



Introduction
Ischemic stroke results from the occlusion of an afferent blood vessel and the subsequent reduction of the blood and oxygen supply to the affected brain regions. It accounts for roughly 80% of all clinical strokes [1]. Overall, oxidative stress and cell death (apoptosis and necrosis) are the key molecular processes that are involved in ischemic stroke.
Interestingly, exposure to genotoxic stressors such as toxic chemicals or radiation also causes oxidative stress and apoptosis [2-3]. Indeed, it has been long-established that naïve cells that have been in direct contact with genotoxic stress-exposed cells or have received a certain distress signal from exposed cells exhibit signs of genome destabilization [4]. Such an exposure communication process is termed a bystander effect. This effect is well-documented in cells in vitro upon radiation exposure. Recent studies have also shown that chemotherapy can lead to bystander effects in the neighboring cells [5].The bystander effect also manifests itself in vivo in the tissue and whole-organism context. It was recently shown that cranial irradiation leads to a wide variety of cellular effects in the distal unexposed spleens, testes, and livers of laboratory animals [6-7].
Does ischemic stroke cause a similar bystander-like effect? This question remains to be answered. We hypothesized that ischemic stroke will affect the entire organism and will result in bystander-like effects in the livers, kidneys, and hearts of the animals. We also predicted that, similarly to the irradiation bystander phenomenon [6, 8], the stroke-induced distal bystander-like effects might be epigenetic in nature.
Epigenetic changes are meiotically and mitotically stable alterations in gene expression that are not based on DNA sequence changes and that involve processes that affectchromatin structure, such as DNA methylation, histone modifications, and small RNA-induced silencing [9]. Cytosine DNA methylation is the most extensively studied epigenetic process[10]. It is known to be associated with an inactive chromatin state and repressedgene expression [11-12]. In mammals, three DNA methyltransferases (DNMT1, DNMT3a, and DNMT3b) partake in establishing (DNMT3a and DNMT3b) and maintaining (DNMT1) DNA methylation patterns [13-14]. Proper maintenance of DNA methylation is important for maintaining genome stability, and altered DNA methylation has been well documented in cancer, immunological, cardiovascular, developmental, neurological and psychiatric disorder and aging [15-17].
Changes in DNA methylation do not appear to be isolated, independent events; they often accompany chemical modifications to histone proteins[12, 18-19]. Numerous histone modifications include (not exclusively) acetylation, methylation, and phosphorylation. Histone acetylation leads to more relaxed chromatin packaging and increased gene expression. Histone deacetylation has an opposite effect. Histone methylation is not as straightforward and may lead to both chromatin compaction and relaxation depending upon the residue that is modified [20-21].
Furthermore, classical, genotoxic stress-induced bystander effects are mediated in part through small RNAs — specifically, microRNAs [22]. MicroRNAs are small, single-stranded non-coding RNAs that regulate gene expression at the post-transcriptional level [23]. To control translation of target mRNAs, microRNAs associate with RNA-induced silencing complex (RISC) proteins and bind to the 3' UTR of mRNAs, thus serving as translational suppressors and thereby regulating the production of proteins and affecting many cellular functions, including proliferation, differentiation, cell death, senescence and others [24, 25].
Here, we analyzed epigenetic changes in the liver, kidney, and heart tissue of control and stroked animals. These organs were chosen based on some of the scarce, relevant clinical reports suggesting the indirect impact of stroke on their function in affected patients.
Results and Discussion
Lack of DNA methylation changes in liver, kidney, and heart tissue of stroked animals
The focal, permanent devascularization (stroke) has been described in detail elsewhere [26]. Figure 1 provides a schematic description of the stroke model used in our study. To test if changes in global DNA methylation wereobserved in liver, kidney and heart tissues of control and stroked animals, we employed the well-established HpaII-based cytosine extension assay. This assay measures the proportion of unmethylated CCGG sites in genomic DNA [8]. We found that all tissues of the control and stroked animals had comparable DNA methylation levels (Figure 2).
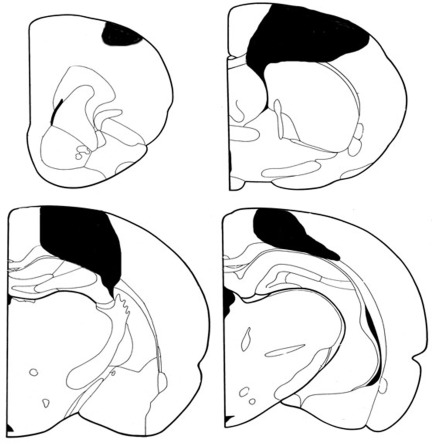
Figure 1. Coronal sections representing the extent of a typical stroke Though this was a somewhat surprising finding, we note that the cytosine extension assay measures global net changes in DNA methylation and does not provide us with the locus-specific DNA methylation details. Thus, we cannot exclude the possibility that there still may be loci that have lost or gained DNA methylation in the tissues of stroked animals. Indeed, recently it has been shown that alterations in DNA methylation in cancer cells occur in defined regions, suggesting localized, and not random, global deregulation of DNA methylation [27-28]. A detailed analysis of locus-specific DNA methylation changes may be needed to investigate the precise gene and locus-specific nature of DNA methylation changes.
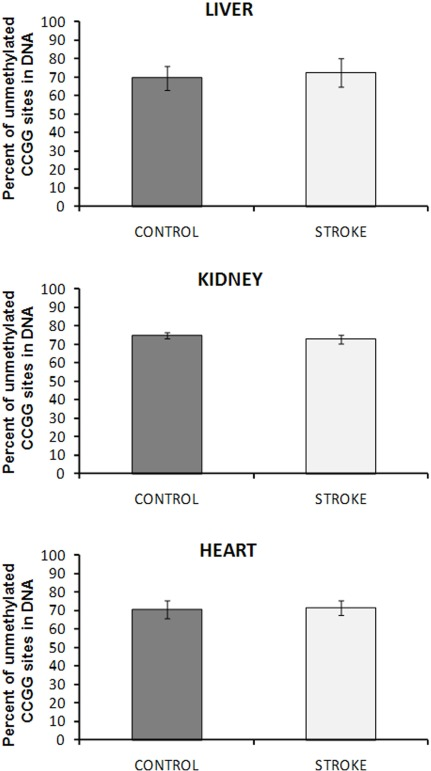
Figure 2. DNA methylation in liver, heart and kidney tissues of control and stroked rats The absolute percent of double-stranded unmethylated CCGG sites was calculated by relating the data of HpaII and MspI digests. Data are presented as mean values ± SD.
Subtle changes in the levels of DNA methyltransferases and methyl-binding protein MeCP2 in the distal somatic tissues of stroked animals
Analysis of the levels of DNMT1 and DNMT3a the liver, kidney, and heart tissues of control and stroked animals revealed interesting and tissue-specific protein expression changes. We noted that DNMT1 levels were subtly (p<0.10) decreased in the heart tissue of stroked animals as compared to controls. In the kidneys of stroked animals, contrarily, the levels of DNMT3a were slightly increased (p<0.05). In the liver tissue of stroked animals, both DNMT1 and DNMT3a levels remained unchanged (Figure 3).
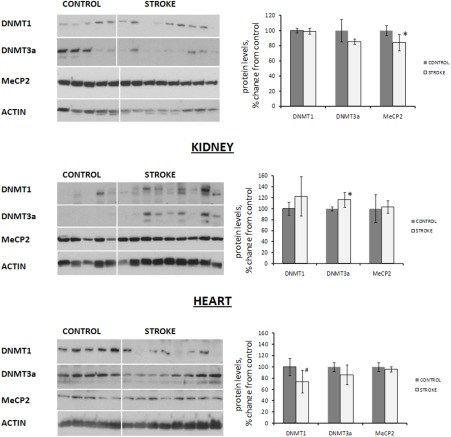
Figure 3. Expression of DNA methyltransferases and methyl-binding protein MeCP2 in liver, heart and kidney tissues of control and stroked rats Protein levels relative to those of control animals are shown as the mean ± SD, ±significant, p < 0.05, Student's t-test. Representative blots from among three independent repeats are shown.
In addition to altered levels of DNMTs, we noted a subtle change in the levels of methyl-binding protein MeCP2 in the liver tissue of stroked rats (Figure 3).
This protein was shown to interact with methylated DNA and affect the methylation-mediated chromatin remodeling and gene silencing [29]. The contributions of DNMTs and MeCP2 to bystander-like effects induced by stroke in distal somatic organs have yet to be analyzed, as those may contribute to establishing some locus-specific DNA methylation patterns.
Histone methylation and acetylation changes in the distal tissues of stroked animals
Changes in histone modifications directly affect chromatin packaging and gene expression [30]. Among numerous histone modifications, trimethylation of histone H3 lysine 9 (H3K9) is associated with chromatin compaction and decreased expression [31]. Acetylation of H3K3, contrarily, leads to increased expression and chromatin relaxation [30]. These modifications may be mutually exclusive. Furthermore, trimethylation of lysine 4 on histone H3 (H3K4) is also known to correlate with increased expression and relaxed chromatin.
We analyzed the levels of these H3K9 and H3K4 modifications using western immunoblotting. While no changes were seen in the liver, and only one modification occurred in the heart, the changes in the kidney tissues of stroked animals were the most pronounced.
Indeed, on the one hand, no histone acetylation or methylation changes were seen in the liver tissue of stroked rats, and in the heart tissue of stroked animals, we noted a significant decrease in the level of trimethylated H3K4. On the other hand, we noted a statistically significant 15% increase in the levels of acetylated H3K9, and a corresponding decrease in methylated H3K9, in the kidney tissue of stroked animals as compared to control animals (Figure 4). Likewise, we noted a significant 16% increase in the levels of H3K4 trimethylation in the kidney tissue of stroked rats compared to the controls (Figure 4).
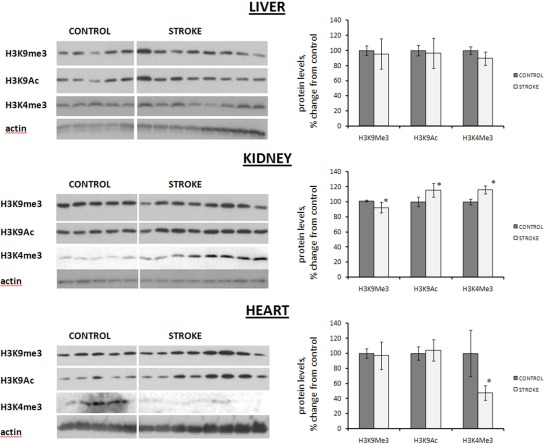
Figure 4. Histone modification levels in liver, heart and kidney tissues of control and stroked rats Levels relative to those of control animals are shown as the mean ± SD. Representative blots from among three independent repeats are shown. ±significant, p < 0.05, Student's t-test.
Interestingly, H3K4 and H3K9 methylation levels are mutually exclusive, and this relationship is conserved from yeast to humans. Recent findings show that increased H3K4 trimethylation plays an important part in the renal inflammatory response and is associated with proximal tubal injury, acute renal failure, and ischemic renal injury. On the other hand, decreased levels of H3K4 trimethylation were reported to be a negative prognostic indicator for renal cell carcinoma. Moreover, recent studies show that the acetylation of histone H3K9 and loss of H3K9 trimethylation may be important in progressive glomerulosclerosis [32-33]. Stroke-induced effects in the kidney may in turn be relatively similar to those induced by hemorrhage and glomerulosclerosis.
In sum, the observed histone changes may be indicative of the alteration in chromatin structure and gene expression in the kidneys of stroked rats. These new intriguing findings deserve special attention in the future. It is especially important to discern the locus specificity of the observed global histone changes and any relationship between the observed changes and inflammation.
Altered gene expression in the distal kidney tissues of stroked animals
Decreased levels of methylated H3K9, increased levels of acetylated H3K9, and methylated H3K4 may in turn lead to the activation of gene expression. Therefore we conducted an analysis of differential gene expression inthe kidney tissue of control and stroked rats. Our analysis revealed that 22 genes were differentially expressed in the kidney tissues of stroked rats as compared to the controls. Interestingly, all of these genes were up-regulated. The expression of four randomly selected genes—Cml3, Rps27a, RGD1562073, and Tnfsf13—was verified by qRT-PCR (Table 1).
| Gene ID | Gene name | Cellular function/process | Fold induction (array) | Fold induction (qRT-PCR) | Condition/disease association |
|---|---|---|---|---|---|
| Alad | δ-aminolevulinic acid dehydrase | heme biosynthesis | 1.3 | kidney response to toxic compounds | |
| Cln6_predicted | ceroid-lipofuscinosis, neuronal 6 | 1.3 | NCL | ||
| Cml3 | camello-like 3, transcription factor | transcription | 1.6 | 1.9 | |
| Degs1 | degenerative spermatocyte homolog 1, lipid desaturase | membrane fatty acid desaturase | 1.4 | cancer, metastasis | |
| Exosc7 | exosome component 7 | exosome function | 1.3 | ||
| Krt2-7 | a member of the type I (acidic) keratin family | cytoskeleton | 1.5 | nephropathy | |
| LOC289715 | Rattus norvegicus similar to ribosomal protein L37 | translation | 1.3 | cancer | |
| Mrps36_predicted | mammalian mitochondrial ribosomal protein | translation | 1.3 | ||
| Plac8_predicted | Rattus norvegicus placenta-specific 8 (predicted) | 1.6 | |||
| RGD1310925_predicted | small nuclear ribonucleoprotein 27 (U4/U6.U5) | splicing | 1.3 | ||
| RGD1559951_predicted | similar to 60S ribosomal protein L37a | translation | 1.4 | cancer, DNA damage response | |
| RGD1559960_predicted | similar to sulfotransferase K1 | 1.9 | |||
| RGD1561195_predicted | similar to ribosomal protein L31 | translation | 1.3 | rat cerebral ischemia | |
| RGD1561310_predicted | similar to ribosomal protein L37 | translation | 1.4 | cancer | |
| RGD1562073_predicted | similar to ribosomal protein S17 | translation | 1.9 | 1.6 | |
| RGD1563547_predicted | hypothetical protein LOC360478 | 1.4 | |||
| Rpl39 | 60S ribosomal protein L39 | translation | 1.5 | ||
| Rps27a | ribosomal protein S27a | translation | 1.4 | 1.5 | |
| Tnfsf13 | tumor necrosis factor ligand superfamily member 13 | ligand, TNF ligand family | 1.3 | 1.3 | cancer, lupus nephritis |
| Umps | uridine 5'-monophosphate synthase | UMP synthesis | 1.4 |
Among the upregulated genes, eight encode ribosomal proteins. The products of ribosomal protein genes—ribosomal proteins—are involved in translation and constitute integral parts of ribosomes [34]. The small 40S ribosomal subunit contains approximately 32 ribosomal proteins (RPS proteins), while the large 60S ribosomal subunit harbors approximately 47 ribosomal proteins (RPL proteins) [34]. Ribosomal proteins are crucial for normal cellular functioning, yet not much is known about their roles in various cellular processes. Recent reports show that the increased expression of ribosomal proteins has also been associated with increased cellular proliferation and growth. Furthermore, the increased expression of ribosomal proteins, such as RPS8, RPL12, RPL23A, RPL27, RPL37, and RPL30, has been found in various different tumor types [34-35]. Furthermore, RPS27 protein partakes in the genotoxic stress response [36-37]. Notwithstanding this, it is unclear if these changes in ribosomal protein expression are causally related to tumorigenesis. In our study, we found a significant increase in the expression of genes encoding for ribosomal proteins. The cellular and organismal reper-cussions of these changes need to be analyzed further.
Another gene that we found to be significantly upregulated in the kidneys of stroked animals was aminolevulinate dehydratase (ALAD)[38]. ALAD is one of the key heme biosynthesis enzymes. This enzyme is important for normal kidney function and is involved in kidney responses to heavy metals and other toxicants [39]. In our study, ALAD expression was 1.3 times higher in the kidneys of stroked rats as compared to controls. The role of ALAD in post-stroke acute kidney injuries needs to be further analyzed.
We also noted an upregulation of the Tnfsf13 gene, also known as APRIL (a proliferation-inducing ligand). Tnfsf13/APRIL belongs to the tumor necrosis factor superfamily and is important for B-cell development, maturation, and survival. It contributes to human autoimmune diseases and cancer. Furthermore, recent studies have shown that Tnfsf13/APRIL is significantly upregalated in the glomeruli and tubulointerstitium in the course of human proliferative lupus nephritis [40-41].
Overall, based on the literature and database analysis, many genes that were upregulated in the kidney tissue of the stroked rats were previously shown to be associated with various neoplasms, including urinary neoplasms, kidney inflammation, and kidney responses to toxic compounds (Table 1).
Altered expression of microRNAs in kidney tissue of stroked rats
Altered levels of histone modifications may also affect expression of microRNAs. Therefore, next we analyzed microRNA expression in the kidneys of control and stroked animals, using microRNA microarrays. Cluster analysis revealed that the kidney tissue of stroked animals was characterized by altered microRNA expression. Specifically, we noted a significant decrease in the levels of miR-221 and a significant increase in the levels of miR-486 and miR-714 (p<0.05) (Figure 5A). Altered expression of miR-221 and miR-486 was independently confirmed by quantitative real-time PCR analysis (Figure 5B).
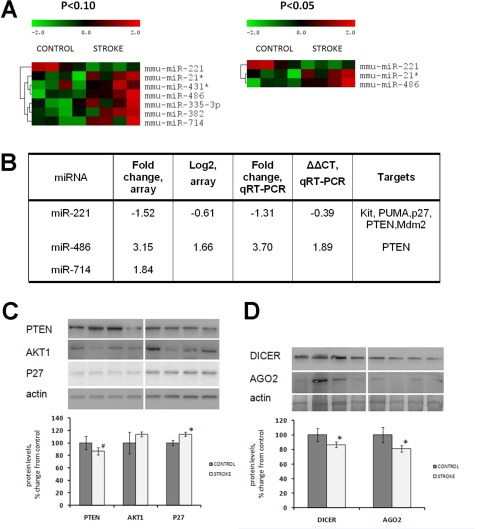
Figure 5. Altered expression of miRNAs, microRNA target proteins and microRNAome machinery in the kidney tissue of control and stroked rats (A) Hierarchical clusters of differentially expressed miRNA genes in the kidney tissue of control and stroked rats (as determined by t-test). (B) Fold changes of miRNA expression as measured by both microarray and qRT-PCR. (C) Levels of miRNA targets PTEN, AKT1 and P27. (D) Levels of miRNA processing enzymes DICER and AGO2. Protein levels relative to those of control animals are shown as the means ± SD, ±significant, p < 0.05, Student's t-test. Representative western blots from among three technical repeats are shown.
MicroRNAs that exhibited altered expression in the kidney tissue of stroked rats are known to regulate a variety of cellular processes by targeting important proteins. miR-221 was shown to target and control expression of several key proteins such as p27 [42-43], phosphatase, tensin homolog (PTEN) [44], Kit oncogene, and others. The most important targets of miR-486 are PTEN and Foxo1a, which negatively affect phosphoinositide-3-kinase (PI3K)/Akt signaling [45].
We noted that levels of p27 were significantly elevated in the kidneys of the stroked rats (Figure 5C). Interestingly, we saw a subtle (p<0.10) decrease in the level of PTEN. This protein is targeted by both miR-221 and miR-486, which exhibit opposite expression tendencies: miR-221 is down-regulated and miR-486 is unregulated in the kidney tissue of stroked animals compared with the control ones. Since PTEN is a target of both of these microRNAs, and miR-221 was 1.5-fold down-regulated, and miR-486 was 3.15-fold up-regulated, PTEN expression in the stroked kidneys was slightly down-regulated as compared to the control kidneys.
The observed decrease of the PTEN level in the kidneys of stroked rats is interesting because it was recently shown that PTEN downregulation may contribute to renal cell hypertrophy and matrix expansion. Also, a decrease of PTEN and elevated levels have been reported in diabetes kidney injury [46]. Recent studies have also reported increased levels of p27 and concurrent activation of Akt signaling in renal cell carcinoma [47].
Having seen some deregulation of microRNA expression in the kidneys of stroked animals, we next analyzed the levels of the microRNA-processing machinery proteins Dicer and Argonaute 2 (Ago2) in the kidney tissue of control and stroked rats. Our analysis revealed a slightly decreased level of both Dicer and Ago2 in the kidneys of stroked rats (Figure 5D). Dicer, a key enzyme for microRNA production, plays important roles in the kidneys. Mice that lacked Dicer in the proximal kidney tubular cells exhibited normal renal function and histology despite a global down-regulation of microRNAs in the renal cortex [48]. Furthermore, these mice were very resistant to renal ischemia-reperfusion injury. They exhibited better renal function, less tissue damage, lower tubular apoptosis, and improved survival as compared to the wild-type animals with normal Dicer levels [48]. Overall, Dicer is very important for kidney function, especially for the juxtaglomerular apparatus [49]. Dicer and microRNAs play important roles in kidney disease [50-51]. However, the role of Dicer and Ago2 in stroke-induced bystander-like responses in the kidneys need to be further substantiated.
Conclusions and outlook
Animals that suffered from ischemic stroke exhibited molecular epigenetic changes in the distal kidney, as well as in heart and liver, tissues. Interestingly, the aforementioned epigenetic changes were the most pronounced in the kidney tissue. The most striking changes were seen in the levels of H3K4 methylation, H3K3 methylation, and acetylation in the kidneys of stroked animals. Additionally, we also noted intriguing gene and microRNA expression changes in the kidney tissues of stroked rats. The sensitivity of kidney tissue to the distal effects of stroke is an important novel finding.
Recent data clearly showed that kidney damage and acute kidney failure is one of the very common, yet overlooked, post-stroke complications [52-53]. Epidemiological evidence suggests that acute kidney damage contributes significantly to post-stroke mortality, yet the mechanisms of post-stroke kidney damage have not been completely understood [53]. Our animal model-based data are the first to show the existence of altered levels of histone modification in the kidney tissues of stroked animals.
These epigenetic changes may significantly affect gene expression and chromatin organization in the kidney cells. Even though the observed epigenetic changes were seemingly subtle, even subtle epigenetic changes may often lead to altered gene expression and altered genome stability. The mechanisms of the occurrence, locus specificity, and cellular repercussions of stroke-induced bystander effect-like epigenetic changes need to be further analyzed. Our study serves as a roadmap for the future analysis of the stroke-induced bystander-like effects in distal tissues and organs.
Materials and Methods
Animal model
Male Long-Evans rats (N= 16, 120-150 days old) were used as subjects. Eight rats received a motor cortex stroke and eight rats served as normal controls.
Surgery: Under isofluorane anaesthesia, a rectangular hole was drilled into the frontal and parietal bones running from + 3 to -4mm anterior/posterior to the Bregma and running laterally from 1.5 to 4.5mm from midline. The focal, permanent devascularization (stroke) has been described in detail elsewhere [26]. Specifically, the dura was removed and a sterile saline-soaked cotton swab was used to wipe the pia and attached blood vessels from the cortical surface (Figure 1). Surgery was performed on the hemisphere contralateral to the preferred paw in a skilled reaching test (data not shown) and was roughly equally divided between the left and right hemispheres. There was no mortality after surgeries; however, for 24 h after the devascularization animals were lethargic and sensitive to touch, after which they returned to their pre-surgical behaviour. Animals were humanely euthanized and kidney, liver and heart were harvested and immediately frozen for analysis.
Western immunoblotting analysis
Western immuno-blotting was conducted on tissue lysates as described before [8] using antibodies against DNMT1 (1:1000; Santa Cruz Biotechnology) MeCP2, DNMT3a, ac-H3K9, me3-H3K9, me3-H3K4, Dicer, Ago2 (1:1000; Abcam), PTEN, p27, Akt1 (1:1000; Cell Signalling). The protein levels were related to those in controls. Two or three technical replicates were used for each immunoblotting.
DNA methylation analysis
Genomic DNA was isolated from tissues, and the cytosine extension assay for determination of the absolute percent of double-stranded unmethylated CCGG sites were conducted as described previously [8].
microRNA expression analysis
MicroRNA microarray analysis was conducted using kidney tissue of control and stroked rats. Total RNA was extracted using TRIzol Reagent (Invitrogen, Burlington, ON). Microarray analysis was performed by LC Sciences (Houston, TX) as described before [8].
Quantitative real-time PCR (qRT-PCR) expression enalysis
qRT-PCRs were performed on samples from all groups using TaqMan Primer sets specific for miR-486 and miR-221. SnoRNA202 served as a control.
Statistical tests
Results are presented as mean±S.D. Statistical analyses were conducted by the Student`s t-test.
Acknowledgments
Work was supported by the CIHR, NSERC and Canadian Stroke Network grants to Olga Kovalchuk and to Bryan Kolb. Anna Kovalchuk was a recipient of the AIHS Summer Research Studentship. We are thankful to Andrey Golubov, Jody Filkowski and Igor Koturbash for technical assistance and to Karen Dow-Cazal for animal care.
Conflicts of Interest
The authors of this manuscript have no conflict of interest to declare.
References
- 1. Donnan GA, Fisher M, Macleod M, Davis SM. Stroke. Lancet. 2008; 371: 1612 -1623. [PubMed] .
- 2. Eriksson D and Stigbrand T. Radiation-induced cell death mechanisms. Tumour Biol. 2010; 31: 363 -372. [PubMed] .
- 3. Ferrer I. Cell death in the normal developing brain, and following ionizing radiation, methyl-azoxymethanol acetate, and hypoxia-ischaemia in the rat. Neuropathol Appl Neurobiol. 1996; 22: 489 -494. [PubMed] .
- 4. Morgan WF and Sowa MB. Non-targeted bystander effects induced by ionizing radiation. Mutat Res. 2007; 616: 159 -164. [PubMed] .
- 5. Hodny Z, Hubackova S, Bartek J. Cytokines shape chemotherapy-induced and 'bystander' senescence. Aging (Albany NY). 2010; 2: 375 -376. [PubMed] .
- 6. Kovalchuk O and Baulch JE. Epigenetic changes and nontargeted radiation effects--is there a link? Environ Mol Mutagen. 2008; 49: 16 -25. [PubMed] .
- 7. Brooks AL. Evidence for 'bystander effects' in vivo. Hum Exp Toxicol. 2004; 23: 67 -70. [PubMed] .
- 8. Koturbash I, Boyko A, Rodriguez-Juarez R, McDonald RJ, Tryndyak VP, Kovalchuk I, Pogribny IP, Kovalchuk O. Role of epigenetic effectors in maintenance of the long-term persistent bystander effect in spleen in vivo. Carcinogenesis. 2007; 28: 1831 -1838. [PubMed] .
- 9. Bird A. Perceptions of epigenetics. Nature. 2007; 447: 396 -398. [PubMed] .
- 10. Feinberg AP. The epigenetics of cancer etiology. Semin Cancer Biol. 2004; 14: 427 -432. [PubMed] .
- 11. Klose RJ and Bird AP. Genomic DNA methylation: the mark and its mediators. Trends Biochem Sci. 2006; 31: 89 -97. [PubMed] .
- 12. Jaenisch R and Bird A. Epigenetic regulation of gene expression: how the genome integrates intrinsic and environmental signals. Nat Genet. 2003; 33 Suppl 245 -254. [PubMed] .
- 13. Rountree MR, Bachman KE, Herman JG, Baylin SB. DNA methylation, chromatin inheritance, and cancer. Oncogene. 2001; 20: 3156 -3165. [PubMed] .
- 14. Goll MG and Bestor TH. Eukaryotic cytosine methyltransferases. Annu Rev Biochem. 2005; 74: 481 -514. [PubMed] .
- 15. Agrawal A, Tay J, Yang GE, Agrawal S, Gupta S. Age-associated epigenetic modifications in human DNA increase its immunogenicity. Aging (Albany NY). 2010; 2: 93 -100. [PubMed] .
- 16. Koch CM and Wagner W. Epigenetic-aging-signature to determine age in different tissues. Aging (Albany NY). 2011; 3: 1018 -1027. [PubMed] .
- 17. Murgatroyd C, Wu Y, Bockmuhl Y, Spengler D. The Janus face of DNA methylation in aging. Aging (Albany NY). 2010; 2: 107 -110. [PubMed] .
- 18. Jenuwein T and Allis CD. Translating the histone code. Science. 2001; 293: 1074 -1080. [PubMed] .
- 19. Schellenberg A, Lin Q, Schuler H, Koch CM, Joussen S, Denecke B, Walenda G, Pallua N, Suschek CV, Zenke M, Wagner W. Replicative senescence of mesenchymal stem cells causes DNA-methylation changes which correlate with repressive histone marks. Aging (Albany NY). 2011; 3: 873 -888. [PubMed] .
- 20. Cheung P and Lau P. Epigenetic regulation by histone methylation and histone variants. Mol Endocrinol. 2005; 19: 563 -573. [PubMed] .
- 21. Saha A, Wittmeyer J, Cairns BR. Chromatin remodelling: the industrial revolution of DNA around histones. Nat Rev Mol Cell Biol. 2006; 7: 437 -447. [PubMed] .
- 22. Dickey JS, Zemp FJ, Martin OA, Kovalchuk O. The role of miRNA in the direct and indirect effects of ionizing radiation. Radiat Environ Biophys. 2011; 50: 491 -499. [PubMed] .
- 23. Sevignani C, Calin GA, Siracusa LD, Croce CM. Mammalian microRNAs: a small world for fine-tuning gene expression. Mamm Genome. 2006; 17: 189 -202. [PubMed] .
- 24. Mack GS. MicroRNA gets down to business. Nat Biotechnol. 2007; 25: 631 -638. [PubMed] .
- 25. Jensen DK and Lal A. Micromanaging senescence. Aging (Albany NY). 2010; 2: 322 -323. [PubMed] .
- 26. Gonzalez CL, Whishaw IQ, Kolb B. Complete sparing of spatial learning following posterior and posterior plus anterior cingulate cortex lesions at 10 days of age in the rat. Neuroscience. 2003; 122: 563 -571. [PubMed] .
- 27. Schubeler D. Epigenomics: Methylation matters. Nature. 2009; 462: 296 -297. [PubMed] .
- 28. Weber M, Hellmann I, Stadler MB, Ramos L, Paabo S, Rebhan M, Schubeler D. Distribution, silencing potential and evolutionary impact of promoter DNA methylation in the human genome. Nat Genet. 2007; 39: 457 -466. [PubMed] .
- 29. Dhasarathy A and Wade PA. The MBD protein family-reading an epigenetic mark? Mutat Res. 2008; 647: 39 -43. [PubMed] .
- 30. Bartova E, Krejci J, Harnicarova A, Galiova G, Kozubek S. Histone modifications and nuclear architecture: a review. J Histochem Cytochem. 2008; 56: 711 -721. [PubMed] .
- 31. Brenner C and Fuks F. A methylation rendezvous: reader meets writers. Dev Cell. 2007; 12: 843 -844. [PubMed] .
- 32. Zacharias N, Sailhamer EA, Li Y, Liu B, Butt MU, Shuja F, Velmahos GC, de Moya M, Alam HB. Histone deacetylase inhibitors prevent apoptosis following lethal hemorrhagic shock in rodent kidney cells. Resuscitation. 2010; .
- 33. Sayyed SG, Gaikwad AB, Lichtnekert J, Kulkarni O, Eulberg D, Klussmann S, Tikoo K, Anders HJ. Progressive glomerulosclerosis in type 2 diabetes is associated with renal histone H3K9 and H3K23 acetylation, H3K4 dimethylation and phosphorylation at serine 10. Nephrol Dial Transplant. 2010; 25: 1811 -1817. [PubMed] .
- 34. van Riggelen J, Yetil A, Felsher DW. MYC as a regulator of ribosome biogenesis and protein synthesis. Nat Rev Cancer. 2010; 10: 301 -309. [PubMed] .
- 35. Kondoh N, Shuda M, Tanaka K, Wakatsuki T, Hada A, Yamamoto M. Enhanced expression of S8, L12, L23a, L27 and L30 ribosomal protein mRNAs in human hepatocellular carcinoma. Anticancer Res. 2001; 21: 2429 -2433. [PubMed] .
- 36. Sun Y, Xiong X, Zhao Y, He H. Ribosomal protein S27-like and S27 interplay with p53-MDM2 axis as a target, a substrate and a regulator. Oncogene. 2011; 30: 1798 -1811. [PubMed] .
- 37. Yu Q, Li J, Tan J, Zhuang L, Banerjee B, Yang XJ, Chau JFL, Lee PL, Hande MP, Li BJ. Ribosomal protein s27-like, a p53-inducible modulator of cell fate in response to genotoxic stress. Cancer Research. 2007; 67: 11317 -11326. [PubMed] .
- 38. Roth KS, Spencer PD, Moses LC, Carter BE. Delta-Aminolevulinic-Acid Dehydratase - Is There a Form Unique to Renal-Cortex. Enzyme. 1990; 43: 17 -25. [PubMed] .
- 39. Oskarsson A and Fowler BA. Effects of Lead on the Heme Biosynthetic-Pathway in Rat-Kidney. Exp Mol Pathol. 1985; 43: 409 -417. [PubMed] .
- 40. Neusser MA, Lindenmeyer M, Edenhofer I, Gaiser S, Kretzler M, Segerer S, Cohen CD. Intrinsic APRIL and BLyS production in human lupus nephritis (LN). Swiss Med Wkly. 2010; 140: 8S -8S. .
- 41. Cohen CD, Neusser MA, Lindenmeyer MT, Edenhofer I, Gaiser S, Kretzler M, Regele H, Segerer S. Intrarenal production of B-cell survival factors in human lupus nephritis. Modern Pathol. 2011; 24: 98 -107. .
- 42. Gillies JK and Lorimer IA. Regulation of p27Kip1 by miRNA 221/222 in glioblastoma. Cell Cycle. 2007; 6: 2005 -2009. [PubMed] .
- 43. le Sage C, Nagel R, Egan DA, Schrier M, Mesman E, Mangiola A, Anile C, Maira G, Mercatelli N, Ciafre SA, Farace MG, Agami R. Regulation of the p27(Kip1) tumor suppressor by miR-221 and miR-222 promotes cancer cell proliferation. EMBO J. 2007; 26: 3699 -3708. [PubMed] .
- 44. Chun-Zhi Z, Lei H, An-Ling Z, Yan-Chao F, Xiao Y, Guang-Xiu W, Zhi-Fan J, Pei-Yu P, Qing-Yu Z, Chun-Sheng K. MicroRNA-221 and microRNA-222 regulate gastric carcinoma cell proliferation and radioresistance by targeting PTEN. BMC Cancer. 2010; 10: 367 [PubMed] .
- 45. Small EM, O'Rourke JR, Moresi V, Sutherland LB, McAnally J, Gerard RD, Richardson JA, Olson EN. Regulation of PI3-kinase/Akt signaling by muscle-enriched microRNA-486. Proc Natl Acad Sci U S A. 2010; 107: 4218 -4223. [PubMed] .
- 46. Dey N, Das F, Mariappan MM, Mandal CC, Ghosh-Choudhury N, Kasinath BS, Choudhury GG. MicroRNA-21 Orchestrates High Glucose-induced Signals to TOR Complex 1, Resulting in Renal Cell Pathology in Diabetes. J Biol Chem. 2011; 286: 25586 -25603. [PubMed] .
- 47. Kim J, Jonasch E, Alexander A, Short JD, Cai S, Wen S, Tsavachidou D, Tamboli P, Czerniak BA, Do KA, Wu KJ, Marlow LA, Wood CG, et al. Cytoplasmic sequestration of p27 via AKT phosphorylation in renal cell carcinoma. Clin Cancer Res. 2009; 15: 81 -90. [PubMed] .
- 48. Wei Q, Bhatt K, He HZ, Mi QS, Haase VH, Dong Z. Targeted deletion of Dicer from proximal tubules protects against renal ischemia-reperfusion injury. J Am Soc Nephrol. 2010; 21: 756 -761. [PubMed] .
- 49. Sequeira-Lopez ML, Weatherford ET, Borges GR, Monteagudo MC, Pentz ES, Harfe BD, Carretero O, Sigmund CD, Gomez RA. The microRNA-processing enzyme dicer maintains juxtaglomerular cells. J Am Soc Nephrol. 2010; 21: 460 -467. [PubMed] .
- 50. Saal S and Harvey SJ. MicroRNAs and the kidney: coming of age. Curr Opin Nephrol Hypertens. 2009; 18: 317 -323. [PubMed] .
- 51. Kato M, Arce L, Natarajan R. MicroRNAs and their role in progressive kidney diseases. Clin J Am Soc Nephrol. 2009; 4: 1255 -1266. [PubMed] .
- 52. Schiller A and Covic A. Kidney and brain--a renal perspective of ‘Les Liaisons Dangereuses’. Nephrol Dial Transplant. 2010; 25: 1370 -1373. [PubMed] .
- 53. Covic A, Schiller A, Mardare NG, Petrica L, Petrica M, Mihaescu A, Posta N. The impact of acute kidney injury on short-term survival in an Eastern European population with stroke. Nephrol Dial Transplant. 2008; 23: 2228 -2234. [PubMed] .