GJ, Ming GL, Song H. In vivo clonal analysis reveals self-renewing and multipotent adult neural stem cell characteristics. Cell. 2011; 145: 1142 -1155. [PubMed] .
- 48. Blagosklonny MV. Cell cycle arrest is not senescence. Aging. 2011; 3: 94 -101. [PubMed] .
- 49. Kronenberg G, Bick-Sander A, Bunk E, Wolf C, Ehninger D, Kempermann G. Physical exercise prevents age-related decline in precursor cell activity in the mouse dentate gyrus. Neurobiol Aging. 2006; 27: 1505 -1513. [PubMed] .
- 50. Erickson KI, Voss MW, Prakash RS, Basak C, Szabo A, Chaddock L, Kim JS, Heo S, Alves H, White SM, Wojcicki TR, Mailey E, Vieira VJ, et al. Exercise training increases size of hippocampus and improves memory. Proc Natl Acad Sci U S A. 2011; 108: 3017 -3022. [PubMed] .
- 51. Kim YP, Kim H, Shin MS, Chang HK, Jang MH, Shin MC, Lee SJ, Lee HH, Yoon JH, Jeong IG, Kim CJ. Age-dependence of the effect of treadmill exercise on cell proliferation in the dentate gyrus of rats. Neurosci Lett. 2004; 355: 152 -154. [PubMed] .
- 52. Rao MS, Hattiangady B, Shetty AK. Status epilepticus during old age is not associated with enhanced hippocampal neurogenesis. Hippocampus. 2008; 18: 931 -944. [PubMed] .
- 53. Darsalia V, Heldmann U, Lindvall O, Kokaia Z. Stroke-induced neurogenesis in aged brain. Stroke. 2005; 36: 1790 -1795. [PubMed] .
- 54. Yagita Y, Kitagawa K, Ohtsuki T, Takasawa K, Miyata T, Okano H, Hori M, Matsumoto M. Neurogenesis by progenitor cells in the ischemic adult rat hippocampus. Stroke. 2001; 32: 1890 -1896. [PubMed] .
- 55. Walter J, Keiner S, Witte OW, Redecker C. Differential stroke-induced proliferative response of distinct precursor cell subpopulations in the young and aged dentate gyrus. Neuroscience. 2010; 169: 1279 -1286. [PubMed] .
- 56. Rao MS, Hattiangady B, Abdel-Rahman A, Stanley DP, Shetty AK. Newly born cells in the ageing dentate gyrus display normal migration, survival and neuronal fate choice but endure retarded early maturation. Eur J Neurosci. 2005; 21: 464 -476. [PubMed] .
- 57. Rao MS, Hattiangady B, Shetty AK. The window and mechanisms of major age-related decline in the production of new neurons within the dentate gyrus of the hippocampus. Aging cell. 2006; 5: 545 -558. [PubMed] .
- 58. McDonald HY and Wojtowicz JM. Dynamics of neurogenesis in the dentate gyrus of adult rats. Neurosci Lett. 2005; 385: 70 -75. [PubMed] .
- 59. Palmer TD, Willhoite AR, Gage FH. Vascular niche for adult hippocampal neurogenesis. J Comp Neurol. 2000; 425: 479 -494. [PubMed] .
- 60. Tavazoie M, Van der Veken L, Silva-Vargas V, Louissaint M, Colonna L, Zaidi B, Garcia-Verdugo JM, Doetsch F. A specialized vascular niche for adult neural stem cells. Cell Stem Cell. 2008; 3: 279 -288. [PubMed] .
- 61. Mirzadeh Z, Merkle FT, Soriano-Navarro M, Garcia-Verdugo JM, Alvarez-Buylla A. Neural stem cells confer unique pinwheel architecture to the ventricular surface in neurogenic regions of the adult brain. Cell Stem Cell. 2008; 3: 265 -278. [PubMed] .
- 62. Shen Q, Wang Y, Kokovay E, Lin G, Chuang SM, Goderie SK, Roysam B, Temple S. Adult SVZ stem cells lie in a vascular niche: a quantitative analysis of niche cell-cell interactions. Cell Stem Cell. 2008; 3: 289 -300. [PubMed] .
- 63. Riddle DR, Sonntag WE, Lichtenwalner RJ. Microvascular plasticity in aging. Ageing Res Rev. 2003; 2: 149 -168. [PubMed] .
- 64. Leenders KL, Perani D, Lammertsma AA, Heather JD, Buckingham P, Healy MJ, Gibbs JM, Wise RJ, Hatazawa J, Herold S, et al. Cerebral blood flow, blood volume and oxygen utilization. Normal values and effect of age. Brain. 1990; 113: 27 -47. [PubMed] .
- 65. Ajmani RS, Metter EJ, Jaykumar R, Ingram DK, Spangler EL, Abugo OO, Rifkind JM. Hemodynamic changes during aging associated with cerebral blood flow and impaired cognitive function. Neurobiol Aging. 2000; 21: 257 -269. [PubMed] .
- 66. Bentourkia M, Bol A, Ivanoiu A, Labar D, Sibomana M, Coppens A, Michel C, Cosnard G, De Volder AG. Comparison of regional cerebral blood flow and glucose metabolism in the normal brain: effect of aging. J Neurol Sci. 2000; 181: 19 -28. [PubMed] .
- 67. Heo S, Prakash RS, Voss MW, Erickson KI, Ouyang C, Sutton BP, Kramer AF. Resting hippocampal blood flow, spatial memory and aging. Brain Res. 2010; 1315: 119 -127. [PubMed] .
- 68. Studer L, Csete M, Lee SH, Kabbani N, Walikonis J, Wold B, McKay R. Enhanced proliferation, survival, and dopaminergic differentiation of CNS precursors in lowered oxygen. J Neurosci. 2000; 20: 7377 -7383. [PubMed] .
- 69. Morrison SJ, Csete M, Groves AK, Melega W, Wold B, Anderson DJ. Culture in reduced levels of oxygen promotes clonogenic sympathoadrenal differentiation by isolated neural crest stem cells. J Neurosci. 2000; 20: 7370 -7376. [PubMed] .
- 70. Storch A, Lester HA, Boehm BO, Schwarz J. Functional characterization of dopaminergic neurons derived from rodent mesencephalic progenitor cells. J Chem Neuroanat. 2003; 26: 133 -142. [PubMed] .
- 71. Storch A, Paul G, Csete M, Boehm BO, Carvey PM, Kupsch A, Schwarz J. Long-term proliferation and dopaminergic differentiation of human mesencephalic neural precursor cells. Exp Neurol. 2001; 170: 317 -325. [PubMed] .
- 72. Pistollato F, Chen HL, Rood BR, Zhang HZ, D'Avella D, Denaro L, Gardiman M, te Kronnie G, Schwartz PH, Favaro E, Indraccolo S, Basso G, Panchision DM. Hypoxia and HIF1alpha repress the differentiative effects of BMPs in high-grade glioma. Stem Cells. 2009; 27: 7 -17. [PubMed] .
- 73. Zheng X, Linke S, Dias JM, Zheng X, Gradin K, Wallis TP, Hamilton BR, Gustafsson M, Ruas JL, Wilkins S, Bilton RL, Brismar K, Whitelaw ML, et al. Interaction with factor inhibiting HIF-1 defines an additional mode of cross-coupling between the Notch and hypoxia signaling pathways. Proc Natl Acad Sci U S A. 2008; 105: 3368 -3373. [PubMed] .
- 74. Zhang K, Zhu L, Fan M. Oxygen, a Key Factor Regulating Cell Behavior during Neurogenesis and Cerebral Diseases. Front Mol Neurosci. 2011; 4: 5 [PubMed] .
- 75. Mazumdar J, O'Brien WT, Johnson RS, LaManna JC, Chavez JC, Klein PS, Simon MC. O2 regulates stem cells through Wnt/beta-catenin signalling. Nat Cell Biol. 2010; 12: 1007 -1013. [PubMed] .
- 76. Rabie T, Kunze R, Marti HH. Impaired hypoxic response in senescent mouse brain. Int J Dev Neurosci. 2011; 29: 655 -661. [PubMed] .
- 77. Benderro GF and Lamanna JC. Hypoxia-induced angiogenesis is delayed in aging mouse brain. Brain Res. 2011; 1389: 50 -60. [PubMed] .
- 78. Villeda SA, Luo J, Mosher KI, Zou B, Britschgi M, Bieri G, Stan TM, Fainberg N, Ding Z, Eggel A, Lucin KM, Czirr E, Park JS, et al. The ageing systemic milieu negatively regulates neurogenesis and cognitive function. Nature. 2011; 477: 90 -94. [PubMed] .
- 79. Herbert J, Goodyer IM, Grossman AB, Hastings MH, de Kloet ER, Lightman SL, Lupien SJ, Roozendaal B, Seckl JR. Do corticosteroids damage the brain? J Neuroendocrinol. 2006; 18: 393 -411. [PubMed] .
- 80. Reul JM and de Kloet ER. Two receptor systems for corticosterone in rat brain: microdistribution and differential occupation. Endocrinology. 1985; 117: 2505 -2511. [PubMed] .
- 81. Han F, Ozawa H, Matsuda K, Nishi M, Kawata M. Colocalization of mineralocorticoid receptor and glucocorticoid receptor in the hippocampus and hypothalamus. Neurosci Res. 2005; 51: 371 -381. [PubMed] .
- 82. Garcia A, Steiner B, Kronenberg G, Bick-Sander A, Kempermann G. Age-dependent expression of glucocorticoid- and mineralocorticoid receptors on neural precursor cell populations in the adult murine hippocampus. Aging Cell. 2004; 3: 363 -371. [PubMed] .
- 83. Gould E, Cameron HA, Daniels DC, Woolley CS, McEwen BS. Adrenal hormones suppress cell division in the adult rat dentate gyrus. J Neurosci. 1992; 12: 3642 -3650. [PubMed] .
- 84. Cameron HA and Gould E. Adult neurogenesis is regulated by adrenal steroids in the dentate gyrus. Neuroscience. 1994; 61: 203 -209. [PubMed] .
- 85. Cameron HA and McKay RD. Restoring production of hippocampal neurons in old age. Nat Neurosci. 1999; 2: 894 -897. [PubMed] .
- 86. Montaron MF, Petry KG, Rodriguez JJ, Marinelli M, Aurousseau C, Rougon G, Le Moal M, Abrous DN. Adrenalectomy increases neurogenesis but not PSA-NCAM expression in aged dentate gyrus. Eur J Neurosci. 1999; 11: 1479 -1485. [PubMed] .
- 87. Garrido P, de Blas M, Del Arco A, Segovia G, Mora F. Aging increases basal but not stress-induced levels of corticosterone in the brain of the awake rat. Neurobiol Aging. 2012; 33: 375 -382. [PubMed] .
- 88. Lai M, Hibberd CJ, Gluckman PD, Seckl JR. Reduced expression of insulin-like growth factor 1 messenger RNA in the hippocampus of aged rats. Neurosci Lett. 2000; 288: 66 -70. [PubMed] .
- 89. Shetty AK, Hattiangady B, Shetty GA. Stem/progenitor cell proliferation factors FGF-2, IGF-1, and VEGF exhibit early decline during the course of aging in the hippocampus: role of astrocytes. Glia. 2005; 51: 173 -186. [PubMed] .
- 90. Bernal GM and Peterson DA. Phenotypic and gene expression modification with normal brain aging in GFAP-positive astrocytes and neural stem cells. Aging Cell. 2011; 10: 466 -482. [PubMed] .
- 91. Chadashvili T and Peterson DA. Cytoarchitecture of fibroblast growth factor receptor 2 (FGFR-2) immunoreactivity in astrocytes of neurogenic and non-neurogenic regions of the young adult and aged rat brain. J Comp Neurol. 2006; 498: 1 -15. [PubMed] .
- 92. Sun LY, Al-Regaiey K, Masternak MM, Wang J, Bartke A. Local expression of GH and IGF-1 in the hippocampus of GH-deficient long-lived mice. Neurobiol Aging. 2005; 26: 929 -937. [PubMed] .
- 93. Trejo JL, Llorens-Martin MV, Torres-Aleman I. The effects of exercise on spatial learning and anxiety-like behavior are mediated by an IGF-I-dependent mechanism related to hippocampal neurogenesis. Mol Cell Neurosci. 2008; 37: 402 -411. [PubMed] .
- 94. Aberg MA, Aberg ND, Hedbacker H, Oscarsson J, Eriksson PS. Peripheral infusion of IGF-I selectively induces neurogenesis in the adult rat hippocampus. J Neurosci. 2000; 20: 2896 -2903. [PubMed] .
- 95. Lichtenwalner RJ, Forbes ME, Bennett SA, Lynch CD, Sonntag WE, Riddle DR. Intracerebroventricular infusion of insulin-like growth factor-I ameliorates the age-related decline in hippocampal neurogenesis. Neuroscience. 2001; 107: 603 -613. [PubMed] .
- 96. Zhao M, Li D, Shimazu K, Zhou YX, Lu B, Deng CX. Fibroblast growth factor receptor-1 is required for long-term potentiation, memory consolidation, and neurogenesis. Biol Psychiatry. 2007; 62: 381 -390. [PubMed] .
- 97. Kuhn HG, Winkler J, Kempermann G, Thal LJ, Gage FH. Epidermal growth factor and fibroblast growth factor-2 have different effects on neural progenitors in the adult rat brain. J Neurosci. 1997; 17: 5820 -5829. [PubMed] .
- 98. Wagner JP, Black IB, DiCicco-Bloom E. Stimulation of neonatal and adult brain neurogenesis by subcutaneous injection of basic fibroblast growth factor. J Neurosci. 1999; 19: 6006 -6016. [PubMed] .
- 99. Jin K, Sun Y, Xie L, Batteur S, Mao XO, Smelick C, Logvinova A, Greenberg DA. Neurogenesis and aging: FGF-2 and HB-EGF restore neurogenesis in hippocampus and subventricular zone of aged mice. Aging Cell. 2003; 2: 175 -183. [PubMed] .
- 100. Rai KS, Hattiangady B, Shetty AK. Enhanced production and dendritic growth of new dentate granule cells in the middle-aged hippocampus following intracerebroventricular FGF-2 infusions. European J Neurosci. 2007; 26: 1765 -1779. [PubMed] .
- 101. Schanzer A, Wachs FP, Wilhelm D, Acker T, Cooper-Kuhn C, Beck H, Winkler J, Aigner L, Plate KH, Kuhn HG. Direct stimulation of adult neural stem cells in vitro and neurogenesis in vivo by vascular endothelial growth factor. Brain Pathol. 2004; 14: 237 -248. [PubMed] .
- 102. Sun Y, Jin K, Childs JT, Xie L, Mao XO, Greenberg DA. Vascular endothelial growth factor-B (VEGFB) stimulates neurogenesis: evidence from knockout mice and growth factor administration. Dev Biol. 2006; 289: 329 -335. [PubMed] .
- 103. Jin K, Zhu Y, Sun Y, Mao XO, Xie L, Greenberg DA. Vascular endothelial growth factor (VEGF) stimulates neurogenesis in vitro and in vivo. Proc Natl Acad Sci U S A. 2002; 99: 11946 -11950. [PubMed] .
- 104. Cao L, Jiao X, Zuzga DS, Liu Y, Fong DM, Young D, During MJ. VEGF links hippocampal activity with neurogenesis, learning and memory. Nat Genet. 2004; 36: 8 827 -835. [PubMed] .
- 105. Sonntag WE, Ramsey M, Carter CS. Growth hormone and insulin-like growth factor-1 (IGF-1) and their influence on cognitive aging. Ageing Res Rev. 2005; 4: 195 -212. [PubMed] .
- 106. Mudo G, Bonomo A, Di Liberto V, Frinchi M, Fuxe K, Belluardo N. The FGF-2/FGFRs neurotrophic system promotes neurogenesis in the adult brain. J Neural Transm. 2009; 116: 995 -1005. [PubMed] .
- 107. van Praag H, Kempermann G, Gage FH. Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nat Neurosci. 1999; 2: 266 -270. [PubMed] .
- 108. van Praag H. Exercise and the brain: something to chew on. Trends Neurosci. 2009; 32: 283 -290. [PubMed] .
- 109. Trejo JL, Carro E, Torres-Aleman I. Circulating insulin-like growth factor I mediates exercise-induced increases in the number of new neurons in the adult hippocampus. J Neurosci. 2001; 21: 1628 -1634. [PubMed] .
- 110. Glasper ER, Llorens-Martin MV, Leuner B, Gould E, Trejo JL. Blockade of insulin-like growth factor-I has complex effects on structural plasticity in the hippocampus. Hippocampus. 2010; 20: 706 -712. [PubMed] .
- 111. Fabel K, Tam B, Kaufer D, Baiker A, Simmons N, Kuo CJ, Palmer TD. VEGF is necessary for exercise-induced adult hippocampal neurogenesis. Eur J Neurosci. 2003; 18: 2803 -2812. [PubMed] .
- 112. Bishop NA and Guarente L. Genetic links between diet and lifespan: shared mechanisms from yeast to humans. Nat Rev Genet. 2007; 8: 835 -844. [PubMed] .
- 113. Blagosklonny MV. Calorie restriction: decelerating mTOR-driven aging from cells to organisms (including humans). Cell Cycle. 2010; 9: 683 -688. [PubMed] .
- 114. Blagosklonny MV. Aging, stem cells, and mammalian target of rapamycin: a prospect of pharmacologic rejuvenation of aging stem cells. Rejuvenation Res. 2008; 11: 801 -808. [PubMed] .
- 115. Castilho RM, Squarize CH, Chodosh LA, Williams BO, Gutkind JS. mTOR mediates Wnt-induced epidermal stem cell exhaustion and aging. Cell Stem Cell. 2009; 5: 279 -289. [PubMed] .
- 116. Lee J, Seroogy KB, Mattson MP. Dietary restriction enhances neurotrophin expression and neurogenesis in the hippocampus of adult mice. J Neurochem. 2002; 80: 539 -547. [PubMed] .
- 117. Bondolfi L, Ermini F, Long JM, Ingram DK, Jucker M. Impact of age and caloric restriction on neurogenesis in the dentate gyrus of C57BL/6 mice. Neurobiol Aging. 2004; 25: 333 -340. [PubMed] .
- 118. Levenson CW and Rich NJ. Eat less, live longer? New insights into the role of caloric restriction in the brain. Nutrition Rev. 2007; 65: 412 -415. [PubMed] .
- 119. Kim JY, Duan X, Liu CY, Jang MH, Guo JU, Pow-anpongkul N, Kang E, Song H, Ming GL. DISC1 regulates new neuron development in the adult brain via modulation of AKT-mTOR signaling through KIAA1212. Neuron. 2009; 63: 761 -773. [PubMed] .
- 120. Prolla TA and Mattson MP. Molecular mechanisms of brain aging and neurodegenerative disorders: lessons from dietary restriction. Trends Neurosci. 2001; 24: 11 Suppl S21 -31. [PubMed] .
- 121. Fabel K and Kempermann G. Physical activity and the regulation of neurogenesis in the adult and aging brain. Neuromolecular Med. 2008; 10: 59 -66. [PubMed] .
- 122. Cotman CW, Berchtold NC, Christie LA. Exercise builds brain health: key roles of growth factor cascades and inflammation. Trends Neurosci. 2007; 30: 464 -472. [PubMed] .
- 123. Suh H, Deng W, Gage FH. Signaling in adult neurogenesis. Annu Rev Cell Dev Biol. 2009; 25: 253 -275. [PubMed] .
- 124. Doetsch F. A niche for adult neural stem cells. Curr Opin Genet Dev. 2003; 13: 543 -550. [PubMed] .
- 125. Ninkovic J and Gotz M. Signaling in adult neurogenesis: from stem cell niche to neuronal networks. Curr Opin Neurobiol. 2007; 17: 338 -344. [PubMed] .
- 126. Ohnuma S and Harris WA. Neurogenesis and the cell cycle. Neuron. 2003; 40: 199 -208. [PubMed] .
- 127. Dehay C and Kennedy H. Cell-cycle control and cortical development. Nat Rev Neurosci. 2007; 8: 438 -450. [PubMed] .
- 128. Salomoni P and Calegari F. Cell cycle control of mammalian neural stem cells: putting a speed limit on G1. Trends Cell Biol. 2010; 5: 332 -342. .
- 129. Beukelaers P, Vandenbosch R, Caron N, Nguyen L, Moonen G, Malgrange B. Cycling or not cycling: cell cycle regulatory molecules and adult neurogenesis. Cell Mol Life Sci. 2011; DOI 10.1007/s00018-011-0880-6. .
- 130. Krishnamurthy J, Torrice C, Ramsey MR, Kovalev GI, Al-Regaiey K, Su L, Sharpless NE. Ink4a/Arf expression is a biomarker of aging. J Clin Invest. 2004; 114: 1299 -1307. [PubMed] .
- 131. Molofsky AV, Slutsky SG, Joseph NM, He S, Pardal R, Krishnamurthy J, Sharpless NE, Morrison SJ. Increasing p16INK4a expression decreases forebrain progenitors and neurogenesis during ageing. Nature. 2006; 443: 448 -452. [PubMed] .
- 132. Molofsky AV, He S, Bydon M, Morrison SJ, Pardal R. Bmi-1 promotes neural stem cell self-renewal and neural development but not mouse growth and survival by repressing the p16Ink4a and p19Arf senescence pathways. Genes Dev. 2005; 19: 1432 -1437. [PubMed] .
- 133. Kohman RA, Rodriguez-Zas SL, Southey BR, Kelley KW, Dantzer R, Rhodes JS. Voluntary wheel running reverses age-induced changes in hippocampal gene expression. PloS One. 2011; 6: e22654 [PubMed] .
- 134. Olariu A, Cleaver KM, Cameron HA. Decreased neurogenesis in aged rats results from loss of granule cell precursors without lengthening of the cell cycle. J Comp Neurol. 2007; 501: 659 -667. [PubMed] .
- 135. Stoll EA, Habibi BA, Mikheev AM, Lasiene J, Massey SC, Swanson KR, Rostomily RC, Horner PJ. Increased re-entry into cell cycle mitigates age-related neurogenic decline in the murine subventricular zone. Stem Cells. 2011; 29: 2005 -2017. [PubMed] .
- 136. Lange C, Huttner WB, Calegari F. Cdk4/cyclinD1 overexpression in neural stem cells shortens G1, delays neurogenesis, and promotes the generation and expansion of basal progenitors. Cell Stem Cell. 2009; 5: 320 -331. [PubMed] .
- 137. Artegiani B, Lindemann D, Calegari F. Overexpression of cdk4 and cyclinD1 triggers greater expansion of neural stem cells in the adult mouse brain. J Exp Med. 2011; 208: 937 -948. [PubMed] .
- 138. Beukelaers P, Vandenbosch R, Caron N, Nguyen L, Belachew S, Moonen G, Kiyokawa H, Barbacid M, Santamaria D, Malgrange B. Cdk6-dependent regulation of G(1) length controls adult neurogenesis. Stem Cells. 2011; 29: 713 -724. [PubMed] .
- 139. Kowalczyk A, Filipkowski RK, Rylski M, Wilczynski GM, Konopacki FA, Jaworski J, Ciemerych MA, Sicinski P, Kaczmarek L. The critical role of cyclin D2 in adult neurogenesis. J Cell Biol. 2004; 167: 209 -213. [PubMed] .
- 140. Calegari F and Huttner WB. An inhibition of cyclin-dependent kinases that lengthens, but does not arrest, neuroepithelial cell cycle induces premature neurogenesis. J Cell Sci. 2003; 116: 4947 -4955. [PubMed] .
- 141. Lange C and Calegari F. Cdks and cyclins link G(1) length and differentiation of embryonic, neural and hematopoietic stem cells. Cell Cycle. 2010; 9: 1893 -1900. [PubMed] .
- 142. Kempermann G, Gast D, Kronenberg G, Yamaguchi M, Gage FH. Early determination and long-term persistence of adult-generated new neurons in the hippocampus of mice. Development. 2003; 130: 391 -399. [PubMed] .
- 143. Morgenstern NA, Lombardi G, Schinder AF. Newborn granule cells in the ageing dentate gyrus. J Physiol. 2008; 586: 3751 -3757. [PubMed] .
- 144. Kempermann G, Kuhn HG, Gage FH. More hippocampal neurons in adult mice living in an enriched environment. Nature. 1997; 386: 493 -495. [PubMed] .
- 145. Sahay A, Scobie KN, Hill AS, O'Carroll CM, Kheirbek MA, Burghardt NS, Fenton AA, Dranovsky A, Hen R. Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature. 2011; 472: 466 -470. [PubMed] .
- 146. Bernal GM and Peterson DA. Neural stem cells as therapeutic agents for age-related brain repair. Aging cell. 2004; 3: 345 -351. [PubMed] .
- 147. Lindvall O and Kokaia Z. Stem cells for the treatment of neurological disorders. Nature. 2006; 441: 1094 -1096. [PubMed] .
- 148. Lie DC, Song H, Colamarino SA, Ming GL, Gage FH. Neurogenesis in the adult brain: new strategies for central nervous system diseases. Annu Rev Pharmacol Toxicol. 2004; 44: 399 -421. [PubMed] .
- 149. Minger SL, Ekonomou A, Carta EM, Chinoy A, Perry RH, Ballard CG. Endogenous neurogenesis in the human brain following cerebral infarction. Regen Med. 2007; 2: 69 -74. [PubMed] .
- 150. Jin K, Wang X, Xie L, Mao XO, Zhu W, Wang Y, Shen J, Mao Y, Banwait S, Greenberg DA. Evidence for stroke-induced neurogenesis in the human brain. Proc Natl Acad Sci U S A. 2006; 103: 13198 -13202. [PubMed] .
- 151. Macas J, Nern C, Plate KH, Momma S. Increased generation of neuronal progenitors after ischemic injury in the aged adult human forebrain. J Neurosci. 2006; 26: 13114 -13119. [PubMed] .
- 152. Dunnett SB and Rosser AE. Clinical translation of cell transplantation in the brain. Curr Opin Organ Transplant. 2011; 16: 632 -639. [PubMed] .