



Introduction
Hypospadias is a common congenital genitourinary deformity characterized by a ventrally opened urethral meatus, chordee and hooded foreskin with ventral deficiency [1]. The incidence of hypospadias varies among populations, regions and ethnicities. Besides, there is an increasing trend of hypospadias in many countries. The mean prevalence was: Worldwide 20.9%, Asia 0.6–69%, Europe 18.6%, North America 34.2%, South America 5.2%, Africa 5.9%, and Australia 17.1–34.8% [2]. Hypospadias is caused by incomplete urethral fusion during gestational weeks 8 to 16 [3]. Hypospadias can be clinically divided into two subgroups according to the abnormal meatus location, namely, mild/moderate and severe hypospadias [4]. Despite rapid progress in surgical repair over recent decades, the etiology of hypospadias remains largely unknown [1, 5–7].
Genetic factors have been demonstrated to be a key element during the development of hypospadias [8, 9]. By case-control analysis, a genome-wide association study (GWAS) identified that a few genes cause genetic susceptibility, including AR, SRD5A2, ATF3 and BMP. [10] Moreover, it has been reported that some SNPs correlate with hypospadias, but most relationships have been found in small studies without consistent replication [11, 12]. For instance, GREM1 rs3743104 (NC_000015.9:g.33023985A>G) was identified by a GWAS [13] in a large European population (1,972 cases and 1,812 controls), though the results were not replicated by Chen et al. in a small Chinese population (197 cases and 933 controls) [14].
GREM1, which encodes a member of the bone morphogenic protein antagonist family, is located at 15q13.3, and it is reported to be closely related to a variety of malignant tumors [15]. It is worth mentioning that the epithelial-mesenchymal transition (EMT) in colon cancer cells can be suppressed by silencing of GREM1 via short hairpin RNA (shRNA), [16] and this process is considered an essential event inducing morphogenetic changes during the development of embryos [17]. Hence, to identify the association of GREM1 with hypospadias and the reproducibility of the link between GREM1 rs3743104 A>G previously derived in European GWASs of hypospadias, a replication investigation was conducted on ethnically homogeneous patients (577 cases and 654 controls) from the largest Southern Han Chinese population. The relationship between GREM1 and subtypes of hypospadias was studied. We also further demonstrated the functional relevance of GREM1 rs3743104 on Hypospadias and the mechanism of the SNP-related MicroRNAs(miRNA) involved in regulating GREM1 expression.
Materials and Methods
Study population
A total of 557 patients undergoing repair of hypospadias diagnosed by pediatric urologists at Guangzhou Women and Children’s Medical Center from 2016 to 2018 were collected, all of whom were Southern Han Chinese. Patients with only penile curvature were excluded from this study. The cases were divided into two subtypes according to the location of the urethral meatus: mild/moderate (281 patients with coronal or glanular or shaft penis hypospadias) and severe (247 patients with penoscrotal, scrotal and perineal hypospadias) hypospadias. A total of 29 cases could not be classified because the patients underwent the first surgical repairs at other hospitals. A total of 654 healthy children who underwent a physical examination in our hospital were recruited as the healthy control group, but children who had a history of hypospadias or any experience with hypospadias repair were excluded. Each patient was informed of the study purpose, and written consent was obtained from all participants or their parents/legal guardians. In addition, ethical approval was obtained from Guangzhou Women and Children’s Medical Center in China.
DNA extraction and genotyping
TIANamp Blood DNA kits (Catalog No. DP335-02; TIANGEN Biotech Co. Ltd., Beijing, China) were utilized to extract genomic DNA from venous blood samples of patients with hypospadias following the manufacturer’s instructions [18]. DNA concentration and purity were assessed using NanoPhotometer® N50 (Implen GmbH., Munich, Germany). An ABI-7900 real-time quantitative PCR instrument (Applied Biosystems, Foster City, CA, USA) was used to amplify genomic DNA, which was subsequently subjected to TaqMan genotyping for the GREM1 rs3743104 [19–21]. PCR reactions were run in duplicates in a total volume of 5 μl containing 1 μl (40 ng) genomic DNA, 0.04 μl TaqMan® SNP Genotyping Assays (Catalog No: 4351379, C_180222_20, Thermo Fisher, Grand Island, NY, USA), 2.5 μl TIANexact genotyping qPCR PreMix(Probe) (Catalog No. FP211-02; TIANGEN Biotech Co. Ltd., Beijing, China) and 1.46 μl DNase/RNase-free H2O with 98% concordance rate for genotype calls. Moreover, 10% of DNA samples were selected at random for another genotyping analysis. All of the replicated samples were entirely consistent to ensure that the data were accurate [22].
Bioinformatics analysis
To further investigate the potential impact of GREM1 rs3743104, we used TargetScan (http://www.targetscan.org) and MirSNP databases (http://bioinfo.bjmu.edu.cn/mirsnp/search/) to identify that rs3743104 at the 3′-UTR of GREM1 might contain a microRNA (miRNA) binding site. TargetScan is a web server that predicts biological targets of microRNAs (miRNAs) by searching for the presence of sites that match the seed region of each miRNA. MirSNP (http://bioinfo.bjmu.edu.cn/mirsnp/search/) allows for searching miRNA-related SNP sites.
The majority of GWAS risk loci are present in noncoding genomic regions with many gene regulatory signals, suggesting that most disease-causing SNPs exert their phenotypic effects by altering gene expression [23, 24]. GREM1 rs3743104 is in the 3’-UTR of GREM1 (noncoding genomic regions). Human cell population-based or tissue-based expression quantitative trait locus (eQTL) mapping studies, such as Genotype-Tissue Expression Project (GTEx), have provided an unprecedented opportunity to explore context-dependent regulatory patterns surrounding these loci [25]. To examine the eQTL SNP GREM1 rs3743104 in different tissues, we downloaded significant SNP-gene associations from GTEx V4 from the GTEx portal (http://www.gtexportal.org/), which contains eQTL mapping results for expression of different genotypes in different tissues.
Luciferase assays
The wild-type GREM1 dual reporting vector was obtained by the GREM1 PCR product and the psiCHECK™-2 (Catalog No: C8011, Promega, Madison, WI, USA) vector though digested with XhoI/NotI (XhoI: Catalog No: D1094A, TaKaRa Biotech Co. Ltd., Beijing, China; NotI: Catalog No: D1166A, TaKaRa Biotech Co. Ltd., Beijing, China). The GREM1 mutant sites vector was successfully constructed by PCR and DpnI enzyme (Catalog No: R6231, Promega, Madison, WI, USA). Sequences of the wild-type GREM1 dual reporting vector and the GREM1 mutant sites vector were shown in Supplementary Table 1. Each of the DNA products was separately subcloned into a luciferase reporter plasmid at the downstream of luciferase coding region (psiCHECK2; Promega, Madison, WI, USA) and the sequence validity was confirmed by Sanger sequencing. Next, 2×104 HEK293T cells were seeded into each well of 24-well plate and simultaneously transfected with respective construct, either miR-182, miR-212, miR-221, miR-3128 mimics and correspond MicroRNA inhibitor(Sequences of miR-182, miR-212, miR-221, miR-3128 mimics were shown in Supplementary Table 2) using lipofectamine 2000 (Catalog No: 116680119, Invitrogen, Carlsbad, CA, USA). After being cultured for 48 h, luciferase activity of the cells was measured with a Dual-Luciferase Reporter Assay Kit (Catalog No: E1910, Promega, Madison, WI, USA) according to the manufacturer’s protocol.
Statistical analysis
GraphPad Prism version 8 (GraphPad Software, Inc., La Jolla, CA, USA) and SAS 9.4 software (SAS Institute Inc., Cary, NC, USA) was used to perform statistical analyses in this study. A chi-squared goodness-of-fit test was conducted to examine the consistency between the genotype frequencies of the controls and Hardy–Weinberg equilibrium (HWE). The SNP was analyzed for association with the disease by comparing the risk of allele frequency (allelic test) in patients and controls as well as other tests using PLINK 1.9 (test of dominant and recessive models, genotype test) [26]. Association stratified by subphenotype was analyzed by comparing cases with a certain subphenotype with controls. A P-value of 0.05 was considered statistically significant.
Results
Association between GREM1 polymorphism rs3743104 and hypospadias susceptibility
In the present study, 525 of 557 cases and 594 of 654 controls were successfully genotyped. Table 1 shows the crude and adjusted odds/P values for the GREM1 polymorphism rs3743104 and hypospadias susceptibility. The frequency distribution of Hardy-Weinberg equilibrium (P_HWE=0.32). Comparison between Hypospadias patients and controls revealed that AA and AG genotypes were significantly higher in controls. In contrast, the GG genotype was more frequent in patients (65.71% in case versus 32.57% in controls, P=3.25×10-12/Adjusted P=1.99×10-12). Consistently, the G allele was more prevalent among patients (77.28%) compared to control group (55.12%). The GREM1 risk allele rs3743104[G] was associated with an increased risk of hypospadias. Our results also illustrated that GREM1 rs3743104[G] conferred increased risk to hypospadias under homozygote comparison based on different genotypic models, including dominant and recessive models (GG+AG versus AA: Adjusted OR=3.07, Adjusted P=1.32×10-06;GG versus AG+AA: Adjusted OR=4.43, Adjusted P=1.62×10-28).
Table 1. Association between GREM1 polymorphism rs3743104 and hypospadias susceptibility.
| Genotype | Case (n=557) | Control (N=654) | Crude OR (95% CI) | P | Adjusted OR (95% CI)1 | P1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| AA | 30 (53.86) | 86 (13.15) | Ref | 1.00 | Ref | 1.00 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| AG | 129 (23.16) | 295 (45.11) | 1.25(0.79-1.99) | 0.34 | 1.35(0.92-2.22) | 0.24 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GG | 366 (65.71) | 213 (32.57) | 4.93(3.15-7.72) | 3.25×10-12 | 5.30(3.33-8.43) | 1.99×10-12 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Genotypic model | NA | 1.91×10-27 | NA | 1.63×10-27 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Dominant model (GG+AG versus AA) | 495/30 | 508/86 | 2.79(1.81-4.31) | 3.44×10-06 | 3.07(1.95-4.83) | 1.32×10-06 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Recessive model (GG versus AG+AA) | 366/159 | 213/381 | 4.12(3.21-5.29) | 1.73×10-28 | 4.43(3.41-5.77) | 1.62×10-28 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Values are shown as number (%); AA, homozygous protective; AG, heterozygous; GG, homozygous risk for rs3743104; OR (95% CI), odds ratio and confidence interval; 1Adjusted for age with omission of the corresponding stratification factor; P value surpassing statistical significance (0.05) were boldfaced. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Subtype analysis of the relationship between GREM1 polymorphism rs3743104 and hypospadias risk
Hypospadias subtypes are classified according to the meatus location, including mild/moderate and severe hypospadias. The GREM1 risk allele rs3743104[G] notably increased the risk of mild/moderate and severe hypospadias (P<0.01, 0.28≤OR≤0.66, Table 2).
Table 2. Stratification analysis for the association between GREM1 risk allele rs3743104[G] and hypospadias susceptibility (by subgroup).
| rs3743104 | Case | Control | Case-only | Severe-only | Mild/moderate -only | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Severe | Mild/moderate | P | OR(CI95) | P | OR(CI95) | P | OR(CI95) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0.1533 | 0.2065 | 0.3931 | <0.01 | 0.66(0.47-0.92) | <0.01 | 0.28(0.21~0.36 | <0.01 | 0.40(0.32~0.51) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Case-only: All the patients with hypospadias. Severe-only: patients with penoscrotal, scrotal and perineal hypospadias. Mild/moderate-only: patients with coronal or glanular or shaft penis hypospadias. P value surpassing statistical significance (0.05) were boldfaced. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Regulatory element analysis of miRNAs targeting rs3743104
As rs3743104 is located at the 3′-UTR of GREM1, it is suspected that rs3743104 may affect the binding of miRNAs. Using the TargetScan and MirSNP databases, we identified that hsa-miR-182-5p, hsa-miR-212-5p, hsa-miR-221-5p and hsa-miR-3128 might bind to a site at the GREM1 3'-UTR (Table 3). In fact, based on TargetScan and MirSNP predictions, different alleles of rs3743104 would affect the binding of microRNAs to GREM1.
Table 3. rs3743104 regulates GREM1 though miRNAs.
| SNP | Gene | miRNA | Effect | Allele | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs3743104 | GREM1 | hsa-miR-182-5p | create | G/A | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| hsa-miR-212-5p | create | G/A | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| hsa-miR-221-5p | break | G/A | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| hsa-miR-3128 | break | G/A | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| The microRNA (miRNA) binding site rs3743104 in the 3′ UTR of GREM1 were predicted by TargetScan (http://www.targetscan.org) and MirSNP databases (http://bioinfo.bjmu.edu.cn/mirsnp/search/). Create means that the occurrence of the SNP leads to a miRNA binding site in the mRNA. Break means that the occurrence of the SNP causes the loss of miRNA binding site. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
GTEx eQTL analysis of rs3743104 and GREM1 expression
Loci in which expression level is influenced by specific genetic variation are termed expression quantitative trait loci (eQTL), which refers to genetic variants that affect expression in an allele-specific manner, with implications on an underlying mechanism [27, 28]. We sought to illustrate SNP associations with expression of GREM1 in different tissues, and GTEx eQTL data revealed more associations of different genotypes of eQTL SNP rs3743104 and GREM1 targets in pituitary tissues (Figure 1). Compared with the GREM1 rs3743104 GG genotype, the AG/AA genotype resulted in reduced expression of GREM1.
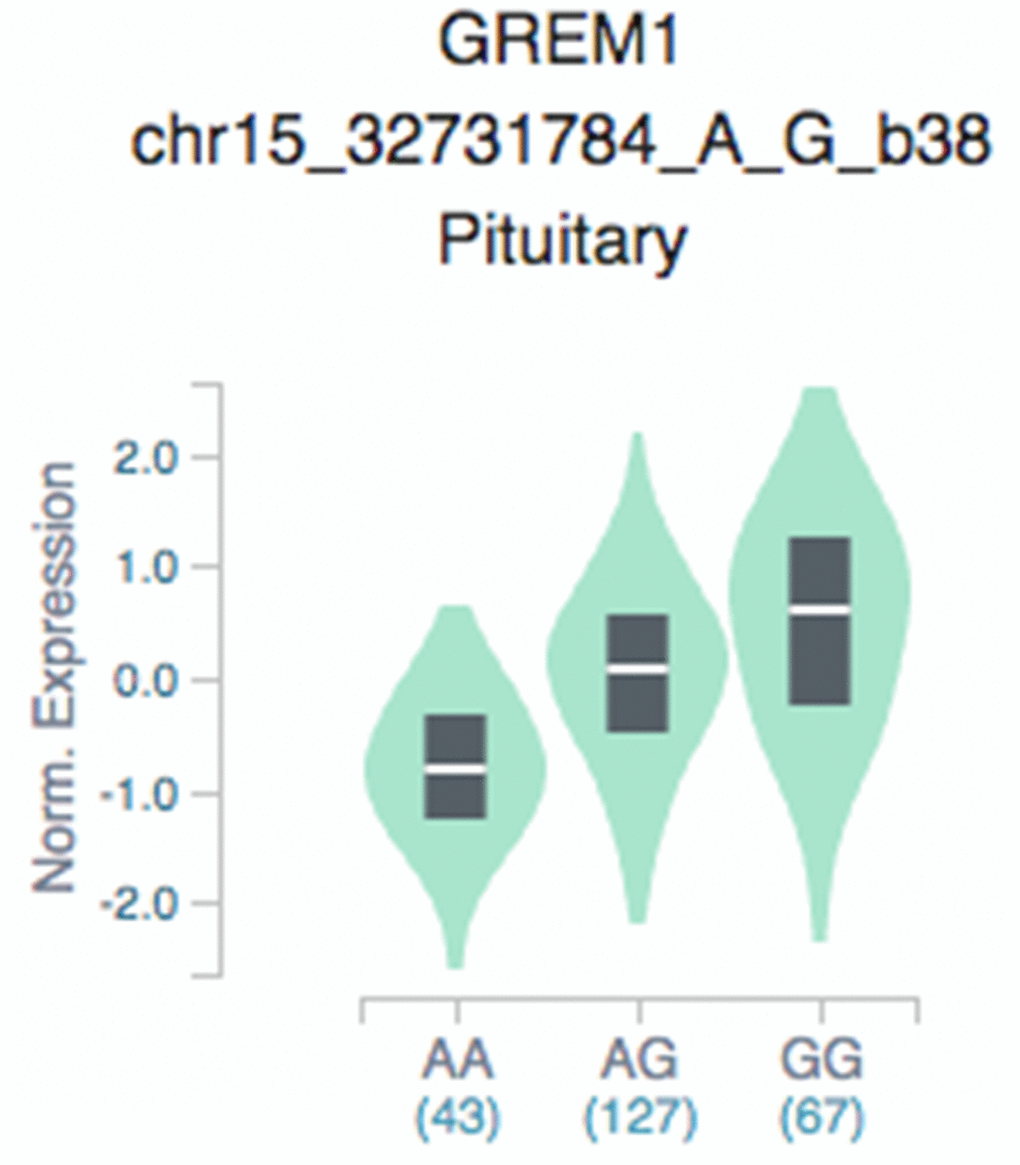
Figure 1. GTEx eQTL analysis of rs3743104 and GREM1 expression in pituitary tissues. GTEx: Genotype-Tissue Expression, GREM1 rs3743104 (chr15:33023985;GRCh37/hg19, A/G); AA, homozygous protective; AG, heterozygous; GG, homozygous risk for rs3743104.
Hsa-miR-182-5p targeted rs3743104
To further investigate how rs3743104 regulates upstream GREM1, we identified that hsa-miR-182-5p, hsa-miR-212-5p, hsa-miR-221-5p and hsa-miR-3128 might bind to a site at the GREM1 3'-UTR (Table 3). We identified that hsa-miR-182-5p might target a miRNA binding site at the 3′-UTR of RPA1, using TargetScan. Luciferase reporter assays showed that the luciferase activity of GREM1 3′-UTR was significantly reduced in the 293T cells transfected with miR-182 mimics compared to the control group in a dose-dependent manner (Figure 2). The data for Figure 2 were shown in Supplementary Table 3.
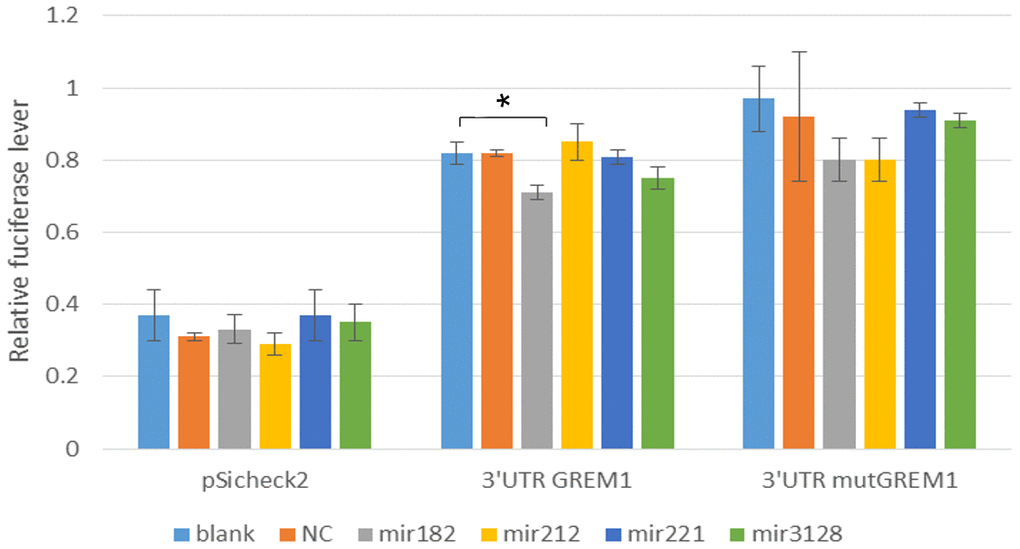
Figure 2. Luciferase activities in HEK293T cells transfected with respective construct, either miR-182, miR-212, miR-221, miR-3128 mimics and correspond MicroRNA inhibitor. NC: Negative control. Comparing each group with the blank group, *:P=0.0478, P<0.05 significant difference.
Discussion
Hypospadias is a common congenital urethral anomaly. In previous studies, [13] it was confirmed that SNP rs3743104 is associated with hypospadias susceptibility. Kojima et al. [29] replicated 17 SNPs of HAAO identified by European GWASs [13] and showed that only one SNP was strongly associated with hypospadias susceptibility in the Japanese population, indicating different allele frequencies of causative variants for hypospadias among different ethnic groups. However, according to a replication study, rs3743104 had no relationship with hypospadias susceptibility in a small Chinese population. Comparing to the previous GWAS study, the recruited population and region were different. Although the populations of the previous Chinese GWAS study and our study belonged to the Han population, the distribution of the population was different from that our population mainly located in the Southern area of China. Except for genetic factors, the environmental factor is also contributing to hypospadias. That may be the reason why the result was different from the research of Chen et al. [14] Thus, another replication study was conducted on a larger Southern Han Chinese population to assess the association of SNP rs3743104 with hypospadias susceptibility.
This is the largest Chinese population-based study to examine the correlation of GREM1 genetic polymorphisms with hypospadias risk, enrolling 557 cases and 645 unrelated controls. In this study, the associated of GREM1 rs3743104 with hypospadias was replicated (Table 1). Further analysis of subclinical manifestations clarified SNP rs3743104 and revealed associations with mild/moderate and severe hypospadias (Table 2). Given that rs3743104 is located at the 3′-UTR of GREM1, we determined a series of miRNAs predicted to bind to this region, suggesting that rs3743104 has a regulatory effect on expression of GREM1 (Table 3). eQTL analysis suggests that GREM1 rs3743104 may reduce androgen production by the hypothalamic-pituitary-gonadal axis (HPG axis, Figure 1). The HPG axis in males has a central role in developing the genitalia, including spermatogenesis and testosterone production [30]. The testosterone of hypospadias patients was significantly lower than that of the normal control population after human chorionic gonadotrophin stimulation [31]. Given that rs3743104 is located at the 3′-UTR of GREM1, we provided a series of experimental evidence showing that rs3743104 has a regulatory effect on the expression of upstream GREM1, which thereby might contribute to the progression of Hypospadias. we identified that miR-182 could target the seeding region spanning rs3743104 at the 3′-UTR of GREM1, which suppresses the transcription of GREM1. miR-182 has been reported as a biomarker for kidney injury and bladder cancer, and a regulatory miRNA binding to 3′-UTR of gene targets to the progression of diseases [32, 33]. It's worthy of further work that how miR-182 is regulated. Our study has some strengths. In addition to the largest sample size in Asia, our discovery withstood a strict threshold for statistical significance corrected by multiple tests. The clinical associations remained significant after adjusting for known prognostic factors.
GREM1 is located at 15q13.3 and encodes a member of the bone morphogenic protein antagonist family. GREM1 has been reported to have a close relationship with a variety of malignant tumors in recent years. For example, Gu et al. [15] reported that overexpression of GREM1 in osteosarcoma cells in vitro and in vivo suppresses migration, invasion, angiogenesis and proliferation. It has also been shown that SNPs near GREM1 and SCG5 are significantly related to increased risk of colorectal cancer (CRC) [34]. Jang et al. [35] mentioned that GERM1-expressing fibroblasts were closely associated with lower invasiveness and better prognosis in CRC, suggesting that stromal GREM1 may represent a novel target for the treatment of CRCs and a potential biomarker. Neckmann et al. [36] showed that GREM1 is related to metastasis and poor prognosis in patients with ER-negative breast cancer and that it is a potential target for treatment. These studies reveal that GREM1 plays a vital role in tumorigenesis and can serve as a biomarker for prognosis prediction and therapeutic targets. It is worth mentioning that EMT in colon cancer cells can be suppressed by silencing of GREM1 with shRNA, [16] and EMT is considered a critical event inducing morphogenetic changes during the process of embryonic development [17]. Church et al. [37] found that among GREM1-knockout mice with a C57BL/6 background, most died in a short period of time after birth due to lung defects and kidney dysplasia. Wang [38] et al. revealed a significant correlation between GREM1 polymorphism rs1258763 and the risk of nonsyndromic orofacial clefts, and Viena CS [39] found that interaction between NTN1 and GREM1 may be associated with the pathogenesis of nonsyndromic cleft lip with or without cleft palate. It was also reported that GREM1 inhibits BMP2-mediated differentiation of MSCs to osteoblasts [40]. In human esophageal squamous cell carcinoma, EMT can be promoted by GREM1 derived from mesenchymal stromal cells [41]. In mice, hypospadias can be induced by dibutyl phthalate (DBP), which is capable of inhibiting EMT in urethral epithelial cells and blocking fusion of the urethral meatus via oxidative stress [42]. Thus, it was hypothesized that GREM1 might have an impact on urethral development through EMT. Nevertheless, the biological mechanism underlying the correlation between GREM1 polymorphism and hypospadias sensitivity remains unclear.
This study demonstrated the association of rs3743104 with hypospadias in mild/moderate and severe cases, whereas previous research failed to allocate patients into subgroups. In addition, this study has some limitations. First, the effect of gene-environment interactions was not analyzed. Second, more precise ORs should be adjusted based on fetal exposure. Third, in-depth analyses are warranted to explore the biological mechanisms of GREM1 rs3743104 and hypospadias sensitivity.
Conclusion
In summary, GREM1 risk allele rs3743104[G] increase susceptibility to hypospadias in mild/moderate and severe cases from the Southern Han Chinese population and regulates GREM1 expression by altering the binding affinity of miR-182 to their locus. Further studies are required to understand the biological mechanism by which variations in GREM1 contribute to the pathogenesis of hypospadias.
Supplementary Materials
Abbreviations
eQTL: expression quantitative trait locus; SNP: single-nucleotide polymorphism; miRNAs: microRNAs; shRNA: short hairpin RNA; GWAS: genome-wide association study; EMT: epithelial-mesenchymal transition; GTEx: Genotype-Tissue Expression Project; HWE: Hardy–Weinberg equilibrium; CI: confidence interval; CRC: colorectal cancer; DBP: dibutyl phthalate.
Author Contributions
Wen Fu and Guochang Liu designed experiment. Fuming Deng, Jinglu Zhao, Wei Jia, Kai Fu, Xiaoyu Zuo, Ning Wang, Huiming Xia, Yan Zhang, Wen Fu and Guochang Liu collected samples and conducted the study. Fuming Deng, Jinglu Zhao and Wei Jia analyzed the data and wrote the paper. All authors read and approved the manuscript.
Acknowledgments
We thank Tuqun Hu for their assistance in DNA extraction and the collection of medical histories. The authors have no conflicts of interest to declare. All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional review board of Guangzhou Women and Children’s Medical Center (NO. 39401) and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
Huimin Xia thanks the supporting of National Natural Science Foundation of China (Grant NO.8177629) and Guangdong Provincial Key Laboratory of Research in Structural Birth Defect Disease (Grant No. 2019B030301004). Lihua Huang thanks the grant National Natural Science Foundation of China (Grant NO.81600400) and the China Postdoctoral Science Foundation (Grant NO.2017T100621). Yan Zhang thanks the supporting by the grant of National Natural Science Foundation of China (Grant NO.81970450) and the Science and Technology Project of Guangzhou (Grant NO.201903010074). Fuming Deng thanks the fund from Guangzhou Institute of Pediatrics/Guangzhou Women and Children’s Medical Center (Grant NO. 0190026).
Editorial Note
This corresponding author has a verified history of publications using a personal email address for correspondence
References
- 1. Carmichael SL, Shaw GM, Lammer EJ. Environmental and genetic contributors to hypospadias: a review of the epidemiologic evidence. Birth Defects Res A Clin Mol Teratol. 2012; 94:499–510. https://doi.org/10.1002/bdra.23021 [PubMed]
- 2. Yu X, Nassar N, Mastroiacovo P, Canfield M, Groisman B, Bermejo-Sánchez E, Ritvanen A, Kiuru-Kuhlefelt S, Benavides A, Sipek A, Pierini A, Bianchi F, Källén K, et al. Hypospadias Prevalence and Trends in International Birth Defect Surveillance Systems, 1980-2010. Eur Urol. 2019; 76:482–90. https://doi.org/10.1016/j.eururo.2019.06.027 [PubMed]
- 3. Fredell L, Kockum I, Hansson E, Holmner S, Lundquist L, Läckgren G, Pedersen J, Stenberg A, Westbacke G, Nordenskjöld A. Heredity of hypospadias and the significance of low birth weight. J Urol. 2002; 167:1423–27. https://doi.org/10.1016/S0022-5347(05)65334-7 [PubMed]
- 4. Duckett JW
Jr . Hypospadias. Pediatr Rev. 1989; 11:37–42. https://doi.org/10.1542/pir.11-2-37 [PubMed] - 5. Cunha GR, Sinclair A, Risbridger G, Hutson J, Baskin LS. Current understanding of hypospadias: relevance of animal models. Nat Rev Urol. 2015; 12:271–80. https://doi.org/10.1038/nrurol.2015.57 [PubMed]
- 6. Leung AK, Robson WL. Hypospadias: an update. Asian J Androl. 2007; 9:16–22. https://doi.org/10.1111/j.1745-7262.2007.00243.x [PubMed]
- 7. Shih EM, Graham JM
Jr . Review of genetic and environmental factors leading to hypospadias. Eur J Med Genet. 2014; 57:453–63. https://doi.org/10.1016/j.ejmg.2014.03.003 [PubMed] - 8. George M, Schneuer FJ, Jamieson SE, Holland AJ. Genetic and environmental factors in the aetiology of hypospadias. Pediatr Surg Int. 2015; 31:519–27. https://doi.org/10.1007/s00383-015-3686-z [PubMed]
- 9. Söderhäll C, Körberg IB, Thai HT, Cao J, Chen Y, Zhang X, Shulu Z, van der Zanden LF, van Rooij IA, Frisén L, Roeleveld N, Markljung E, Kockum I, Nordenskjöld A. Fine mapping analysis confirms and strengthens linkage of four chromosomal regions in familial hypospadias. Eur J Hum Genet. 2015; 23:516–22. https://doi.org/10.1038/ejhg.2014.129 [PubMed]
- 10. van der Zanden LF, van Rooij IA, Feitz WF, Franke B, Knoers NV, Roeleveld N. Aetiology of hypospadias: a systematic review of genes and environment. Hum Reprod Update. 2012; 18:260–83. https://doi.org/10.1093/humupd/dms002 [PubMed]
- 11. Kojima Y, Kohri K, Hayashi Y. Genetic pathway of external genitalia formation and molecular etiology of hypospadias. J Pediatr Urol. 2010; 6:346–54. https://doi.org/10.1016/j.jpurol.2009.11.007 [PubMed]
- 12. van der Zanden LF, van Rooij IA, Feitz WF, Knight J, Donders AR, Renkema KY, Bongers EM, Vermeulen SH, Kiemeney LA, Veltman JA, Arias-Vásquez A, Zhang X, Markljung E, et al. Common variants in DGKK are strongly associated with risk of hypospadias. Nat Genet. 2011; 43:48–50. https://doi.org/10.1038/ng.721 [PubMed]
- 13. Geller F, Feenstra B, Carstensen L, Pers TH, van Rooij IA, Körberg IB, Choudhry S, Karjalainen JM, Schnack TH, Hollegaard MV, Feitz WF, Roeleveld N, Hougaard DM, et al. Genome-wide association analyses identify variants in developmental genes associated with hypospadias. Nat Genet. 2014; 46:957–63. https://doi.org/10.1038/ng.3063 [PubMed]
- 14. Chen Z, Lin X, Lei Y, Chen H, Finnell RH, Wang Y, Xu J, Lu D, Xie H, Chen F. Genome-wide association study in Chinese cohort identifies one novel hypospadias risk associated locus at 12q13.13. BMC Med Genomics. 2019; 12:196. https://doi.org/10.1186/s12920-019-0642-0 [PubMed]
- 15. Gu Q, Luo Y, Chen C, Jiang D, Huang Q, Wang X. GREM1 overexpression inhibits proliferation, migration and angiogenesis of osteosarcoma. Exp Cell Res. 2019; 384:111619. https://doi.org/10.1016/j.yexcr.2019.111619 [PubMed]
- 16. Liu Y, Li Y, Hou R, Shu Z. Knockdown GREM1 suppresses cell growth, angiogenesis, and epithelial-mesenchymal transition in colon cancer. J Cell Biochem. 2019; 120:5583–96. https://doi.org/10.1002/jcb.27842 [PubMed]
- 17. Kim DH, Xing T, Yang Z, Dudek R, Lu Q, Chen YH. Epithelial Mesenchymal Transition in Embryonic Development, Tissue Repair and Cancer: A Comprehensive Overview. J Clin Med. 2017; 7:1. https://doi.org/10.3390/jcm7010001 [PubMed]
- 18. Lu T, Li L, Zhu J, Liu J, Lin A, Fu W, Liu G, Xia H, Zhang T, He J. AURKA rs8173 G>C Polymorphism Decreases Wilms Tumor Risk in Chinese Children. J Oncol. 2019; 2019:9074908. https://doi.org/10.1155/2019/9074908 [PubMed]
- 19. He J, Wang F, Zhu J, Zhang R, Yang T, Zou Y, Xia H. Association of potentially functional variants in the XPG gene with neuroblastoma risk in a Chinese population. J Cell Mol Med. 2016; 20:1481–90. https://doi.org/10.1111/jcmm.12836 [PubMed]
- 20. He J, Yang T, Zhang R, Zhu J, Wang F, Zou Y, Xia H. Potentially functional polymorphisms in the LIN28B gene contribute to neuroblastoma susceptibility in Chinese children. J Cell Mol Med. 2016; 20:1534–41. https://doi.org/10.1111/jcmm.12846 [PubMed]
- 21. He J, Zhang X, Zhang J, Zhang R, Yang T, Zhu J, Xia H, Zou Y. LMO1 super-enhancer polymorphism rs2168101 G>T correlates with decreased neuroblastoma risk in Chinese children. J Cancer. 2018; 9:1592–97. https://doi.org/10.7150/jca.24326 [PubMed]
- 22. Liu J, Jia W, Hua RX, Zhu J, Zhang J, Yang T, Li P, Xia H, He J, Cheng J. APEX1 Polymorphisms and Neuroblastoma Risk in Chinese Children: A Three-Center Case-Control Study. Oxid Med Cell Longev. 2019; 2019:5736175. https://doi.org/10.1155/2019/5736175 [PubMed]
- 23. Schaub MA, Boyle AP, Kundaje A, Batzoglou S, Snyder M. Linking disease associations with regulatory information in the human genome. Genome Res. 2012; 22:1748–59. https://doi.org/10.1101/gr.136127.111. [PubMed]
- 24. Maurano MT, Humbert R, Rynes E, Thurman RE, Haugen E, Wang H, Reynolds AP, Sandstrom R, Qu H, Brody J, Shafer A, Neri F, Lee K, et al. Systematic localization of common disease-associated variation in regulatory DNA. Science. 2012; 337:1190–95. https://doi.org/10.1126/science.1222794 [PubMed]
- 25. Li MJ, Li M, Liu Z, Yan B, Pan Z, Huang D, Liang Q, Ying D, Xu F, Yao H, Wang P, Kocher JA, Xia Z, et al. cepip: context-dependent epigenomic weighting for prioritization of regulatory variants and disease-associated genes. Genome Biol. 2017; 18:52. https://doi.org/10.1186/s13059-017-1177-3 [PubMed]
- 26. Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA, Bender D, Maller J, Sklar P, de Bakker PI, Daly MJ, Sham PC. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet. 2007; 81:559–75. https://doi.org/10.1086/519795 [PubMed]
- 27. Benzer S. FINE STRUCTURE OF A GENETIC REGION IN BACTERIOPHAGE. Proc Natl Acad Sci USA. 1955; 41:344–54. https://doi.org/10.1073/pnas.41.6.344 [PubMed]
- 28. Hasin-Brumshtein Y, Hormozdiari F, Martin L, van Nas A, Eskin E, Lusis AJ, Drake TA. Allele-specific expression and eQTL analysis in mouse adipose tissue. BMC Genomics. 2014; 15:471. https://doi.org/10.1186/1471-2164-15-471 [PubMed]
- 29. Kojima Y, Koguchi T, Mizuno K, Sato Y, Hoshi S, Hata J, Nishio H, Hashimoto D, Matsushita S, Suzuki K, Miyagawa S, Hui CC, Tanikawa C, et al. Single Nucleotide Polymorphisms of HAAO and IRX6 Genes as Risk Factors for Hypospadias. J Urol. 2019; 201:386–92. https://doi.org/10.1016/j.juro.2018.07.050 [PubMed]
- 30. Kuiri-Hänninen T, Sankilampi U, Dunkel L. Activation of the hypothalamic-pituitary-gonadal axis in infancy: minipuberty. Horm Res Paediatr. 2014; 82:73–80. https://doi.org/10.1159/000362414 [PubMed]
- 31. Knorr D, Beckmann D, Bidlingmaier F, Helmig FJ, Sippell WG. Plasma testosterone in male puberty. II. hCG stimulation test in boys with hypospadia. Acta Endocrinol (Copenh). 1979; 90:365–71. https://doi.org/10.1530/acta.0.0900365 [PubMed]
- 32. Xie F, Li Y, Wang M, Huang C, Tao D, Zheng F, Zhang H, Zeng F, Xiao X, Jiang G. Circular RNA BCRC-3 suppresses bladder cancer proliferation through miR-182-5p/p27 axis. Mol Cancer. 2018; 17:144. https://doi.org/10.1186/s12943-018-0892-z [PubMed]
- 33. Ding C, Ding X, Zheng J, Wang B, Li Y, Xiang H, Dou M, Qiao Y, Tian P, Xue W. miR-182-5p and miR-378a-3p regulate ferroptosis in I/R-induced renal injury. Cell Death Dis. 2020; 11:929. https://doi.org/10.1038/s41419-020-03135-z [PubMed]
- 34. Jaeger E, Webb E, Howarth K, Carvajal-Carmona L, Rowan A, Broderick P, Walther A, Spain S, Pittman A, Kemp Z, Sullivan K, Heinimann K, Lubbe S, et al, and CORGI Consortium. Common genetic variants at the CRAC1 (HMPS) locus on chromosome 15q13.3 influence colorectal cancer risk. Nat Genet. 2008; 40:26–28. https://doi.org/10.1038/ng.2007.41 [PubMed]
- 35. Jang BG, Kim HS, Chang WY, Bae JM, Oh HJ, Wen X, Jeong S, Cho NY, Kim WH, Kang GH. Prognostic significance of stromal GREM1 expression in colorectal cancer. Hum Pathol. 2017; 62:56–65. https://doi.org/10.1016/j.humpath.2016.12.018 [PubMed]
- 36. Neckmann U, Wolowczyk C, Hall M, Almaas E, Ren J, Zhao S, Johannessen B, Skotheim RI, Bjørkøy G, Ten Dijke P, Holien T. GREM1 is associated with metastasis and predicts poor prognosis in ER-negative breast cancer patients. Cell Commun Signal. 2019; 17:140. https://doi.org/10.1186/s12964-019-0467-7 [PubMed]
- 37. Church RH, Ali I, Tate M, Lavin D, Krishnakumar A, Kok HM, Hombrebueno JR, Dunne PD, Bingham V, Goldschmeding R, Martin F, Brazil DP. Gremlin1 plays a key role in kidney development and renal fibrosis. Am J Physiol Renal Physiol. 2017; 312:F1141–57. https://doi.org/10.1152/ajprenal.00344.2016 [PubMed]
- 38. Wang X, Song H, Jiao X, Hao Y, Zhang W, Gao Y, Li Y, Mi N, Yan J. Association between a single-nucleotide polymorphism in the GREM1 gene and non-syndromic orofacial cleft in the Chinese population. J Oral Pathol Med. 2018; 47:206–10. https://doi.org/10.1111/jop.12662 [PubMed]
- 39. Viena CS, Machado RA, Persuhn DC, Martelli-Júnior H, Medrado AP, Coletta RD, Reis SR. Understanding the participation of GREM1 polymorphisms in nonsyndromic cleft lip with or without cleft palate in the Brazilian population. Birth Defects Res. 2019; 111:16–25. https://doi.org/10.1002/bdr2.1405 [PubMed]
- 40. Olivares-Navarrete R, Hyzy SL, Haithcock DA, Cundiff CA, Schwartz Z, Boyan BD. Coordinated regulation of mesenchymal stem cell differentiation on microstructured titanium surfaces by endogenous bone morphogenetic proteins. Bone. 2015; 73:208–16. https://doi.org/10.1016/j.bone.2014.12.057 [PubMed]
- 41. Hong D, Liu T, Huang W, Liao Y, Wang L, Zhang Z, Chen H, Zhang X, Xiang Q. Gremlin1 Delivered by Mesenchymal Stromal Cells Promoted Epithelial-Mesenchymal Transition in Human Esophageal Squamous Cell Carcinoma. Cell Physiol Biochem. 2018; 47:1785–99. https://doi.org/10.1159/000491060 [PubMed]
- 42. Zhu YP, Zhao W, Sun WL, Zhao S, Chen M, Pan L, Zhou Z, Xia SJ, Jiang JT. Di-n-butyl phthalate (DBP) reduces epithelial-mesenchymal transition via IP3R in hypospadias during maternal exposure. Ecotoxicol Environ Saf. 2020; 192:110201. https://doi.org/10.1016/j.ecoenv.2020.110201 [PubMed]