



Introduction
Polycystic ovary syndrome (PCOS) is one of the most prevalent endocrine and metabolic disorders, affecting up to one in five women of childbearing age [1]. Its cardinal features are hyperandrogenism, menstrual irregularity, and polycystic ovary morphology [2]. PCOS is associated with insulin resistance (IR), metabolic syndrome, increased risk of endometrial cancer, ovulatory dysfunction, infertility, pregnancy complications, type 2 diabetes, and cardiovascular disease [3–7]. Although the pathogenesis of PCOS is complex and remains unclear, there is growing scientific consensus in that genetic factors play a key role in PCOS occurrence and development.
Long non-coding RNAs (lncRNAs) represent a class of non-coding RNA transcripts greater than 200 nucleotides in length [8]. Although lncRNAs lack protein-encoding capacity, an increasing body of evidence suggests that they act a pivotal part in many biological processes, including genetic imprinting, X-chromosome inactivation, transcriptional and post-transcriptional regulation, recruitment of epigenetic modifiers, control of mRNA decay, organelle biogenesis, and subcellular trafficking, among others [9, 10]. Accumulating evidence also suggests that lncRNA dysregulation is closely associated with numerous human diseases, including PCOS and PCOS-related conditions. For instance, results from microarray analyses showed that some lncRNAs were abnormally expressed in the granulosa cells of PCOS patients, which suggested their involvement in PCOS development [10]. Examples include HCG26, an lncRNA that participates in the regulation of ovarian granulosa cell proliferation and steroidogenesis [11], and RP11-151A6.4, which was found to be upregulated in granulosa cells from PCOS patients and suggested to promote insulin resistance, androgen excess, and adipose dysfunction [12]. However, only a few lncRNAs have been so far verifiably related to PCOS.
The competitive endogenous RNA (ceRNA) hypothesis describes a regulatory network involving mutually interacting protein-coding (mRNA) and non-coding (e.g. lncRNA, miRNA, etc.) transcripts to modulate gene and protein expression, A central tenet of the ceRNA hypothesis is that lncRNAs can regulate other RNA transcripts by competitive binding to miRNAs via shared miRNA response elements [13]. ceRNA activities have been confirmed to influence the development of several diseases, and ceRNA networks have been built for lung cancer [14], cardiac hypertrophy [15], and implantation failure [16], among other conditions. Considering the potential relevance of lncRNAs in the regulation of PCOS through the ceRNA mechanism, in this study we used bioinformatics tools to construct a PCOS-related lncRNA-mRNA network (PCLMN) to identify key lncRNAs that might impact the development and serve as biomarkers of PCOS.
Results
Identification of differentially expressed lncRNAs and mRNAs
LncRNA and mRNA expression profiles from granulosa cells from seven PCOS patients and seven controls were retrieved from the GSE95728 dataset available on the NCBI-GEO repository. Based on FC > 2 and adjusted P < 0.05, 86 differentially expressed lncRNAs (DELs) and 112 differentially expressed mRNAs (DEMs) were identified.
Construction of a global lncRNA-miRNA-mRNA triple network
A total of 50,2653 miRNA-mRNA interaction pairs were downloaded from miRTarBase and starBase. Subsequently, the lncRNASNP2 tool was applied to select candidate miRNAs targeting the 86 DELs identified in the GSE95728 dataset. DELs’ sequences were obtained from UCSC Genome Browser, and miRNA/lncRNA associations with a prediction score > 160 and binding energy < -20 were selected. A total of 66,059 miRNA-lncRNA interaction pairs were thus obtained. All selected miRNA-mRNA and miRNA-lncRNA pairs were then merged to construct a global triple network which was used as a background network to construct the PCLMN.
Construction of the PCOS-related lncRNA-mRNA network
After mapping all the DELs and DEMs into the global triple network, a hypergeometric test was performed to extract 334 lncRNA-miRNA-mRNA triplets. From these, 41 lncRNAs and 41mRNAs, linked by 203 edges, were chosen to construct the PCLMN (Figure 1B).
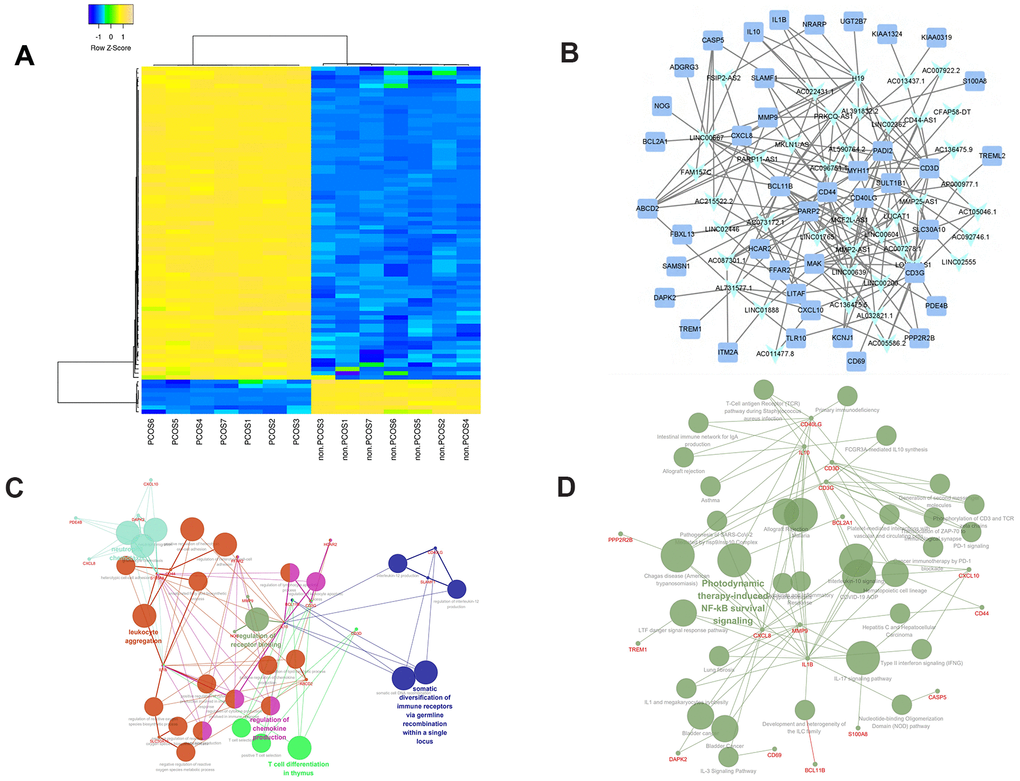
Figure 1. Clustering and enrichment analysis of the PCOS-related lncRNA-mRNA network (PCLMN). (A) Unsupervised clustering of PCLMN genes. (B) PCLMN visualization. Light-blue arrowheads indicate lncRNAs and blue squares indicate mRNAs. Grey edges indicate lncRNA-mRNA interactions. (C) Network map of enriched GO terms in the PCLMN. (D) Network map of enriched KEGG pathways in the PCLMN.
Hierarchical clustering and functional characteristics of the PCLMN
The 82 differentially expressed genes (DEGs) included in the PCLMN were grouped by unsupervised hierarchical clustering, suggesting distinct gene expression profiles for control and PCOS (Figure 1A). We next performed GO classification and KEGG pathway analysis on the DELs and DEMs in the PCLMN. GO analysis results indicated significant enrichment in 27 GO terms (P≤0.05, Benjamini-Hochberg-corrected) as listed in Figure 1C and Table 1. Meanwhile, as shown in Figure 2 and Table 2, 13 pathways were found to be enriched (P≤0.05) in the target genes.
Table 1. Enriched GO terms in the PCLMN.
| GO term | P-value | Associated genes (%) |
| regulation of receptor binding | 2.67E-05 | 10.71 |
| T cell selection | 1.64E-04 | 5.88 |
| positive T cell selection | 6.78E-05 | 7.89 |
| T cell differentiation in thymus | 2.20E-05 | 5.06 |
| granulocyte chemotaxis | 4.15E-07 | 4.32 |
| neutrophil migration | 2.80E-07 | 4.62 |
| neutrophil chemotaxis | 1.09E-07 | 5.41 |
| interleukin-12 production | 3.23E-04 | 4.69 |
| regulation of interleukin-12 production | 2.94E-04 | 4.84 |
| somatic diversification of immune receptors | 4.75E-04 | 4.11 |
| somatic cell DNA recombination | 4.75E-04 | 4.11 |
| chemokine production | 4.93E-05 | 4.12 |
| regulation of cytokine production involved in immune response | 5.56E-05 | 4.00 |
| regulation of chemokine production | 4.01E-05 | 4.35 |
| regulation of leukocyte apoptotic process | 4.01E-05 | 4.35 |
| regulation of lymphocyte apoptotic process | 2.67E-04 | 5.00 |
| heterotypic cell-cell adhesion | 3.38E-04 | 4.62 |
| chemokine production | 4.93E-05 | 4.12 |
| regulation of lipid catabolic process | 3.38E-04 | 4.62 |
| leukocyte aggregation | 3.03E-06 | 21.43 |
| regulation of reactive oxygen species biosynthetic process | 5.56E-05 | 4.00 |
| negative regulation of reactive oxygen species metabolic process | 3.53E-04 | 4.55 |
| regulation of cytokine production involved in immune response | 5.56E-05 | 4.00 |
| regulation of chemokine production | 4.01E-05 | 4.35 |
| regulation of heterotypic cell-cell adhesion | 2.39E-05 | 11.11 |
| negative regulation of reactive oxygen species biosynthetic process | 4.42E-05 | 9.09 |
| positive regulation of cytokine production involved in immune response | 3.69E-04 | 4.48 |
| positive regulation of chemokine production | 4.03E-04 | 4.35 |
| positive regulation of heterotypic cell-cell adhesion | 4.65E-06 | 18.75 |
| unsaturated fatty acid biosynthetic process | 3.38E-04 | 4.62 |
| regulation of lymphocyte apoptotic process | 2.67E-04 | 5.00 |
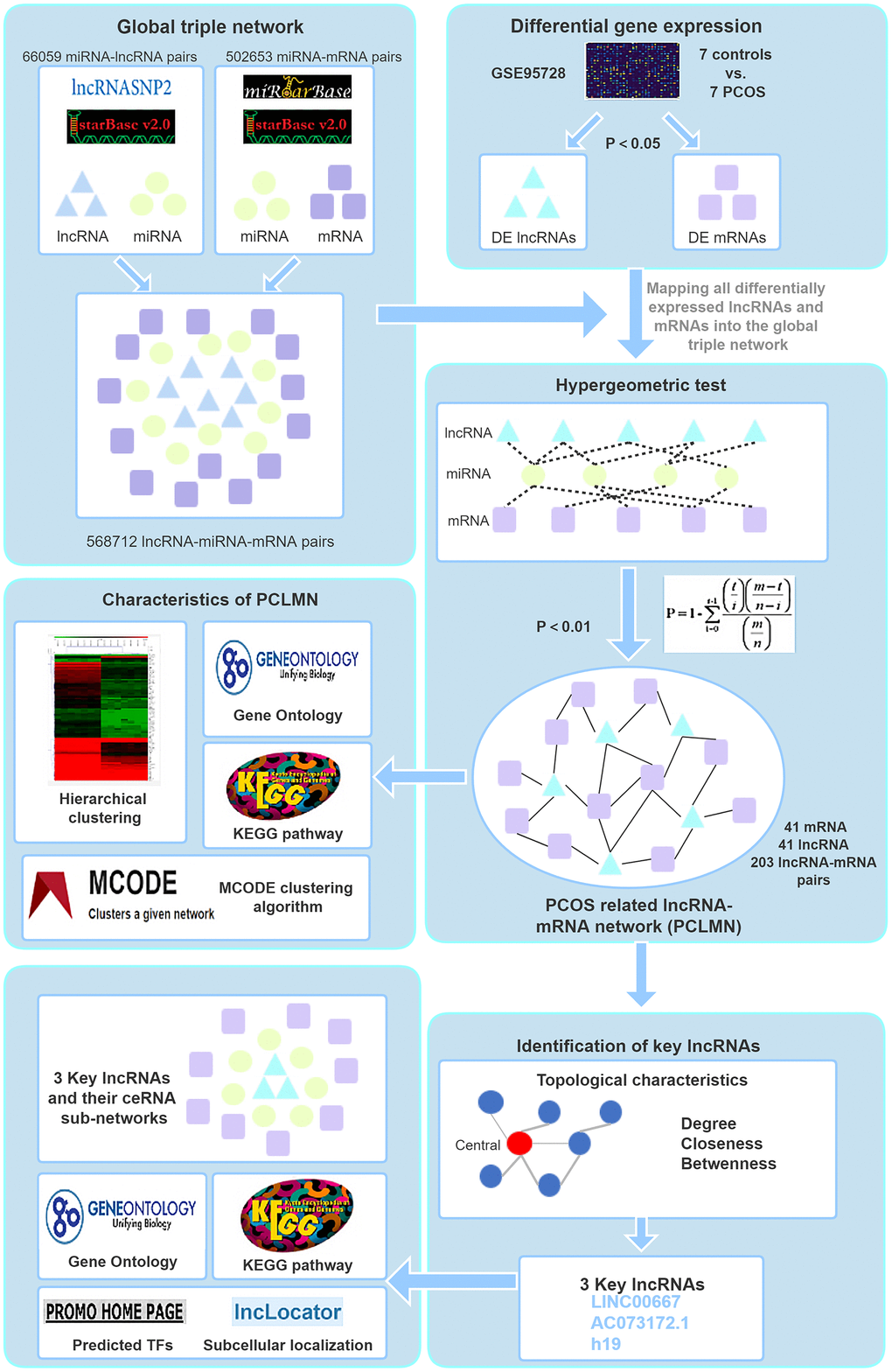
Figure 2. Study workflow. First, we constructed a global background network based on predicted lncRNA-miRNA and miRNA-mRNA pairs. Second, we applied a hypergeometric test to construct the PCLMN and performed network topology analysis to determine the lncRNAs with the highest centroid variability. Lastly, we explored the subcellular localization of the key lncRNAs thus identified, performed functional module analyses, and identified putative transcription factors regulating the expression of the candidate lncRNAs.
Table 2. Enriched KEGG pathways in the PCLMN.
| GO term | P-value | Associated genes% |
| Hematopoietic cell lineage | 2.09E-04 | 4.04 |
| IL-17 signaling pathway | 8.26E-06 | 5.32 |
| Intestinal immune network for IgA production | 9.35E-03 | 4.08 |
| Chagas disease (American trypanosomiasis) | 5.22E-07 | 5.88 |
| African trypanosomiasis | 5.42E-03 | 5.41 |
| Malaria | 1.42E-05 | 8.00 |
| Bladder cancer | 2.43E-04 | 7.32 |
| Asthma | 3.82E-03 | 6.45 |
| Allograft rejection | 5.71E-03 | 5.26 |
| Primary immunodeficiency | 5.71E-03 | 5.26 |
| Phosphorylation of CD3 and TCR zeta chains | 1.93E-03 | 9.09 |
| Translocation of ZAP-70 to Immunological synapse | 1.44E-03 | 10.53 |
| Generation of second messenger molecules | 4.59E-03 | 5.88 |
| PD-1 signaling | 2.11E-03 | 8.70 |
| Interleukin-10 signaling | 1.11E-05 | 8.51 |
| FCGR3A-mediated IL10 synthesis | 7.26E-03 | 4.65 |
| Nucleotide-binding Oligomerization Domain (NOD) pathway | 6.62E-03 | 4.88 |
| Allograft Rejection | 1.44E-04 | 4.44 |
| Bladder Cancer | 2.43E-04 | 7.32 |
| IL-3 Signaling Pathway | 9.35E-03 | 4.08 |
| IL1 and megakaryocytes in obesity | 2.50E-03 | 8.00 |
| Photodynamic therapy-induced NF-kB survival signaling | 5.56E-08 | 14.29 |
| Lung fibrosis | 8.64E-04 | 4.76 |
| Hepatitis C and Hepatocellular Carcinoma | 9.72E-03 | 4.00 |
| T-Cell antigen Receptor pathway during Staphylococcus aureus infection | 8.25E-04 | 4.84 |
| Development and heterogeneity of the ILC family | 4.07E-03 | 6.25 |
| Platelet-mediated interactions with vascular and circulating cells | 1.15E-03 | 11.76 |
| LTF danger signal response pathway | 2.71E-05 | 15.00 |
| Cancer immunotherapy by PD-1 blockade | 2.30E-03 | 8.33 |
| Pathogenesis of SARS-CoV-2 Mediated by nsp9/nsp10 Complex | 1.93E-03 | 9.09 |
| COVID-19 AOP | 9.07E-08 | 26.67 |
| Cytokines and Inflammatory Response | 2.91E-03 | 7.41 |
| Type II interferon signaling (IFNG) | 5.42E-03 | 5.41 |
Topological characteristics of the PCLMN and subcellular localization of key lncRNAs
The degree distribution of the PCLMN was examined and all the nodes were found to conform to a power-law distribution (R2 = 0.80), indicating that the network was scale-free (Figure 3A). Next, we analyzed the topological characteristics of the PCLMN to predict the biological functions of the network’s lncRNAs. Centralization metrics of degree, betweenness, and closeness were calculated. The top ten hits for each topological parameter were then selected, and three lncRNAs, i.e. LINC00667, AC073172.1, and H19, were found to intersect across the three features (Figure 3B).
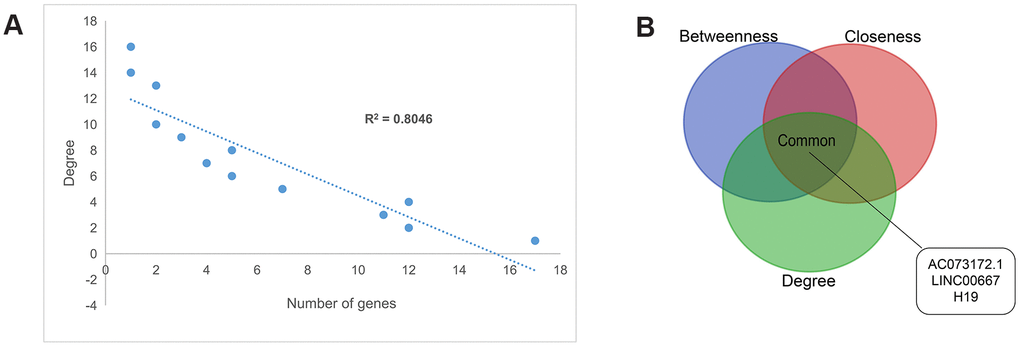
Figure 3. Topology features of the PCLMN. (A) Degree distributions of the network. All degrees followed a power-law distribution. (B) Identification of 3 lncRNA genes simultaneously ranking within the top-10 lists across degree, betweenness, and closeness.
The subcellular localization of lncRNAs provides important clues about their potential functions. To explore the subcellular localization of the key lncRNAs identified in the PCLMN, we accessed two online platforms, i.e. LncLocator and RNAlocate, and verified the results against iLoc-lncRNA, a sequence-based subcellular localization prediction tool. Summary analysis results for LINC00667, AC073172.1, and H19 are displayed in Figures 4–6, respectively. LINC00667 showed a predominant cytosolic localization, confirmed also by iLoc-LncRNA (probability score = 0.853 for ‘cytosol/cytoplasm’) (Figure 4A). As shown in Figure 4B, there were 13 nearest mRNA neighbors for LINC00667 in the PCLMN. GO analysis indicated significant associations of LINC00667 with 4 GO terms under Biological Process (BP), one term under Molecular Function (MF), and one term under Cellular Component (CC) (Figure 4C). Meanwhile, 10 KEGG pathways showed significant enrichment in LINC00667 (Figure 4D).
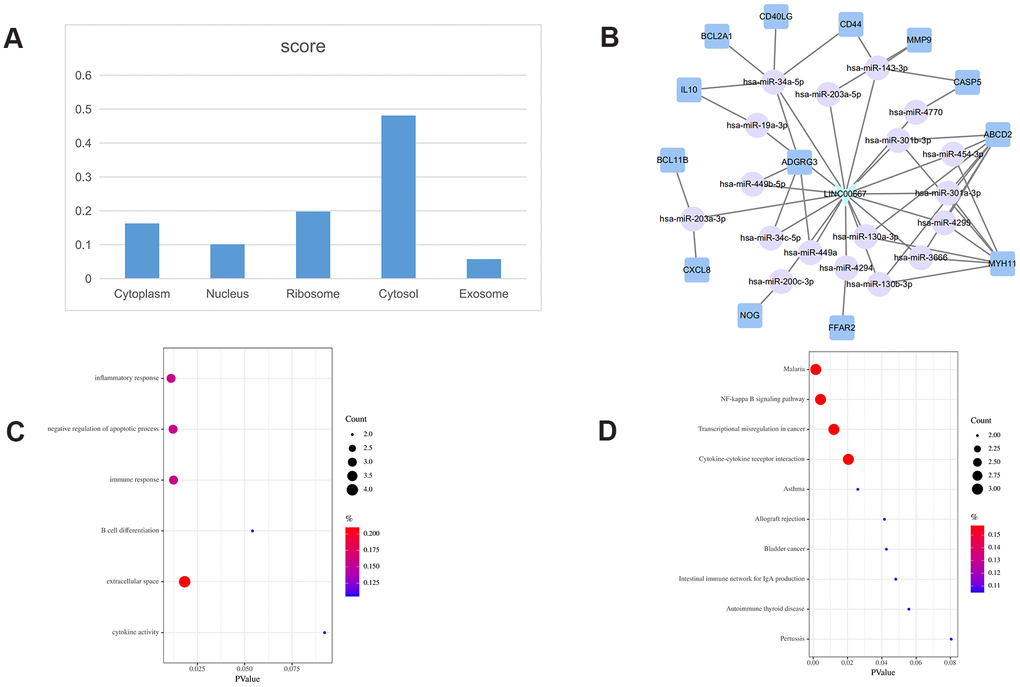
Figure 4. LINC00667-related ceRNA sub-network analysis. (A) Subcellular location analysis for LINC00667. (B) ceRNA network of LINC00667. (C) GO biological process enrichment results for LINC00667. (D) KEGG pathways enriched in LINC00667.
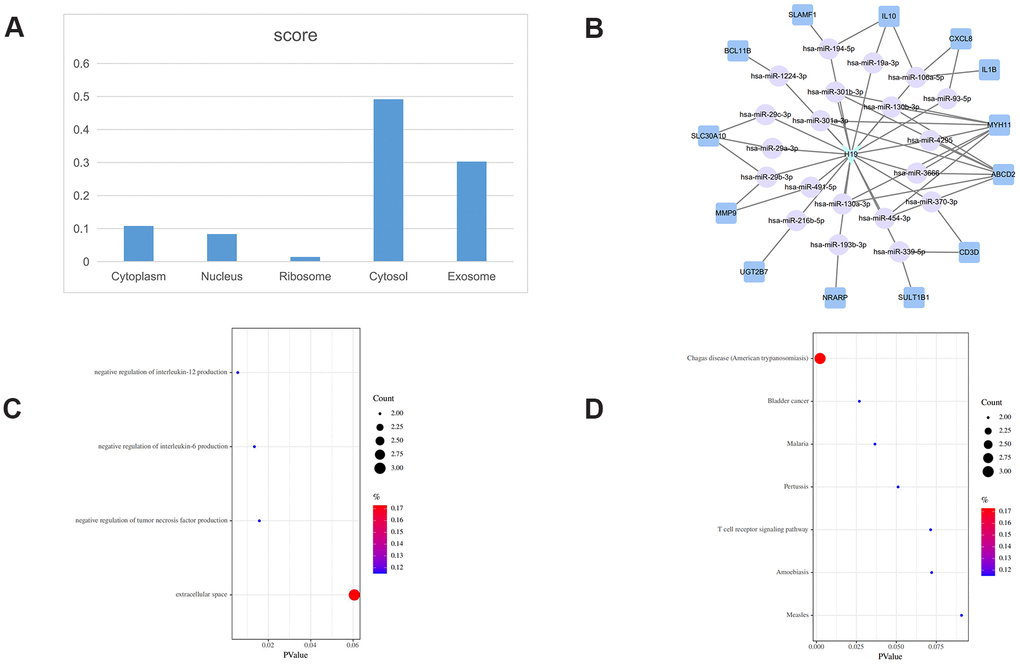
Figure 5. H19-related ceRNA sub-network. (A) Subcellular location analysis for H19. (B) ceRNA network of H19. (C) GO biological process enrichment for H19. (D) KEGG pathways enriched in H19.
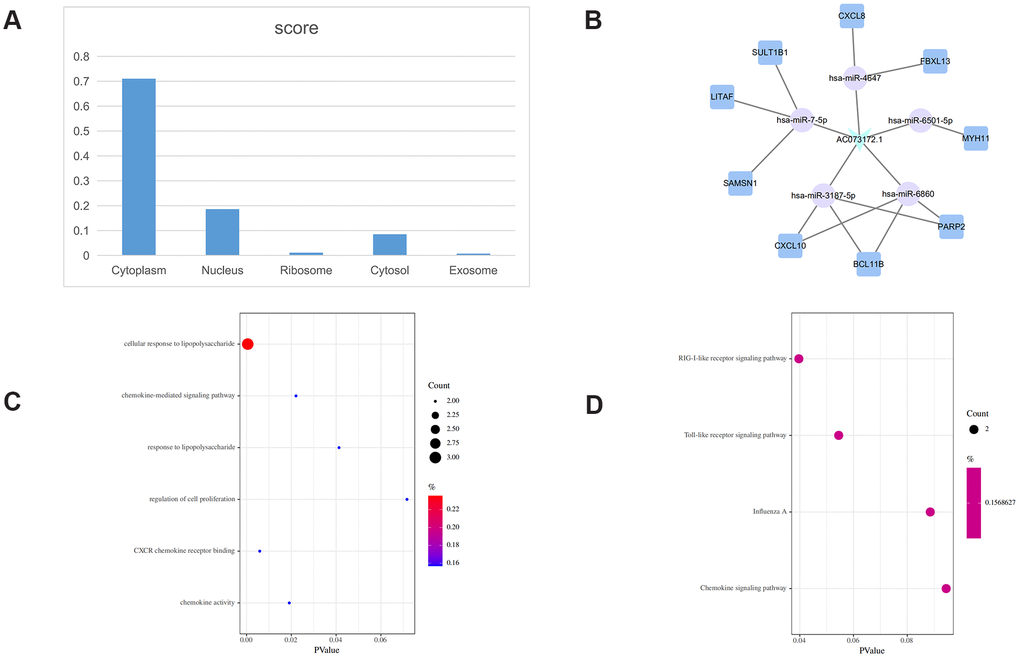
Figure 6. AC073172.1-related ceRNA sub-network. (A) Subcellular location analysis for AC073172.1. (B) ceRNA network of AC073172.1. (C) GO biological process enrichment analysis for AC073172.1. (D) KEGG pathways enriched in AC073172.1.
H19 localized also mainly to the cytoplasm (Figure 5A), with iLoc-LncRNA yielding a probability score of 0.603 for the ‘cytosol/cytoplasm’ term. Meanwhile, 13 mRNAs were identified as nearest mRNA neighbors of H19 in the PCLMN (Figure 5B). GO classification detected enrichment by H19 in 3 terms under BP and one term under CC (Figure 5C). H19 was also enriched in 7 KEGG pathways, as shown in Figure 5D.
AC073172.1 showed also a predominant cytoplasmic localization (Figure 6A), with the iLoc-LncRNA tool indicating a probability score of 0.868 for the ‘cytosol/cytoplasm’ category. Seventeen mRNAs were detected as the nearest mRNA neighbors of H19 in the PCLMN (Figure 6B). Upon GO classification, 4 BP and 2 MF terms were found to be enriched in H19 (Figure 6C). In turn, 4 enriched KEGG pathways were identified for this lncRNA (Figure 6D).
Identification of putative transcription factors regulating key lncRNAs in the PCLMN
Promoter sequence-based prediction of candidate regulatory TFs of the key lncRNAs was carried out using PROMO software with maximum matrix dissimilarity rate < 3. A total of 27 common TFs targeting the promoters of the 3 key lncRNAs were identified (Figure 7A). Of these, 4 TFs, including ER-alpha, PRA, PRB, and AR (marked in red in Figure 7A), had been previously associated with PCOS.
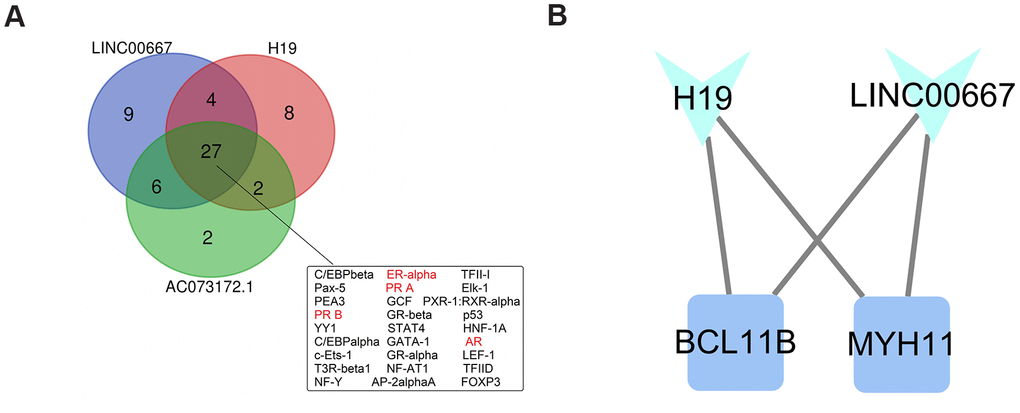
Figure 7. Identification of putative transcription factors and module analysis. (A) Venn diagram of predicted transcription factors (TFs) regulating the expression of the three key lncRNAs in the PCMLN. Among 27 TFs associated with all three lncRNAs, four (marked in red) are known regulators of the expression of sexual hormones. (B) Representation of the functional module identified in the PCLMN.
Identification of functional modules in the PCLMN
We performed functional module prediction on the PCLMN using the MCODE clustering algorithm implemented in Cytoscape. A functional module containing H19, LINC00667, and two protein-coding genes, namely MYH11 (which encodes the smooth muscle myosin heavy chain) and BCL11B (which encodes a zinc finger protein TF) was thus identified (Figure 7B).
Discussion
PCOS represents a prevalent disorder of follicular development characterized by excessive early follicular growth, arrested growth of antral follicles, and disrupted dominant follicle selection. In this study we performed a network-based computational analysis to identify and characterize lncRNAs potentially influencing PCOS. RNA/RNA interaction data from starBase, lncRNASNP2, and miRTarBase databases were used to construct a global lncRNA-miRNA-mRNA network based on the ceRNA theory. Then, a PCOS-related lncRNA-mRNA network (PCLMN) containing 41 mRNA nodes, 41 lncRNA nodes, and 203 edges was extracted by mapping DEGs in the GSE95728 dataset into the global triple network. The resulting PCLMN was then analyzed to identify putative functional biomarkers for PCOS.
GO classification and KEGG pathway analysis showed that many inflammation-related sub-categories, including ‘cytokines and inflammatory response’, ‘chemokine production’, ‘neutrophil chemotaxis’, ‘leukocyte aggregation’, ‘interleukin-10 signaling pathway’, and ‘IL-17 signaling pathway’ were enriched in the PCLMN. Previous studies underscored the contribution of chronic, low-grade inflammation to the occurrence of IR, hyperandrogenemia, and metabolic syndrome in PCOS [17, 18]. IR is highly prevalent in women with POCS and may be induced by pro-inflammatory cytokines acting directly on insulin-like receptor molecules [19]. A study found that increased serum CRP and IL-6 levels correlated with elevated insulin levels and higher insulin resistance index (HOMA-IR) scores, further suggesting that inflammatory factors are closely related to IR [20]. Along these lines, it was reported that inflammatory factor-mediated signaling inhibits the tyrosine kinase activity of the insulin receptor and interferes with insulin substrate 1 (IRS-1) synthesis, thus leading to IR by blocking insulin-dependent signaling [21]. In this regard, it was reported that α-trinositol (D-myo-inositol-1,2,6-trisphosphate), a synthetic inositol phosphate analog with significant anti-inflammatory properties [22] showed good safety and efficacy in decreasing IR in PCOS patients by acting as an insulin-sensitizer [23–26].
’Regulation of reactive oxygen species biosynthetic process’ was another GO term enriched in our PCLMN. Common features of oxidative stress in PCOS patients include increased lipid peroxidation and protein hydroxyl content and decreased antioxidant capacity [27]. These changes, paralleled and induced by exacerbated cellular production of reactive oxygen species, contribute to a proinflammatory state conducive to IR and hyperandrogenism [28].
Several immune-related biological processes and pathways, including ‘T cell selection’, ‘T cell differentiation in thymus’, ‘regulation of cytokine production involved in immune response’, ‘regulation of lymphocyte apoptotic process’, and ‘IL1 and megakaryocytes in obesity’ were also enriched in the PCLMN. Ovulation disorder is one of the common clinical manifestations and the main cause of infertility in women affected by PCOS. Several leukocyte subsets participate in the regulation of follicle growth and maturation. Lymphocytes have a promoting role on follicle growth, ovulation, and luteinization; however, abnormal inflammatory conditions can cause lymphocytes to oversecrete pro-inflammatory factors that trigger cytotoxicity and induce follicular apoptosis. This in turn leads to phagocytosis by macrophages, follicular development stagnation, and ovulation failure [29].
We further performed network topology analysis and run the MCODE clustering algorithm to identify crucial lncRNAs with central topology features in the PCLMN. We identified 3 lncRNAs (LINC00667, AC073172.1, and H19) with potentially crucial roles in regulating key pathways in PCOS. The NF-kappa B signaling pathway was enriched in the ceRNA sub-network of LINC00667. NF-kB is an important TF that initiates and regulates the expression of various inflammatory mediators, modulates the development of inflammatory responses, and is also closely involved in IR [30, 31]. Suggesting yet another link between PCOS and chronic inflammation, increased serum NF-κB levels have been reported in PCOS patients [32]. Indeed, the main functional module identified in our PCLMN contained, along with LINC00667 and H19, BCL11B, a TF expressed by all T-cell subsets that was shown to enhance TCR/CD28-triggered NF-kB activation [33].
Another biological process enriched in PCLMN genes was ‘negative regulation of apoptotic process’. Apoptosis is the mechanism responsible for follicular atresia and the basis for the cyclical growth and regression of human ovarian follicles [34]. Abnormal regulation of apoptosis has been suggested as a central mechanism of implantation failure by a previous ceRNA network analysis [35]. A recent study suggested also that reduced serum levels of caspase 9, an apoptotic marker, might be correlated to the pathogenesis of PCOS [36]. Moreover, compared with normally ovulating women, reduced expression of caspases 3, 8, and 9 and overexpression of the anti-apoptotic regulators cIAP-2 and Hsp27 were detected in oocytes of women with PCOS [37, 38].
The GO term ’negative regulation of tumor necrosis factor production’ was enriched in the ceRNA sub-network of H19. Tumor necrosis factor-α (TNF-α) is a crucial mediator of IR through its capacity to weaken the tyrosine kinase activity of the insulin receptor [19]. Expression levels of both TNF-α and TNF-α receptor 2 are upregulated during obesity and correlate with hyperinsulinemia [39]. Indeed, serum levels of TNF-α were found to be significantly elevated in PCOS patients with BMI > 27, compared to matched healthy controls [40].
The BP GO terms ‘cellular response to lipopolysaccharide’, and ‘Toll-like receptor signaling pathway’ were enriched in the ceRNA sub-network of AC073172.1. Lipopolysaccharide (LPS) is the main pathogenic component of gram-negative bacteria and a powerful inducer of inflammatory responses. LPS binds to Toll-like receptor 4 (TLR4) to activate NF-kB, promoting the transcription of TNF-α, IL-1β, and IL-6 [41]. Notably, saturated fatty acids can also activate TLR2 and TLR4 signaling and lead to IR [42]. In this regard, a recent study found that lipid-induced LPS-mediated inflammation through TLR4 is associated with obesity and worsened by PCOS [43]. On the other hand, recent studies suggested that single-nucleotide polymorphisms identified in TLR2 and TLR4 genes in PCOS patients might influence metabolic variables and increase susceptibility to PCOS [44, 45].
Finally, we identified several TFs that might regulate the expression of the 3 key lncRNAs identified in our PCMLN. Four TFs (ER-alpha, PRA, PRB, and AR) involved in sex steroid functions were predicted to have high binding affinity to transcriptional control elements in the DNA sequences of the 3 lncRNAs. Sex steroid hormones play a fundamental role in fertility by regulating reproductive function in the ovary through specific nuclear receptors. Several studies indicated that lncRNAs can synergize with TFs to regulate sex steroid functions. For example, a regulatory circuit composed of androgen receptors and PlncRNA-1 was shown to promote prostate cancer [46]. Another lncRNA, HOTAIR, is a direct target of ER-mediated transcriptional repression and its upregulation promotes ligand-independent ER activities [47]. Estrogens act by binding to estrogen receptor-alpha (ERα) and beta (ERβ) [48], both of which are expressed in the human ovary [49]. Studies in mice showed that ERα knockout leads to a PCOS phenotype defined by the presence of polycystic ovaries and increased luteinizing hormone (LH) levels, present impaired glucose tolerance, and eventually develop IR [50, 51]. In these mice, such condition was aggravated by accumulation of bioactive lipid intermediates and inflammation became more severe upon high-fat feeding, suggesting that ERα is also essential to protect against tissue inflammation [52]. The progesterone receptor (PR) and the androgen receptor (AR) belong to the nuclear hormone receptor family, which are associated with the regulation of eukaryotic gene expression and influence cellular proliferation and differentiation in target tissues. A previous developmental study on PRA- and PRB-knockout mice suggested that PRA is necessary for ovulation [53]. PRA and PRB are expressed in human granulosa cells and were found to be significantly downregulated in PCOS patients [54]. Human in vitro and in vivo studies found that the number of CAG repeats, which encode for an amino-acid sequence in the receptor’s transactivation domain, associates inversely the AR activity [55]. It was reported that short CAG repeats were more frequent in PCOS, possibly eliciting androgenic effects, while longer CAG repeats were more recurrent in the control group, involving probably a protective effect [56].
In summary, our study unmasked a regulatory network involving novel interactions between lncRNAs, sex steroids, and TFs with potential influence in the occurrence and development of PCOS. Further functional studies are warranted to validate the present findings and to explore their therapeutic implications.
Materials and Methods
miRNA-lncRNA and miRNA-mRNA interaction data
Predicted lncRNA-miRNA pairs were obtained from starBase V3.0 [57] and lncRNASNP2 [58] databases. The latter was also used for predicting potential miRNAs targeted by DELs. Predicted miRNA-mRNA pairs were in turn obtained from miRTarBase [59] and starbase V3.0. Next, the global triple network was constructed as the background network to identify gene interactions.
Gene expression profile
Gene expression data from the GSE95728 dataset, based on the GPL16956 platform (Agilent-045997 Arraystar human lncRNA microarray V3), was downloaded from the Gene Expression Omnibus (GEO) database. GSE95728 included lncRNA and mRNA expression profiles from granulosa cells from seven PCOS patients and seven controls (women with normal ovarian reserve).
Differential gene expression analysis and probe re-annotation
Expression data was imported into R-studio (https://rstudio.com) and normalized with the RMA algorithm [60]. Bioconductor’s limma package was applied to identify DEGs between control and PCOS samples, based on |log2(fold change)| > 2 and adjusted P <0.05. Probe annotation data provided by Agilent were aligned to both human long non-coding and protein-coding transcript sequences retrieved from the GENCODE database by running the SeqMap program [61]. Alignment results were filtered as follows: 1) probes matched to one transcript were retained, whereas probes simultaneously matched to protein-coding and long non-coding transcripts were deleted. Two sets of probes-transcripts pairs were finally obtained; 2) for each probe–transcript pair, probes matched to more than one transcript were removed; 3) transcripts were finally selected if they matched at least three probes.
Hierarchical clustering
Genes with comparable expression profiles were grouped by unsupervised hierarchical clustering using Multiple Experiment Viewer (MeV V4.9) software. The data were normalized and processed using Pearson’s correlation as distance metric and average linkage clustering algorithm.
Enrichment analysis
Functional analysis of DEGs in the PCLMN and in the ceRNA sub-networks was performed using Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses using DAVID V6.8 and Cytoscape V3.8.0 with the ClueGo V2.3.7 plug-in. P values were calculated by two-sided hypergeometric test and Benjamini-Hochberg adjustment. GO terms and KEGG pathways with p <0.05 were considered statistically significant. Results were visualized with Cytoscape.
Topological analysis and selection of key lncRNAs
We performed topological analysis of DELs DEMs to identify the central nodes of the PCLMN network. Topological parameters, including closeness, betweenness, and degree, were assessed using Cytoscape with the CentiScaPe V2.2 plug-in. The top 10 genes ranked by each measure were compared and those overlapping across all three topological parameters were chosen as key genes for further ceRNA analysis.
Subcellular localization analysis
We predicted the subcellular localization of key lncRNAs via LncLocator [62], a public platform based on a stacked ensemble classifier. The lncRNA subcellular localization information used in LncLocator, backed by experimental evidence, was extracted from RNAlocate database (http://www.rnasociety.org/rnalocate). Furthermore, we used iLoc-lncRNA, a sequence-based tool for subcellular localization prediction, to validate the above results [63]. The sequences of the key lncRNAs were downloaded from UCSC Genome Browser database.
Construction of ceRNA sub-networks
We extracted all the key lncRNAs and their nearest mRNA neighbors from transcript clusters in the PCLMN. miRNAs associated with lncRNA/mRNA pairs were also extracted from the global triple network and used to identify candidate lncRNA-miRNA-mRNA triplets. Then we constructed the ceRNA networks based on the ceRNA theory and visualized them with Cytoscape software.
Identification of putative transcription factors
Transcription factors (TFs) can bind to the DNA regulatory elements of lncRNAs to activate or inhibit their expression. To assess potential linkages between the key lncRNAs, we identified the TFs that might regulate them. Promoters were defined as DNA regions within 2 kb upstream of lncRNA transcriptional start sites. We used PROMO V3.0.2 software with maximum matrix dissimilarity rate < 3 to scan the predicted TFs [64, 65], and then used a Venn diagram to identify overlapping TFs targeting all the key lncRNAs.
Identification of functional modules in the PCLMN
LncRNAs participate in biological processes as members of functional modules encompassing other genes. To explore lncRNA-related functional modules in our network, we run Cytoscape with the MCODE plug-in applying the “Haircut,” “Fluff,” and Node Score Cutoff: 0.2 options.
Author Contributions
Y.M. conducted all the analyses, interpreted data, and wrote the manuscript. J.Z., L.M., and Y.C. provided critical revision of the final manuscript. All authors read and approved the final manuscript.
Acknowledgments
The authors thank members of their laboratory and collaborators for their research work.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
The study was supported by the National Natural Science Foundation of China (NSFC) (grant no. 82071649). The funders had no role in the study design, data collection and analysis, or decision to submit the article for publication.
References
- 1. Shi Y, Zhao H, Shi Y, Cao Y, Yang D, Li Z, Zhang B, Liang X, Li T, Chen J, Shen J, Zhao J, You L, et al. Genome-wide association study identifies eight new risk loci for polycystic ovary syndrome. Nat Genet. 2012; 44:1020–25. https://doi.org/10.1038/ng.2384 [PubMed]
- 2. Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome. Fertil Steril. 2004; 81:19–25. https://doi.org/10.1016/j.fertnstert.2003.10.004 [PubMed]
- 3. Christakou C, Diamanti-Kandarakis E. Polycystic ovary syndrome--phenotypes and diagnosis. Scand J Clin Lab Invest Suppl. 2014; 244:18–22 https://doi.org/10.3109/00365513.2014.936675 [PubMed]
- 4. Ranasinha S, Joham AE, Norman RJ, Shaw JE, Zoungas S, Boyle J, Moran L, Teede HJ. The association between Polycystic Ovary Syndrome (PCOS) and metabolic syndrome: a statistical modelling approach. Clin Endocrinol (Oxf). 2015; 83:879–87. https://doi.org/10.1111/cen.12830 [PubMed]
- 5. Kakoly NS, Khomami MB, Joham AE, Cooray SD, Misso ML, Norman RJ, Harrison CL, Ranasinha S, Teede HJ, Moran LJ. Ethnicity, obesity and the prevalence of impaired glucose tolerance and type 2 diabetes in PCOS: a systematic review and meta-regression. Hum Reprod Update. 2018; 24:455–67. https://doi.org/10.1093/humupd/dmy007 [PubMed]
- 6. Bilal M, Haseeb A, Rehman A. Relationship of polycystic ovarian syndrome with cardiovascular risk factors. Diabetes Metab Syndr. 2018; 12:375–80. https://doi.org/10.1016/j.dsx.2018.01.006 [PubMed]
- 7. Shang K, Jia X, Qiao J, Kang J, Guan Y. Endometrial abnormality in women with polycystic ovary syndrome. Reprod Sci. 2012; 19:674–83. https://doi.org/10.1177/1933719111430993 [PubMed]
- 8. Mattick JS. Non-coding RNAs: the architects of eukaryotic complexity. EMBO Rep. 2001; 2:986–91. https://doi.org/10.1093/embo-reports/kve230 [PubMed]
- 9. An S, Song JJ. The coded functions of noncoding RNAs for gene regulation. Mol Cells. 2011; 31:491–96. https://doi.org/10.1007/s10059-011-1004-8 [PubMed]
- 10. Guttman M, Amit I, Garber M, French C, Lin MF, Feldser D, Huarte M, Zuk O, Carey BW, Cassady JP, Cabili MN, Jaenisch R, Mikkelsen TS, et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature. 2009; 458:223–27. https://doi.org/10.1038/nature07672 [PubMed]
- 11. Liu YD, Li Y, Feng SX, Ye DS, Chen X, Zhou XY, Chen SL. Long noncoding RNAs: potential regulators involved in the pathogenesis of polycystic ovary syndrome. Endocrinology. 2017; 158:3890–99. https://doi.org/10.1210/en.2017-00605 [PubMed]
- 12. Zhao J, Huang J, Geng X, Chu W, Li S, Chen ZJ, Du Y. Polycystic ovary syndrome: novel and hub lncRNAs in the insulin resistance-associated lncRNA-mRNA network. Front Genet. 2019; 10:772. https://doi.org/10.3389/fgene.2019.00772 [PubMed]
- 13. Salmena L, Poliseno L, Tay Y, Kats L, Pandolfi PP. A ceRNA hypothesis: the rosetta stone of a hidden RNA language? Cell. 2011; 146:353–58. https://doi.org/10.1016/j.cell.2011.07.014 [PubMed]
- 14. Jin X, Guan Y, Sheng H, Liu Y. Crosstalk in competing endogenous RNA network reveals the complex molecular mechanism underlying lung cancer. Oncotarget. 2017; 8:91270–80. https://doi.org/10.18632/oncotarget.20441 [PubMed]
- 15. Song C, Zhang J, Liu Y, Pan H, Qi HP, Cao YG, Zhao JM, Li S, Guo J, Sun HL, Li CQ. Construction and analysis of cardiac hypertrophy-associated lncRNA-mRNA network based on competitive endogenous RNA reveal functional lncRNAs in cardiac hypertrophy. Oncotarget. 2016; 7:10827–40. https://doi.org/10.18632/oncotarget.7312 [PubMed]
- 16. Feng C, Shen JM, Lv PP, Jin M, Wang LQ, Rao JP, Feng L. Construction of implantation failure related lncRNA-mRNA network and identification of lncRNA biomarkers for predicting endometrial receptivity. Int J Biol Sci. 2018; 14:1361–77. https://doi.org/10.7150/ijbs.25081 [PubMed]
- 17. Tarkun I, Arslan BC, Cantürk Z, Türemen E, Sahin T, Duman C. Endothelial dysfunction in young women with polycystic ovary syndrome: relationship with insulin resistance and low-grade chronic inflammation. J Clin Endocrinol Metab. 2004; 89:5592–96. https://doi.org/10.1210/jc.2004-0751 [PubMed]
- 18. Blum CA, Müller B, Huber P, Kraenzlin M, Schindler C, De Geyter C, Keller U, Puder JJ. Low-grade inflammation and estimates of insulin resistance during the menstrual cycle in lean and overweight women. J Clin Endocrinol Metab. 2005; 90:3230–35. https://doi.org/10.1210/jc.2005-0231 [PubMed]
- 19. Hotamisligil GS, Peraldi P, Budavari A, Ellis R, White MF, Spiegelman BM. IRS-1-mediated inhibition of insulin receptor tyrosine kinase activity in TNF-alpha- and obesity-induced insulin resistance. Science. 1996; 271:665–68. https://doi.org/10.1126/science.271.5249.665 [PubMed]
- 20. Sarray S, Almawi WY. Levels of CD40L and other inflammatory biomarkers in obese and non-obese women with polycystic ovary syndrome. Am J Reprod Immunol. 2016; 76:285–91. https://doi.org/10.1111/aji.12549 [PubMed]
- 21. Deligeoroglou E, Vrachnis N, Athanasopoulos N, Iliodromiti Z, Sifakis S, Iliodromiti S, Siristatidis C, Creatsas G. Mediators of chronic inflammation in polycystic ovarian syndrome. Gynecol Endocrinol. 2012; 28:974–78. https://doi.org/10.3109/09513590.2012.683082 [PubMed]
- 22. Claxson A, Morris C, Blake D, Sirén M, Halliwell B, Gustafsson T, Löfkvist B, Bergelin I. The anti-inflammatory effects of D-myo-inositol-1.2.6-trisphosphate (PP56) on animal models of inflammation. Agents Actions. 1990; 29:68–70. https://doi.org/10.1007/BF01964724 [PubMed]
- 23. Facchinetti F, Unfer V, Dewailly D, Kamenov ZA, Diamanti-Kandarakis E, Laganà AS, Nestler JE, Soulage CO, and Group of ‘Inositol in PCOS and Reproduction’. Inositols in polycystic ovary syndrome: an overview on the advances. Trends Endocrinol Metab. 2020; 31:435–47. https://doi.org/10.1016/j.tem.2020.02.002 [PubMed]
- 24. Laganà AS, Garzon S, Casarin J, Franchi M, Ghezzi F. Inositol in polycystic ovary syndrome: restoring fertility through a pathophysiology-based approach. Trends Endocrinol Metab. 2018; 29:768–80. https://doi.org/10.1016/j.tem.2018.09.001 [PubMed]
- 25. Facchinetti F, Appetecchia M, Aragona C, Bevilacqua A, Bezerra Espinola MS, Bizzarri M, D’Anna R, Dewailly D, Diamanti-Kandarakis E, Hernández Marín I, Kamenov ZA, Kandaraki E, Laganà AS, et al. Experts’ opinion on inositols in treating polycystic ovary syndrome and non-insulin dependent diabetes mellitus: a further help for human reproduction and beyond. Expert Opin Drug Metab Toxicol. 2020; 16:255–74. https://doi.org/10.1080/17425255.2020.1737675 [PubMed]
- 26. Laganà AS, Rossetti P, Sapia F, Chiofalo B, Buscema M, Valenti G, Rapisarda AM, Vitale SG. Evidence-based and patient-oriented inositol treatment in polycystic ovary syndrome: changing the perspective of the disease. Int J Endocrinol Metab. 2017; 15:e43695. https://doi.org/10.5812/ijem.43695 [PubMed]
- 27. Kuşçu NK, Var A. Oxidative stress but not endothelial dysfunction exists in non-obese, young group of patients with polycystic ovary syndrome. Acta Obstet Gynecol Scand. 2009; 88:612–17. https://doi.org/10.1080/00016340902859315 [PubMed]
- 28. González F, Rote NS, Minium J, Kirwan JP. Reactive oxygen species-induced oxidative stress in the development of insulin resistance and hyperandrogenism in polycystic ovary syndrome. J Clin Endocrinol Metab. 2006; 91:336–40. https://doi.org/10.1210/jc.2005-1696 [PubMed]
- 29. Walusimbi SS, Pate JL. Physiology and endocrinology symposium: role of immune cells in the corpus luteum. J Anim Sci. 2013; 91:1650–59. https://doi.org/10.2527/jas.2012-6179 [PubMed]
- 30. Yamamoto Y, Gaynor RB. IkappaB kinases: key regulators of the NF-kappaB pathway. Trends Biochem Sci. 2004; 29:72–79. https://doi.org/10.1016/j.tibs.2003.12.003 [PubMed]
- 31. Lan W, Petznick A, Heryati S, Rifada M, Tong L. Nuclear factor-κB: central regulator in ocular surface inflammation and diseases. Ocul Surf. 2012; 10:137–48. https://doi.org/10.1016/j.jtos.2012.04.001 [PubMed]
- 32. Liu M, Gao J, Zhang Y, Li P, Wang H, Ren X, Li C. Serum levels of TSP-1, NF-κB and TGF-β1 in polycystic ovarian syndrome (PCOS) patients in northern China suggest PCOS is associated with chronic inflammation. Clin Endocrinol (Oxf). 2015; 83:913–22. https://doi.org/10.1111/cen.12951 [PubMed]
- 33. Cismasiu VB, Duque J, Paskaleva E, Califano D, Ghanta S, Young HA, Avram D. BCL11B enhances TCR/CD28-triggered NF-kappaB activation through up-regulation of Cot kinase gene expression in T-lymphocytes. Biochem J. 2009; 417:457–66. https://doi.org/10.1042/BJ20080925 [PubMed]
- 34. Slot KA, de Boer-Brouwer M, Houweling M, Vaandrager AB, Dorrington JH, Teerds KJ. Luteinizing hormone inhibits Fas-induced apoptosis in ovarian surface epithelial cell lines. J Endocrinol. 2006; 188:227–39. https://doi.org/10.1677/joe.1.06087 [PubMed]
- 35. Maciel GA, Baracat EC, Benda JA, Markham SM, Hensinger K, Chang RJ, Erickson GF. Stockpiling of transitional and classic primary follicles in ovaries of women with polycystic ovary syndrome. J Clin Endocrinol Metab. 2004; 89:5321–27. https://doi.org/10.1210/jc.2004-0643 [PubMed]
- 36. Uyanikoglu H, Sabuncu T, Dursun H, Sezen H, Aksoy N. Circulating levels of apoptotic markers and oxidative stress parameters in women with polycystic ovary syndrome: a case-controlled descriptive study. Biomarkers. 2017; 22:643–47. https://doi.org/10.1080/1354750X.2016.1265004 [PubMed]
- 37. Cai L, Ma X, Liu S, Liu J, Wang W, Cui Y, Ding W, Mao Y, Chen H, Huang J, Zhou Z, Liu J. Effects of upregulation of Hsp27 expression on oocyte development and maturation derived from polycystic ovary syndrome. PLoS One. 2013; 8:e83402. https://doi.org/10.1371/journal.pone.0083402 [PubMed]
- 38. Das M, Djahanbakhch O, Hacihanefioglu B, Saridogan E, Ikram M, Ghali L, Raveendran M, Storey A. Granulosa cell survival and proliferation are altered in polycystic ovary syndrome. J Clin Endocrinol Metab. 2008; 93:881–87. https://doi.org/10.1210/jc.2007-1650 [PubMed]
- 39. Hotamisligil GS, Arner P, Atkinson RL, Spiegelman BM. Differential regulation of the p80 tumor necrosis factor receptor in human obesity and insulin resistance. Diabetes. 1997; 46:451–55. https://doi.org/10.2337/diab.46.3.451 [PubMed]
- 40. Samy N, Hashim M, Sayed M, Said M. Clinical significance of inflammatory markers in polycystic ovary syndrome: their relationship to insulin resistance and body mass index. Dis Markers. 2009; 26:163–70. https://doi.org/10.3233/DMA-2009-0627 [PubMed]
- 41. Fitzgerald-Bocarsly P. Natural interferon-alpha producing cells: the plasmacytoid dendritic cells. Biotechniques. 2002; (Suppl): 16-20, 22, 24-29. https://doi.org/10.2144/Oct0203 [PubMed]
- 42. Yin J, Peng Y, Wu J, Wang Y, Yao L. Toll-like receptor 2/4 links to free fatty acid-induced inflammation and β-cell dysfunction. J Leukoc Biol. 2014; 95:47–52. https://doi.org/10.1189/jlb.0313143 [PubMed]
- 43. González F, Considine RV, Abdelhadi OA, Acton AJ. Saturated Fat Ingestion Promotes Lipopolysaccharide-Mediated Inflammation and Insulin Resistance in Polycystic Ovary Syndrome. J Clin Endocrinol Metab. 2019; 104:934–46. https://doi.org/10.1210/jc.2018-01143 [PubMed]
- 44. Allauddin N, Rozati R, Jaideep M. Single Nucleotide Polymorphisms in CD14 and Toll-like Receptor 4 Genes in Patients with Polycystic Ovarian Syndrome. Journal of SAFOG. 2017; 9:304–07. https://doi.org/10.5005/jp-journals-10006-1518
- 45. Ojeda-Ojeda M, Martínez-García MÁ, Alpañés M, Luque-Ramírez M, Escobar-Morreale HF. Association of TLR2 S450S and ICAM1 K469E polymorphisms with polycystic ovary syndrome (PCOS) and obesity. J Reprod Immunol. 2016; 113:9–15. https://doi.org/10.1016/j.jri.2015.09.072 [PubMed]
- 46. Fang Z, Xu C, Li Y, Cai X, Ren S, Liu H, Wang Y, Wang F, Chen R, Qu M, Wang Y, Zhu Y, Zhang W, et al. A feed-forward regulatory loop between androgen receptor and PlncRNA-1 promotes prostate cancer progression. Cancer Lett. 2016; 374:62–74. https://doi.org/10.1016/j.canlet.2016.01.033 [PubMed]
- 47. Xue X, Yang YA, Zhang A, Fong KW, Kim J, Song B, Li S, Zhao JC, Yu J. LncRNA HOTAIR enhances ER signaling and confers tamoxifen resistance in breast cancer. Oncogene. 2016; 35:2746–55. https://doi.org/10.1038/onc.2015.340 [PubMed]
- 48. Hillisch A, Peters O, Kosemund D, Müller G, Walter A, Schneider B, Reddersen G, Elger W, Fritzemeier KH. Dissecting physiological roles of estrogen receptor alpha and beta with potent selective ligands from structure-based design. Mol Endocrinol. 2004; 18:1599–609. https://doi.org/10.1210/me.2004-0050 [PubMed]
- 49. Hegele-Hartung C, Siebel P, Peters O, Kosemund D, Müller G, Hillisch A, Walter A, Kraetzschmar J, Fritzemeier KH. Impact of isotype-selective estrogen receptor agonists on ovarian function. Proc Natl Acad Sci USA. 2004; 101:5129–34. https://doi.org/10.1073/pnas.0306720101 [PubMed]
- 50. Schomberg DW, Couse JF, Mukherjee A, Lubahn DB, Sar M, Mayo KE, Korach KS. Targeted disruption of the estrogen receptor-alpha gene in female mice: characterization of ovarian responses and phenotype in the adult. Endocrinology. 1999; 140:2733–44. https://doi.org/10.1210/endo.140.6.6823 [PubMed]
- 51. Heine PA, Taylor JA, Iwamoto GA, Lubahn DB, Cooke PS. Increased adipose tissue in male and female estrogen receptor-alpha knockout mice. Proc Natl Acad Sci USA. 2000; 97:12729–34. https://doi.org/10.1073/pnas.97.23.12729 [PubMed]
- 52. Ribas V, Nguyen MT, Henstridge DC, Nguyen AK, Beaven SW, Watt MJ, Hevener AL. Impaired oxidative metabolism and inflammation are associated with insulin resistance in ERalpha-deficient mice. Am J Physiol Endocrinol Metab. 2010; 298:E304–19. https://doi.org/10.1152/ajpendo.00504.2009 [PubMed]
- 53. Shimada M, Yamashita Y, Ito J, Okazaki T, Kawahata K, Nishibori M. Expression of two progesterone receptor isoforms in cumulus cells and their roles during meiotic resumption of porcine oocytes. J Mol Endocrinol. 2004; 33:209–25. https://doi.org/10.1677/jme.0.0330209 [PubMed]
- 54. Artimani T, Saidijam M, Aflatoonian R, Amiri I, Ashrafi M, Shabab N, Mohammadpour N, Mehdizadeh M. Estrogen and progesterone receptor subtype expression in granulosa cells from women with polycystic ovary syndrome. Gynecol Endocrinol. 2015; 31:379–83. https://doi.org/10.3109/09513590.2014.1001733 [PubMed]
- 55. Zitzmann M, Depenbusch M, Gromoll J, Nieschlag E. X-chromosome inactivation patterns and androgen receptor functionality influence phenotype and social characteristics as well as pharmacogenetics of testosterone therapy in klinefelter patients. J Clin Endocrinol Metab. 2004; 89:6208–17. https://doi.org/10.1210/jc.2004-1424 [PubMed]
- 56. Schüring AN, Welp A, Gromoll J, Zitzmann M, Sonntag B, Nieschlag E, Greb RR, Kiesel L. Role of the CAG repeat polymorphism of the androgen receptor gene in polycystic ovary syndrome (PCOS). Exp Clin Endocrinol Diabetes. 2012; 120:73–79. https://doi.org/10.1055/s-0031-1291343 [PubMed]
- 57. Yang JH, Li JH, Shao P, Zhou H, Chen YQ, Qu LH. starBase: a database for exploring microRNA-mRNA interaction maps from argonaute CLIP-seq and degradome-seq data. Nucleic Acids Res. 2011; 39:D202–09. https://doi.org/10.1093/nar/gkq1056 [PubMed]
- 58. Gong J, Liu W, Zhang J, Miao X, Guo AY. lncRNASNP: a database of SNPs in lncRNAs and their potential functions in human and mouse. Nucleic Acids Res. 2015; 43:D181–86. https://doi.org/10.1093/nar/gku1000 [PubMed]
- 59. Chou CH, Shrestha S, Yang CD, Chang NW, Lin YL, Liao KW, Huang WC, Sun TH, Tu SJ, Lee WH, Chiew MY, Tai CS, Wei TY, et al. miRTarBase update 2018: a resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2018; 46:D296–302. https://doi.org/10.1093/nar/gkx1067 [PubMed]
- 60. Harbron C, Chang KM, South MC. RefPlus: an R package extending the RMA algorithm. Bioinformatics. 2007; 23:2493–94. https://doi.org/10.1093/bioinformatics/btm357 [PubMed]
- 61. Jiang H, Wong WH. SeqMap: mapping massive amount of oligonucleotides to the genome. Bioinformatics. 2008; 24:2395–96. https://doi.org/10.1093/bioinformatics/btn429 [PubMed]
- 62. Cao Z, Pan X, Yang Y, Huang Y, Shen HB. The lncLocator: a subcellular localization predictor for long non-coding RNAs based on a stacked ensemble classifier. Bioinformatics. 2018; 34:2185–94. https://doi.org/10.1093/bioinformatics/bty085 [PubMed]
- 63. Su ZD, Huang Y, Zhang ZY, Zhao YW, Wang D, Chen W, Chou KC, Lin H. iLoc-lncRNA: predict the subcellular location of lncRNAs by incorporating octamer composition into general PseKNC. Bioinformatics. 2018; 34:4196–204. https://doi.org/10.1093/bioinformatics/bty508 [PubMed]
- 64. Laguna-Bercero MA, Larrea A, Merino RI, Peña JI, Orera VM. Crystallography and thermal stability of textured Co-YSZ cermets from eutectic precursors. Journal of the European Ceramic Society. 2008; 28:2325–29. https://doi.org/10.1016/j.jeurceramsoc.2008.01.019
- 65. Farré D, Roset R, Huerta M, Adsuara JE, Roselló L, Albà MM, Messeguer X. Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res. 2003; 31:3651–53. https://doi.org/10.1093/nar/gkg605 [PubMed]