



Introduction
In current aging research, Sir2 is one of the most prominent longevity genes and at the same time the most intensively discussed one. Kaeberlein et al. described Sir2 (silent information regulator 2) as a NAD+-dependent histone deacetylase that extends replicative lifespan of yeast [1]. It is highly conserved in eukaryotes and lent its name to the family of Sirtuins (Sir2- like genes), which has five members in Drosophila (Sir2, Sirt2, Sirt4, Sirt6 and Sirt7) and seven members in mammals.
Findings in C. elegans and Drosophila confirmed a role of Sir2 on lifespan. Tissenbaum and colleagues showed that an increased copy number of Sir2.1 extends lifespan of worms by up to 50 % [2]. In Drosophila, ubiquitous, constitutive overexpression as well as panneuronal overexpression only in the adult animal was shown to extend lifespan [3, 4]. Studies in mice have demonstrated that the Sir2 homolog Sirt1 does not increase life expectancy but promotes a longer health span by prevention of age associated diseases [5-7].
Due to the apparent evolutionarily conserved effect of Sir2/Sirt1 on health and ageing, the physiological functions and protein targets of the enzyme have been intensively studied in mammals in order to cure age-related diseases. SIRT1 was shown to increase insulin sensitivity and regulate energy expenditure through stimulation of oxidative metabolism like fatty acid oxidation and oxidative phosphorylation in mice [5, 8-10]. This predestines the enzyme as a novel target for the treatment of diabetes [11]. In addition, it has been implicated in the etiology and/or cure of cancer, Alzheimer's and other age-related diseases [12-14]. In contrast to mammals, in Drosophila only few studies tried to elucidate physiological functions of Sir2 and the molecular mechanisms, which may mediate its effect on life span. A screen for obesity-inducing genes in Drosophila larvae pointed to a role for Sir2 in regulating fat metabolism and response to amino-acid starvation [15]. Moreover, Sir2 apparently regulates expression of genes involved in fat metabolism and the lack of Sir2 increases fat deposition under normal conditions and consequently impairs starvation survival of flies [16].
It has been hypothesized that Sir2 mediates the effects of dietary restriction (DR) on fly lifespan because decreased activity of the histone deacetylase RPD3, which is down-regulated in DR, increases SIR2 activity [3]. This hypothesis is corroborated by the recent results that revealed a diet-dependent effect of Sir2 on lifespan of Drosophila [17]. In addition, P53 is discussed as a downstream target of RPD3 and SIR2 in mediating longevity [4].
Strong doubts over the lifespan prolonging effect of Sir2 overexpression have recently been expressed [18]. As different genetic backgrounds of control and experimental animals can be a confounder in lifespan experiments, Burnett et al. [19] aimed to validate results for C. elegans and Drosophila with genetically matched controls and could not confirm the positive effect on lifespan. However, a very recent study has supported the initial observation by showing that genetic manipulation of Sir2 in the fly's fat body elongates lifespan in Drosophila [17]. Thus, an urgent need to either confirm or rebut these effects of Sir2 expression levels on lifespan exists. We used the inducible GeneSwitch Gal4/UAS system [20] to drive Sir2 overexpression in the fat body only, which offers two major advantages: first, it is possible to restrict overexpression to the adult animals without affecting development and second, Sir2 overexpressing flies and control flies have the same genetic background since overexpression is induced by treatment with mifepristone (RU486). We found that Sir2 overexpression in the fat body only can promote longevity in both sexes. Furthermore, a detailed analysis of the fat tissue revealed no overt structural changes. However transcriptional profiles of fat bodies after DR or Sir2 overexpression suggest that the interventions promote longevity through independent mechanisms.
Results
Moderate Overexpression of Sir2 in the Fat Body Elongates Lifespan of Drosophila
To avoid differences in genotypes between control and experimental animals, we used the inducible GeneSwitch System [20] to overexpress Sir2 in the fat body of adult flies. Here, tissue-specific overexpression of Sir2 is achieved by administration of mifepristone (RU486) to the F1 generation of a cross of UAS-Sir2 with the driver line S1106-Gal4 while genetically and developmentally identical controls feed on normal medium supplemented with the same dose of the diluent ethanol. We first examined the expression level of Sir2 under the experimental conditions. UAS-Sir2>S1106-Gal4 flies were fed 400 μM RU486 or diluent only for 24 h. Fat bodies were dissected and qPCR for Sir2 mRNA was performed. We detected an approx. 3.5 fold and 2.5 fold up-regulation of Sir2 gene expression in the fat bodies of females and males, respectively (Fig. S1). This system induces a moderate increase of Sir2 expression, which is presumably in the physiological range of expression changes.
Until recently, studies that aimed to investigate the effect of Sir2 on lifespan mainly manipulated its gene expression in whole animals or the nervous system. We decided to study the effect of overexpressing Sir2 in the fat body as it is strongly expressed in this tissue [3]. Moreover, the fat body integrates the response to both immune and metabolic stimuli, which are both important modulators of the aging process [21, 22]. To do so, we determined lifespan of mated UAS-Sir2>S1106-Gal4 flies fed on medium with or without RU486 throughout adult life. Survival curves of females (Fig. 1A) and males (Fig. 1C) overexpressing Sir2 are significantly shifted to the right. As shown in Table 1, median lifespan of females and males treated with RU486 is increased by 12 % and 13 % compared to controls, respectively. These data show that even the relatively weak overexpression of Sir2 induced by the S1106-Gal4 line in the adult fat body promotes longevity in flies, largely by delaying the onset of cohort aging. To rule out the possibility that the GAL4 activator RU486 itself may elongate lifespan of Drosophila, we performed lifespan experiments with flies from a crossing of the w1118 strain and the driver line S1106-Gal4. Flies were treated as described above and no significant shift in survival curves was observed for either females (Fig. 1B) or males (Fig. 1D) compared to controls.
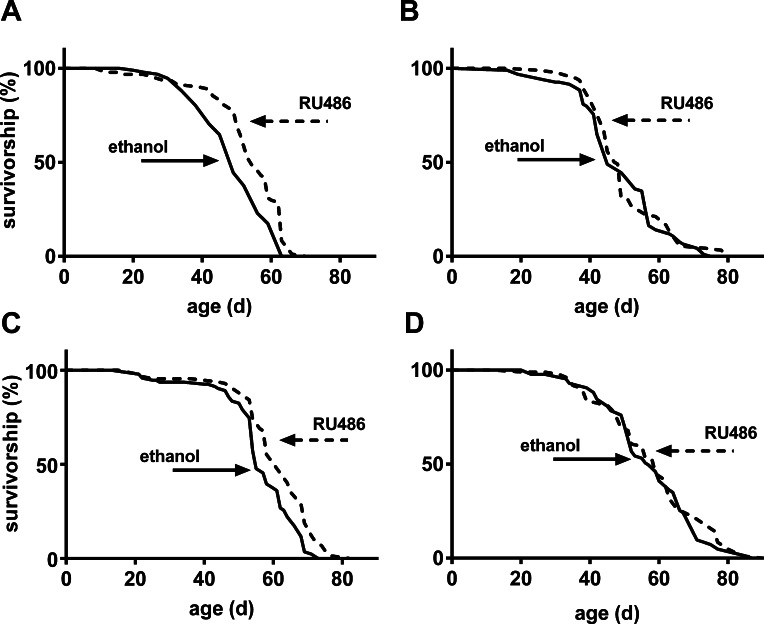
Figure 1. Increased expression of Sir2 in the fat body extended lifespan of male and female flies Mated female (A) or male (C) UAS-Sir2>S1106-Gal4 flies were fed 400 μM RU486 or diluent (ethanol) throughout adult life and lifespan was determined. As a control for a potential lifespan extending effect of RU486, the driver line was crossed to w1118 and female (B) or male control flies (D) of the F1 generation were treated as the experimental flies. For p-values of log rank test see Table 1, n = 100 for each group.
Table 1. Elevated expression levels of Sir2 increased life expectancy of flies
| Genotype | Sex | RU486 | Median lifespan (d) | Median lifespan extension (% of control) | Maximum lifespan (d) | χ2 | p-value | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| UAS-Sir2>S1106-Gal4 | ♀ | − | 49 | 12.3 | 59 | 8.796 | 0.003 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| UAS-Sir2>S1106-Gal4 | ♀ | + | 55 | 62 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| UAS-Sir2>S1106-Gal4 | ♂ | − | 55 | 12.7 | 68 | 13.15 | 0.0003 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| UAS-Sir2>S1106-Gal4 | ♂ | + | 62 | 71 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| w1118>S1106-Gal4 | ♀ | − | 45 | 6.7 | 63 | 0.352 | ns | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| w1118>S1106-Gal4 | ♀ | + | 48 | 63 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| w1118>S1106-Gal4 | ♂ | − | 57 | 3.4 | 70 | 0.015 | ns | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| w1118>S1106-Gal4 | ♂ | + | 59 | 76 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Data derived from log rank analysis of the survival curves shown in Fig. 1. The maximum lifespan is the median of 10 % survival. Log rank test was performed using GraphPad Prism 6.00. ns, not significant. Experiments shown are representatives of two independent experiments. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Gene Expression Profiles Induced by Sir2 Overexpression and Dietary Restriction Do not Match
The finding that fat body Sir2 can promote a long life led us to examine which candidate mechanisms may play a role in conferring Sir2-dependent longevity. As several studies suggest that Sir2 mediates at least parts of the response to dietary restriction (DR)[3, 17] we aimed to elucidate how these two interventions alter gene expression patterns of the fat body and to what extent transcriptional responses match. We performed microarray analysis with RNA isolated from fat bodies of mated, age-matched females either subjected to DR for 7 d or with fat body specific Sir2 overexpression after 24 h induction by RU486. The control and DR food was prepared according to recommendations by Ja et al. [23] to prevent desiccation, which robustly resulted in prolonged lifespan of male and female w1118 flies (Fig. S2). Surprisingly, when comparing gene expression profiles we noticed that only roughly 10 % of up- (p value 0.355) or down-regulated genes (p value 0.383) overlap between both samples (Fig 2A, B). This is a surprising small overlap, indicative for almost completely independent processes, especially when taking into account that fat tissue of age- and gender-matched flies was used for both analyses.
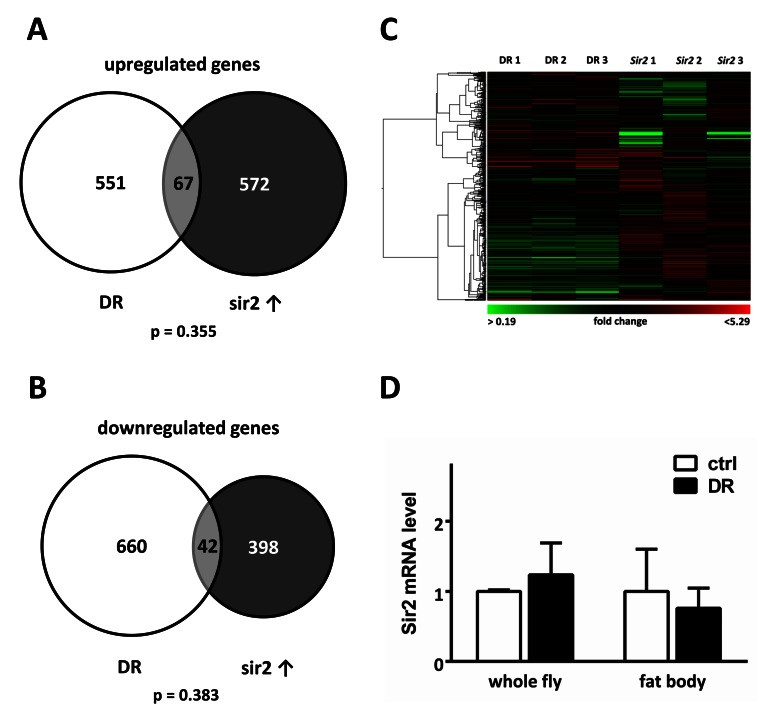
Figure 2. Fat body Sir2 does not play a major role in the DR response Transcriptional profiles of fat bodies from either w1118 females after a 7 d DR period or UAS-Sir2>S1106-Gal4 females after 24 h of Sir2 overexpression were compared. The scaled Venn diagrams depict the actual relationship of the numbers of up-regulated (A) and down-regulated genes (B) of both samples and the number of overlapping genes. Only roughly 10 % of differentially expressed genes are overlapping between DR samples and Sir2 overexpression. Statistical analyses were performed with Fisher's exact test. Genes, which were up-or downregulated > 1.5 fold in at least 2 of 3 microarrays were included in the diagrams shown. Hierarchical clustering of DR and Sir2 Arrays reveals that transcriptional patterns elicited by DR and Sir2 overexpression in the fat body do not show substantial similarities. (D) mRNA levels of Sir2 in female w1118 whole animals and in their fat bodies were unchanged after DR. Shown are mean values of 3 replicates + SEM.
Furthermore, hierarchical clustering of the microarray data shows substantial differences in gene expression patterns induced by both interventions (Fig 2C). Especially genes down-regulated in DR sets are up-regulated in Sir2 gene sets. Additionally, we did not detect an increase of Sir2 transcription in response to DR as reported by other groups, neither in whole animals nor in their fat tissues (Fig. 2D). Taken together, these results imply that fat body Sir2 evokes other cellular processes than those elicited by DR.
Fat Body Sir2 Regulates Lipid Droplet Proteome Associated Genes in a Gender-specific Manner
Based on the transcriptome studies performed with isolated fat tissue from Sir2 overexpressing males and females, we focused on those genes that are relevant for fat tissues. Within this tissue lipid droplets are of central importance to regulate fat utilization [24]. Thus, we hypothesized that fat body Sir2 modulates the composition of the lipid droplet proteome and compared the gene expression data from fat bodies overexpressing Sir2 with a list of Gene IDs corresponding to the lipid droplet proteome as defined by Beller et al. [25]. Corresponding genes of 39 out of 254 lipid droplet associated proteins were found to be differentially regulated in fat bodies overexpressing Sir2 which corresponds to 15 % of the lipid droplet proteome (Fig. 3A; p value 0.007). The (putative) functions of these genes are very diverse and range from lipid storage and fatty acid metabolism, stress response and energy metabolism, to chromatin assembly (Table 2).
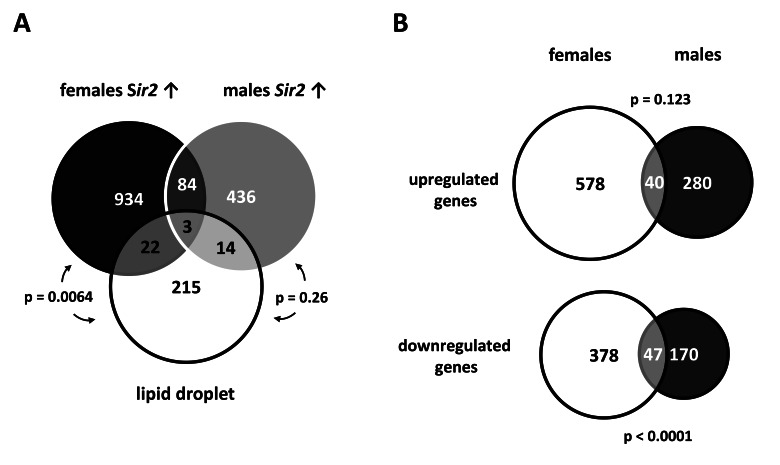
Figure 3. Gene expression profiles of fat bodies overexpressing Sir2 show notable intersections with the lipid droplet proteome (A) Differentially regulated genes (up-and down-regulated) found in Sir2 overexpressing males and females were compared with the lipid droplet proteome as defined by Beller et al. [25]. The Venn diagram shows considerable intersections between these different sample types (statistical analyses were performed with Fisher's exact test; see Table 2 for a list of intersecting genes). (B) To view gender-specific effects, the transcriptional profiles of fat bodies from UAS-Sir2>S1106-Gal4 males and females after 24 h of Sir2 overexpression were compared. The scaled Venn diagrams depict the actual relationship of the numbers of up- and down-regulated genes of both samples and the number of overlapping genes. Genes which were up- or down-regulated > 1.5 fold in at least 2 of 3 microarrays were included in the diagrams shown in (A) and (B).
Table 2. Gene IDs of proteins found to be associated with lipid droplets and genes found to be differentially regulated after Sir2 overexpression in the fat body
| Gene ID | Name | (Putative) function | Gender-specific regulation | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CG31618 | His2A:CG31618 | chromatin assembly or disassembly | ♂ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG5170 | Dodeca-satellite-binding protein 1 | chromosome segregation | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG2512 | alpha-Tubulin at 84D | cytokinesis | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG18000 | short wing | cytoplasmic dynein intermediate chain | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG1049 | CTP:phosphocholine cytidylyltransferase 1 | endocytosis | ♀ ↓ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG5915 | Rab7 | endosome to lysosome transport | ♂ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG11642 | TRAM | phagocytosis | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG6543 | CG6543 | fatty acid β-oxidation | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG12262 | CG12262 | fatty acid β-oxidation | ♀ ↓ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG3523 | Fatty acid synthase | fatty acid synthesis | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG1112 | alpha-Esterase-7 | lipid storage | ♂ ↓ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG5119 | polyA-binding protein | mRNA 3'-UTR binding | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG9748 | belle | ATP-dependent RNA helicase activity | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG9325 | hu li tai shao | axon guidance | ♀ ↓ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG7920 | CG7920 | neurogenesis | ♂ ↓ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG3725 | Calcium ATPase at 60A | neuromuscular synaptic transmission | ♂ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG8322 | ATP citrate lyase | citrate metabolism | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG4233 | Glutamate oxaloacetate transaminase 2 | L-aspartate:2-oxoglutarate aminotransferase activity | ♀ ↑ | ♂↑ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG7998 | Malate dehydrogenase 2 | TCA cycle | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG2968 | lethal (1) G0230 | hydrogen-exporting ATPase activity | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG1683 | Adenine nucleotide translocase 2 | mitochondrial ATP:ADP antiporter activity | ♀ ↑ | ♂↓ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG7834 | CG7834 | oxidative phosphorylation | ♂ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG10639 | CG10639 | oxidoreductase activity | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG5590 | CG5590 | oxidoreductase activity | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG5809 | calcium-binding protein 1 | cell redox homeostasis | ♂ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG6988 | Protein disulfide isomerase | protein disulfide isomerase activity | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG5834 | Hsp70Bbb | stress response | ♂ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG5436 | Heat shock protein 68 | stress response | ♂ ↓ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG8937 | Heat shock protein cognate 1 | unfolded protein response | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG7808 | Ribosomal protein S8 | structural constituent of ribosomes | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG1524 | Ribosomal protein S14a | structural constituent of ribosomes | ♂ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG4087 | Ribosomal protein LP1 | structural constituent of ribosomes | ♂ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG7014 | Ribosomal protein S5b | structural constituent of ribosomes | ♂ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG9282 | Ribosomal protein L24 | structural constituent of ribosomes | ♂ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG3373 | Hemomucin | biosynthetic process | ♀ ↓ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG6415 | CG6415 | glycine catabolism | ♀ ↑ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG2852 | CG2852 | reproductive process | ♀ ↓ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG18212 | aluminum tubes | unknown | ♀ ↑ | ♂↓ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG9186 | CG9186 | unknown | ♂ ↓ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gene IDs of proteins which are associated with lipid droplets [25] and of genes which were differentially regulated in 2 of 3 microarrays of fat bodies of female or male UAS-Sir2>S1106-Gal4 flies after feeding of RU486. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
When comparing gene expression patterns of male and female fat bodies overexpressing Sir2 we found great differences between genders. Only 40 of 619 genes up-regulated in females were also found within the 320 genes up-regulated in males (Fig. 3B, p value 0.123). This overlap corresponds to 6.5 % and 12.5 % of up-regulated genes in females and males, respectively. The situation was different for down-regulated genes where 11 % and 22 % were overlapping between females and males, respectively (p value < 0.0001). These gender-specific differences are mirrored in the comparison of genes affected by Sir2 overexpression and the genes corresponding to proteins associated with lipid droplets. Considering the data for males and females separately 17 (7 %) and 25 (10 %) of the lipid droplet proteome genes are regulated, respectively (Fig. 3A). But only 1 gene (Glutamate oxaloacetate transaminase 2) is up-regulated in both (Table 2) whereas all other genes are not or inversely regulated in the respective other gender. Taken together, these results point to a role for fat body Sir2 in modulating the composition of the lipid droplet proteome, which is obviously gender-specific.
Fat Body Sir2 Increases Starvation Resistance in Male Flies
The fat body is the major storage organ of the fly, which regulates the response to differing nutrient conditions. Sir2 is capable of sensing cellular energy levels, as its enzymatic activity is NAD+- dependent [26]. We hypothesized that fat body Sir2 participates in regulating the response to changes in nutrient conditions when promoting longevity. One measure for the ability to react to changing nutrient supply is to determine starvation resistance. Mated UAS-Sir2>S1106-Gal4 flies fed on 1 % agar with or without RU486 and starvation medium was changed every 2-3 days. Dead flies were counted 1-3 times a day. Fat body Sir2 appeared to have no impact on female starvation resistance as median lifespan of control and Sir2 overexpressing flies showed no difference (Fig. 4A, C). However, fat body Sir2 increased starvation survival in males by 13 % (Fig. 4B, C). In line with the transcript-tome analysis we found a small gender-specific reaction to starvation after Sir2 overexpression in the fat body.
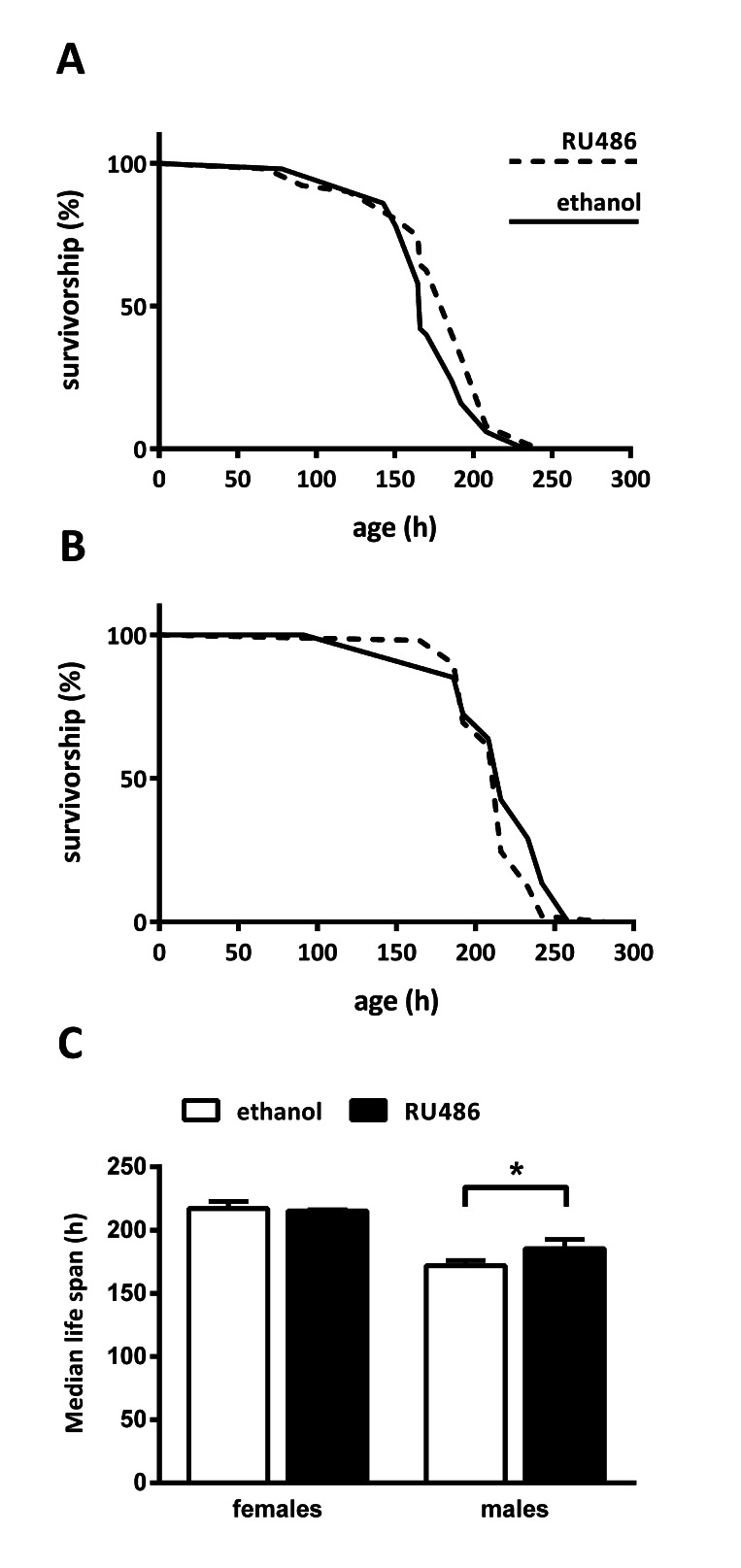
Figure 4. Fat body Sir2 increases starvation survival in males Mated female (A) or male (B) UAS-Sir2>S1106-Gal4 flies fed on 1 % agar containing 400 μM RU486 or diluent (ethanol) and dead flies were scored 1-3 times a day. (C) Median lifespan of females was 217 ± 6 h and 215 ± 1 h in control and experimental flies, respectively. For males median lifespans were 172 ± 4 h and 185 ± 7 h for control and Sir2 overexpressing flies, respectively. Data are represented as means + SEM. Log rank analysis was performed using GraphPad Prism 6.00, n = 50.
No Influence of Sir2 on Fat Storage and Fat Cell Morphology
As Sir2 overexpression in the fat body did not only extend lifespan of the flies but also influenced the relative abundance of lipid droplet proteome associated genes and starvation resistance in a gender-specific manner we wanted to elucidate the role of fat body Sir2 on fat storage and morphology of the fat cells. We tested if Sir2 overexpression affects relative body fat content, body weight, the morphology of the fat cells and the size of the fat droplets in males and females.
Mated UAS-Sir2>S1106-Gal4 males or females were fed on medium containing RU486 or ethanol for 7 d before they were weighed and relative body fat content was quantified by a coupled colorimetric assay according to Hildebrandt et al. [27]. Neither sex showed fat body Sir2 - dependent effects on fat storage (Fig. 5A) and body weight (Fig. 5B). To analyze changes in the morphology of the fat body we performed Bodipy staining with L3 larvae (Fig. 5 D, G), adult females (Fig. 5 E, H) and males (Fig. 5 F, I). Although fat cell size and filling levels of fat stores obviously varied substantially between all sample types, no effect of Sir2 overexpression (Fig. 5 D-I, bottom row) was observed on these parameters compared to controls (Fig. 4 D-I, top row). We randomly selected 14 pictures comprising 7 biological replicates and measured diameters of fat droplets of the larval samples (Fig. 5C) to confirm the results of the qualitative analysis of the tissues. Altogether, these results do not reveal an obvious phenotype in fat body cells following Sir2 over-expression, regardless of developmental stage or sex.
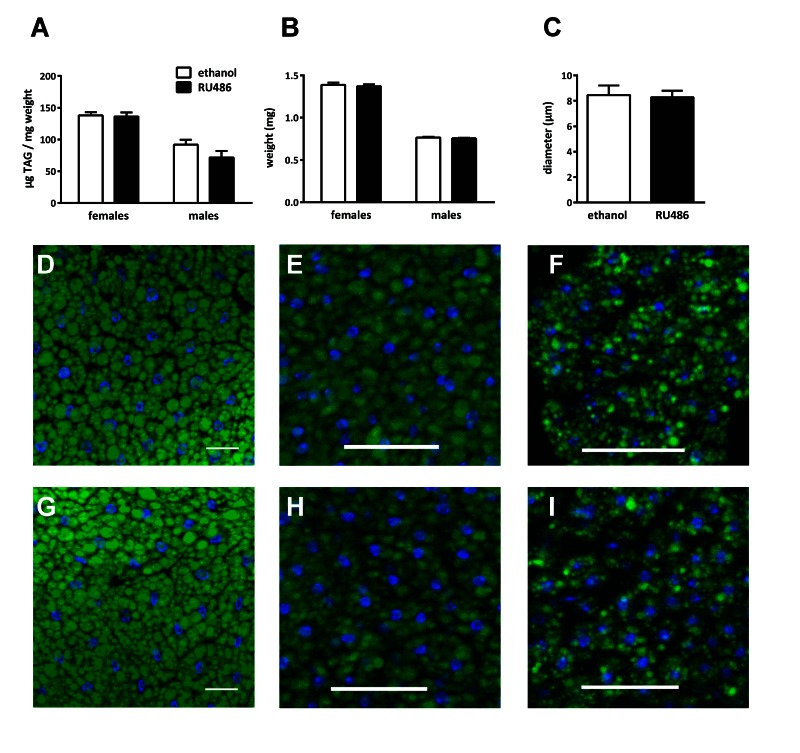
Figure 5. Sir2 overexpression in the fat body did not affect fat storage and fat cell morphology Relative body fat content as quantified by coupled colorimetric assay (A) and body weight (B) of male and female UAS-Sir2>S1106-Gal4 flies feeding on medium with or without 400 μM RU486 showed no significant differences. Diameters of fat droplets were measured in fat bodies of UAS-Sir2>S1106-Gal4 L3 larvae after Bodipy staining (C) and no differences were observed. Bodipy staining furthermore reveals no differences in size and structure of fat cells as well as droplet size in L3 larvae (D, G), adult females (E, H) or males (F, I) after feeding on ethanol (top row) or RU486 (bottom row). A-C, Data are represented as means + SEM. Mann-Whitney-U-Test was performed using GraphPad Prism 6.00, n = 4 for coupled colorimetric assay and body weight, n = 7 for fat droplet diameter measurements. D-I, scale bar represents 50 μm.
Discussion
The aim of this study was to test whether tissue-specific overexpression of Sir2 in the fat body of adult flies can elongate lifespan and to elucidate the mechanisms by which fat body Sir2 promotes longevity. We showed that moderate (roughly 3-fold) Sir2 overexpression in the fat body can extend median lifespan of both male and female Drosophila melanogaster by about 13 %. With this study, we provide additional independent evidence for the concept of Sir2 as a longevity gene in Drosophila as suggested by earlier studies [3, 4, 16, 17]. The UAS-line used in our study has not been employed for lifespan experiments so far and does not show increased expression of dnaJ-H, which has been shown to affect apoptosis control of Sir2 and may also interfere with Sir2 functions with regards to longevity [28]. Importantly, we used the inducible GeneSwitch Gal4 System; this removes the use of different genetic backgrounds in control and Sir2 overexpressing flies, a potentially severe confounder in lifespan studies. Geneswitch also allows overexpression during adulthood only, thus removing any developmental testing potential gerontogenes from a therapeutic perspective. However, our results contradict the study from Burnett et al. [19] who did not observe an effect of ubiquitous overexpression of Sir2 on Drosophila lifespan. This may be due to antagonistic effects on lifespan caused by Sir2 overexpression during development, and/or due to the site of overexpression; by restricting overexpression of Sir2 to the fat body, possibly harmful effects caused by overexpression in other tissues may be excluded. In this context it is notable that overexpression of Sir2 in the nervous system prolonged lifespan of flies [3]. These results are in line with the observation that only these two tissues express SIR2 protein at high levels [3].
We also aimed to test different hypotheses of how fat body Sir2 may confer longevity. Our first working hypothesis was to presume that Sir2 overexpression mimics dietary restriction, which has been proposed by other studies [3, 17, 29]. This led us to compare microarray data generated from fat bodies overexpressing Sir2 with those obtained from fat bodies of animals subjected to DR. As the induced transcriptional changes of both interventions were different our data did not confirm this hypothesis. Instead they support the alternative hypothesis that DR and Sir2 overexpression are indendent processes. Furthermore, dietary restriction had no effect on Sir2 mRNA levels in fat tissue. Our results join a canon of studies presenting conflicting results regarding this topic. Previous findings in yeast [30-33], flies [3, 17, 19] and rodents [34, 35] are inconsistent, which may be due to application of different dietary /caloric restriction regimes, different sites of ectopic expression or knockout of Sir2 and/or genetic backgrounds. We used the food preparations recommended by Ja et al. [23] to circumvent adverse side effects caused by desiccation of the flies, which robustly produced a DR response in both sexes and the food used in the Sir2 experiments was identical except for addition of RU486 dissolved in ethanol or ethanol alone. However, the data presented here suggest that fat body Sir2 does not mediate the effects of DR on lifespan of Drosophila.
It has also been proposed that Sir2 in the fat body plays a role in regulating fat storage and mobilization as Sir2 / Sirt1 has been implicated in regulation of fat metabolism in flies and mammals. Firstly, we compared the part of the fat body's transcriptome that is altered by Sir2 overexpression with those genes coding for the lipid droplet proteome as these organelles are known to be central players for the control of fat storage and mobilization [36-39]. A considerable overlap between these data sets suggested a role for fat body Sir2 in modulating the composition of the lipid droplet proteome, a hypothesis that is supported by the statistical data. Nevertheless, as lipid droplets have been shown to regulate aging processes [40] our data provide a good starting point to unravel the putative role of Sir2 in lipid droplet biology. The overlap with genes coding for lipid droplet proteins was highly sex specific, a feature that was also seen for starvation resistance and the Sir2 induced transcriptional changes in fat tissues.
Our findings that fat storage, body weight and lipid droplet size were not changed after Sir2 overexpression, contrast with other recent studies which demonstrated an impact of RNAi-mediated knockout of Sir2 in the fat body or constitutive whole body knockout, respectively, on fat metabolism [16]. Furthermore, data from mouse studies indicate a role for Sir2 in the regulation of lipid metabolism [8, 10]. These divergences may result from the different expression system used in our study. We aimed to overexpress Sir2 only moderately in the fat body and nowhere else, and only during adulthood. This strategy should have only minimal confounding effects on e.g. fat storage caused by complete silencing of Sir2 in either the fat body/liver or the whole animal.
Overall, we conclusively showed that Sir2 is a longevity gene in Drosophila. With this study we provide additional independent evidence for its ability to extend lifespan of male and female flies when moderately overexpressed in the adult fat body. We also obtained transcriptional profiles elicited by this overexpression and propose a role for Sir2 in lipid droplet biology especially under conditions of starvation. However, the precise conditions under which Sir2 affects fat utilization and the molecular mechanisms involved in this function remain to be elucidated. Furthermore, our transcriptional data do not support the idea of Sir2 mediating the response to dietary restriction. This study again highlights the relevance of Sir2 for aging processes and its potential as a pharmacological target to cure age-related diseases.
Methods
Fly Strains and Husbandry
Males of the driver line S1106-Gal4 (gift from R. Kuehnlein, MPI Goettingen, Germany [20]) were crossed to virgin females of UAS-Sir2 (gift from Kyung-Tai Min,[28]) to generate flies overexpressing Sir2 in the fat body after induction by RU486. As a control for RU486-dependent effects on lifespan a control crossing using the driver line and the w1118 strain (Bloomington Drosophila Stock Center, # 5905) was performed. Fly stocks were raised on standard cornmeal-agar medium at 25 °C under a 12 h/12 h light/dark cycle.
Lifespan Experiments
For lifespan experiments the F1 generation of the crossings mentioned above were allowed to mate for two days after eclosion before they were set on medium containing 5 % (w/v) yeast extract (Becton Dickinson), 5 % (w/v) sucrose, 8.6 % cornmeal, 0.5 % (w/v) agar, 0.03 % (v/v) propionic acid, 0.3 % (v/v) methyl-4-hydroxybenzoat and either 400 μM RU486 (diluted from a 10 mM stock in 90 % ethanol) for the experimental or the corresponding volume of 90 % ethanol for the control groups. Flies were kept at a density of 25 animals per vial (68 ml, Greiner Bio One) at 25 °C, 40-60 % humidity and a 12 h/12h light/dark cycle. Medium was changed every 3 – 4 days and dead flies were counted.
Dietary Restriction (DR)
The DR experiments were performed with medium prepared as recommended in Ja et al. to provide sufficient water supply [23]. Control medium contained 5 % (w/v) yeast extract (Becton Dickinson), 5 % (w/v) sucrose, 8.6 % cornmeal, 0.5 % (w/v) agar, 0.03 % (v/v) propionic acid, 0.3 % (v/v) methyl-4-hydroxybenzoat. DR medium contained only 0.25 % yeast extract. w1118 flies were allowed to mate for 2 days before genders were separated and either used for lifespan studies as described above or fed on control or DR medium for 7 d prior to dissection of fat bodies and RNA isolation.
Microarray Analysis
Microarray analyses were essentially performed as described earlier [41, 42]. In brief, cDNA synthesis from RNA isolated from fat bodies was performed with Prime Script RT (Takara) using the following primers: OdT T7 I (5'-GAG AGA GGA TCC AAG TAC TAA TAC GAC TCA CTA TAG GGA GAT TTT TTT TTT TTT TTT TTT TTT T G/A/C-3') and CapFinder Sp6rG (5'-CAG CGG CCG CAG ATT TAG GTG ACA CTA TAG A rGrGrG-3'). cDNA was amplified with OdT T7 II (5'-GAG AGA GGA TCC AAG TAC TAA TAC GAC TCA CTA TAG G-3') and Adaptor Sp6rG (5'-GAC GCC TGC AGG CGA TGA ATT TAG G-3') and LA Taq polymerase [43]. cDNA was transcribed with MEGAscript® T7 (Ambion) including aminoallyl-UTP (Ambion) and subsequently labeled with Alexa Fluor 555 or 647 (Invitrogen) for control or experimental sample, respectively. Samples were hybridized to Drosophila 14 k V2 (Canadian Drosophila Microarray Centre, Toronto, Canada) and scanned in a GenePix 4000B Microarray Scanner (Axon Instruments, Molecular Devices). Data acquisition, normalization and analysis including hierarchical clustering were carried out with the corresponding programs GenePix 6.0 and Acuity 4.1 (Axon Instruments, Molecular Devices). Scaled Venn diagrams were drawn with Wolfram Mathematica [44]. The microarray data have been deposited at the GEO database (Gene Expression Omnibus (http://www.ncbi.nlm.nih.gov/geo/)) with the accession number GSE45974.
Quantitative real-time PCR
Mated Sir2>S1106-Gal4 flies fed on medium containing RU486 or ethanol as described for lifespan experiments for 24 h. RNA was isolated from the abdomen of 3 animals for one biological replicate and reverse transcription was performed using OdT-T7I Primer and SuperScript III Reverse Transcriptase (Invitrogen). qPCR was performed in a StepOne Real-Time PCR System (Applied Biosystems) using the DyNAmo Flash SYBR Green qPCR Kit (Finnzymes) and the following primers: Sir2 forward (5'-GCC TCC AGG ACA GTT AGC AG-3'), Sir2 reverse (5'- CGT GAT ACC GGG AAT TGA AG-3'), RpL32 forward (5'- TTG GCT TCG GTT TCC GGC AAG-3'), RpL32 reverse (5'-ATC GAT CCG ACT GGT GGC GGAT-3'). RpL32 was used as a reference gene and expression data were analyzed using the method described by Pfaffl [45].
Relative Body Fat Quantification and Body Weight Measurement
A coupled colorimetric assay was performed using the Triglycerides Kit from Thermo Fisher Scientific (# 981786) according to the protocol from Hildebrandt et al. [27]. Briefly, after mating and feeding on medium containing RU486 or ethanol for 7 d, for one biological replicate 8 males or 5 females were weighed on a digital accuracy balance and homogenized in 1 ml 0.05 % Tween-20 with 3 zirconium beads (2,8 mm) for 2 min at 3.25 m/s in a Bead Ruptor 24 (Omni International). Lysates were heat-inactivated for 5 min at 70 °C, centrifuged and supernatant was incubated with the Triglyceride Solution for 30 min at 37 °C. Absorbance was measured at 562 nm and glyceryl trioleate served as a standard.
BODIPY Staining and Determination of Diameters of Lipid Droplets
After feeding on RU486 or control medium for 24 h L3 larvae or adults generated by crossing UAS-Sir2 flies to the driver line S1106-Gal4 were dissected, fixed and fat bodies were stained with a solution containing 1 μg/ml BODIPY (Invitrogen) and 5 μg/ml Hoechst (Sigma) for 30 min at room temperature. Slides were mounted in Mowiol/DABCO (Roth) and photographed with a Zeiss Axio imager 7.1 microscope equipped with a Zeiss Axiocam MRM camera within the next two days. 14 photographs taken from 7 biological replicates were analyzed per group with AxioVision 4.1 (Zeiss). To measure fat droplet diameters a line was drawn across each picture and only droplets intersecting with the line were measured using a measurement tool implemented in the program.
Starvation Resistance
Mated Sir2>S1106-Gal4 flies were kept on 1 % agar containing either 400 μM RU486 or ethanol and vials were changed every 2-3 days. Dead flies were scored 1-3 times a day.
Statistics and Bioinformatics
All statistics were performed using GraphPad Prism version 6.00 for Windows, GraphPad Software, La Jolla California USA, www.graphpad.com. Lifespan and starvation survival data were analyzed by log rank test and data from qPCR, body fat quantification, body weight and diameters of lipid droplets by Mann-Whitney-U Test. Statistical analyses of Venn-diagram data were performed with Fisher's exact test.
Supplementary Materials
Acknowledgments
The authors would like to thank David Clancy for helpful discussions and critically reading the manuscript. We also thank Annekatrin Daniel and Britta Laubenstein for technical assistance. We thank Kyung Tai-Min and Ronald Kuehnlein for the kind gift of fly stocks. This work was supported by a grant from the German Ministry of Education and Research in the framework of “Ernährungsforschung – für ein gesundes Leben, Modul: Biomedizinische Ernährungsforschung – Vision Epifood”, and by the Excellence Cluster of the German Research Foundation (DFG) “Inflammation@ Interfaces”.
Conflicts of Interest
The authors have no conflicting interest to declare.
References
- 1. Kaeberlein M, McVey M, Guarente L. The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes Dev. 1999; 13: 2570 -2580. [PubMed] .
- 2. Tissenbaum HA and Guarente L. Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans. Nature. 2001; 410: 227 -230. [PubMed] .
- 3. Rogina B and Helfand SL. Sir2 mediates longevity in the fly through a pathway related to calorie restriction. Proc Natl Acad Sci U S A. 2004; 101: 15998 -16003. [PubMed] .
- 4. Bauer JH, Morris SN, Chang C, Flatt T, Wood JG, Helfand SL. dSir2 and Dmp53 interact to mediate aspects of CR-dependent lifespan extension in D. melanogaster. Aging (Albany NY). 2009; 1: 38 -48. [PubMed] .
- 5. Bordone L, Cohen D, Robinson A, Motta MC, van Veen E, Czopik A, Steele AD, Crowe H, Marmor S, Luo J, Gu W, Guarente L. SIRT1 transgenic mice show phenotypes resembling calorie restriction. Aging Cell. 2007; 6: 759 -767. [PubMed] .
- 6. Banks AS, Kon N, Knight C, Matsumoto M, Gutierrez-Juarez R, Rossetti L, Gu W, Accili D. SirT1 gain of function increases energy efficiency and prevents diabetes in mice. Cell Metab. 2008; 8: 333 -341. [PubMed] .
- 7. Herranz D, Munoz-Martin M, Canamero M, Mulero F, Martinez-Pastor B, Fernandez-Capetillo O, Serrano M. Sirt1 improves healthy ageing and protects from metabolic syndrome-associated cancer. Nat Commun. 2010; 1: 3 [PubMed] .
- 8. Feige JN, Lagouge M, Canto C, Strehle A, Houten SM, Milne JC, Lambert PD, Mataki C, Elliott PJ, Auwerx J. Specific SIRT1 activation mimics low energy levels and protects against diet-induced metabolic disorders by enhancing fat oxidation. Cell Metab. 2008; 8: 347 -358. [PubMed] .
- 9. Milne JC, Lambert PD, Schenk S, Carney DP, Smith JJ, Gagne DJ, Jin L, Boss O, Perni RB, Vu CB, Bemis JE, Xie R, Disch JS, et al. Small molecule activators of SIRT1 as therapeutics for the treatment of type 2 diabetes. Nature. 2007; 450: 712 -716. [PubMed] .
- 10. Wang RH, Kim HS, Xiao C, Xu X, Gavrilova O, Deng CX. Hepatic Sirt1 deficiency in mice impairs mTorc2/Akt signaling and results in hyperglycemia, oxidative damage, and insulin resistance. J Clin Invest. 2011; 121: 4477 -4490. [PubMed] .
- 11. Houtkooper RH, Pirinen E, Auwerx J. Sirtuins as regulators of metabolism and healthspan. Nat Rev Mol Cell Biol. 2012; 13: 225 -238. [PubMed] .
- 12. Albani D, Polito L, Forloni G. Sirtuins as novel targets for Alzheimer's disease and other neurodegenerative disorders: experimental and genetic evidence. J Alzheimers Dis. 2010; 19: 11 -26. [PubMed] .
- 13. Yi J and Luo J. SIRT1 and p53, effect on cancer, senescence and beyond. Biochim Biophys Acta. 2010; 1804: 1684 -1689. [PubMed] .
- 14. Dali-Youcef N, Lagouge M, Froelich S, Koehl C, Schoonjans K, Auwerx J. Sirtuins: the ‘magnificent seven’, function, metabolism and longevity. Ann Med. 2007; 39: 335 -345. [PubMed] .
- 15. Reis T, Van Gilst MR, Hariharan IK. A buoyancy-based screen of Drosophila larvae for fat-storage mutants reveals a role for Sir2 in coupling fat storage to nutrient availability. PLoS Genet. 2010; 6: e1001206 [PubMed] .
- 16. Banerjee KK, Ayyub C, Sengupta S, Kolthur-Seetharam U. dSir2 deficiency in the fatbody, but not muscles, affects systemic insulin signaling, fat mobilization and starvation survival in flies. Aging (Albany NY). 2012; 4: 206 -223. [PubMed] .
- 17. Banerjee KK, Ayyub C, Ali SZ, Mandot V, Prasad NG, Kolthur-Seetharam U. dSir2 in the adult fat body, but not in muscles, regulates life span in a diet-dependent manner. Cell Rep. 2012; 2: 1485 -1491. [PubMed] .
- 18. Couzin-Frankel J. Genetics. Aging genes: the sirtuin story unravels. Science. 2011; 334: 1194 -1198. [PubMed] .
- 19. Burnett C, Valentini S, Cabreiro F, Goss M, Somogyvari M, Piper MD, Hoddinott M, Sutphin GL, Leko V, McElwee JJ, Vazquez-Manrique RP, Orfila AM, Ackerman D, et al. Absence of effects of Sir2 overexpression on lifespan in C. elegans and Drosophila. Nature. 2011; 477: 482 -485. [PubMed] .
- 20. Roman G, Endo K, Zong L, Davis RL. P[Switch], a system for spatial and temporal control of gene expression in Drosophila melanogaster. Proc Natl Acad Sci U S A. 2001; 98: 12602 -12607. [PubMed] .
- 21. Arrese EL and Soulages JL. Insect fat body: energy, metabolism, and regulation. Annu Rev Entomol. 2010; 55: 207 -225. [PubMed] .
- 22. Eleftherianos I and Castillo JC. Molecular mechanisms of aging and immune system regulation in Drosophila. Int J Mol Sci. 2012; 13: 9826 -9844. [PubMed] .
- 23. Ja WW, Carvalho GB, Zid BM, Mak EM, Brummel T, Benzer S. Water- and nutrient-dependent effects of dietary restriction on Drosophila lifespan. Proc Natl Acad Sci U S A. 2009; 106: 18633 -18637. [PubMed] .
- 24. Kuhnlein RP. The contribution of the Drosophila model to lipid droplet research. Prog Lipid Res. 2011; 50: 348 -356. [PubMed] .
- 25. Beller M, Riedel D, Jansch L, Dieterich G, Wehland J, Jackle H, Kuhnlein RP. Characterization of the Drosophila lipid droplet subproteome. Mol Cell Proteomics. 2006; 5: 1082 -1094. [PubMed] .
- 26. Imai S, Armstrong CM, Kaeberlein M, Guarente L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature. 2000; 403: 795 -800. [PubMed] .
- 27. Hildebrandt A, Bickmeyer I, Kuhnlein RP. Reliable Drosophila body fat quantification by a coupled colorimetric assay. PLoS One. 2011; 6: e23796 [PubMed] .
- 28. Griswold AJ, Chang KT, Runko AP, Knight MA, Min KT. Sir2 mediates apoptosis through JNK-dependent pathways in Drosophila. Proc Natl Acad Sci U S A. 2008; 105: 8673 -8678. [PubMed] .
- 29. Antosh M, Whitaker R, Kroll A, Hosier S, Chang C, Bauer J, Cooper L, Neretti N, Helfand SL. Comparative transcriptional pathway bioinformatic analysis of dietary restriction, Sir2, p53 and resveratrol life span extension in Drosophila. Cell Cycle. 2011; 10: 904 -911. [PubMed] .
- 30. Kaeberlein M, Steffen KK, Hu D, Dang N, Kerr EO, Tsuchiya M, Fields S, Kennedy BK. Comment on “HST2 mediates SIR2-independent life-span extension by calorie restriction”. Science. 2006; 312: 1312 author reply 1312 [PubMed] .
- 31. Kaeberlein M, Kirkland KT, Fields S, Kennedy BK. Sir2-independent life span extension by calorie restriction in yeast. PLoS Biol. 2004; 2: E296 [PubMed] .
- 32. Lin SJ, Defossez PA, Guarente L. Requirement of NAD and SIR2 for life-span extension by calorie restriction in Saccharomyces cerevisiae. Science. 2000; 289: 2126 -2128. [PubMed] .
- 33. Lamming DW, Latorre-Esteves M, Medvedik O, Wong SN, Tsang FA, Wang C, Lin SJ, Sinclair DA. HST2 mediates SIR2-independent life-span extension by calorie restriction. Science. 2005; 309: 1861 -1864. [PubMed] .
- 34. Chen D, Bruno J, Easlon E, Lin SJ, Cheng HL, Alt FW, Guarente L. Tissue-specific regulation of SIRT1 by calorie restriction. Genes Dev. 2008; 22: 1753 -1757. [PubMed] .
- 35. Boily G, Seifert EL, Bevilacqua L, He XH, Sabourin G, Estey C, Moffat C, Crawford S, Saliba S, Jardine K, Xuan J, Evans M, Harper ME, et al. SirT1 regulates energy metabolism and response to caloric restriction in mice. PLoS One. 2008; 3: e1759 [PubMed] .
- 36. Athenstaedt K and Daum G. The life cycle of neutral lipids: synthesis, storage and degradation. Cell Mol Life Sci. 2006; 63: 1355 -1369. [PubMed] .
- 37. Beller M, Bulankina AV, Hsiao HH, Urlaub H, Jackle H, Kuhnlein RP. PERILIPIN-dependent control of lipid droplet structure and fat storage in Drosophila. Cell Metab. 2010; 12: 521 -532. [PubMed] .
- 38. Londos C, Sztalryd C, Tansey JT, Kimmel AR. Role of PAT proteins in lipid metabolism. Biochimie. 2005; 87: 45 -49. [PubMed] .
- 39. Olofsson SO, Bostrom P, Andersson L, Rutberg M, Levin M, Perman J, Boren J. Triglyceride containing lipid droplets and lipid droplet-associated proteins. Curr Opin Lipidol. 2008; 19: 441 -447. [PubMed] .
- 40. Goldberg AA, Bourque SD, Kyryakov P, Boukh-Viner T, Gregg C, Beach A, Burstein MT, Machkalyan G, Richard V, Rampersad S, Titorenko VI. A novel function of lipid droplets in regulating longevity. Biochem Soc Trans. 2009; 37: 1050 -1055. [PubMed] .
- 41. Wagner C, Isermann K, Fehrenbach H, Roeder T. Molecular architecture of the fruit fly's airway epithelial immune system. BMC Genomics. 2008; 9: 446 [PubMed] .
- 42. Abdelsadik A and Roeder T. Chronic activation of the epithelial immune system of the fruit fly's salivary glands has a negative effect on organismal growth and induces a peculiar set of target genes. BMC Genomics. 2010; 11: 265 [PubMed] .
- 43. Schramm G, Bruchhaus I, Roeder T. A simple and reliable 5'-RACE approach. Nucleic Acids Res. 2000; 28: E96 [PubMed] .
- 44. Chandler S and Weisstein EW. The perfect Venn diagram. Wolfram Demonstration Project. 2012; .
- 45. Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001; 29: e45 [PubMed] .