




Introduction
Aging is a multifaceted process that results in increases in inflammation, oxidative stress (OS) and contributes to cellular senescence, eventually leading to a decline in organ function and death [1–4]. Aging and other neurological or neurodegenerative disorders result in deficits in cognitive and motor performance as a result of enhanced susceptibility to the long-term effects of increased OS and inflammation [5,6]. A decrease in endogenous antioxidant mechanisms with aging increases the vulnerability of the brain to oxidative damage as a result of exacerbated OS [7]. In addition, the aged brain is much more susceptible to the impact of inflammation compared to the young brain [8]. Aging results in an increase in inflammatory mediators (i.e., cytokines, chemokines, etc.) liberated by microglia and astrocytes in the brain and an infiltration of peripheral inflammatory cells that contribute to the cognitive and physical deficits [9]. These changes in OS and inflammation result in alteration in membrane structure, modifications in DNA, RNA, protein and lipids that contribute to age-related adjustments in brain vulnerability [10,11].
Because of the potential impact of OS and inflammatory vulnerability in aging, research has focused on approaches to ameliorate these effects and attenuate the deficits in cognitive function [12–14]. A number of studies have concentrated on the use of exogenous and synthetic anti-oxidants and anti-inflammatory agents, including plant-derived compounds that protect against stress-related phenomenon [15,16]. Over the years, a number of compounds have been isolated from various plant species that protect the plants from OS and inflammation [17,18]. Studies have identified polyphenols, carotenoids, flavonoids, and sulfur compounds as natural anti-oxidants and anti-inflammatory agents that protect the plant from susceptibility to harmful conditions [19]. Finally, studies have been investigating the role of these compounds in reducing the age-related sensitivity of the brain to OS and inflammation [20].
Our previous studies have shown that dietary supplementation with GrandFusion® (GF), which is a nutritional supplement that contains the natural ingredients from whole fruits and vegetables demonstrated anti-oxidant, anti-inflammatory, and neuroprotective properties [21]. We showed that when mice were fed a diet supplemented GF for two months followed by cerebral ischemia were protected from infarct damage in the brain and demonstrated improved behavioral outcomes. In addition, the diet attenuated inflammatory processes by reducing microglial and astrocytic activation. The study implicated GF in the neuroprotective effects against the impact of cerebral ischemia and reperfusion injury.
Previous studies have shown that extracts from blueberries or strawberries can attenuate age-related motor and cognitive deficits in aged rat [22]. When young rats were fed a diet enriched in AIN-93 supplemented with spinach or strawberry extracts, the animals did not show age-related changes in cognitive function when compared to control rats [23]. In addition, older rats fed diets enriched in blueberry or strawberry extracts over an 8 week period, resulted in a reversal of cognitive and behavioral deficits [24]. Other studies using NT-020 (enriched in polyphenols from blueberry extracts) reduced inflammation, OS and improved Nrf-2 signaling [25]. These studies suggest that extracts from fruits and vegetables contain compounds that not only reduce age-related inflammation and OS, but help to enhance neurogenesis in the brain that may contribute to improved outcomes [25].
The present study was carried out to further assess the behavioral and biochemical benefits of a diet rich in vegetables and fruits. Our results demonstrate that these diets show a positive influence on behavioral outcomes associated with the aging phenomenon [21]. In addition, advantageous changes in biochemical parameters such as inflammation, reactive oxygen species (ROS) and cellular signaling are present in animals fed a diet enriched in fruits and vegetables. Finally, we have demonstrated that the supplemented diets enhanced neurogenesis in the brain of aged animals. These data suggest that these diets can affect the pronounced changes seen in aging to slow the process and improve outcome.
Results
Overall appearance and weights of animals in the study
Mean body weights of all groups over the 16 weeks of experimental period were illustrated in Figure 1A. The control group showed a steady weight over the time course of the study. The rats on the GrandFusion® (GF) diets, showed a slight decrease in weight with the diets that was significant. The calorie content between the different diets was the same and this was not the result of the body changes. All four groups are present, there is significant overlap and appear together. The average food intake was also determined in the rats over the course of the 16 weeks (Figure 1B). In that time, there were no significant differences in food consumption between the control rats and the rats on the GF diets.
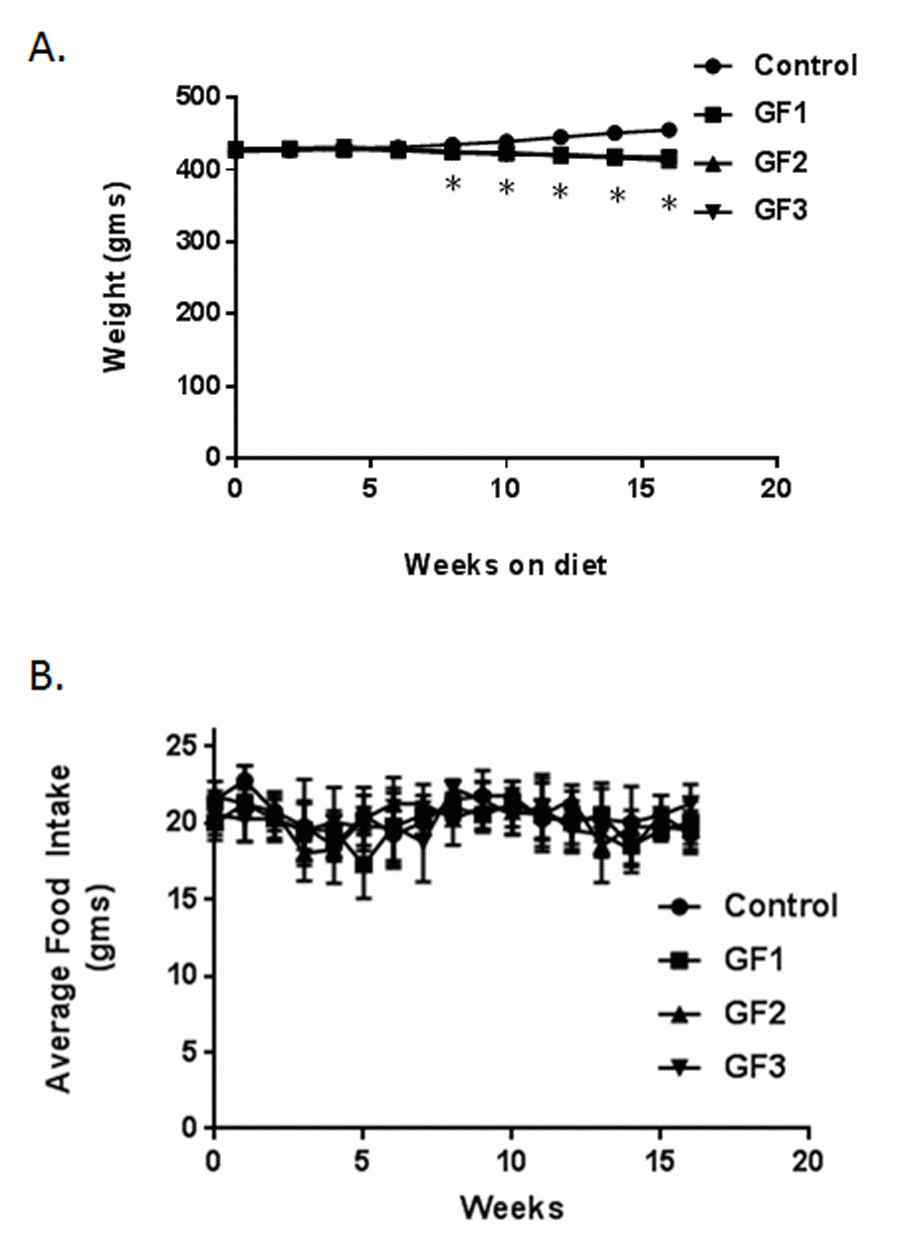
Figure 1. Animal weights and food intake. Animals were fed a standard diet of NIH-31 or diets enriched in GrandFusion (NF-216,GF1; NF-316, GF2; or NF-416, GF3). (A) Animals were weighed starting on month 18 and for 16 weeks after starting the diet. (B) Average food intake was determined every week after start of the diets. Values are means, with their standard errors represented by vertical bars. N = 20 per group. *P<0.001.
Improvement in psychomotor test in aged rats
The aged rats that consumed the GF diets performed significantly better on the behavioral tests than did control aged rats; and all of the GF diets resulted in similar effectiveness for all the behavioral endpoints. In addition, both male and female aged rat groups showed improved performance on tests of psychomotor performance compared with control aged rats (regular diet). On the rod walk (Fig. 2A), latency to fall was significantly higher in the GF groups than in the control group (P < 0.05) the aged males showed a slightly better improvement in the rod walk compared to the females but this was not significantly different. In the rotarod test (Fig. 2B) latency to fall was significantly better in the GF groups than in the control group (P ≤ 0.05). There was no difference between the aged male and aged female rats. Finally, group differences were seen on the large plank test (Fig. 2C). The GF groups showed significant improvement when compared with the control group on the large plank assay. Therefore, GF supplementation improved performance on motor tests, which rely on balance, coordination, and strength.
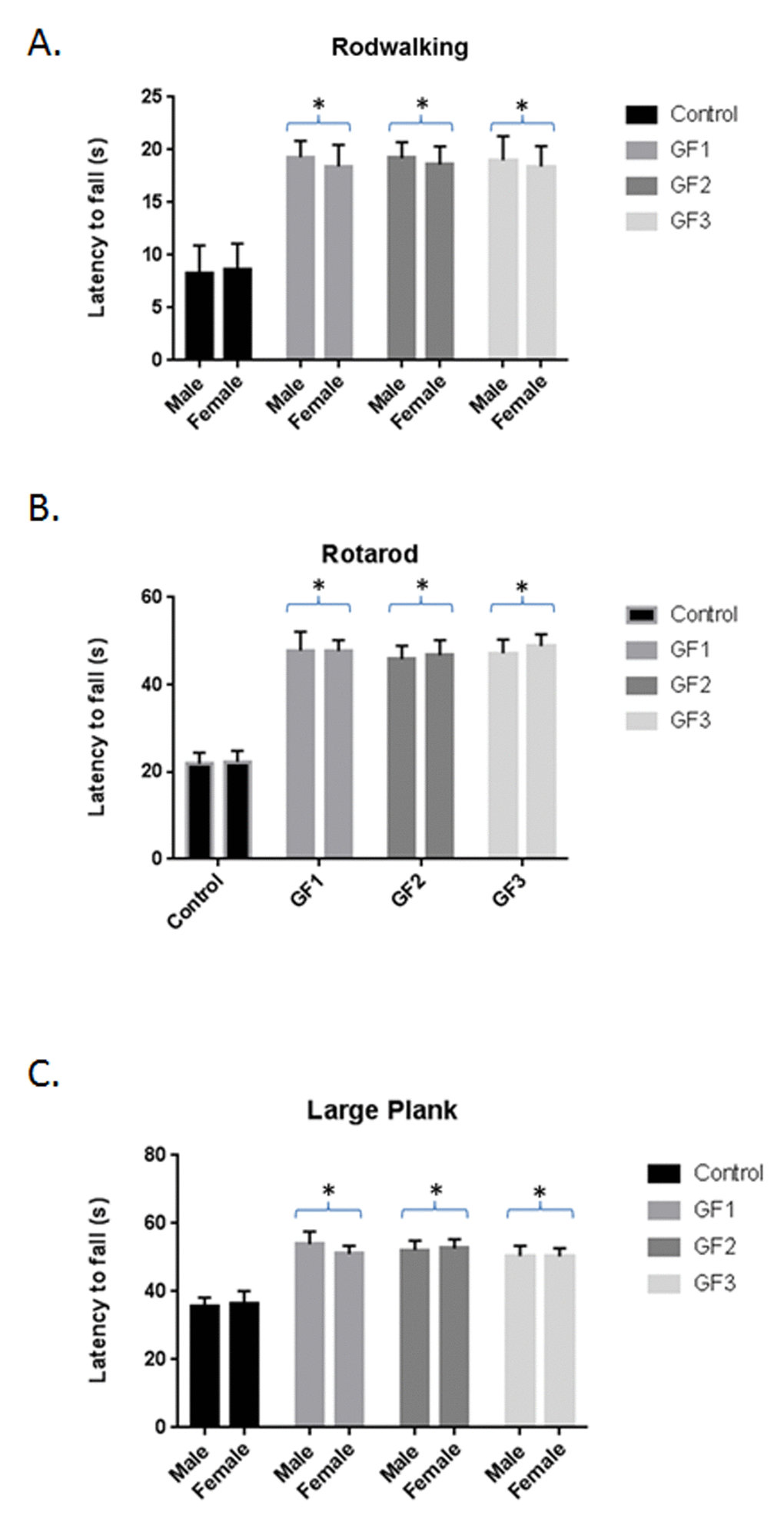
Figure 2. Behavioral testing following diet feeding. Performance (latency to fall, in seconds) on rod walk (A), rotarod (B) and large plank (C) for the various diet groups. Male and female 18 month old Fisher 344 rats were fed chow with and without GF diets and examined for various behavioral tests. Values are means, with their standard errors represented by vertical bars. N = 10 per group. *P<0.001.
Enhanced cognitive testing in aged rats
When examining cognitive performance, the GF groups showed improved performance over the control group in the Morris water maze test (Fig. 3A and B). We performed t-tests between the trial latencies and distances for each group to determine whether the different groups significantly improved their performance, which would demonstrate improved working memory. The GF groups showed significant (P < 0.05) differences in latencies (Fig. 3A) and distances (Fig. 3B) compared to the control group. These differences were not due to learning ability or swim speed because there were no differences between groups on these parameter (data not shown). There were no differences between the male and female aged rats in the GF groups and subsequent data analysis combined the two groups.
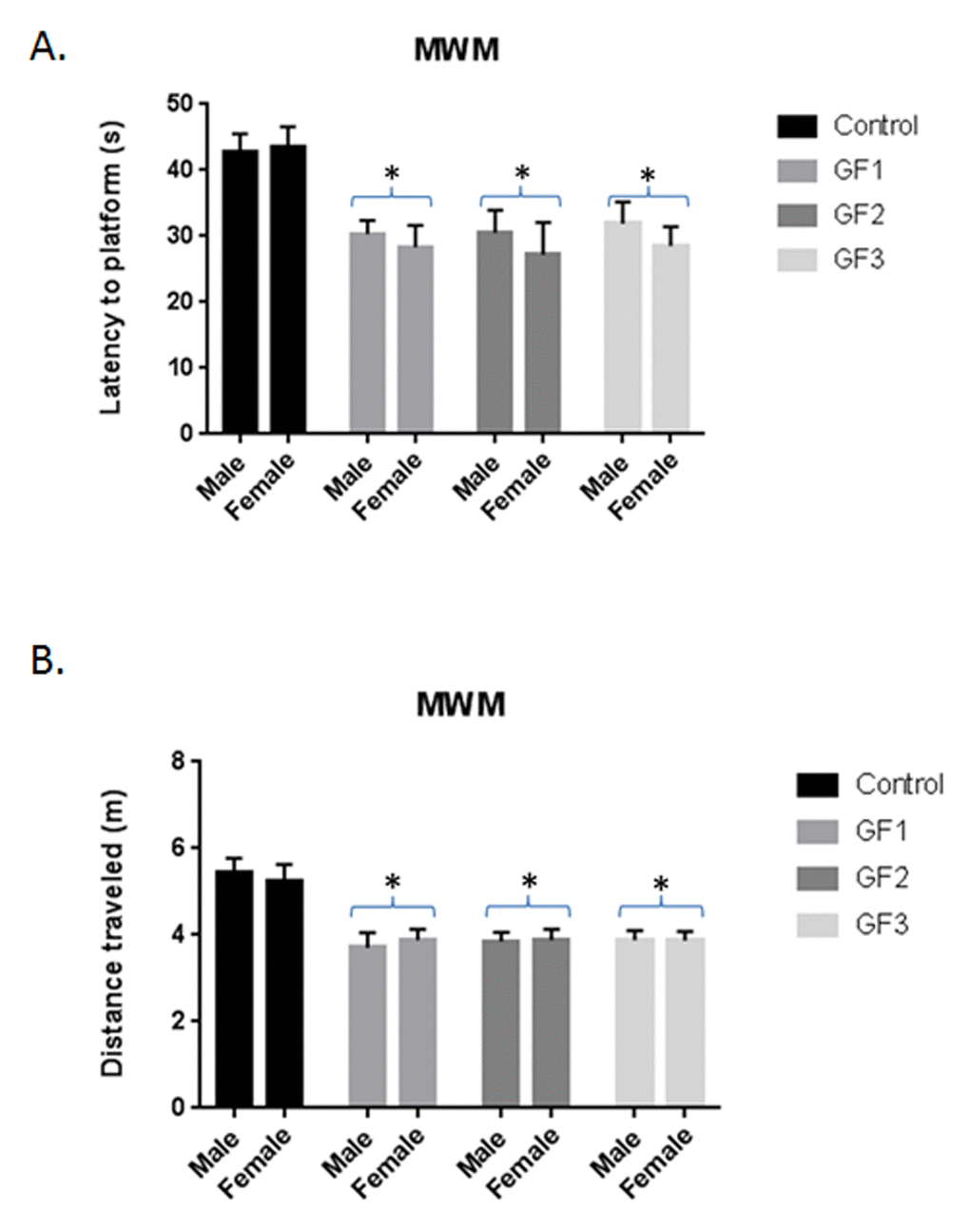
Figure 3. Morris water maze testing. Morris water maze performance assessed as latency in seconds (A) and distance (B) to find the hidden platform over days 4 and 5 of testing for animals in control, GF1, GF2 and GF3 diet groups. After 16 weeks on the GF diets, the animals were subjected to Morris water maze. The final trial day is presented as seconds to find the hidden platform. Both males and females were used in the studies. Values are means, with their standard errors represented by vertical bars. N = 10 per group. *P<0.001.
Enhanced neurogenesis
Differences were seen in cell survival among the diet groups compared to the control group (P < 0.05; Fig. 4A), with rats in the GF groups showing a significant increase in the number of cells surviving in the dentate gyrus of the hippocampus compared with the control diet group (P < 0.05). In addition, the rats in the GF groups also showed higher numbers of proliferating precursor cells (Fig. 4B), these differences did reach significance (P < 0.05). In both measures, rats in both the male and female GF diet groups were significantly different from the control group (Fig. 4A and B). Correlations between neurogenesis and cognitive performance revealed that, as the number of proliferating cells increased, the mean difference in latency was shorter in the GF groups (Fig. 3 and 4). The difference in the score is a measure of working memory, and the score implies that the rats were able to find the platform more quickly, remembering where it was on earlier trials. All of the GF groups showed significant correlations between proliferation and working memory performance (P < 0.05). In addition, the cognitive performance did significantly correlate with surviving cells, and all of the motor tests showed a positive correlation with neurogenesis.
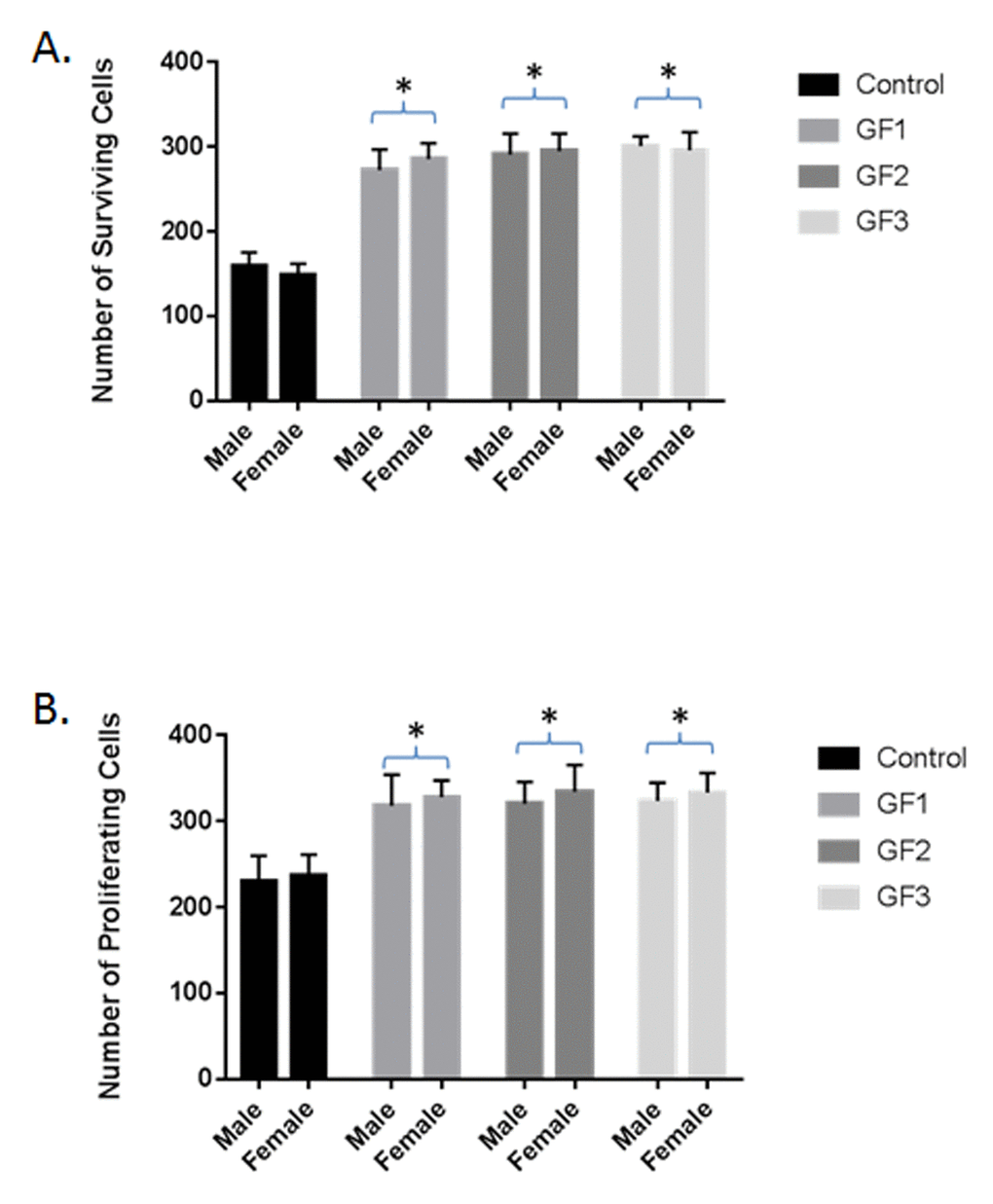
Figure 4. Neuroprotection and neurogenesis. The number of surviving (A) and proliferating (B) precursor cells in the dentate gyrus of the hippocampus of rats in the control, GF1, GF2 or GF3 diet groups. Values are means, with their standard errors represented by vertical bars. N = 20 per group. *P<0.05.
Reduced oxidative stress in diet treated rats
Supplementation of the GF diets demonstrated changes in malondialdehyde (MDA) or thiobarbituric acid reactive substances (TBARS) (Fig. 5A). The diets showed a significant effect on MDA/TBARS levels in the brain in control aged and GF supplemented animals. Significant differences were present in the brain with all of the diets demonstrating an effect in reducing lipid peroxidation products. The results showed that the control rats had greater values when compared with the GF groups. The diets showed ∼75% reduction in TBARS. DCF measurements in the brain of aged animals treated with GF diets lead to a reduction in free radical levels compared with control aged animals (Fig. 5C). The GF diets showed improvement with approximately ~70% reduction in free radicals. The impact of the diets on superoxide dismutase (SOD) activity was detected in different groups. Aged rats demonstrated a significant decrease in SOD activity in the brain at the end of the study (Fig. 5B). In the GF supplemented aged rats, there was an increase in the SOD activity compared with the control rats. The diets caused an increase in SOD activity at the end of the experiment. These results suggest that aging exacerbates oxidative stress in the brain and the diets where capable of suppressing the oxidative stress and enhancing anti-oxidant activity.
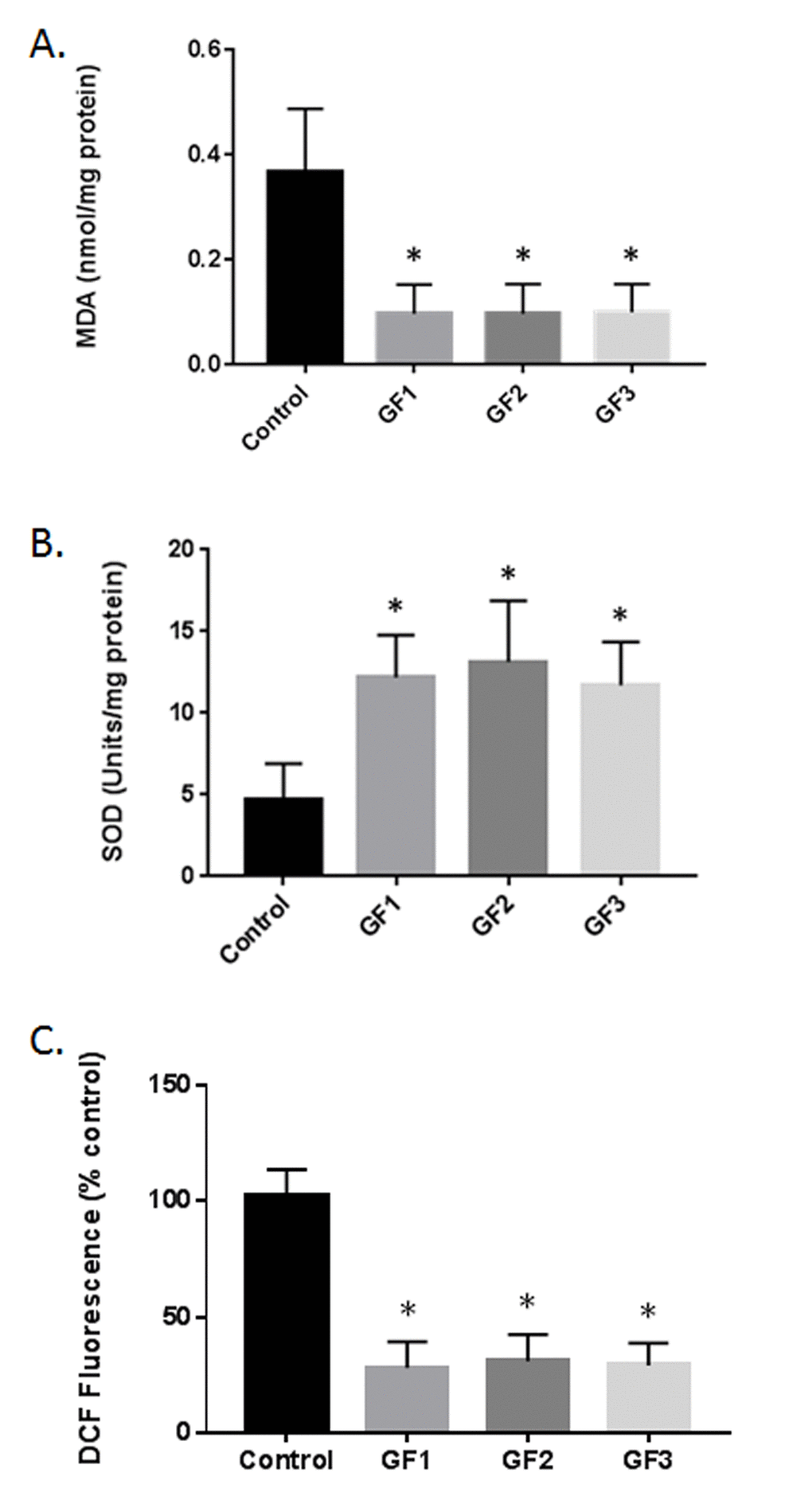
Figure 5. Reduction in oxidative stress following GF diets. (A) MDA levels in the brains of aged rats subjected to control or GF diets. (B) SOD activity in the brains of aged rats following feeding control diets or diets enriched with GF formulas. (C) DCF assay in the brains of aged rats following feeding control diets or diets enriched with GF formulas. N = 20 per group. P<0.001.
Reduced inflammatory markers in diet fed aged rats
Quantification of inflammatory markers (IL-1β, TNF-α and IL-6) by ELISA in the brain showed that aged animals fed the GF diets exhibited significantly lower overall cytokine activity than the control aged rats (Fig. 6A, P < 0.01). Consumption of a GF diets in the aged rats was associated with decreased expression of IL-1β, TNF-α and IL-6. Interestingly, all of the GF diet supplementation showed a significant attenuated of cytokine expression. Additionally, the brains were analyzed for the expression of brain derived neurotrophic factor (BDNF) and the GF diets showed a significant increase in BDNF levels compared to the control diet rats (Fig. 6B). Overall, there is a significant decrease in inflammatory markers with an increase in trophic factor expression.
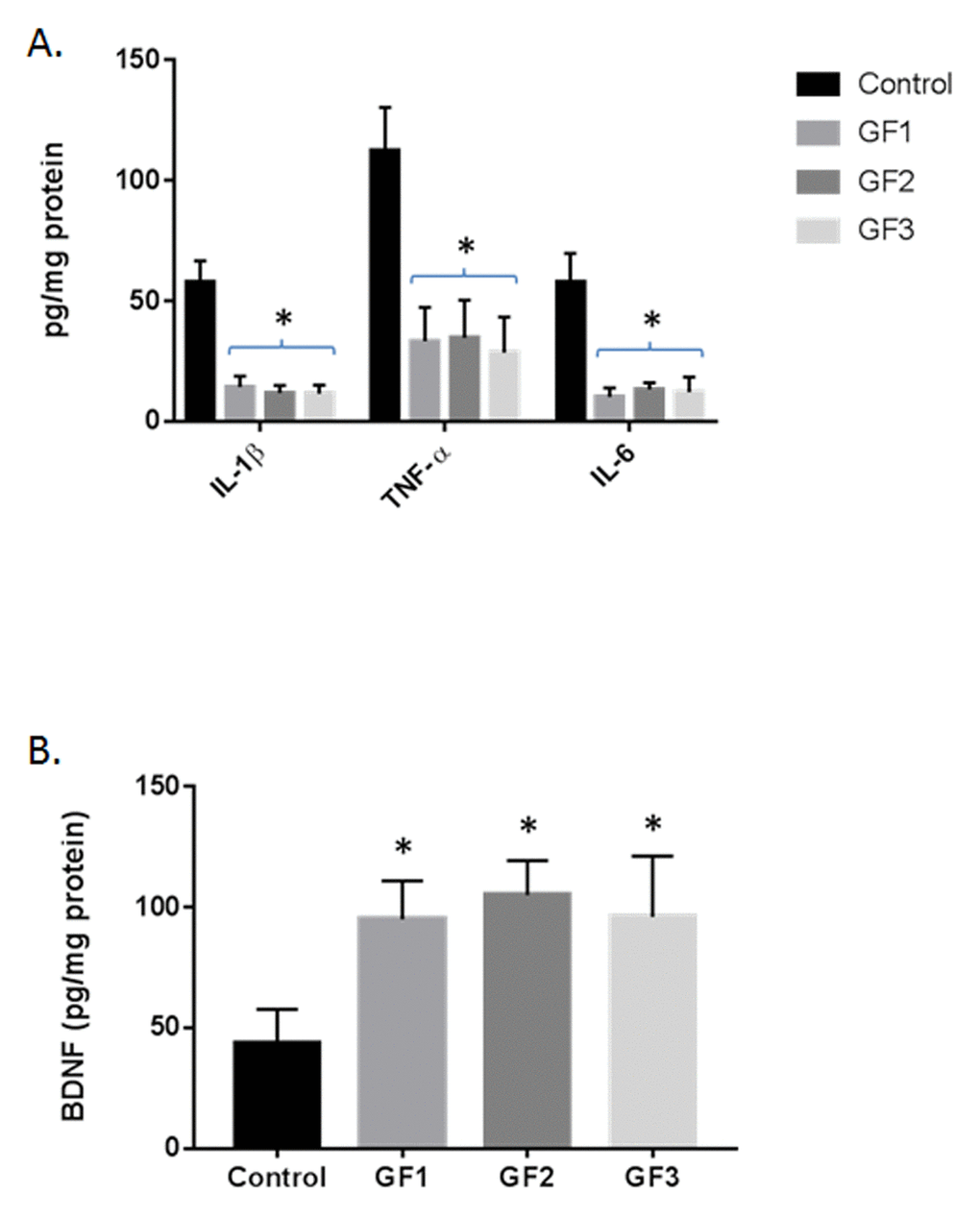
Figure 6. Cytokine and BDNF levels in the brains of aged rats. (A) IL-1β, TNF−α and IL-6 were measured in the brains of aged rats subjected to control or GF diets. (B) BDNF was measured in the brains of aged rats subjected to control or GF diets. N = 20 per group. P<0.001.
Microglial and astrocyte activation following GF diets
As seen in Fig. 7B-C, both microglial (iba-1) and glial fibrillary acidic protein (GFAP) were increased in the GF supplemented groups compared to the control group. In Figure 7A, a representative western blot for iba-1 immunoreactivity is shown. At 12 week supplementation with GF diets, all the aged rats exhibited lower iba-1 and GFAP expression compared to that seen in aged control rats as evidenced by semi-quantitative densitometry measurements. In the brains from GF supplemented animals, iba-1 expression was normalized to 100% while the GF treated animals decreased to around 23 +/- 8% post treatment (Fig. 7A and B). In addition, GFAP expression was 100% while the GF treated animals decreased to 14 ± 7% at 12 weeks post-supplementation, representing significantly reduced astrogliosis compared to GFAP in aged rats on a control diet (Fig. 7C).
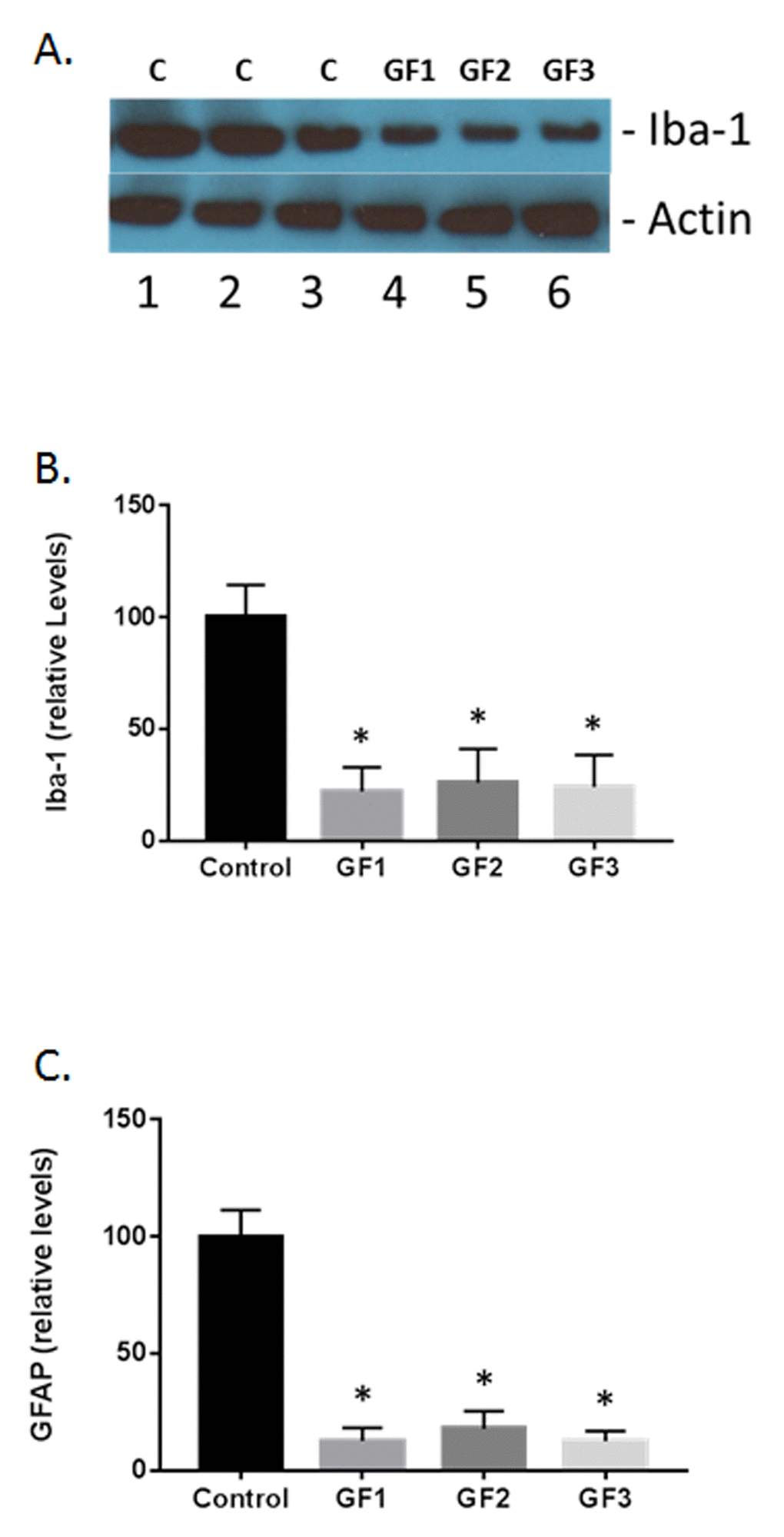
Figure 7. Measurement of glial activation in the aged brain and brains of animals fed GF diets. (A) Representative western blot of iba-1 levels in the brains of aged rats subjected to control or GF diets. C, control; GF1, GF2, GF3, GF diets. Actin is used as a control for loading. (B) Quantified levels of iba-1 in the brains of aged rats following feeding control diets or diets enriched with GF formulas. (C) Quantified levels of iba-1 in the brains of aged rats following feeding control diets or diets enriched with GF formulas. N = 20 per group. P<0.001.
Nrf2 expression in aged rats following nutraceutical treatment
A number of studies have shown that polyphenolic chemicals are capable of activating the adaptive stress response pathways in various cells [26]. Of these responses, Nrf2 has been shown to play a critical role in the stress pathway by attenuation of the inflammatory process [27]. Western blot analysis of the Nrf2 in the brain of aged animals in the presence of GF diets or control diet was examined. As seen in Figure 8, the GF diets significantly increased Nrf2 expression in the brain. Quantification of the levels of Nrf2 showed that the GF diets increased Nrf2 by 300-400% over the 16 weeks of treatment (Fig. 8B). As demonstrated in Figure 7, both iba-1 and GFAP as well as other inflammatory cytokines were decreased in the presence of the GF diets suggesting an impact for Nrf2 on stress in aging. Decreases in MDA and DCF fluorescence and an increase in SOD are also indicators of Nrf2 activity. These data support the assumption that the activity is derived from the activation of antioxidant response element pathways from most cell types in the brain.
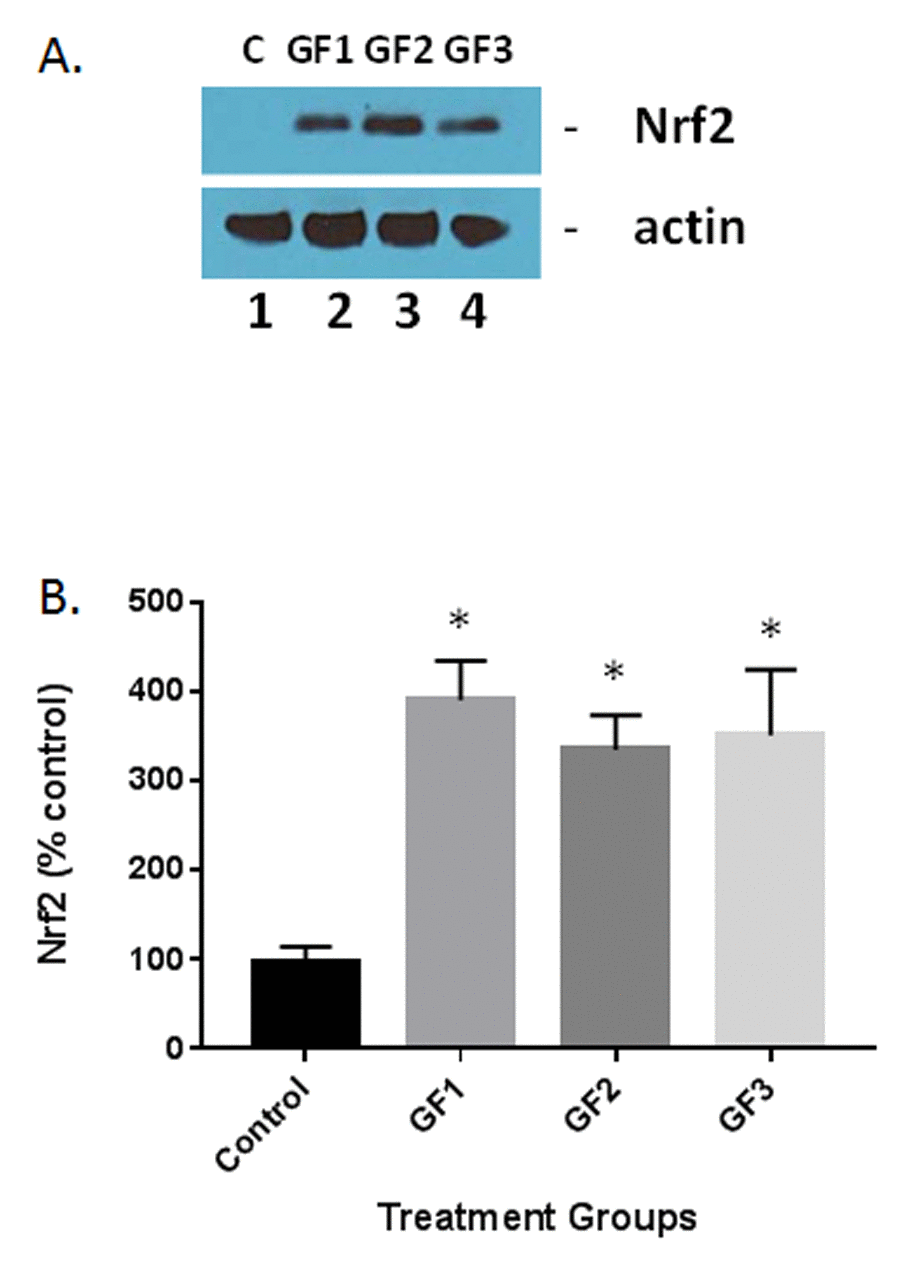
Figure 8. Measurement of Nrf2 expression in the aged brain and brains of animals fed GF diets. (A) Representative western blot of Nrf2 levels in the brains of aged rats subjected to control or GF diets. C, control; GF1, GF2, GF3, GF diets. Actin is used as a control for loading. (B) Quantified levels of Nrf2 in the brains of aged rats following feeding control diets or diets enriched with GF formulas. N = 20 per group. P<0.001.
Discussion
The current study has shown that a diet enriched with GrandFusion® can reduce inflammation and oxidative stress, neural stem cell proliferation and improved cognitive function in the aged rat brain. In addition, the treatment showed a significant increase in stem cell survival and activation of BDNF and Nrf2 to stimulate the expression of antioxidant genes. No significant differences were detected between male and female animals in the study. The overall premise is that aging exacerbates inflammation and that impacts the neuronal cells in the brain to alter their function and to stimulate gliosis that results in cognitive impairment.
The processes underlying age-related cognitive deficits persist to be fully depicted, it is clear that inflammation and oxidative stress play a significant role [28,29]. During the aging process, the CNS endogenous anti-inflammatory defenses deteriorate, causing the brain to be vulnerable to inflammatory insults [30–32]. The brain has limited regenerative capacity and various insults lead to aggregate cellular impairment that compromises normal neuronal and glial function. Mitochondrial dysfunction, lipid peroxidation, protein oxidation and altered autophagy are only some of the mechanisms responsible for intracellular damaged protein/lipid complexes, protein oligomers/aggregates, and cellular organelles [33]. These processes are well-regulated whose disruption leads to neuronal dystrophy and death, and are potential mechanisms of the aging process and the development of neurodegenerative diseases [34,35]. Neuroinflammation has been identified as a cause of neuronal dysregulation in aged rats [36]. Therefore, a potentially valuable approach to combat age-related deficits in behavior is to alter inflammation.
In the realm of aging, changes such as increased oxidative stress, inflammation characterized by reactive astrocytes and activated microglia and altered neurogenesis characterized by atypical migration of newly born neurons have been determined [37–41]. Memory and mood impairments in the aging are been attributed to the decline in neurogenesis and loss of neurons in the hippocampus [42,43]. Anti-aging therapies (AAT) can slow the aging process in most instances but cannot satisfactorily suppress age-induced detrimental changes described above [44–46]. Since these changes contribute to the aging process, anti-aging therapies have mostly failed to prevent the evolution of aging into chronic aging. Therefore, the best approach for aging should be capable of limiting neuronal loss, oxidative stress, inflammation and abnormal neurogenesis [47–50]. In this context, complexes and drugs having neuroprotective and/or anti-aging properties are excellent for preventing age-induced chronic neuronal dysfunction typified by aging, cognitive and mood impairments.
Other potential mechanisms for the impact of aging on cognition and memory may involve activation of cell signaling pathways such as p38 mitogen-activated protein kinase (MAPK), c-jun NH2-terminal kinase (JNK), caspase 1, and NFkB [51–56]. These kinases and factors occur within, and between, multiple signaling cascades that play a role in inflammation-associated cognitive deficits [57]. Interestingly, these pathways also interconnect with other pathways that are used by the neurotrophic factors, specifically BDNF. BDNF plays a critical role in the survival and development of certain populations of neurons [58]. In addition, BDNF can be neuroprotective, through the mitigation of damaging effects of a number of insults [59]. Finally, BDNF can play an important role in synaptic plasticity and cognition similar to inflammatory mechanisms associated with consolidation of hippocampus-dependent memory [60–62].
Studies from our group and others have implicated protection from aging and neurological disorders by increasing the dietary intake of fruits and vegetables [21–25,63]. Diets enriched in polyphenolics, which are high in antioxidant and anti-inflammatory compounds, such as blueberries, strawberries and spinach, can slow or even prevent the changes in neurochemical and behavioral parameters that characterize the aged brain [23,64–70]. We have demonstrated in several studies that diets supplemented with GrandFusion®, a mixture of fruits and vegetables, highly enriched in vitamins were able to limit the extent of cerebral ischemia injury and reverse several parameters of stroke, such as inflammation and oxidative stress and behavioral changes [21]. Finally, in addition to having anti-oxidant and anti-inflammatory properties, diets containing vegetables and fruits may have direct and indirect effects on cell signaling to augment neuronal communication, the ability to stimulate neurogenesis, enhancement of neurotrophic factor expression and the diminution of NF-κB [71–78]. Neurogenesis occurs in a number of locations including the dentate gyrus [68,69,72,77]. These cells migrate to areas of injury due to trauma or aging that helps to enhance neuronal function embracing cognition and motor function [72,77]. Therefore, neurogenesis in the dentate gyrus may lead to improved outcomes.
In summary, over the years, research has shown that changes in inflammation, oxidative stress and neurogenesis are involved in the aging process and therapeutics that alters this process can improve outcomes. The process of aging results in the elaboration of inflammatory cytokines and chemokines both present in the circulation as well as at the tissue level, which express their influence on neural stem cell population limiting their potential for regeneration and repair of the aging nervous system [23]. In addition, the presence of the inflammatory cytokines can affect neuronal function, induce microglial activation and astrocyte stimulation which can enhance the aging process [21]. The presence of GrandFusion® enriched in phyotchemicals and anti-inflammatory molecules help to attenuate inflammation and enhance neural stem cell survival and proliferation. This process helps to improve organ function, enrich the aging process and enhance cognitive ability in the animals.
Materials and Methods
Animals and diets
Male and female Fischer 344 rats were purchased from Charles River Labs at 17 months of age. Rats were used at eighteen months of age. The rats were maintained on a 12-hour light/dark schedule in standard cages with ad libitum food and water. After 4 weeks of acclimation, rats were weight-matched and randomly assigned to diet groups (n = 15/group for both male and female groups). Food intake was recorded weekly and body weights were recorded every other week. Rats were monitored for signs of injury or disease in accordance with the Medical University of South Carolina (MUSC) and Ralph H. Johnson VA Institutional Animal Care and Use Committee. Dr. Kindy moved to the University of South Florida after the studies were completed. Animals were fed for 4 months (old animals) on a normal diet or a normal diet with ~2% supplementation of the different materials NF-216 (GrandFusion – Fruit and Veggie #1 Blend, GF1), NF-316 (GrandFusion – Fruit #2 Blend, GF2), NF-416 (GrandFusion – Vegetable #3 Blend, GF3) as described previously (21). The amount of supplementation was based on the recommended intake of the diet for human consumption. The supplementation was pulverized and incorporated into the pelleted diet. GrandFusion® supplements were prepared by NutriFusion, LLC (www.nutrifusion.com). Average food intake was 0.020 ± 0.1 kg/kg body weight/day; and the average consumption of diets was 0.0004 ± 0.00005 kg/kg body weight/day. A food intake assessment (over a 96-h period) was made during the 8th week of the experimental feeding. During the course of the study, no rats had to be removed from the study due to animal issues. All studies were approved that the Institutional Animal Care and Use Committee at the Medical University of South Carolina and the Veterans Affairs Medical Center.
Histological examination
For histological examination, the animals were anesthetized with an intraperitoneal injection of sodium pentobarbital (50 mg/kg) at the indicated times. The brains were transcardially perfused with 4°C, 10% phosphate buffered saline (PBS). The brains were removed and chilled for 15 min at −20°C before being placed in a Rodent Brain Matrix. Coronal sections (2-mm thickness) were prepared and placed in OCT medium and frozen [25]. 30 μm sections were prepared for immunohistochemical analysis.
ELISA analysis
For quantitative analysis of cytokines, an enzyme-linked immunosorbent assay (ELISA) was used to measure the levels of tumor necrosis factor-α, interleukin-1β, or interleukin-6 in brain tissue [79,80]. For BDNF assays, we used the ELISA kit provided by Abnova. Cytokines were extracted from rat brains as follows: frozen hemibrains were placed in Tissue Homogenization Buffer containing Protease Inhibitor Cocktail (PIC, Sigma, St Louis, MO, USA) 1:1,000 dilution immediately before use, and homogenized using polytron. Tissue sample suspensions were distributed in aliquots and snap frozen in liquid nitrogen for later measurements. ELISA kits were then used, according to manufacturer directions (Abnova, Taipei City, Taiwan).
Behavioral assessment
The battery of motor tests and the cognitive test selected for this study have been previously shown by our group to be age-sensitive [21]. For most of the measures, the decline in performance is observed as early as 12 to 15 months of age. Previous work has shown that an N of 15 per group will give us sufficient power to find statistically significant (P < .05) differences on tests of behavior. Power calculations show that, for the Morris water maze (MWM), a difference in means of 10 (e.g., 32 vs 22 seconds in latency to find the platform), when the SD is 10, would yield a power of 78%, while for the rod walking test, a difference in means of 4.5 (e.g., 8.1 vs 12.6 s in latency to fall), when the standard deviation is 4, would yield a power of 85%, using a sample size of 15.
Motor testing
After 2 months of dietary intervention, rats underwent a battery of motor tests to assess for (1) Balance: rats were placed on a large plank (38 mm; counterbalanced) and a narrow rod (26 mm) horizontally suspended 23 cm above a thick foam-core pad and latency to fall was recorded (max 60 seconds); and (2) Fine motor coordination and stamina: rats were placed on an accelerating rotarod (San Diego Instruments, San Diego, CA) consisting of a slowly accelerating (+2 rpm/30 s; 20 rpm max) rotating dowel (7 cm diameter) and latency to fall was recorded (max 300 seconds).
Cognitive testing
To assess spatial learning and memory, rats completed a working memory version of the MWM (wMWM) at weeks 9–10 of dietary intervention [81,82]. The test was given daily for 4 consecutive days, 2 sessions per day and 2 trials each session: a reference memory or acquisition trial (trial 1) and a working memory or retrieval trial (trial 2). During testing, rats were placed in a large water-filled pool (134 cm diameter) and allowed 120 s to escape onto a platform hidden 2 cm below the water surface. If the rat failed to escape within this time, it was guided to the platform. Once the rat reached the platform, it was allowed to remain there for 15 s (trial 1). The rat was then returned to its home cage for 10 minutes (inter-trial interval). Trial 2 used the same platform location and start position as trial 1. The platform was moved to one of four locations, chosen to frustrate a number of non-place learning strategies that rats may adopt, at the beginning of each session. wMWM files were analyzed with image tracking software (San Diego Instruments, La Jolla, CA).
Analysis of inflammation
Brain iba-1, GFAP, and actin (control) protein levels were determined at time of euthanasia. Animals were sham or treated with vehicle or supplemental diets. Relative levels of iba-1, GFAP, and actin in the supernatant fraction from the brain extract were determined by western blot analysis (polyclonal antibodies: iba-1 (sc-28530); GFAP, (sc-6170); β-actin (sc-130657); Santa Cruz Biotechnology, Santa Cruz, CA), as described previously [21]. For Nrf2 analysis, we used antibodies provided by Abcam (ab137550). Relative intensities of western blot bands were assessed by densitometry in triplicate for each sample. Densitometric analysis was done using IQTL software (GE Life Sciences, Piscataway, NJ). For protein studies, the entire lesional area was harvested for western blot analysis. In control or sham animals, a similar region was harvested.
Thiobarbituric acid reactive substances
MDA (malondialdehyde) was evaluated by thiobarbituric acid reactive substances (TBARS) test [83]. Aliquots of samples were incubated with 10% trichloroacetic acid and 0.67% thiobarbituric acid. The mixture was heated (30 minutes) on a boiling water bath. Afterwards, n-butanol was added and the mixture was centrifuged (1,000 x g for 10 minutes). The organic phase was collected to measure fluorescence at excitation and emission wavelengths of 515 and 553 nm, respectively. 1,1,3,3-tetramethoxypropane, which is converted to MDA (malondialdehyde), was used as standard. Results are expressed as pmol MDA/mg protein and reported as percentage of control.
Free radical levels
To assess the free radicals content, we used 20-70-dichlorofluorescein diacetate (DCFH-DA) as a probe [84]. A sample aliquot was incubated with DCFH-DA (100 mM) at 37°C for 30 minutes; the reaction was terminated by chilling the reaction mixture in ice. The formation of the oxidized fluorescent derivative (DCF) was monitored at excitation and emission wavelengths of 488 and 525 nm, respectively, using a fluorescence spectrophotometer (Hitachi F-2000, Hitachi High-Tech, Northridge, CA, USA). The free radicals content was quantified using a DCF standard curve and the results were expressed as pmol of DCF formed/mg protein. All procedures were performed in the dark and blanks containing DCFH-DA (no homogenate) were processed for measurement of autofluorescence.
SOD assay
For the analysis of superoxide dismutase (SOD) activity, tissues were homogenized on ice in cold 25 mM HEPES buffer (pH 7.4) containing 250 mM sucrose and 1 mM EDTA and then centrifuged at 1500× g for 5 min at 4 °C. The activity of SOD was measured in supernatants using a superoxide dismutase assay kit following the manufacturer's instructions and expressed as units per mg of protein (Cayman Chemicals, Ann Arbor, MI). One unit is defined as the amount of enzyme needed to produce 50% dismutation of the superoxide radical.
Protein determination
Protein was measured by the Coomassie blue method using bovine serum albumin as standard.
Statistical analysis
The results were expressed as the mean±standard deviation (SD). The statistical significance of the results in the brain by ELISA, behavioral studies, physiological and histological data were analyzed using a T-test or one-way analysis of variance (ANOVA) followed by Fisher's post hoc test. Repeated-measures ANOVA were computed on the monitoring data and the significance of the difference among groups were evaluated by Fisher's post hoc test.
Abbreviations
GF: GrandFusion; OS: oxidative stress; ROS: reactive oxygen species; MDA: malondialdehyde; TBARS: thiobarbituric acid reactive species; SOD: superoxide dismutase; BDNF: brain derived neurotrophic factor; GFAP: glial fibrillary acidic protein; AAT: anti-aging therapies.
Author Contributions
J.Y, and M.S.K. provided the study concept and the design. J.Y., H.Z., S.T. and M.S.K. acquired the data. S.P. and M.S.K. provided the analysis and interpretation of the data. M.S.K. drafted the manuscript. All authors critically reviewed the manuscript for important intellectual content. J.Y. and M.S.K. supervised the study.
Acknowledgements
The authors wish to acknowledge NutriFusion, LLC for providing the GrandFusion® diets for the studies. We thank Mr. William Grand for reviewing the manuscript prior to submission. GrandFusion® is a patent pending product of NutriFusion, LLC.
Conflicts of Interest
Dr. Stephen Perry is a technical and science consultant for NutriFusion, LLC.
Funding
This work was partially supported by grants from the National Institutes of Health (R01 ES016774-01, R21AG043718, 1P20GM109091, 2P20GM103444, and 5P30GM103342), VA Merit Award, a grant from the National Science Foundation (IIP-0903795), an AHA SFRN grant and VA Merit Review (M.S.K.). Dr Kindy is a Senior Research Career Scientist in the VA.
References
- 1. Schafer MJ, Miller JD, LeBrasseur NK. Cellular senescence: implications for metabolic disease. Mol Cell Endocrinol. 2016S0303-7207(16)30354-9; Epub ahead of print. [PubMed]
- 2. Calçada D, Vianello D, Giampieri E, Sala C, Castellani G, de Graaf A, Kremer B, van Ommen B, Feskens E, Santoro A, Franceschi C, Bouwman J. The role of low-grade inflammation and metabolic flexibility in aging and nutritional modulation thereof: a systems biology approach. Mech Ageing Dev. 2014; 136-137:138–47. https://doi.org/10.1016/j.mad.2014.01.004 [PubMed]
- 3. Cevenini E, Caruso C, Candore G, Capri M, Nuzzo D, Duro G, Rizzo C, Colonna-Romano G, Lio D, Di Carlo D, Palmas MG, Scurti M, Pini E, et al. Age-related inflammation: the contribution of different organs, tissues and systems. How to face it for therapeutic approaches. Curr Pharm Des. 2010; 16:609–18. https://doi.org/10.2174/138161210790883840 [PubMed]
- 4. Bourassa K, Sbarra DA. Body mass and cognitive decline are indirectly associated via inflammation among aging adults. Brain Behav Immun. 2017; 60:63–70. https://doi.org/10.1016/j.bbi.2016.09.023 [PubMed]
- 5. Norden DM, Muccigrosso MM, Godbout JP. Microglial priming and enhanced reactivity to secondary insult in aging, and traumatic CNS injury, and neurodegenerative disease. Neuropharmacology. 2015; 96:29–41. https://doi.org/10.1016/j.neuropharm.2014.10.028 [PubMed]
- 6. Wang J, Lei H, Hou J, Liu J. Involvement of oxidative stress in SAMP10 mice with age-related neurodegeneration. Neurol Sci. 2015; 36:743–50. https://doi.org/10.1007/s10072-014-2029-5 [PubMed]
- 7. Paul A, Belton A, Nag S, Martin I, Grotewiel MS, Duttaroy A. Reduced mitochondrial SOD displays mortality characteristics reminiscent of natural aging. Mech Ageing Dev. 2007; 128:706–16. https://doi.org/10.1016/j.mad.2007.10.013 [PubMed]
- 8. del Zoppo GJ. Aging and the neurovascular unit. Ann N Y Acad Sci. 2012; 1268:127–33. https://doi.org/10.1111/j.1749-6632.2012.06686.x [PubMed]
- 9. Kohman RA. Aging microglia: relevance to cognition and neural plasticity. Methods Mol Biol. 2012; 934:193–218. https://doi.org/10.1007/978-1-62703-071-7_11 [PubMed]
- 10. DiSabato DJ, Quan N, Godbout JP. Neuroinflammation: the devil is in the details. J Neurochem. 2016 (Suppl 2); 139:136–53. https://doi.org/10.1111/jnc.13607 [PubMed]
- 11. Cornejo F, von Bernhardi R. Age-dependent changes in the activation and regulation of microglia. Adv Exp Med Biol. 2016; 949:205–26. https://doi.org/10.1007/978-3-319-40764-7_10 [PubMed]
- 12. Campuzano O, Castillo-Ruiz MM, Acarin L, Gonzalez B, Castellano B. Decreased myeloperoxidase expressing cells in the aged rat brain after excitotoxic damage. Exp Gerontol. 2011; 46:723–30. https://doi.org/10.1016/j.exger.2011.05.003 [PubMed]
- 13. Calabrese V, Cornelius C, Mancuso C, Lentile R, Stella AM, Butterfield DA. Redox homeostasis and cellular stress response in aging and neurodegeneration. Methods Mol Biol. 2010; 610:285–308. https://doi.org/10.1007/978-1-60327-029-8_17 [PubMed]
- 14. Chakrabarti S, Mohanakumar KP. Aging and neurodegeneration: a tangle of models and mechanisms. Aging Dis. 2016; 7:111–13. https://doi.org/10.14336/AD.2016.0312 [PubMed]
- 15. Pusic KM, Pusic AD, Kraig RP. Environmental enrichment stimulates immune cell secretion of exosomes that promote CNS myelination and may regulate inflammation. Cell Mol Neurobiol. 2016; 36:313–25. https://doi.org/10.1007/s10571-015-0269-4 [PubMed]
- 16. Jia Z, Xue R, Ma S, Xu J, Guo S, Li S, Zhang E, Wang J, Yang J. Erythropoietin attenuates the memory deficits in aging rats by rescuing the oxidative stress and inflammation and promoting BDNF releasing. Mol Neurobiol. 2016; 53:5664–70. https://doi.org/10.1007/s12035-015-9438-1 [PubMed]
- 17. Ricordi C, Garcia-Contreras M, Farnetti S. Diet and inflammation: possible effects on immunity, chronic diseases, and life span. J Am Coll Nutr. 2015 (Suppl 1); 34:10–13. https://doi.org/10.1080/07315724.2015.1080101 [PubMed]
- 18. Lemon JA, Aksenov V, Samigullina R, Aksenov S, Rodgers WH, Rollo CD, Boreham DR. A multi-ingredient dietary supplement abolishes large-scale brain cell loss, improves sensory function, and prevents neuronal atrophy in aging mice. Environ Mol Mutagen. 2016; 57:382–404. https://doi.org/10.1002/em.22019 [PubMed]
- 19. Pribis P, Shukitt-Hale B. Cognition: the new frontier for nuts and berries. Am J Clin Nutr. 2014 (Suppl 1); 100:347S–52S. https://doi.org/10.3945/ajcn.113.071506 [PubMed]
- 20. Williams RJ, Mohanakumar KP, Beart PM. Neuro-nutraceuticals: the path to brain health via nourishment is not so distant. Neurochem Int. 2015; 89:1–6. https://doi.org/10.1016/j.neuint.2015.08.012 [PubMed]
- 21. Yu J, Zhu H, Gattoni-Celli S, Taheri S, Kindy MS. Dietary supplementation of GrandFusion(®) mitigates cerebral ischemia-induced neuronal damage and attenuates inflammation. Nutr Neurosci. 2016; 19:290–300. https://doi.org/10.1179/1476830515Y.0000000021 [PubMed]
- 22. Flowers A, Lee JY, Acosta S, Hudson C, Small B, Sanberg CD, Bickford PC, Grimmig B. NT-020 treatment reduces inflammation and augments Nrf-2 and Wnt signaling in aged rats. J Neuroinflammation. 2015; 12:174. https://doi.org/10.1186/s12974-015-0395-4 [PubMed]
- 23. Acosta S, Jernberg J, Sanberg CD, Sanberg PR, Small BJ, Gemma C, Bickford PC. NT-020, a natural therapeutic approach to optimize spatial memory performance and increase neural progenitor cell proliferation and decrease inflammation in the aged rat. Rejuvenation Res. 2010; 13:581–88. https://doi.org/10.1089/rej.2009.1011 [PubMed]
- 24. Bickford PC, Gould T, Briederick L, Chadman K, Pollock A, Young D, Shukitt-Hale B, Joseph J. Antioxidant-rich diets improve cerebellar physiology and motor learning in aged rats. Brain Res. 2000; 866:211–17. https://doi.org/10.1016/S0006-8993(00)02280-0 [PubMed]
- 25. Bickford PC, Shukitt-Hale B, Joseph J. Effects of aging on cerebellar noradrenergic function and motor learning: nutritional interventions. Mech Ageing Dev. 1999; 111:141–54. https://doi.org/10.1016/S0047-6374(99)00063-9 [PubMed]
- 26. Ajami M, Pazoki-Toroudi H, Amani H, Nabavi SF, Braidy N, Vacca RA, Atanasov AG, Mocan A, Nabavi SM. Therapeutic role of sirtuins in neurodegenerative disease and their modulation by polyphenols. Neurosci Biobehav Rev. 2017; 73:39–47. https://doi.org/10.1016/j.neubiorev.2016.11.022 [PubMed]
- 27. McNeilly AD, Gallagher JR, Dinkova-Kostova AT, Hayes JD, Sharkey J, Ashford ML, McCrimmon RJ. Nrf2-mediated neuroprotection against recurrent hypoglycemia is insufficient to prevent cognitive impairment in a rodent model of type 1 Diabetes. Diabetes. 2016; 65:3151–60. https://doi.org/10.2337/db15-1653 [PubMed]
- 28. Shukitt-Hale B. The effects of aging and oxidative stress on psychomotor and cognitive behavior. Age (Omaha). 1999; 22:9–17. https://doi.org/10.1007/s11357-999-0002-7 [PubMed]
- 29. Hauss-Wegrzyniak B, Vannucchi MG, Wenk GL. Behavioral and ultrastructural changes induced by chronic neuroinflammation in young rats. Brain Res. 2000; 859:157–66. https://doi.org/10.1016/S0006-8993(00)01999-5 [PubMed]
- 30. Carney JM, Smith CD, Carney AM, Butterfield DA. Aging- and oxygen-induced modifications in brain biochemistry and behavior. Ann N Y Acad Sci. 1994; 738:44–53. https://doi.org/10.1111/j.1749-6632.1994.tb21788.x [PubMed]
- 31. Olanow CW. A radical hypothesis for neurodegeneration. Trends Neurosci. 1993; 16:439–44. https://doi.org/10.1016/0166-2236(93)90070-3 [PubMed]
- 32. Joseph JA, Denisova N, Fisher D, Bickford P, Prior R, Cao G. Age-related neurodegeneration and oxidative stress: putative nutritional intervention. Neurol Clin. 1998; 16:747–55. https://doi.org/10.1016/S0733-8619(05)70092-X [PubMed]
- 33. Thakurta IG, Chattopadhyay M, Ghosh A, Chakrabarti S. Dietary supplementation with N-acetyl cysteine, α-tocopherol and α-lipoic acid reduces the extent of oxidative stress and proinflammatory state in aged rat brain. Biogerontology. 2012; 13:479–88. https://doi.org/10.1007/s10522-012-9392-5 [PubMed]
- 34. Meijer AJ, Codogno P. Signalling and autophagy regulation in health, aging and disease. Mol Aspects Med. 2006; 27:411–25. https://doi.org/10.1016/j.mam.2006.08.002 [PubMed]
- 35. Rajawat YS, Hilioti Z, Bossis I. Aging: central role for autophagy and the lysosomal degradative system. Ageing Res Rev. 2009; 8:199–213. https://doi.org/10.1016/j.arr.2009.05.001 [PubMed]
- 36. Poulose SM, Bielinski DF, Carrihill-Knoll K, Rabin BM, Shukitt-Hale B. Exposure to 16O-particle radiation causes aging-like decrements in rats through increased oxidative stress, inflammation and loss of autophagy. Radiat Res. 2011; 176:761–69. https://doi.org/10.1667/RR2605.1 [PubMed]
- 37. Norden DM, Trojanowski PJ, Walker FR, Godbout JP. Insensitivity of astrocytes to interleukin 10 signaling following peripheral immune challenge results in prolonged microglial activation in the aged brain. Neurobiol Aging. 2016; 44:22–41. https://doi.org/10.1016/j.neurobiolaging.2016.04.014 [PubMed]
- 38. Niraula A, Sheridan JF, Godbout JP. Microglia priming with aging and stress. Neuropsychopharmacology. 2017; 42:318–33. https://doi.org/10.1038/npp.2016.185 [PubMed]
- 39. Kovacs GG, Adle-Biassette H, Milenkovic I, Cipriani S, van Scheppingen J, Aronica E. Linking pathways in the developing and aging brain with neurodegeneration. Neuroscience. 2014; 269:152–72. https://doi.org/10.1016/j.neuroscience.2014.03.045 [PubMed]
- 40. Conover JC, Todd KL. Development and aging of a brain neural stem cell niche. Exp Gerontol. 2016S0531-5565(16)30398-9; Epub ahead of print. [PubMed]
- 41. Valero J, Paris I, Sierra A. Lifestyle shapes the dialogue between environment, microglia, and adult neurogenesis. ACS Chem Neurosci. 2016; 7:442–53. https://doi.org/10.1021/acschemneuro.6b00009 [PubMed]
- 42. Hattiangady B, Kuruba R, Shetty AK. Acute seizures in old age leads to a greater loss of CA1 pyramidal neurons, an increased propensity for developing chronic TLE and a severe cognitive dysfunction. Aging Dis. 2011; 2:1–17. [PubMed]
- 43. Coras R, Siebzehnrubl FA, Pauli E, Huttner HB, Njunting M, Kobow K, Villmann C, Hahnen E, Neuhuber W, Weigel D, Buchfelder M, Stefan H, Beck H, et al. Low proliferation and differentiation capacities of adult hippocampal stem cells correlate with memory dysfunction in humans. Brain. 2010; 133:3359–72. https://doi.org/10.1093/brain/awq215 [PubMed]
- 44. Mendelsohn AR, Larrick JW. Pharmaceutical rejuvenation of age-associated decline in spatial memory. Rejuvenation Res. 2016; 19:521–24. https://doi.org/10.1089/rej.2016.1903 [PubMed]
- 45. Haas CB, Kalinine E, Zimmer ER, Hansel G, Brochier AW, Oses JP, Portela LV, Muller AP. Brain Insulin administration triggers distinct cognitive and neurotrophic responses in young and aged rats. Mol Neurobiol. 2016; 53:5807–17. https://doi.org/10.1007/s12035-015-9494-6 [PubMed]
- 46. Parmalee NL, Aschner M. Manganese and aging. Neurotoxicology. 2016; 56:262–68. https://doi.org/10.1016/j.neuro.2016.06.006 [PubMed]
- 47. Wood JG, Rogina B, Lavu S, Howitz K, Helfand SL, Tatar M, Sinclair D. Sirtuin activators mimic caloric restriction and delay ageing in metazoans. Nature. 2004; 430:686–89. https://doi.org/10.1038/nature02789 [PubMed]
- 48. Baur JA, Pearson KJ, Price NL, Jamieson HA, Lerin C, Kalra A, Prabhu VV, Allard JS, Lopez-Lluch G, Lewis K, Pistell PJ, Poosala S, Becker KG, et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature. 2006; 444:337–42. https://doi.org/10.1038/nature05354 [PubMed]
- 49. Chung JH, Manganiello V, Dyck JR. Resveratrol as a calorie restriction mimetic: therapeutic implications. Trends Cell Biol. 2012; 22:546–54. https://doi.org/10.1016/j.tcb.2012.07.004 [PubMed]
- 50. Pardo J, Uriarte M, Cónsole GM, Reggiani PC, Outeiro TF, Morel GR, Goya RG. Insulin-like growth factor-I gene therapy increases hippocampal neurogenesis, astrocyte branching and improves spatial memory in female aging rats. Eur J Neurosci. 2016; 44:2120–28. https://doi.org/10.1111/ejn.13278 [PubMed]
- 51. Patterson SL. Immune dysregulation and cognitive vulnerability in the aging brain: interactions of microglia, IL-1β, BDNF and synaptic plasticity. Neuropharmacology. 2015; 96:11–18. https://doi.org/10.1016/j.neuropharm.2014.12.020 [PubMed]
- 52. Ji M, Dong L, Jia M, Liu W, Zhang M, Ju L, Yang J, Xie Z, Yang J. Epigenetic enhancement of brain-derived neurotrophic factor signaling pathway improves cognitive impairments induced by isoflurane exposure in aged rats. Mol Neurobiol. 2014; 50:937–44. https://doi.org/10.1007/s12035-014-8659-z [PubMed]
- 53. Curran BP, Murray HJ, O’Connor JJ. A role for c-Jun N-terminal kinase in the inhibition of long-term potentiation by interleukin-1beta and long-term depression in the rat dentate gyrus in vitro. Neuroscience. 2003; 118:347–57. https://doi.org/10.1016/S0306-4522(02)00941-7 [PubMed]
- 54. Kelly A, Vereker E, Nolan Y, Brady M, Barry C, Loscher CE, Mills KH, Lynch MA. Activation of p38 plays a pivotal role in the inhibitory effect of lipopolysaccharide and interleukin-1 beta on long term potentiation in rat dentate gyrus. J Biol Chem. 2003; 278:19453–62. https://doi.org/10.1074/jbc.M301938200 [PubMed]
- 55. Vereker E, Campbell V, Roche E, McEntee E, Lynch MA. Lipopolysaccharide inhibits long term potentiation in the rat dentate gyrus by activating caspase-1. J Biol Chem. 2000; 275:26252–58. https://doi.org/10.1074/jbc.M002226200 [PubMed]
- 56. Vereker E, O’Donnell E, Lynch MA. The inhibitory effect of interleukin-1beta on long-term potentiation is coupled with increased activity of stress-activated protein kinases. J Neurosci. 2000; 20:6811–19. [PubMed]
- 57. Tong L, Balazs R, Soiampornkul R, Thangnipon W, Cotman CW. Interleukin-1 beta impairs brain derived neurotrophic factor-induced signal transduction. Neurobiol Aging. 2008; 29:1380–93. https://doi.org/10.1016/j.neurobiolaging.2007.02.027 [PubMed]
- 58. Mariga A, Zavadil J, Ginsberg SD, Chao MV. Withdrawal of BDNF from hippocampal cultures leads to changes in genes involved in synaptic function. Dev Neurobiol. 2015; 75:173–92. https://doi.org/10.1002/dneu.22216 [PubMed]
- 59. Yang Z, Qiao H, Sun Z, Li X. Effect of BDNF-plasma-collagen matrix controlled delivery system on the behavior of adult rats neural stem cells. J Biomed Mater Res A. 2013; 101:599–606. https://doi.org/10.1002/jbm.a.34331 [PubMed]
- 60. Bramham CR, Messaoudi E. BDNF function in adult synaptic plasticity: the synaptic consolidation hypothesis. Prog Neurobiol. 2005; 76:99–125. https://doi.org/10.1016/j.pneurobio.2005.06.003 [PubMed]
- 61. Lu B. BDNF and activity-dependent synaptic modulation. Learn Mem. 2003; 10:86–98. https://doi.org/10.1101/lm.54603 [PubMed]
- 62. Tyler WJ, Alonso M, Bramham CR, Pozzo-Miller LD. From acquisition to consolidation: on the role of brain-derived neurotrophic factor signaling in hippocampal-dependent learning. Learn Mem. 2002; 9:224–37. https://doi.org/10.1101/lm.51202 [PubMed]
- 63. Joseph JA, Shukitt-Hale B, Casadesus G. Reversing the deleterious effects of aging on neuronal communication and behavior: beneficial properties of fruit polyphenolic compounds. Am J Clin Nutr. 2005 (Suppl ); 81:313S–16S. [PubMed]
- 64. Shukitt-Hale B, Lau FC, Carey AN, Galli RL, Spangler EL, Ingram DK, Joseph JA. Blueberry polyphenols attenuate kainic acid-induced decrements in cognition and alter inflammatory gene expression in rat hippocampus. Nutr Neurosci. 2008; 11:172–82. https://doi.org/10.1179/147683008X301487 [PubMed]
- 65. Olszanecki R, Gebska A, Kozlovski VI, Gryglewski RJ. Flavonoids and nitric oxide synthase. J Physiol Pharmacol. 2002; 53:571–84. [PubMed]
- 66. Rice-Evans CA, Miller NJ, Bolwell PG, Bramley PM, Pridham JB. The relative antioxidant activities of plant-derived polyphenolic flavonoids. Free Radic Res. 1995; 22:375–83. https://doi.org/10.3109/10715769509145649 [PubMed]
- 67. Bickford PC, Gould T, Briederick L, Chadman K, Pollock A, Young D, Shukitt-Hale B, Joseph J. Antioxidant-rich diets improve cerebellar physiology and motor learning in aged rats. Brain Res. 2000; 866:211–17. https://doi.org/10.1016/S0006-8993(00)02280-0 [PubMed]
- 68. Casadesus G, Shukitt-Hale B, Stellwagen HM, Zhu X, Lee HG, Smith MA, Joseph JA. Modulation of hippocampal plasticity and cognitive behavior by short-term blueberry supplementation in aged rats. Nutr Neurosci. 2004; 7:309–16. https://doi.org/10.1080/10284150400020482 [PubMed]
- 69. Joseph JA, Shukitt-Hale B, Denisova NA, Bielinski D, Martin A, McEwen JJ, Bickford PC. Reversals of age-related declines in neuronal signal transduction, cognitive, and motor behavioral deficits with blueberry, spinach, or strawberry dietary supplementation. J Neurosci. 1999; 19:8114–21. [PubMed]
- 70. Joseph JA, Denisova NA, Arendash G, Gordon M, Diamond D, Shukitt-Hale B, Morgan D. Blueberry supplementation enhances signaling and prevents behavioral deficits in an Alzheimer disease model. Nutr Neurosci. 2003; 6:153–62. https://doi.org/10.1080/1028415031000111282 [PubMed]
- 71. Shukitt-Hale B, Galli RL, Meterko V, Carey A, Bielinski DF, McGhie T, Joseph JA. Dietary supplementation with fruit polyphenolics ameliorates age-related deficits in behavior and neuronal markers of inflammation and oxidative stress. Age (Dordr). 2005; 27:49–57. https://doi.org/10.1007/s11357-005-4004-9 [PubMed]
- 72. Youdim KA, Shukitt-Hale B, Martin A, Wang H, Denisova N, Bickford PC, Joseph JA. Short term dietary supplementation of blueberry polyphenolics: beneficial effects on aging brain performance and peripheral tissue function. Nutr Neurosci. 2000; 3:383–97. https://doi.org/10.1080/1028415X.2000.11747338
- 73. Goyarzu P, Malin DH, Lau FC, Taglialatela G, Moon WD, Jennings R, Moy E, Moy D, Lippold S, Shukitt-Hale B, Joseph JA. Blueberry supplemented diet: effects on object recognition memory and nuclear factor-kappa B levels in aged rats. Nutr Neurosci. 2004; 7:75–83. https://doi.org/10.1080/10284150410001710410 [PubMed]
- 74. Joseph JA, Denisova NA, Arendash G, Gordon M, Diamond D, Shukitt-Hale B, Morgan D. Blueberry supplementation enhances signaling and prevents behavioral deficits in an Alzheimer disease model. Nutr Neurosci. 2003; 6:153–62. https://doi.org/10.1080/1028415031000111282 [PubMed]
- 75. Joseph JA, Fisher DR, Carey AN. Fruit extracts antagonize Abeta- or DA-induced deficits in Ca2+ flux in M1-transfected COS-7 cells. J Alzheimers Dis. 2004; 6:403–11. [PubMed]
- 76. Galli RL, Bielinski DF, Szprengiel A, Shukitt-Hale B, Joseph JA. Blueberry supplemented diet reverses age-related decline in hippocampal HSP70 neuroprotection. Neurobiol Aging. 2006; 27:344–50. https://doi.org/10.1016/j.neurobiolaging.2005.01.017 [PubMed]
- 77. Andres-Lacueva C, Shukitt-Hale B, Galli RL, Jauregui O, Lamuela-Raventos RM, Joseph JA. Anthocyanins in aged blueberry-fed rats are found centrally and may enhance memory. Nutr Neurosci. 2005; 8:111–20. https://doi.org/10.1080/10284150500078117 [PubMed]
- 78. Manna SK, Mukhopadhyay A, Aggarwal BB. Resveratrol suppresses TNF-induced activation of nuclear transcription factors NF-kappa B, activator protein-1, and apoptosis: potential role of reactive oxygen intermediates and lipid peroxidation. J Immunol. 2000; 164:6509–19. https://doi.org/10.4049/jimmunol.164.12.6509 [PubMed]
- 79. Yun X, Maximov VD, Yu J, Zhu H, Vertegel AA, Kindy MS. Nanoparticles for targeted delivery of antioxidant enzymes to the brain after cerebral ischemia and reperfusion injury. J Cereb Blood Flow Metab. 2013; 33:583–92. https://doi.org/10.1038/jcbfm.2012.209 [PubMed]
- 80. Hook GR, Yu J, Sipes N, Pierschbacher MD, Hook V, Kindy MS. The cysteine protease cathepsin B is a key drug target and cysteine protease inhibitors are potential therapeutics for traumatic brain injury. J Neurotrauma. 2014; 31:515–29. https://doi.org/10.1089/neu.2013.2944 [PubMed]
- 81. Lebel CP, Bondy SC. Sensitive and rapid quantitation of oxygen reactive species formation in rat synaptosomes. Neurochem Int. 1990; 17:435–40. https://doi.org/10.1016/0197-0186(90)90025-O [PubMed]
- 82. Driver AS, Kodavanti PR, Mundy WR. Age-related changes in reactive oxygen species production in rat brain homogenates. Neurotoxicol Teratol. 2000; 22:175–81. https://doi.org/10.1016/S0892-0362(99)00069-0 [PubMed]
- 83. Keller JN, Hanni KB, Kindy MS. Oxidized high-density lipoprotein induces neuron death. Exp Neurol. 2000; 161:621–30. https://doi.org/10.1006/exnr.1999.7276 [PubMed]
- 84. Gary DS, Bruce-Keller AJ, Kindy MS, Mattson MP. Ischemic and excitotoxic brain injury is enhanced in mice lacking the p55 tumor necrosis factor receptor. J Cereb Blood Flow Metab. 1998; 18:1283–87. https://doi.org/10.1097/00004647-199812000-00001 [PubMed]