



Introduction
A tremendous growth in the proportion of elderly people raises a range of challenges to societies worldwide. Healthy aging should therefore be a main priority for all countries across the globe. However, science behind the study of age-associated diseases is increasing and common molecular mechanisms that could be used to dissect longevity pathways and develop safe and effective interventions for aging are being explored. In this regard, novel methodologies using the power of artificial intelligence (AI) are emerging to cope with the massive amount of data that are becoming available [1]. A collaborative effort based on transfer of technology and knowledge between academia and industry is also needed to accelerate aging discoveries and facilitate better transition of effective interventions into clinics. To accelerate this, the Aging Research and Drug Discovery (ARDD) meeting was founded seven years ago in Basel, Switzerland. This year’s meeting, organized by Alex Zhavoronkov, Insilico Medicine, Morten Scheibye-Knudsen, University of Copenhagen and Daniela Bakula, University of Copenhagen, was particularly challenging due to the ongoing COVID-19 pandemic. The 7th ARDD meeting, 1st to 4th of September 2020, moved online with local hosting at the University of Copenhagen. We were very fortunate to have 65 fantastic speakers and more than 2200 ‘ARDDists’ (Figure 1). This report provides an overview of the presentations covering topics on some of the latest methodologies to study aging, molecular characterization of longevity pathways, existing aging interventions and the importance of aging research for the global society and economy.
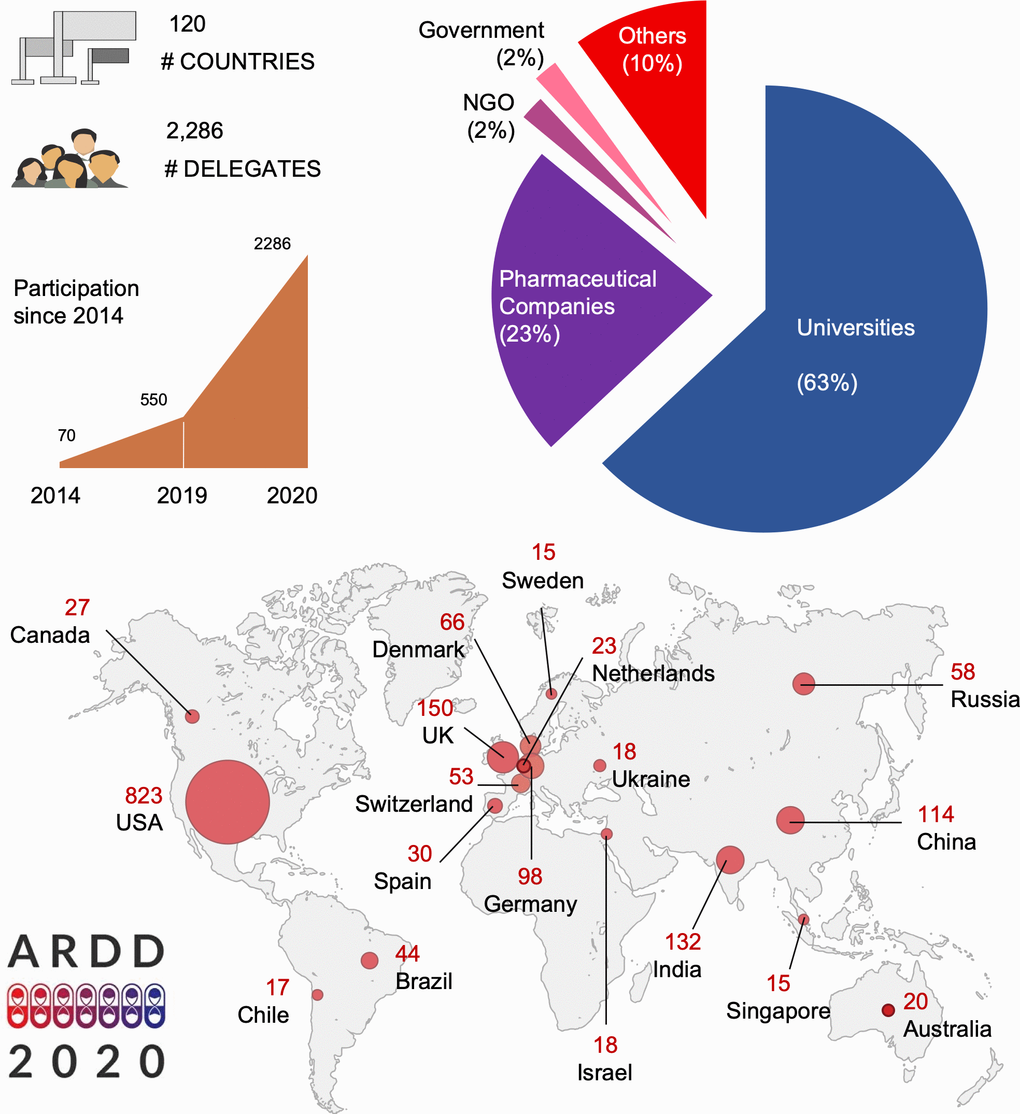
Figure 1. Statistics from ARDD2020.
Novel approaches to study aging
The progress in discoveries of basic mechanisms of aging as well as development of novel interventional strategies depends primarily on approaches and tools that are used in the lab. During the last couple of decades, methodological strategies for emerging big data from various cell-, tissue- and organ-types have accelerated towards development of high-throughput screening techniques and computational approaches using the power of artificial intelligence [1]. However, despite the capacity of data analysis, existing methods are constantly improving and becoming integrated as a part of intervention-screening platforms. Martin Borch Jensen, Gordian Biotechnology, San Francisco, USA, presented their approach for conducting high-throughput screens of many therapies in a single animal, using single-cell sequencing. By identifying cells within a diseased tissue that appear healthy after receiving one of many interventions, they are able to test in vivo efficacy of therapies much faster than traditional drug development. Another achievement in the development of advanced technology has been made in the area of proteomics, discussed by Andreas Mund, University of Copenhagen, Denmark. In particular, single cell proteomes can be analysed to extract valuable information regarding disease mechanisms. Its integration with multiplexed imaging of human-derived tissue samples and deep learning techniques enables the creation of an advanced deep visual proteomics pipeline for discovery of novel biomarkers and effective therapeutics that could potentially be used in clinics [2]. Further, Ieva Bagdonaite, University of Copenhagen, Denmark, underlined the value of investigating protein glycosylation that undergoes dynamic changes in age-associated diseases, as well as highlighted the importance of mapping complex glycoproteome using advanced quantitative proteomics for both developing efficient therapies and biomarker discovery [3].
Benefits of omics-based big data analysis were also presented by Christian Riedel, Karolinska Institutet, Sweden, who developed an advanced screening approach for geroprotector discovery. In this approach, screening of novel compounds is based on the transcriptome analysis of human tissues from people of varying ages and the application of machine learning to create age classifiers that predict biological age [4]. He looks for compounds that can shift transcriptomes of “older” towards “younger” tissues and thereby identifies drug candidates with potential geroprotective capabilities. Transcriptomics and other omics-based tools are also actively used by Vadim Gladyshev’s team, Brigham and Women’s Hospital, Harvard Medical School, USA. Particularly, comparative analysis of global transcriptomics and metabolomics data across species with different lifespan, across known longevity interventions, and across cell types with different lifespan can be used to develop an unbiased approach for the discovery of novel longevity interventions [5, 6]. Such approach can be intensely applied to determine the relationship among different types of interventions and its association with lifespan extension [7]. Identification of common signatures of longevity also raises great opportunities for high-throughput screening for novel compounds that are candidates for lifespan extension. Besides interventions, genetic studies enable the identification of novel factors contributing to aging. Using available genotyping data and genetic traits from two human cohort studies, ultra-rare damaging mutations were identified including rarest protein-truncating variants that negatively affect lifespan and healthspan [8]. Interestingly, genetic variation that supports longevity is also protective against COVID-19, which emerges as a disease of aging [9, 10]. A genetic approach to understand aging in humans was also described by Yousin Suh, Columbia University, USA. Studies of common and rare genetic variants in centenarians can be used to dissect longevity-associated genetic variants that may potentially be applied to understand the molecular basis of healthy aging as well as to develop new therapies for improving healthspan [11, 12].
Novel animal models are also being explored for the study of aging. Dario Riccardo Valenzano, Max Planck Institute for Biology of Ageing, Cologne, Germany, demonstrated how host-microbiota interactions can be important determinants for maintaining homeostasis and modulating lifespan using the short-lived model organism African turquoise killifish [13, 14]. He showed that fecal transplantation of young microbiota to old animals rescued age-dependent decrease in abundance of microbiota and extended the lifespan of the fish. Modulation of microbial function can also impact the effect of potential pharmacological interventions in hosts, as was highlighted by Filipe Cabreiro, Imperial College London, UK, who suggested that age-dependent changes in gut microbial composition can be potentially considered one of the hallmarks of aging [15]. Using the well-known pro-longevity drug metformin and a combination of model organisms such as C. elegans and D. melanogaster together with computational approaches for modelling human microbiome data, Filipe’s team showed that diet can impact the beneficial effect of metformin. His team developed a high-throughput screening platform to identify dietary metabolites and microbial molecular pathways that are responsible for the effect of metformin on health- and lifespan [16]. In worms, this and other potential aging interventions can be tested using a novel approach to measure health by atomic force microscopy based on the assessment of worm stiffness and cuticle senescence (roughness) across age spectrum [17].
Deep aging clocks
Application of artificial intelligence for the development of novel therapeutics and biomarker discovery have been actively highlighted during the ARDD meeting. In particular, for the last couple of years several aging clocks have been developed to predict chronological and biological age based on certain clinical parameters [18]. Steve Horvath, University of California, Los Angeles, USA, demonstrated their importance in predicting not only chronological age, but also mortality risks across mammalian species referring to second generation epigenetic clocks, including PhenoAge and GrimAge [19, 20]. The translational value of epigenetic clocks was also highlighted in regard to testing aging interventions and their application in clinical trials [21]. The assessment of health state and life expectancy using deep learned clocks was further presented by Alice Kane, Harvard, USA. She developed a deep learned aging measure using a frailty index as a fast non-invasive mortality predictor for mice [22]. Another talk on estimation of chronological age was given by Anastasia Georgievskaya, Haut.AI, Tallinn, Estonia. Anastasia took an advantage of face and hand images from different age groups to create multimodal age prediction analyses as a part of a pipeline for the development of non-invasive visual biomarkers of human aging [23]. The importance of deep learning applications in healthcare and biomarker discovery was also discussed by Polina Mamoshina, Deep Longevity, Hong Kong. In particular, because different biological aging clocks may be associated with different aging processes, Deep Longevity aims to combine multiple clocks to estimate and monitor biological age over time [18, 24].
Interestingly, in humans, drug repurposing and its efficiency can be estimated not only by applying various aging clocks, but also using life-course trajectories and health-to-disease transition analyses presented by Søren Brunak from Novo Nordisk Foundation Center for Protein Research, University of Copenhagen, Denmark [25]. Using large datasets on millions of patients, this in silico approach enables the understanding of how one disease follows another and estimates if certain genes are linked to diseases [26]. The use of single patient disease trajectories spanning up to 20 years when predicting intensive care mortality highlighted how aging data and machine learning can be made actionable at the bedside as opposed to statistical assessment of larger groups of individuals [27].
Genome maintenance in aging and longevity
Age-dependent alterations in cellular pathways lead to a decline in organ function and progression of disease. Understanding what causes aging and examining common patterns of changes on gene, protein and post-translational levels during the lifespan and across multiple tissues is one of the challenging tasks for aging researchers. Jan Vijg, Albert Einstein College of Medicine, USA, underlined that somatic mutations, including point mutations and genomic rearrangements, accumulate during aging and may contribute to mortality and disease [28]. Applying single-cell sequencing allows the identification of an age-dependent exponential increase in mutation frequency in human B lymphocytes [29] and liver hepatocytes [30], and propose mechanisms of how de novo mutations accumulate from early embryogenesis to adulthood and old age, and lead to the development of disease [28]. A topic of chromosomal aging was also highlighted in regard to reproductive lifespan by Eva Hoffmann, University of Copenhagen, Denmark. Investigations of changes in fertility rate during aging showed that chromosome errors and aberrations in oocytes control natural fertility in humans [31]. Based on current knowledge of genetic regulation of reproductive aging, interventions for pregnancy loss are being developed and were discussed during the ARDD meeting.
Interestingly, a genetic network that controls reproductive aging and somatic maintenance is primarily related to pathways associated with DNA repair and cell cycle regulation. Substantial research in aging has been done towards investigating nuclear DNA damage, which associates with multiple hallmarks of aging [32], as well as developing interventional strategies for protecting the aging genome [33]. Björn Schumacher, University of Cologne, Germany, characterized molecular consequences of DNA damage either in germline or somatic cells [34, 35] and examined molecular pathways that regulate somatic maintenance using C. elegans as a model organism. His recent discovery of epigenetic modifiers that are required for maintaining lifespan after DNA damage [36] shed light on novel molecular pathways linking DNA damage, epigenetics and longevity. A connection between DNA damage/repair, post-translational modifications and longevity was also presented by Vera Gorbunova, University of Rochester, USA, who applies a comparative biology approach to study short- and long-lived animals. Her team showed that the DNA double strand break (DSB) repair efficiency shows strong positive correlation with maximum lifespan across mammalian species. Higher DNA repair efficiency in long-lived species was, in large part, due to the higher activity of the histone deacetylase and mono-ADP-rybosylase, Sirtuin 6 (SIRT6) [37]. Vera’s talk also included unpublished data in humans showing that rare missense mutations in SIRT6 sequence identified in human centenarians are associated with more efficient DNA DSB repair. The functional role of SIRT6 in DNA repair also includes acting as a sensor of DSB, observed by Debra Toiber, Ben Gurion University of the Negev, Israel. Her data illustrates that SIRT6 directly binds to DNA, is recruited to the site of damage independently of PARP, MRE11 and KU80 and triggers activation of a DNA damage response [38]. Considering that SIRT6 depletion leads to accelerated aging and neurodegeneration phenotypes in mice, targeting it could be a potential strategy for the development of novel neuroprotective therapeutics [39, 40].
Importantly, genome maintenance and age-dependent changes in gene expression patterns are primarily dependent on chromatin state and epigenetic modifications [41]. David Sinclair, Harvard, USA, showed that DSBs may drive age-dependent epigenetic alterations and loss of cellular identity. Using a transgenic mouse system for inducible creation of DSBs, he revealed that loss of epigenetic structures, an accumulation of epigenetic noise and increased predicted DNA methylation changes increase with age and DNA damage [42, 43]. Importantly, the introduction of an engineered vector expressing Yamanaka transcription factors, excluding c-Myc, regenerated axons after optic nerve crush injury and restored vision in old mice [44]. The effect was dependent on the DNA demethylases Tet1, Tet2 and TDG and was accompanied by a reversal of methylation patterns and resetting the DNA methylation clock. Interestingly, epigenetic modifications in aging were studied also in other model organisms. In worms, one of the euchromatin-associated epigenetic marks known to affect lifespan is H3K4me3, controlled by the COMPASS complex [45]. Carlos Silva Garcia, Harvard, USA, showed that COMPASS-related longevity in C. elegans is dependent on activation of SREBP1, a master regulator of lipid metabolism leading to an increase in monosaturated fatty acids that is required for lifespan extension in COMPASS-deficient C.elegans [45], and presented novel data on the involvement of CREB-regulated transcriptional coactivator CRTC1 in COMPASS-mediated lifespan extension [46]. Here, Alexey Moskalev, the Russian Academy of Sciences, Russia, discussed mechanisms associated with lifespan-prolonging effects of chromatin modifier E(z) histone methyltransferases in D. melanogaster. The increase in lifespan of heterozygous E(z) mutant flies is associated with higher resistance to different stressors and changes in expression of genes related to immune response, cell cycle, and ribosome biogenesis [47].
Longevity pathways
Longevity-associated molecular pathways are actively being explored in different model organisms. In rodents, differential gene expression in multiple tissues were described by David Glass from Regeneron, Inc., USA. Studies revealed an increase in gene expression variability during aging where bioenergetics pathways were identified to be significantly down-regulated in kidney, skeletal muscle and liver, while inflammatory signaling was upregulated in these tissues [48]. While old animals demonstrated increased activity of mammalian Target Of Rapamycin (mTOR) in skeletal muscle, it was highlighted that mTOR needs to be down-regulated for healthspan benefits but not completely inhibited. By using rapalogs, inhibitors of mTOR, the authors observed improvements in the kidney of old rats and identified key regulators, including c-Myc, that are involved in the beneficial effect of mTOR inhibition [49]. In a mouse study, partial mTOR down-regulation was also shown to be beneficial for skeletal muscle – decreasing degeneration/regeneration, and, surprisingly, increasing skeletal muscle mass [50]. Another mechanism by which down-regulation of mTOR signaling leads to longevity, was presented by Collin Ewald, ETH Zurich, Switzerland. Collin’s team discovered a hydrogen sulfide pathway as a potential longevity mechanism that is up-regulated by ATF4 in dietary restriction (DR) as a stress response to a decrease in global mRNA translation [51]. Interestingly, hydrogen sulfide has already been shown to possess beneficial effects in age-related diseases with currently running clinical trials for cardio-vascular improvements (NCT02899364 and NCT02278276). However, in addition to activation of hydrogen sulfide signaling, decreased mRNA translation and ATF4 expression have a strong regulatory link to the mitochondrial translation machinery that was also shown to impact longevity [52]. Riekelt Houtkooper, Amsterdam UMC, Netherlands showed that down-regulation of mitochondrial ribosomal proteins extends lifespan [52] and presented novel data that disruption of mitochondrial dynamics [53] synergizes with reduced mitochondrial mRNA translation in C.elegans [54]. Marte Molenaars, Amsterdam UMC, Netherlands demonstrated that there is a balance between mRNA translation in the cytosol and in mitochondria, and inhibiting mitochondrial translation leads to the repression of cytosolic translation and lifespan extension via atf-5/ATF4 [55].
Another molecular mechanism tightly related with mitochondria and other organelles maintenance that is impaired during aging is autophagy [56]. Ana Maria Cuervo, Albert Einstein College of Medicine, USA, presented the importance of selective, and in particular, chaperone-mediated autophagy (CMA) in aging and age-related diseases [57]. Her recently developed mouse model to monitor CMA in vivo revealed that CMA activity is activated upon starvation in multiple organs, but that there are cell-type specific differences in this response [58]. CMA decline in most organs and tissues at different rate and contributes to loss of proteostasis and subsequent cell function. In the case of neurons, reduced CMA in mice leads to gradual alterations in motor-coordination and cognitive function suggesting that targeting selective autophagy can be a treatment option in neurodegenerative diseases [59]. Anais Franco Romero, University of Padova, Italy, presented data on the identification of novel FOXO-dependent genes that are related to longevity. One of the hits, the MYTHO gene, was found to be highly up-regulated in old mice and humans compared to young ones, and is associated with impairments in the autophagy machinery and motor function alterations in C. elegans and D. Rerio model organisms. A novel mechanism linking lysosomal function and mitochondria was presented by Nuno Raimundo, Universitätsmedizin Göttingen, Germany, who studied the consequences of impaired lysosomal acidification in aging and the development of neurodegeneration. Specifically, the data illustrates that blockage of lysosomal acidification via vATPase inhibition leads to the accumulation of iron in lysosomes and cellular iron deficiency resulting in impaired mitochondrial function and development of inflammation both in cultured neurons and in the brain of mice [60].
Importantly, because multiple cellular maintenance pathways associated with longevity requires high energy consumption, mitochondrial function can be considered to possess a pivotal role to affect biological aging. Luigi Ferrucci, National Institute on Aging - NIH, USA, highlighted that restoration of mitochondrial biogenesis and function may be achieved by temporary but not long-term blockage of major energy-consuming regulatory pathways. More specifically, Luigi’s talk was aimed at explaining why mitochondrial function declines with age and presented evidence of reduced resting muscle perfusion, altered lipid biosynthetic pathways and impaired activity of the carnitine shuttle [61, 62].
Lifestyle strategies for metabolic interventions
Identification of longevity-associated molecular pathways and the discovery of novel biomarkers of human aging goes along with the development of effective strategies for interventions. Currently, multiple interventions have been developed to target cellular metabolism, which is considered to possess a critical function in aging process [63]. Lifestyle interventions, including diet and its nutrient composition, regulate metabolic balance and affect lifespan across different species. Andrey Parkhitko, University of Pittsburgh, USA, discussed the role of the non-essential amino acid tyrosine in aging. In fly experiments, tyrosine levels decrease with age, accompanied by an increase of tyrosine-catabolic pathways. Preliminary data revealed that down-regulation of enzymes of tyrosine degradation, including the rate-limiting tyrosine aminotransferase (TAT), extends the lifespan of D. Melanogaster [64]. How protein affects lifespan has been further explored by Dudley Lamming, University of Wisconsin, USA [65]. Dudley’s team dissected the role of essential dietary amino acids in regulating lifespan. Reduction of branched chain amino acids (Leucine, Isoleucine and Valine) increases metabolic health, reducing adiposity and improving glucose tolerance in mice [66]. Further studies revealed that the low BCAA diet possess geroprotective properties, extending the lifespan of two progeroid mouse models, improving metabolic health in wild-type mice throughout their lifespan, and extending the lifespan and reducing the frailty of wild-type male, but not female, mice. These effects may be mediated in part by a sex-specific effect of a low BCAA diet on mTORC1 signaling [67]. Furthermore, Heidi Pak, University of Wisconsin, USA, illustrated that fasting is required for the beneficial effect of caloric restriction on healthspan and lifespan (unpublished data). Specifically, fasting was necessary to detect improvements in insulin sensitivity and to obtain the distinct metabolomic and transcriptomic signatures observed in caloric restricted male mice. Similarly, Pam Taub, UC San Diego, USA described the beneficial role of fasting as a part of lifestyle strategies for patients with cardiometabolic disease [68]. Importantly, the circadian rhythm was highlighted to have an important role for driving metabolism and affecting the efficiency of interventions. One dietary intervention that does not affect the robustness of the circadian rhythm is time-restricted eating (TRE). Pam’s recently published study illustrated that 10 h TRE can be used as a safe and effective lifestyle intervention, together with standard medications that are applied for treatment of cardiometabolic syndrome. However, besides metabolic diseases, fasting and caloric restriction display beneficial effects also in diseases associated with premature aging. Jan Hoeijmakers, Erasmus Medical Center Rotterdam, Netherlands, presented data that caloric restriction positively affects behavior and extends the lifespan in ERCC1-deficient progeroid mice, and reduces tremors and improves the cognitive function in a human patient.
Another lifestyle intervention that has a beneficial role for healthy aging is exercise. Thomas Rando, Stanford, USA, underlined the regenerative potential of skeletal muscles from young species that decline during aging. He showed that exercise in the form of running improves functionality of muscle stem cells almost to the level of young cells and increases the aged muscle capacity to repair injury in mice [69]. The improved function of muscle stem cells in old animals after exercise was associated with up-regulation of Cyclin D1, suppression of TGFbeta signaling and an exit from quiescence [69]. However, besides a decreased capacity of muscle regeneration, a decline in muscle function is also known to occur during aging. Gerard Karsenty, Columbia University, USA highlighted that age-dependent decline in muscle function and exercise capacity can be restored using osteocalcin. Circulating levels of this bone-derived hormone dramatically decreases already in middle age, surges after running and this hormone favors muscle function during exercise without affecting muscle mass, through two mechanisms in part. First, osteocalcin signaling in myofibers promotes uptake of glucose and fatty acids and the catabolism of these nutrients to produce ATP molecules needed for muscle function during exercise. Second, osteocalcin signaling in myofibers up-regulates the release in the circulation of muscle-derived interleukin-6 that in a feed forward loop increases the release of osteocalcin by bone during exercise and thereby exercise capacity [70]. Injection of osteocalcin increases the exercise capacity, fully restores muscle function and increases muscle mass in aged mice [70, 71]. Recent data also revealed that osteocalcin outperforms one of the leading compounds that is being tested for sarcopenia already in late clinical trials.
Benefits of exercise training for muscle function were also described in the context of maintenance of nicotinamide adenine dinucleotide (NAD+) metabolism by Jonas Thue Treebak, University of Copenhagen, Denmark. A rate-limiting enzyme of NAD+ metabolism, nicotinamide phosphoribosyltransferase (NAMPT), declines with age and was shown to be the only enzyme from the NAD+ salvage pathway that is restored by aerobic and resistance exercise training in human skeletal muscle [72]. Recent studies revealed that knockout of NAMPT in mouse skeletal muscle leads to a strong reduction in muscle function, dystrophy and premature death, suggesting a crucial role of NAMPT for maintaining NAD+ levels in skeletal muscle.
Pharmacological approaches to modulate healthspan and lifespan
Molecular and therapeutic importance of NAD+ metabolism for aging was underlined in multiple talks at the ARDD meeting. Eric Verdin, Buck Institute, USA introduced the concept of competition among major NAD+-utilizing enzymes for NAD+ that may explain its age-dependent decline across multiple tissues [73]. The main focus of the talk was CD38, a NAD+-metabolizing enzyme that increases with age in adipose tissues [74]. Verdin’s team discovered that CD38 activity is increased in M1 macrophages during aging and its activation depended on key cytokines from the senescence-associated secretory phenotype (SASP) secreted by senescent cells. Brenna Osborne, University of Copenhagen, Denmark further illustrated that depletion of CD38 appears to exacerbate some of the aging phenotypes in the mouse model of Cockayne syndrome, where another major NAD+-utilizing enzyme poly(ADP) ribose polymerase 1 (PARP1), is hyperactivated. Overall, current data suggest that a crosstalk between NAD+-utilizing enzymes needs to be continuously investigated in order to develop safe and effective interventions targeting NAD+ metabolism. However, precursors of NAD+ are actively being tested in various age-associated disorders. Evandro Fei Fang, University of Oslo, Norway underlined the importance of the NAD+-mitophagy/autophagy axis in aging and neurodegeneration and presented data on how impairment of this axis contributes to the progression in accelerated aging diseases as well as in the most common dementia, the age-predisposed Alzheimer’s disease [75, 76]. Induction of mitophagy either by NAD+ or other mitophagy stimulators inhibits amyloid-beta and p-Tau aggregates, as well as improves memory impairments in several models of Alzheimer’s disease [77, 78]. Similar results were observed by Lene Juel Rasmussen from Center for Healthy Aging, University of Copenhagen, Denmark. Lene’s team uses in vitro and in vivo animal models with a deficiency in the DNA repair gene REV1, which causes replication stress and premature aging. Suppression of REV1 is associated with high PARP1 activity, low endogenous NAD+ and low SIRT1 expression [79]. Presented data showed that mitochondrial dysfunction and morphology changes were suppressed, and autophagy was increased after nicotinamide riboside (NR) supplementation in REV1-deficient cells and that NR increased the lifespan and healthspan of REV1-deficient nematodes. Importantly, the underlying cause of the development of premature aging disorders described before are impairments in genes associated with DNA repair [80]. Morten Scheibye-Knudsen, University of Copenhagen, Denmark, demonstrated the importance of targeting DNA repair for healthy aging and illustrated how the power of AI can be applied to find novel DNA repair stimulators. Particularly, an in silico approach enabled the identification of novel compounds that are able to delay replicative aging and reverse senescent phenotypes in multiple primary cells, as well as improve the behavior and extend the lifespan in wild-type D. Melanogaster (unpublished data).
Another recently uncovered molecule that is able to improve mitochondrial function via mitophagy is Urolithin A, a gut microbiome metabolite known to improve mitochondrial function via mitophagy, increases muscle function and possesses geroprotective features across multiple species [81]. Pénélope Andreux, Amazentis, Switzerland presented results from a double blinded placebo controlled study showing that urolithin A administration in healthy elderly people is safe and was bioavailable after single or multiple doses over a 4-week period [82]. Oral consumption of urolithin A decreased plasma acylcarnitines, a sign of improved systemic mitochondrial function, and displayed transcriptomic signatures of improved mitochondrial and cellular health in muscle. Interventions targeting autophagy pathways were also highlighted by Rafael de Cabo, National Institute on Aging-NIH, USA, in the context of obesity and metabolic health. Recent data showed that disulfiram treatment prevents high-fat diet-induced obesity in mice by reducing feeding efficiency, decreasing body weight, and increasing energy expenditure [83]. Moreover, disulfiram prevents pancreatic islet hyperplasia and protects against high-fat diet-induced hepatic steatosis and fibrosis. Further experiments uncovered common molecular signatures after disulfiram treatment, revealing pathways associated with lipid and energy metabolism, redox, and detoxification and identified autophagy as one of the key targets by which disulfiram mediates its beneficial effects in cell culture [84]. The link between metabolic health and age-related bone loss was highlighted by Moustapha Kassem, Molecular Endocrinology Unit, University of Southern Denmark, Denmark, who suggested targeting skeletal mesenchymal stem cells (MSC) for the treatment of age-related osteoporosis. A decline in bone marrow composition, as well as alterations in the function of MSC in bone remodelling, are known to occur during aging [85]. The Kassem team identified the KIAA1199 protein to be highly secreted from hMSCs during osteoblast differentiation in vitro [86] and is associated with recruitment of hMSC to bone formation sites [85].
Another “classical” pro-longevity pathway that is explored for the development of aging interventions is the IGF signaling pathway. For example, targeting IGFBP-specific PAPP-A protease using genetically modified mouse models leads to lifespan extension [87, 88]. Here, Adam Freund, Calico Life Sciences LLC, USA, investigated targeting PAPP-A using antibodies. RNA sequencing revealed treatment with anti-PAPP-A to down-regulate collagen and extracellular matrix genes across multiple tissues. Further investigations identified MSCs to be a primary responder to PAPP-A inhibition. Restraining MSC activity is likely to be a mechanism driving a systemic response of tissues to PAPP-A inhibition. However, further experiments are required for the development of safe and effective therapeutic strategies for reducing IGF signaling.
Importantly, IGF-1 and other pro-aging factors may trigger activation of the NF-kB signaling cascade leading to inflammation and the development of senescent phenotypes, suggesting that NF-kB plays a key role in modulating the aging process [89, 90]. Lei Zhang, University of Minnesota, USA, applied an in silico approach to screen compounds capable of disrupting IKKβ and NEMO association thereby inhibiting NF-kB transcriptional activation [91]. A small molecule called SR12343 was identified to suppress lipopolysaccharide (LPS)-induced acute pulmonary inflammation in mice and attenuate necrosis and muscle degeneration in a mouse model of Duchenne muscular dystrophy [91]. SR12343 also attenuated senescent cell phenotypes in vitro as well as in mouse models of premature aging. A late life intervention with SR12343 in naturally aged mice demonstrated a decrease in senescent markers in liver and muscle. Hence, pharmacological targeting of NF-kB activation offers considerable potential for improving healthspan.
Interventions targeting senescent cells
Notably, studies of multiple interventions in different aging models include examinations of various markers of cellular senescence. Its significance for the aging process has been shown multiple times across model systems [92]. Senescent cells occur in all organs, including post-mitotic brain tissues, during aging and at sites of age-related pathologies. The SASPs of senescent cells lead to chronic inflammation and may contribute to the development of various cellular phenotypes associated with aging and diseases. Hence, a novel class of drugs targeting senescent cells are emerging, including senolytics (selective elimination of senescent cells) and senomorphics (selective modification of senescent cells). However, it should be considered that cellular senescence is a balancing act between its beneficial and detrimental roles in maintaining tissue homeostasis, as described by Judith Campisi from Buck Institute, USA. For instance, removal of senescent cells by senolytic drugs is one strategy to combat aging phenotypes [93]. However, no single senolytic drug eliminates all senescent cells, likely due to the heterogeneity among cells and distinct cell-type specific differences and variations in the SASP [94]. Moreover, it was highlighted that the SASP also varies depending on the senescence inducer [93] underlining the question: “what drives cells into senescence during natural aging?”. In particular, this question was addressed by Kotb Abdelmohsen, National Institute on Aging - NIH, USA, who presented data on the identification of a transcriptome signature of cellular senescence based on RNA sequencing [95]. His team identified the microRNA miR-340-5p to be highly expressed in senescence triggered by several inducers across multiple cell types. MiR-340-5p promotes senescence through the downstream effector Lamin B receptor (LBR). They also discovered that miR-340-5p is senolytic-associated or senomiR that sensitizes senescent cells to senolytic drugs.
Several strategies were proposed to target senescent cells. Marco Demaria, ERIBA, Netherlands, demonstrated the important role of oxygen in the development of the senescence phenotype [96]. Data illustrated that growth arrest, lysosomal activity and DNA damage signalling were similarly activated in senescent cells cultured at 1% or 5% oxygen, but induction of the SASP was suppressed by low oxygen. Tissues exposed to low oxygen also expressed a lower SASP than more oxygenated ones. It was demonstrated that hypoxia restrains SASP via AMPK activation and mTOR inhibition, and that intermittent treatment with hypoxia mimetic compounds can serve as a potential strategy for the reduction of SASP in vivo. Further, Peter de Keizer, University Medical Center Utrecht, Netherlands underlined again the problem of the existence of distinct subtypes of cellular senescence and the absence of senescence-specific markers. A strategy of FOXO4-p53 targeting using a designed FOXO4 peptide and other FOXO4-p53 inhibitory compounds can be applied to selectively eliminate senescence cells that appear during aging, as well as “senescence-like” chemoresistant cancer cells [97]. Laura Niedernhofer, University of Minnesota, USA, demonstrated a senolytic activity of fisetin, a natural flavonoid that improves the health- and lifespan in mouse models of normal and accelerated aging [98]. It was highlighted that several clinical trials with fisetin, also in regard to COVID-19, are under way.
Another application of small molecules, resveralogues, to target senescent cells by reversing their phenotype was presented by Richard Faragher, University of Brighton, UK [99]. A range of compounds based on resveratrol were able to reverse senescent phenotypes and restore proliferative capacity by altering mRNA splicing and moderating splicing factor levels [100]. Those compounds that were also able to activate SIRT1 demonstrated greater abilities to rescue cells from the senescence state [101].
Interestingly, screening of novel molecules using advanced AI-based tools and targeting senescent cells is also emerging. Carolina Reis, OneSkin, USA, underlined the importance of skin aging and illustrated why targeting senescent cells with novel senotherapeutic compounds can promote skin health in order to delay the onset of age-related diseases. Cell-based drug screening identified a lead compound, OS-1, that was able to reduce senescent cell burden and protect cells from UVB-induced photoaging. In addition, OS-1 was shown to reduce the molecular age of the skin using their developed skin-specific epigenetic clock [102] and showed benefits for skin health in a clinical study. Further experiments are being performed to examine whether OS-1 affects lifespan and healthspan in model organisms.
Besides that application of lifestyle strategies that in many cases can mimic pharmacological therapies or possess synergetic effects for healthspan and lifespan, one should consider more upstream events on the level of prediction of disease. In this regard, development of non-invasive biomarkers for human aging acquires special significance. Majken Jensen, University of Copenhagen, Denmark illustrated the value of investigating high-density lipoprotein (HDL) in the context of cardiovascular diseases and demonstrated HDL containing apoC3 to be the only subtype of HDL that was associated with higher risk of heart disease [103]. The problem of missing stable biomarkers for dementia prediction and Alzheimer disease was also underlined in Jensen’s talk. Recently, published data revealed plasma apoE in HDL and lacking apoC3 was associated with lower dementia risk and better cognitive function [104]. This and other novel biomarkers for Alzheimer disease can be discovered using non-targeted proteomic profiling in cerebrospinal fluid (CSF) [105]. The importance of aging of the tissue producing the CSF was further demonstrated by Nanna MacAulay, University of Copenhagen, Denmark. Alterations associated with dysregulation of CSF can lead to several pathologies, including stroke-related brain edema and hydrocephalus. Water cotransporter mechanisms, rather than conventional osmotic driving forces, were highlighted to play a crucial role in the production of CSF and secretion from the blood to the brain. The Na+/K+/2Cl- cotransporter (NKCC1) was identified to mediate approximately half of the CSF production, and thus provides opportunities for developing novel interventional strategies for pathologies associated with elevated brain fluid levels [106].
Challenges in aging: science, society and economy
Currently, our understanding of the molecular basis of aging and age-associated diseases is improving. However, challenges in the aging field exist and refer to both science and society in general. Nir Barzilai, Albert Einstein College of Medicine, USA, highlighted several concerns including (1) the translational value of identified longevity mechanisms and effective interventions from animals to humans, (2) the discovery of reliable biomarkers for estimation of efficiency of various therapies and (3) the existence of possible antagonistic effects between different gerotherapeutics. Importantly, these challenges may be overcome because evolutionarily conserved molecular signatures of longevity between humans and animals have been identified (unpublished) and novel aging biomarkers that distinguish specific signatures of longevity are emerging [107]. Current knowledge also highlights careful consideration of the combination of geroprotectors that potentially may not lead to synergistic effects, with the example of the known pro-longevity drug metformin [108]. Additionally, COVID-19 research was mentioned in regard to the study of aging as an opportunity to advance geroscience. Aubrey de Grey from SENS Research Foundation, USA, also highlighted this topic and underlined the importance of thinking about COVID-19 in a broader way to target not only the immune system but aging in general, which raises a challenge to disseminate this knowledge to the public to raise awareness of the biology of aging and the possibility of interventions. Hence, novel strategies need to be implemented in order to engage the public into the field of aging.
Another challenging topic discussed by Brian Kennedy, National University of Singapore, Singapore, related to the pros and cons of different ways of testing longevity interventions. Aging interventions would benefit most by applying prevention-based approaches and biomarker discovery [109]. Understanding how different physiological measures and aging clocks correspond to each other could allow targeted testing of different types of aging interventions, including the recently published life-extending molecule 2-oxoglutarate [110]. Further, João Pedro de Magalhães, University of Liverpool, UK, touched upon the topic of longevity interventions and highlighted the exponential growth of pharmacological approaches (DrugAge database), while research in genes associated with longevity have plateaued in recent years (GenAge database). Such shift towards drug discovery is accompanied by the appearance of an anti-aging biotech sector that could bring huge economic benefits in the future [111]. However, the lifespan of anti-aging companies is relatively small due to limitations in the time and ability to validate interventions, likely related to a lack of reliable aging biomarkers. Hence, in silico-based approaches are being applied to overcome such limitations to identify either novel genes associated with aging phenotypes [112] or discover drug candidates for life extension [113].
AI in aging and longevity
At the meeting several talks have been presented showing the power of AI in healthcare and the longevity industry. Kai-Fu Lee, Sinovation Ventures and Sinovation AI Institute, China explained different aspects of artificial intelligence and underlined deep learning to possess amazing attributes and provide great opportunities for the longevity sector. Particularly, deep learning is emerging in every aspect of healthcare and could advance longevity research with new analyses of omics-based big data. Different types of deep learning, including reinforcement learning and transfer learning, were highlighted in the context of building a knowledge-based network for health status prediction. Machine learning techniques are also used to develop a broad range of aging clocks that can be pooled together to conduct AgeMetric, a master predictor of the heath state of an individual [114]. Application of AgeMetric scores were further discussed by Alex Zhavoronkov, Insilico Medicine, Hong Kong who presented how AI-based drug discovery can lead to commercial innovation in longevity. In particular, next generation AI accelerates multiple aspects of biotechnology, including identification of novel targets, a rapid development of small molecules based on known targets [115] and prediction of outcomes of clinical trials. AI outperforms in many aspects humans if enough data is provided. By using different types of data and training a network on relatively healthy people, it is possible to re-train the network on particular disease and identify features that are specific to the diseases. Application of Agemetric scores and other AI-powered preventive medicine strategies in longevity medicine not only enables the prediction of biological age and tracking of healthy state, but also can be used to understand whether an individual is aging faster and how to intervene and slow down aging.
The power of big data was also underlined by Wei-Wu He, Human Longevity Inc., USA, who discussed using big data to personalize aging interventions. These included not only omics data but also whole-body imaging techniques such as MRI. The possibility of combining very large amount of data is a unique opportunity for tailor made longevity solutions for every individual [116].
Sergey Young, Longevity Vision Fund, USA, further unravelled key driving forces within the longevity sector and explained why it is a favorable time to invest in the industry. This includes the appearance of several breakthrough innovations and a growing number of elderly people, along with an exponential increase in the prevalence of chronic age-related disease and unhealthy lifestyle. Further, the integration of AI and technology into medicine, healthcare and the longevity space will likely lead to business opportunities. All this is accompanied by an increase in capital and favourable support in the context of policies and regulations by the government.
Last, Jim Mellon, Juvenescence, UK talked about how Juvenescence focuses on ways for improving human healthspan with the mission to extend healthy lifespan by 8-10 years in the upcoming future. It was highlighted that the biotech world possesses high risks with further complications in predictions of outcomes and achievements. One of the strategies to overcome such limitations is investment in multiple higher risk projects, with the likelihood that one or two of them will succeed. An important aspect of funded projects is the application of AI to accelerate drug development and bring drugs to market as quickly as possible. A Juvenescence pipeline includes focusing on diseases that have a major commercial impact on pro-longevity effects later on, with the example of targeting chronic kidney disease, Alzheimer disease and liver disease. In addition, a fruitful partnership with leading experts in aging and pharma companies facilitates upcoming fully developed products and FDA-approved clinical trials.
Conclusions
Current knowledge shows that aging is a very complex but plastic process. Conserved molecular pathways underlining aging can be manipulated using genetic, pharmacological and non-pharmacological approaches to significantly improve the healthspan and lifespan in model organisms, and perhaps humans. A collaborative effort between academic research with a growing number of emerging biotech companies, as well as increased investment funds to accelerate discoveries, will most likely bring effective aging pharmaceuticals in the near future. Undoubtedly, we will see more in the coming ARDD meeting in September 2021 in New York City. The future is bright.
Author Contributions
G.V.M. and M.S.K. prepared the original draft of the manuscript. The remaining authors contributed equally for editing and critically reviewing the final version.
Acknowledgments
We dedicate this meeting summary to Jay Mitchell who was a treasured colleague and who passed away too early. We will miss him dearly.
Conflicts of Interest
E.F.F. has CRADA arrangements with ChromaDex, is a consultant to Aladdin Healthcare Technologies and the Vancouver Dementia Prevention Centre. M.S.K. is a consultant for Deep Longevity, AscentaTrio Health, Molecule, Novo Nordisk and the Longevity Vision Fund. M.S.K. is the owner of Soosys.com, Forsoegsperson.dk and a co-founder of Tracked.bio. D.A.S. is a consultant to, inventor of patents licensed to, board member of and equity owner of Iduna Therapeutics, a Life Biosciences company developing epigenetic reprograming therapies. D.A.S. is an advisor to Zymo Research, an epigenetics tools company. Additional disclosures are at https://genetics.med.harvard.edu/sinclair/people/sinclair-other.php.
Funding
We would like to thank Insilico Medicine, Roche, Regeneron, Human Longevity Inc, the Longevity Vision Fund, Haut.ai, Foxo BioScience and Aging for their critical support of this meeting.
S.B. is supported by the Novo Nordisk Foundation (grants NNF17OC0027594 and NNF14CC0001). F.C. acknowledges funding from the Wellcome Trust/Royal Society (102532/Z/12/Z and 102531/Z/13/A). A.M.C is supported in part by grants from JPB and Rainwaters Foundation. C.G.S.G. is funded by the NIH/NIA (K99AG065508). E.F.F. is supported by HELSE SØR-ØST (#2017056), the Research Council of Norway (#262175 and #277813), the National Natural Science Foundation of China (#81971327), an Akershus University Hospital Strategic grant (#269901), and a Rosa Sløyfe grant (#207819) from the Norwegian Cancer Society. V.N.G. is supported by grants from the National Institute on Aging. D.W.L. is supported in part by research grants and funds from the National Institutes of Health (AG041765, AG050135, AG051974, AG056771 and AG062328), the Wisconsin Partnership Program, the Progeria Research Foundation, the American Federation for Aging Research, and the University of Wisconsin-Madison School of Medicine and Public Health and Department of Medicine. The Lamming laboratory is supported in part by the U.S. Department of Veterans Affairs (I01-BX004031), and this work was supported using facilities and resources from the William S. Middleton Memorial Veterans Hospital. H.H.P. is supported by a NIA F31 predoctoral fellowship (NIA F31 AG066311). The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH. This work does not represent the views of the Department of Veterans Affairs or the United States Government. JTT is supported by the Novo Nordisk Foundation Center for Basic Metabolic Research (CBMR). CBMR is an independent Research Center at the University of Copenhagen and partially funded by an unrestricted donation from the Novo Nordisk Foundation (NNF18CC0034900). L.N. is funded by NIH/NIA (R01 AG063543-02S1). L.Z. work is funded by NIH/NIA (U19 AG056278 and P01 AG062412). L.J.R is supported by research grants from Nordea-fonden, Olav Thon Foundation, Novo Nordisk Foundation (NNF17OC0027812). L.J.R. is a member of the Clinical Academic Group: Recovery Capacity After Acute Illness in And Aging Population (RECAP). C.G.R. is supported by the Swedish Research Council (VR) grants (2015-03740, 2017-06088, and 2019-04868), the COST grant BM1408 (GENiE), and an ICMC project grant. D.T. work is funded by The David and Inez Myers foundation and ISF 188/17. D.A.S is supported by the Harvard Medical School Epigenetics Seed Grant and Development Grant, The Paul F. Glenn Foundation for Medical Research, and NIH awards (R01AG019719 and R37AG028730). B.S. acknowledges funding from the Deutsche Forschungsgemeinschaft (SCHU 2494/3-1, SCHU 2494/7-1, SCHU 2494/10-1, SCHU 2494/11-1, CECAD, SFB 829, KFO 286, KFO 329, and GRK2407), the Deutsche Krebshilfe (70112899), and the H2020-MSCA-ITN-2018 (Healthage and ADDRESS ITNs). D.B. is supported by the Lundbeck foundation (# R303-2018-3159). M.S.K. is funded through the Nordea Foundation (#02-2017-1749), the Novo Nordisk Foundation (#NNF17OC0027812), the Neye Foundation, the Lundbeck Foundation (#R324-2019-1492), the Ministry of Higher Education and Science (#0238-00003B) and Insilico Medicine.
References
- 1. Osborne B, Bakula D, Ben Ezra M, Dresen C, Hartmann E, Kristensen SM, Mkrtchyan GV, Nielsen MH, Petr MA, Scheibye-Knudsen M. New methodologies in ageing research. Ageing Res Rev. 2020; 62:101094. https://doi.org/10.1016/j.arr.2020.101094 [PubMed]
- 2. Coscia F, Doll S, Bech JM, Schweizer L, Mund A, Lengyel E, Lindebjerg J, Madsen GI, Moreira JM, Mann M. A streamlined mass spectrometry-based proteomics workflow for large-scale FFPE tissue analysis. J Pathol. 2020; 251:100–12. https://doi.org/10.1002/path.5420 [PubMed]
- 3. Bagdonaite I, Pallesen EM, Ye Z, Vakhrushev SY, Marinova IN, Nielsen MI, Kramer SH, Pedersen SF, Joshi HJ, Bennett EP, Dabelsteen S, Wandall HH. O-glycan initiation directs distinct biological pathways and controls epithelial differentiation. EMBO Rep. 2020; 21:e48885. https://doi.org/10.15252/embr.201948885 [PubMed]
- 4. Janssens GE, Lin XX, Millan-Ariño L, Kavšek A, Sen I, Seinstra RI, Stroustrup N, Nollen EA, Riedel CG. Transcriptomics-based screening identifies pharmacological inhibition of Hsp90 as a means to defer aging. Cell Rep. 2019; 27:467–80.e6. https://doi.org/10.1016/j.celrep.2019.03.044 [PubMed]
- 5. Ma S, Yim SH, Lee SG, Kim EB, Lee SR, Chang KT, Buffenstein R, Lewis KN, Park TJ, Miller RA, Clish CB, Gladyshev VN. Organization of the mammalian metabolome according to organ function, lineage specialization, and longevity. Cell Metab. 2015; 22:332–43. https://doi.org/10.1016/j.cmet.2015.07.005 [PubMed]
- 6. Ma S, Avanesov AS, Porter E, Lee BC, Mariotti M, Zemskaya N, Guigo R, Moskalev AA, Gladyshev VN. Comparative transcriptomics across 14 Drosophila species reveals signatures of longevity. Aging Cell. 2018; 17:e12740. https://doi.org/10.1111/acel.12740 [PubMed]
- 7. Tyshkovskiy A, Bozaykut P, Borodinova AA, Gerashchenko MV, Ables GP, Garratt M, Khaitovich P, Clish CB, Miller RA, Gladyshev VN. Identification and application of gene expression signatures associated with lifespan extension. Cell Metab. 2019; 30:573–93.e8. https://doi.org/10.1016/j.cmet.2019.06.018 [PubMed]
- 8. Shindyapina AV, Zenin AA, Tarkhov AE, Santesmasses D, Fedichev PO, Gladyshev VN. Germline burden of rare damaging variants negatively affects human healthspan and lifespan. Elife. 2020; 9:e53449. https://doi.org/10.7554/eLife.53449 [PubMed]
- 9. Santesmasses D, Castro JP, Zenin AA, Shindyapina AV, Gerashchenko MV, Zhang B, Kerepesi C, Yim SH, Fedichev PO, Gladyshev VN. COVID-19 is an emergent disease of aging. Aging Cell. 2020; 19:e13230. https://doi.org/10.1111/acel.13230 [PubMed]
- 10. Ying K, Zhai R, Pyrkov TV, Mariotti M, Fedichev PO, Shen X, Gladyshev VN. Genetic and Phenotypic Evidence for the Causal Relationship Between Aging and COVID-19. medRxiv. Cold Spring Harbor Laboratory Press; 2020. https://doi.org/10.1101/2020.08.06.20169854
- 11. Zhu Y, Tazearslan C, Suh Y. Challenges and progress in interpretation of non-coding genetic variants associated with human disease. Exp Biol Med (Maywood). 2017; 242:1325–34. https://doi.org/10.1177/1535370217713750 [PubMed]
- 12. Zhu Y, Suh Y. Tri-4C: efficient identification of cis-regulatory loops at hundred base pair resolution. bioRxiv. Cold Spring Harbor Laboratory; 2019.
- 13. Smith P, Willemsen D, Popkes M, Metge F, Gandiwa E, Reichard M, Valenzano DR. Regulation of life span by the gut microbiota in the short-lived African turquoise killifish. Elife. 2017; 6:e27014. https://doi.org/10.7554/eLife.27014 [PubMed]
- 14. Valenzano DR, Benayoun BA, Singh PP, Zhang E, Etter PD, Hu CK, Clément-Ziza M, Willemsen D, Cui R, Harel I, Machado BE, Yee MC, Sharp SC, et al. The African turquoise killifish genome provides insights into evolution and genetic architecture of lifespan. Cell. 2015; 163:1539–54. https://doi.org/10.1016/j.cell.2015.11.008 [PubMed]
- 15. Bana B, Cabreiro F. The microbiome and aging. Annu Rev Genet. 2019; 53:239–61. https://doi.org/10.1146/annurev-genet-112618-043650 [PubMed]
- 16. Pryor R, Norvaisas P, Marinos G, Best L, Thingholm LB, Quintaneiro LM, De Haes W, Esser D, Waschina S, Lujan C, Smith RL, Scott TA, Martinez-Martinez D, et al. Host-microbe-drug-nutrient screen identifies bacterial effectors of metformin therapy. Cell. 2019; 178:1299–312.e29. https://doi.org/10.1016/j.cell.2019.08.003 [PubMed]
- 17. Essmann CL, Martinez-Martinez D, Pryor R, Leung KY, Krishnan KB, Lui PP, Greene ND, Brown AE, Pawar VM, Srinivasan MA, Cabreiro F. Mechanical properties measured by atomic force microscopy define health biomarkers in ageing C. Elegans. Nat Commun. 2020; 11:1043. https://doi.org/10.1038/s41467-020-14785-0 [PubMed]
- 18. Zhavoronkov A, Mamoshina P. Deep aging clocks: the emergence of AI-based biomarkers of aging and longevity. Trends Pharmacol Sci. 2019; 40:546–49. https://doi.org/10.1016/j.tips.2019.05.004 [PubMed]
- 19. Lu AT, Quach A, Wilson JG, Reiner AP, Aviv A, Raj K, Hou L, Baccarelli AA, Li Y, Stewart JD, Whitsel EA, Assimes TL, Ferrucci L, Horvath S. DNA methylation GrimAge strongly predicts lifespan and healthspan. Aging (Albany NY). 2019; 11:303–27. https://doi.org/10.18632/aging.101684 [PubMed]
- 20. Levine ME, Lu AT, Quach A, Chen BH, Assimes TL, Bandinelli S, Hou L, Baccarelli AA, Stewart JD, Li Y, Whitsel EA, Wilson JG, Reiner AP, et al. An epigenetic biomarker of aging for lifespan and healthspan. Aging (Albany NY). 2018; 10:573–91. https://doi.org/10.18632/aging.101414 [PubMed]
- 21. Fahy GM, Brooke RT, Watson JP, Good Z, Vasanawala SS, Maecker H, Leipold MD, Lin DT, Kobor MS, Horvath S. Reversal of epigenetic aging and immunosenescent trends in humans. Aging Cell. 2019; 18:e13028. https://doi.org/10.1111/acel.13028 [PubMed]
- 22. Schultz MB, Kane AE, Mitchell SJ, MacArthur MR, Warner E, Vogel DS, Mitchell JR, Howlett SE, Bonkowski MS, Sinclair DA. Age and life expectancy clocks based on machine learning analysis of mouse frailty. Nat Commun. 2020; 11:4618. https://doi.org/10.1038/s41467-020-18446-0 [PubMed]
- 23. Bobrov E, Georgievskaya A, Kiselev K, Sevastopolsky A, Zhavoronkov A, Gurov S, Rudakov K, Del Pilar Bonilla Tobar M, Jaspers S, Clemann S. PhotoAgeClock: deep learning algorithms for development of non-invasive visual biomarkers of aging. Aging (Albany NY). 2018; 10:3249–59. https://doi.org/10.18632/aging.101629 [PubMed]
- 24. Galkin F, Mamoshina P, Aliper A, Putin E, Moskalev V, Gladyshev VN, Zhavoronkov A. Human gut microbiome aging clock based on taxonomic profiling and deep learning. iScience. 2020; 23:101199. https://doi.org/10.1016/j.isci.2020.101199 [PubMed]
- 25. Hu JX, Thomas CE, Brunak S. Network biology concepts in complex disease comorbidities. Nat Rev Genet. 2016; 17:615–29. https://doi.org/10.1038/nrg.2016.87 [PubMed]
- 26. Westergaard D, Moseley P, Sørup FK, Baldi P, Brunak S. Population-wide analysis of differences in disease progression patterns in men and women. Nat Commun. 2019; 10:666. https://doi.org/10.1038/s41467-019-08475-9 [PubMed]
- 27. Nielsen AB, Thorsen-Meyer HC, Belling K, Nielsen AP, Thomas CE, Chmura PJ, Lademann M, Moseley PL, Heimann M, Dybdahl L, Spangsege L, Hulsen P, Perner A, Brunak S. Survival prediction in intensive-care units based on aggregation of long-term disease history and acute physiology: a retrospective study of the Danish National Patient Registry and electronic patient records. Lancet Digit Health. 2019; 1:e78–89. https://doi.org/10.1016/S2589-7500(19)30024-X [PubMed]
- 28. Vijg J, Dong X. Pathogenic mechanisms of somatic mutation and genome mosaicism in aging. Cell. 2020; 182:12–23. https://doi.org/10.1016/j.cell.2020.06.024 [PubMed]
- 29. Zhang L, Dong X, Lee M, Maslov AY, Wang T, Vijg J. Single-cell whole-genome sequencing reveals the functional landscape of somatic mutations in B lymphocytes across the human lifespan. Proc Natl Acad Sci USA. 2019; 116:9014–19. https://doi.org/10.1073/pnas.1902510116 [PubMed]
- 30. Brazhnik K, Sun S, Alani O, Kinkhabwala M, Wolkoff AW, Maslov AY, Dong X, Vijg J. Single-cell analysis reveals different age-related somatic mutation profiles between stem and differentiated cells in human liver. Sci Adv. 2020; 6:eaax2659. https://doi.org/10.1126/sciadv.aax2659 [PubMed]
- 31. Gruhn JR, Zielinska AP, Shukla V, Blanshard R, Capalbo A, Cimadomo D, Nikiforov D, Chan AC, Newnham LJ, Vogel I, Scarica C, Krapchev M, Taylor D, et al. Chromosome errors in human eggs shape natural fertility over reproductive life span. Science. 2019; 365:1466–69. https://doi.org/10.1126/science.aav7321 [PubMed]
- 32. Schumacher B, Vijg J. Age is in the nucleus. Nat Metab. 2019; 1:931–32. https://doi.org/10.1038/s42255-019-0125-9 [PubMed]
- 33. Petr MA, Tulika T, Carmona-Marin LM, Scheibye-Knudsen M. Protecting the aging genome. Trends Cell Biol. 2020; 30:117–32. https://doi.org/10.1016/j.tcb.2019.12.001 [PubMed]
- 34. Ermolaeva MA, Segref A, Dakhovnik A, Ou HL, Schneider JI, Utermöhlen O, Hoppe T, Schumacher B. DNA damage in germ cells induces an innate immune response that triggers systemic stress resistance. Nature. 2013; 501:416–20. https://doi.org/10.1038/nature12452 [PubMed]
- 35. Ou HL, Kim CS, Uszkoreit S, Wickström SA, Schumacher B. Somatic niche cells regulate the CEP-1/p53-mediated DNA damage response in primordial germ cells. Dev Cell. 2019; 50:167–83.e8. https://doi.org/10.1016/j.devcel.2019.06.012 [PubMed]
- 36. Wang S, Meyer DH, Schumacher B. H3K4me2 regulates the recovery of protein biosynthesis and homeostasis following DNA damage. Nat Struct Mol Biol. 2020; 27:1165–77. https://doi.org/10.1038/s41594-020-00513-1 [PubMed]
- 37. Tian X, Firsanov D, Zhang Z, Cheng Y, Luo L, Tombline G, Tan R, Simon M, Henderson S, Steffan J, Goldfarb A, Tam J, Zheng K, et al. SIRT6 is responsible for more efficient DNA double-strand break repair in long-lived species. Cell. 2019; 177:622–38.e22. https://doi.org/10.1016/j.cell.2019.03.043 [PubMed]
- 38. Onn L, Portillo M, Ilic S, Cleitman G, Stein D, Kaluski S, Shirat I, Slobodnik Z, Einav M, Erdel F, Akabayov B, Toiber D. SIRT6 is a DNA double-strand break sensor. Elife. 2020; 9:e51636. https://doi.org/10.7554/eLife.51636 [PubMed]
- 39. Kaluski S, Portillo M, Besnard A, Stein D, Einav M, Zhong L, Ueberham U, Arendt T, Mostoslavsky R, Sahay A, Toiber D. Neuroprotective functions for the histone deacetylase SIRT6. Cell Rep. 2017; 18:3052–62. https://doi.org/10.1016/j.celrep.2017.03.008 [PubMed]
- 40. Portillo M, Eremenko E, Kaluski S, Onn L, Stein D, Slobodnik Z, Ueberham U, Einav M, Brückner MK, Arendt T, Toiber D. Molecular Mechanism of Nuclear Tau Accumulation and its Role in Protein Synthesis. bioRxiv. Cold Spring Harbor Laboratory; 2020. https://doi.org/10.1101/2020.09.01.276782
- 41. Pal S, Tyler JK. Epigenetics and aging. Sci Adv. 2016; 2:e1600584. https://doi.org/10.1126/sciadv.1600584 [PubMed]
- 42. Yang JH, Griffin PT, Vera DL, Apostolides JK, Hayano M, Meer MV, Salfati EL, Su Q, Munding EM, Blanchette M, Bhakta M, Dou Z, Xu C, et al. Erosion of the Epigenetic Landscape and Loss of Cellular Identity as a Cause of Aging in Mammals. bioRxiv. Cold Spring Harbor Laboratory; 2019. https://doi.org/10.2139/ssrn.3461780
- 43. Hayano M, Yang JH, Bonkowski MS, Amorim JA, Ross JM, Coppotelli G, Griffin PT, Chew YC, Guo W, Yang X, Vera DL, Salfati EL, Das A, et al. DNA Break-Induced Epigenetic Drift as a Cause of Mammalian Aging. bioRxiv. Cold Spring Harbor Laboratory; 2019. https://doi.org/10.2139/ssrn.3466338
- 44. Lu Y, Krishnan A, Brommer B, Tian X, Meer M, Vera DL, Wang C, Zeng Q, Yu D, Bonkowski MS, Yang JH, Hoffmann EM, Zhou S, et al. Reversal of ageing- and injury-induced vision loss by Tet-dependent epigenetic reprogramming. bioRxiv. Cold Spring Harbor Laboratory; 2019. https://doi.org/10.1101/710210
- 45. Han S, Schroeder EA, Silva-García CG, Hebestreit K, Mair WB, Brunet A. Mono-unsaturated fatty acids link H3K4me3 modifiers to C. Elegans lifespan. Nature. 2017; 544:185–90. https://doi.org/10.1038/nature21686 [PubMed]
- 46. Escoubas CC, Silva-García CG, Mair WB. Deregulation of CRTCs in aging and age-related disease risk. Trends Genet. 2017; 33:303–21. https://doi.org/10.1016/j.tig.2017.03.002 [PubMed]
- 47. Moskalev AA, Shaposhnikov MV, Zemskaya NV, Koval LA, Schegoleva EV, Guvatova ZG, Krasnov GS, Solovev IA, Sheptyakov MA, Zhavoronkov A, Kudryavtseva AV. Transcriptome analysis of long-lived Drosophila melanogaster E(z) mutants sheds light on the molecular mechanisms of longevity. Sci Rep. 2019; 9:9151. https://doi.org/10.1038/s41598-019-45714-x [PubMed]
- 48. Shavlakadze T, Morris M, Fang J, Wang SX, Zhu J, Zhou W, Tse HW, Mondragon-Gonzalez R, Roma G, Glass DJ. Age-related gene expression signature in rats demonstrate early, late, and linear transcriptional changes from multiple tissues. Cell Rep. 2019; 28:3263–73.e3. https://doi.org/10.1016/j.celrep.2019.08.043 [PubMed]
- 49. Shavlakadze T, Zhu J, Wang S, Zhou W, Morin B, Egerman MA, Fan L, Wang Y, Iartchouk O, Meyer A, Valdez RA, Mannick JB, Klickstein LB, Glass DJ. Short-term low-dose mTORC1 inhibition in aged rats counter-regulates age-related gene changes and blocks age-related kidney pathology. J Gerontol A Biol Sci Med Sci. 2018; 73:845–52. https://doi.org/10.1093/gerona/glx249 [PubMed]
- 50. Joseph GA, Wang SX, Jacobs CE, Zhou W, Kimble GC, Tse HW, Eash JK, Shavlakadze T, Glass DJ. Partial inhibition of mTORC1 in aged rats counteracts the decline in muscle mass and reverses molecular signaling associated with sarcopenia. Mol Cell Biol. 2019; 39:e00141–19. https://doi.org/10.1128/MCB.00141-19 [PubMed]
- 51. Statzer C, Venz R, Bland M, Robida-Stubbs S, Meng J, Patel K, Emsley R, Petrovic D, Liu P, Morantte I, Haynes C, Filipovic M, Mair WB, et al. ATF-4 and hydrogen sulfide signalling mediate longevity from inhibition of translation or mTORC1. bioRxiv. Cold Spring Harbor Laboratory; 2020. https://doi.org/10.1101/2020.11.02.364703
- 52. Houtkooper RH, Mouchiroud L, Ryu D, Moullan N, Katsyuba E, Knott G, Williams RW, Auwerx J. Mitonuclear protein imbalance as a conserved longevity mechanism. Nature. 2013; 497:451–57. https://doi.org/10.1038/nature12188 [PubMed]
- 53. Liu YJ, McIntyre RL, Janssens GE, Houtkooper RH. Mitochondrial fission and fusion: a dynamic role in aging and potential target for age-related disease. Mech Ageing Dev. 2020; 186:111212. https://doi.org/10.1016/j.mad.2020.111212 [PubMed]
- 54. Liu YJ, McIntyre RL, Janssens GE, Williams EG, Lan J, van Weeghel M, Schomakers B, van der Veen H, van der Wel NN, Yao P, Mair WB, Aebersold R, MacInnes AW, Houtkooper RH. Mitochondrial translation and dynamics synergistically extend lifespan in C. Elegans through HLH-30. J Cell Biol. 2020; 219:e201907067. https://doi.org/10.1083/jcb.201907067 [PubMed]
- 55. Molenaars M, Janssens GE, Williams EG, Jongejan A, Lan J, Rabot S, Joly F, Moerland PD, Schomakers BV, Lezzerini M, Liu YJ, McCormick MA, Kennedy BK, et al. A conserved mito-cytosolic translational balance links two longevity pathways. Cell Metab. 2020; 31:549–63.e7. https://doi.org/10.1016/j.cmet.2020.01.011 [PubMed]
- 56. Kroemer G, Mariño G, Levine B. Autophagy and the integrated stress response. Mol Cell. 2010; 40:280–93. https://doi.org/10.1016/j.molcel.2010.09.023 [PubMed]
- 57. Kaushik S, Cuervo AM. The coming of age of chaperone-mediated autophagy. Nat Rev Mol Cell Biol. 2018; 19:365–81. https://doi.org/10.1038/s41580-018-0001-6 [PubMed]
- 58. Dong S, Aguirre-Hernandez C, Scrivo A, Eliscovich C, Arias E, Bravo-Cordero JJ, Cuervo AM. Monitoring spatiotemporal changes in chaperone-mediated autophagy in vivo. Nat Commun. 2020; 11:645. https://doi.org/10.1038/s41467-019-14164-4 [PubMed]
- 59. Scrivo A, Bourdenx M, Pampliega O, Cuervo AM. Selective autophagy as a potential therapeutic target for neurodegenerative disorders. Lancet Neurol. 2018; 17:802–15. https://doi.org/10.1016/S1474-4422(18)30238-2 [PubMed]
- 60. Yambire KF, Rostosky C, Watanabe T, Pacheu-Grau D, Torres-Odio S, Sanchez-Guerrero A, Senderovich O, Meyron-Holtz EG, Milosevic I, Frahm J, West AP, Raimundo N. Impaired lysosomal acidification triggers iron deficiency and inflammation in vivo. Elife. 2019; 8:e51031. https://doi.org/10.7554/eLife.51031 [PubMed]
- 61. Adelnia F, Cameron D, Bergeron CM, Fishbein KW, Spencer RG, Reiter DA, Ferrucci L. The role of muscle perfusion in the age-associated decline of mitochondrial function in healthy individuals. Front Physiol. 2019; 10:427. https://doi.org/10.3389/fphys.2019.00427 [PubMed]
- 62. Ferrucci L, Zampino M. A mitochondrial root to accelerated ageing and frailty. Nat Rev Endocrinol. 2020; 16:133–34. https://doi.org/10.1038/s41574-020-0319-y [PubMed]
- 63. Parkhitko AA, Filine E, Mohr SE, Moskalev A, Perrimon N. Targeting metabolic pathways for extension of lifespan and healthspan across multiple species. Ageing Res Rev. 2020; 64:101188. https://doi.org/10.1016/j.arr.2020.101188 [PubMed]
- 64. Parkhitko AA, Ramesh D, Wang L, Leshchiner D, Filine E, Binari R, Olsen AL, Asara JM, Cracan V, Rabinowitz JD, Brockmann A, Perrimon N. Downregulation of the tyrosine degradation pathway extends Drosophila lifespan. Elife. 2020; 9:e58053. https://doi.org/10.7554/eLife.58053 [PubMed]
- 65. Green CL, Lamming DW. Regulation of metabolic health by essential dietary amino acids. Mech Ageing Dev. 2019; 177:186–200. https://doi.org/10.1016/j.mad.2018.07.004 [PubMed]
- 66. Fontana L, Cummings NE, Arriola Apelo SI, Neuman JC, Kasza I, Schmidt BA, Cava E, Spelta F, Tosti V, Syed FA, Baar EL, Veronese N, Cottrell SE, et al. Decreased consumption of branched-chain amino acids improves metabolic health. Cell Rep. 2016; 16:520–30. https://doi.org/10.1016/j.celrep.2016.05.092 [PubMed]
- 67. Richardson NE, Konon EN, Schuster HS, Mitchell AT, Boyle C, Rodgers AC, Finke M, Haider LR, Yu D, Flores V, Pak HH, Ahmad S, Ahmed S, et al. Lifelong restriction of dietary branched-chain amino acids has sex-specific benefits for frailty and lifespan in mice, Nature Aging, in press.
- 68. Wilkinson MJ, Manoogian EN, Zadourian A, Lo H, Fakhouri S, Shoghi A, Wang X, Fleischer JG, Navlakha S, Panda S, Taub PR. Ten-hour time-restricted eating reduces weight, blood pressure, and atherogenic lipids in patients with metabolic syndrome. Cell Metab. 2020; 31:92–104.e5. https://doi.org/10.1016/j.cmet.2019.11.004 [PubMed]
- 69. Brett JO, Arjona M, Ikeda M, Quarta M, de Morrée A, Egner IM, Perandini LA, Ishak HD, Goshayeshi A, Benjamin DI, Both P, Rodríguez-Mateo C, Betley MJ, et al. Exercise rejuvenates quiescent skeletal muscle stem cells in old mice through restoration of Cyclin D1. Nat Metab. 2020; 2:307–17. https://doi.org/10.1038/s42255-020-0190-0 [PubMed]
- 70. Mera P, Laue K, Ferron M, Confavreux C, Wei J, Galán-Díez M, Lacampagne A, Mitchell SJ, Mattison JA, Chen Y, Bacchetta J, Szulc P, Kitsis RN, et al. Osteocalcin signaling in myofibers is necessary and sufficient for optimum adaptation to exercise. Cell Metab. 2016; 23:1078–92. https://doi.org/10.1016/j.cmet.2016.05.004 [PubMed]
- 71. Mera P, Laue K, Wei J, Berger JM, Karsenty G. Osteocalcin is necessary and sufficient to maintain muscle mass in older mice. Mol Metab. 2016; 5:1042–47. https://doi.org/10.1016/j.molmet.2016.07.002 [PubMed]
- 72. de Guia RM, Agerholm M, Nielsen TS, Consitt LA, Søgaard D, Helge JW, Larsen S, Brandauer J, Houmard JA, Treebak JT. Aerobic and resistance exercise training reverses age-dependent decline in NAD+ salvage capacity in human skeletal muscle. Physiol Rep. 2019; 7:e14139. https://doi.org/10.14814/phy2.14139 [PubMed]
- 73. Verdin E. NAD+ in aging, metabolism, and neurodegeneration. Science. 2015; 350:1208–13. https://doi.org/10.1126/science.aac4854 [PubMed]
- 74. Covarrubias AJ, Kale A, Perrone R, Lopez-Dominguez JA, Pisco AO, Kasler HG, Schmidt MS, Wiley CD, Iyer SS, Basisty N, Wu Q, Kwok R, Heckenbach I, et al. Aging-related inflammation driven by cellular senescence enhances NAD consumption via activation of CD38+ pro-inflammatory macrophages. bioRxiv. 2019. https://doi.org/10.1101/609438
- 75. Lou G, Palikaras K, Lautrup S, Scheibye-Knudsen M, Tavernarakis N, Fang EF. Mitophagy and neuroprotection. Trends Mol Med. 2020; 26:8–20. https://doi.org/10.1016/j.molmed.2019.07.002 [PubMed]
- 76. Lautrup S, Sinclair DA, Mattson MP, Fang EF. NAD+ in brain aging and neurodegenerative disorders. Cell Metab. 2019; 30:630–55. https://doi.org/10.1016/j.cmet.2019.09.001 [PubMed]
- 77. Fang EF, Hou Y, Palikaras K, Adriaanse BA, Kerr JS, Yang B, Lautrup S, Hasan-Olive MM, Caponio D, Dan X, Rocktäschel P, Croteau DL, Akbari M, et al. Mitophagy inhibits amyloid-β and tau pathology and reverses cognitive deficits in models of Alzheimer’s disease. Nat Neurosci. 2019; 22:401–12. https://doi.org/10.1038/s41593-018-0332-9 [PubMed]
- 78. Fang EF. Mitophagy and NAD+ inhibit Alzheimer disease. Autophagy. 2019; 15:1112–14. https://doi.org/10.1080/15548627.2019.1596497 [PubMed]
- 79. Fakouri NB, Durhuus JA, Regnell CE, Angleys M, Desler C, Hasan-Olive MM, Martín-Pardillos A, Tsaalbi-Shtylik A, Thomsen K, Lauritzen M, Bohr VA, de Wind N, Bergersen LH, Rasmussen LJ. Rev1 contributes to proper mitochondrial function via the PARP-NAD+-SIRT1-PGC1α axis. Sci Rep. 2017; 7:12480. https://doi.org/10.1038/s41598-017-12662-3 [PubMed]
- 80. Keijzers G, Bakula D, Scheibye-Knudsen M. Monogenic diseases of DNA repair. N Engl J Med. 2017; 377:1868–76. https://doi.org/10.1056/NEJMra1703366 [PubMed]
- 81. Ryu D, Mouchiroud L, Andreux PA, Katsyuba E, Moullan N, Nicolet-Dit-Félix AA, Williams EG, Jha P, Lo Sasso G, Huzard D, Aebischer P, Sandi C, Rinsch C, Auwerx J. Urolithin A induces mitophagy and prolongs lifespan in C. Elegans and increases muscle function in rodents. Nat Med. 2016; 22:879–88. https://doi.org/10.1038/nm.4132 [PubMed]
- 82. Andreux PA, Blanco-Bose W, Ryu D, Burdet F, Ibberson M, Aebischer P, Auwerx J, Singh A, Rinsch C. The mitophagy activator urolithin A is safe and induces a molecular signature of improved mitochondrial and cellular health in humans. Nat Metab. 2019; 1:595–603. https://doi.org/10.1038/s42255-019-0073-4 [PubMed]
- 83. Bernier M, Mitchell SJ, Wahl D, Diaz A, Singh A, Seo W, Wang M, Ali A, Kaiser T, Price NL, Aon MA, Kim EY, Petr MA, et al. Disulfiram treatment normalizes body weight in obese mice. Cell Metab. 2020; 32:203–14.e4. https://doi.org/10.1016/j.cmet.2020.04.019 [PubMed]
- 84. Bernier M, Harney D, Koay YC, Diaz A, Singh A, Wahl D, Pulpitel T, Ali A, Guiterrez V, Mitchell SJ, Kim EY, Mach J, Price NL, et al. Elucidating the mechanisms by which disulfiram protects against obesity and metabolic syndrome. NPJ Aging Mech Dis. 2020; 6:8. https://doi.org/10.1038/s41514-020-0046-6 [PubMed]
- 85. Chen L, Shi K, Andersen TL, Qiu W, Kassem M. KIAA1199 is a secreted molecule that enhances osteoblastic stem cell migration and recruitment. Cell Death Dis. 2019; 10:126. https://doi.org/10.1038/s41419-018-1202-9 [PubMed]
- 86. Kristensen LP, Chen L, Nielsen MO, Qanie DW, Kratchmarova I, Kassem M, Andersen JS. Temporal profiling and pulsed SILAC labeling identify novel secreted proteins during ex vivo osteoblast differentiation of human stromal stem cells. Mol Cell Proteomics. 2012; 11:989–1007. https://doi.org/10.1074/mcp.M111.012138 [PubMed]
- 87. Conover CA, Bale LK, Mader JR, Mason MA, Keenan KP, Marler RJ. Longevity and age-related pathology of mice deficient in pregnancy-associated plasma protein-A. J Gerontol A Biol Sci Med Sci. 2010; 65:590–99. https://doi.org/10.1093/gerona/glq032 [PubMed]
- 88. Bale LK, West SA, Conover CA. Inducible knockdown of pregnancy-associated plasma protein-A gene expression in adult female mice extends life span. Aging Cell. 2017; 16:895–97. https://doi.org/10.1111/acel.12624 [PubMed]
- 89. Zhang L, Yousefzadeh MJ, Suh Y, Niedernhofer LJ, Robbins PD. Signal transduction, ageing and disease. Subcell Biochem. 2019; 91:227–47. https://doi.org/10.1007/978-981-13-3681-2_9 [PubMed]
- 90. Salminen A, Kaarniranta K. Insulin/IGF-1 paradox of aging: regulation via AKT/IKK/NF-kappaB signaling. Cell Signal. 2010; 22:573–77. https://doi.org/10.1016/j.cellsig.2009.10.006 [PubMed]
- 91. Zhao J, Zhang L, Mu X, Doebelin C, Nguyen W, Wallace C, Reay DP, McGowan SJ, Corbo L, Clemens PR, Wilson GM, Watkins SC, Solt LA, et al. Development of novel NEMO-binding domain mimetics for inhibiting IKK/NF-κB activation. PLoS Biol. 2018; 16:e2004663. https://doi.org/10.1371/journal.pbio.2004663 [PubMed]
- 92. Campisi J, Robert L. Cell senescence: role in aging and age-related diseases. Interdiscip Top Gerontol. 2014; 39:45–61. https://doi.org/10.1159/000358899 [PubMed]
- 93. Wiley CD, Campisi J. From ancient pathways to aging cells-connecting metabolism and cellular senescence. Cell Metab. 2016; 23:1013–21. https://doi.org/10.1016/j.cmet.2016.05.010 [PubMed]
- 94. Hernandez-Segura A, de Jong TV, Melov S, Guryev V, Campisi J, Demaria M. Unmasking transcriptional heterogeneity in senescent cells. Curr Biol. 2017; 27:2652–60.e4. https://doi.org/10.1016/j.cub.2017.07.033 [PubMed]
- 95. Casella G, Munk R, Kim KM, Piao Y, De S, Abdelmohsen K, Gorospe M. Transcriptome signature of cellular senescence. Nucleic Acids Res. 2019; 47:7294–305. https://doi.org/10.1093/nar/gkz555 [PubMed]
- 96. Casciaro F, Borghesan M, Beretti F, Zavatti M, Bertucci E, Follo MY, Maraldi T, Demaria M. Prolonged hypoxia delays aging and preserves functionality of human amniotic fluid stem cells. Mech Ageing Dev. 2020; 191:111328. https://doi.org/10.1016/j.mad.2020.111328 [PubMed]
- 97. Baar MP, Brandt RM, Putavet DA, Klein JD, Derks KW, Bourgeois BR, Stryeck S, Rijksen Y, van Willigenburg H, Feijtel DA, van der Pluijm I, Essers J, van Cappellen WA, et al. Targeted apoptosis of senescent cells restores tissue homeostasis in response to chemotoxicity and aging. Cell. 2017; 169:132–47.e16. https://doi.org/10.1016/j.cell.2017.02.031 [PubMed]
- 98. Yousefzadeh MJ, Zhu Y, McGowan SJ, Angelini L, Fuhrmann-Stroissnigg H, Xu M, Ling YY, Melos KI, Pirtskhalava T, Inman CL, McGuckian C, Wade EA, Kato JI, et al. Fisetin is a senotherapeutic that extends health and lifespan. EBioMedicine. 2018; 36:18–28. https://doi.org/10.1016/j.ebiom.2018.09.015 [PubMed]
- 99. Faragher RG, Ostler EL. Resveralogues: from novel ageing mechanisms to new therapies? Gerontology. 2020; 66:231–37. https://doi.org/10.1159/000504845 [PubMed]
- 100. Latorre E, Birar VC, Sheerin AN, Jeynes JC, Hooper A, Dawe HR, Melzer D, Cox LS, Faragher RG, Ostler EL, Harries LW. Small molecule modulation of splicing factor expression is associated with rescue from cellular senescence. BMC Cell Biol. 2017; 18:31. https://doi.org/10.1186/s12860-017-0147-7 [PubMed]
- 101. Birar VC, Sheerin AN, Ostler EL, Faragher RG. Novel resveratrol derivatives have diverse effects on the survival, proliferation and senescence of primary human fibroblasts. Biogerontology. 2020; 21:817–26. https://doi.org/10.1007/s10522-020-09896-6 [PubMed]
- 102. Boroni M, Zonari A, Reis de Oliveira C, Alkatib K, Ochoa Cruz EA, Brace LE, Lott de Carvalho J. Highly accurate skin-specific methylome analysis algorithm as a platform to screen and validate therapeutics for healthy aging. Clin Epigenetics. 2020; 12:105. https://doi.org/10.1186/s13148-020-00899-1 [PubMed]
- 103. Jensen MK, Aroner SA, Mukamal KJ, Furtado JD, Post WS, Tsai MY, Tjønneland A, Polak JF, Rimm EB, Overvad K, McClelland RL, Sacks FM. High-density lipoprotein subspecies defined by presence of apolipoprotein C-III and incident coronary heart disease in four cohorts. Circulation. 2018; 137:1364–73. https://doi.org/10.1161/CIRCULATIONAHA.117.031276 [PubMed]
- 104. Koch M, DeKosky ST, Goodman M, Sun J, Furtado JD, Fitzpatrick AL, Mackey RH, Cai T, Lopez OL, Kuller LH, Mukamal KJ, Jensen MK. Association of apolipoprotein E in lipoprotein subspecies with risk of dementia. JAMA Netw Open. 2020; 3:e209250. https://doi.org/10.1001/jamanetworkopen.2020.9250 [PubMed]
- 105. Bader JM, Geyer PE, Müller JB, Strauss MT, Koch M, Leypoldt F, Koertvelyessy P, Bittner D, Schipke CG, Incesoy EI, Peters O, Deigendesch N, Simons M, et al. Proteome profiling in cerebrospinal fluid reveals novel biomarkers of Alzheimer’s disease. Mol Syst Biol. 2020; 16:e9356. https://doi.org/10.15252/msb.20199356 [PubMed]
- 106. Steffensen AB, Oernbo EK, Stoica A, Gerkau NJ, Barbuskaite D, Tritsaris K, Rose CR, MacAulay N. Cotransporter-mediated water transport underlying cerebrospinal fluid formation. Nat Commun. 2018; 9:2167. https://doi.org/10.1038/s41467-018-04677-9 [PubMed]
- 107. Lehallier B, Gate D, Schaum N, Nanasi T, Lee SE, Yousef H, Moran Losada P, Berdnik D, Keller A, Verghese J, Sathyan S, Franceschi C, Milman S, et al. Undulating changes in human plasma proteome profiles across the lifespan. Nat Med. 2019; 25:1843–50. https://doi.org/10.1038/s41591-019-0673-2 [PubMed]
- 108. Kulkarni AS, Gubbi S, Barzilai N. Benefits of metformin in attenuating the hallmarks of aging. Cell Metab. 2020; 32:15–30. https://doi.org/10.1016/j.cmet.2020.04.001 [PubMed]
- 109. Kudryashova KS, Burka K, Kulaga AY, Vorobyeva NS, Kennedy BK. Aging biomarkers: from functional tests to multi-omics approaches. Proteomics. 2020; 20:e1900408. https://doi.org/10.1002/pmic.201900408 [PubMed]
- 110. Asadi Shahmirzadi A, Edgar D, Liao CY, Hsu YM, Lucanic M, Asadi Shahmirzadi A, Wiley CD, Gan G, Kim DE, Kasler HG, Kuehnemann C, Kaplowitz B, Bhaumik D, et al. Alpha-ketoglutarate, an endogenous metabolite, extends lifespan and compresses morbidity in aging mice. Cell Metab. 2020; 32:447–56.e6. https://doi.org/10.1016/j.cmet.2020.08.004 [PubMed]
- 111. de Magalhães JP, Stevens M, Thornton D. The business of anti-aging science. Trends Biotechnol. 2017; 35:1062–73. https://doi.org/10.1016/j.tibtech.2017.07.004 [PubMed]
- 112. Avelar RA, Ortega JG, Tacutu R, Tyler EJ, Bennett D, Binetti P, Budovsky A, Chatsirisupachai K, Johnson E, Murray A, Shields S, Tejada-Martinez D, Thornton D, et al. A multidimensional systems biology analysis of cellular senescence in aging and disease. Genome Biol. 2020; 21:91. https://doi.org/10.1186/s13059-020-01990-9 [PubMed]
- 113. Calvert S, Tacutu R, Sharifi S, Teixeira R, Ghosh P, de Magalhães JP. A network pharmacology approach reveals new candidate caloric restriction mimetics in C. Elegans. Aging Cell. 2016; 15:256–66. https://doi.org/10.1111/acel.12432 [PubMed]
- 114. Zhavoronkov A, Li R, Ma C, Mamoshina P. Deep biomarkers of aging and longevity: from research to applications. Aging (Albany NY). 2019; 11:10771–80. https://doi.org/10.18632/aging.102475 [PubMed]
- 115. Zhavoronkov A, Ivanenkov YA, Aliper A, Veselov MS, Aladinskiy VA, Aladinskaya AV, Terentiev VA, Polykovskiy DA, Kuznetsov MD, Asadulaev A, Volkov Y, Zholus A, Shayakhmetov RR, et al. Deep learning enables rapid identification of potent DDR1 kinase inhibitors. Nat Biotechnol. 2019; 37:1038–40. https://doi.org/10.1038/s41587-019-0224-x [PubMed]
- 116. Hou YC, Yu HC, Martin R, Cirulli ET, Schenker-Ahmed NM, Hicks M, Cohen IV, Jönsson TJ, Heister R, Napier L, Swisher CL, Dominguez S, Tang H, et al. Precision medicine integrating whole-genome sequencing, comprehensive metabolomics, and advanced imaging. Proc Natl Acad Sci USA. 2020; 117:3053–62. https://doi.org/10.1073/pnas.1909378117 [PubMed]