



Introduction
Gastric carcinoma (GC) is a malignant tumor that originates from the gastric mucosa epithelial cells. According to the latest global cancer statistics, GC had the fifth highest global incidence rate and the fourth highest mortality rate in 2020 [1]. In the early stages of GC, there are no obvious symptoms, and clinical manifestations such as weight loss, epigastric pain, and loss of appetite can occur in the middle and late stages [2]. At present, the etiology of GC is not clear, and possible causative factors include Helicobacter pylori (HP) infection, geographical environment, diet, and genetics [3]. Due to the strong concealment of GC, surgery, radiotherapy, and chemotherapy are often ineffective, and tend to cause a range of side effects in the organism. Therefore, people are gradually turning their attention to Traditional Chinese Medicine (TCM).
In recent years, a growing number of studies have shown that certain Chinese herbal monomers and compounds have certain therapeutic effects on gastric carcinoma. Liang et al. [4] found that Galangin can inhibit the proliferation of GC cells by inhibiting NF-κb signaling pathway in vitro and in vivo. Berberine can induce apoptosis of GC cells, promote cell cycle arrest, inhibit cell invasion and migration by regulating the IL-6/JAK2/STAT3 signaling pathway, thus inhibiting the development of GC [5]. Li et al. [6] showed that Si-Jun-Zi decoction (SJZ) could inhibit the proliferation of GC cells by activating CMTM2, in vivo experiments also showed that SJZ could induce apoptosis of GC cells by down-regulating the mRNA expression level of p53 and Bcl-2 [7]. Banxia Xiexin Decoction (BXD) can significantly inhibit the cell viability and induce apoptosis of GC cells, which is mainly related to the inhibition of Wnt/β-catenin signaling pathway [8]. Through in vivo and in vitro experiments, Sun et al. [9] found that the combined application of Zuo Jin Wan (ZJW) and DDP could up-regulate the protein expression levels of Cleaved ROCK and P-PTEN, and down-regulate the expression levels of P-PI3K and p-cofilin-1 to induce apoptosis of GC cells. These results suggest that ZJW may inhibit DDP resistance in GC by regulating the ROCK/PTEN/PI3K signaling pathway. Some studies have found that Danggui Sini Decoction (DSD) can inhibit the Akt/Erk/p53 signaling pathway in a concentration-dependent manner and induce cell cycle arrest and apoptosis, thus inhibiting the proliferation of GC cells [10]. It can be seen that TCM plays an important role in the treatment of GC and is gradually attracting the attention of researchers. The ancient Chinese classical prescription for Shaoyao-Gancao Decoction (SG-D) is derived from Treatise on Febrile and Miscellaneous Diseases by Zhang Zhongjing, which consists of two kinds of Chinese herbal medicine, namely Shaoyao (Paeonia lactiflora.) and Gancao (Glycyrrhiza uralensis Fisch.). Shaoyao can preserve yin to nourish the blood, soften livers and relieve pain. Gancao can tonify middle-Jiao and Qi, relax spasm and relieve pain. Studies have shown that SG-D has a favorable therapeutic effect on digestive system diseases, such as anti-inflammation and protection of gastric mucosa, etc. [11]. However, the relation between SG-D and GC is still poorly studied.
In recent years, network pharmacology has been gradually applied in the field of TCM, which reveals the mechanism of drugs action based on the Chinese herbal medicine-components-targets-pathways-disease network [12]. This coincides with the characteristics of multi-components, multi-targets, and multi-pathways of TCM. Therefore, this research predicted the mechanism of action of SG-D in the treatment of GC through the method of network pharmacology and finally screened out the key active ingredients such as quercetin, core targets such as AKT, and signaling pathways such as PI3K/Akt and MAPK. Subsequently, the results were verified by in vitro cell experiments, which showed that SG-D was able to inhibit the proliferation of AGS cells, induce cell apoptosis and cycle arrest, and decrease the ability of cell clone formation by regulating PI3K/Akt and MAPK signaling pathways. Therefore, this study verified the efficacy of SG-D in the treatment of GC through the combination of network pharmacology and experimental verification, in order to provide a reference for future studies on SG-D in treating GC. The flow chart of this study is shown in Figure 1.
Figure 1. Flow chart.
Results
Active ingredients and targets for SG-D
After screening the active ingredients of SG-D with OB ≥ 30% and DL ≥ 0.18 in TCMSP database, a total of 105 active ingredients were obtained, including 13 from Shaoyao and 92 from Gancao (Supplementary Table 1 for details). At the same time, 1880 related targets were collected through the database, and after normalization processing by the Uniport database and removal of duplicates and non-human targets, a total of 195 targets for SG-D were obtained. Then, the Network Analyzer plug-in was used to calculate the Degree value of the nodes, and the active ingredients of Degree ranked in the top 20 were obtained (Table 1). Among them, quercetin, Kaempferol, and naringenin ranked high, with degrees of 104, 72, and 23, respectively. The construction of “component-target” network of SG-D is shown in Figure 2A, and the detailed names of ingredients are shown in Supplementary Table 2.
Table 1. Active ingredients information.
| Number | Molecule name | Degree | OB (%) | DL | Source |
| 1 | quercetin | 104 | 46.43 | 0.28 | Gancao |
| 2 | kaempferol | 72 | 41.88 | 0.24 | Shaoyao/Gancao |
| 3 | naringenin | 23 | 59.29 | 0.21 | Gancao |
| 4 | 1-Methoxyphaseollidin | 20 | 69.98 | 0.64 | Gancao |
| 5 | formononetin | 19 | 69.67 | 0.21 | Gancao |
| 6 | 7-Methoxy-2-methyl isoflavone | 18 | 42.56 | 0.2 | Gancao |
| 7 | HMO | 18 | 38.37 | 0.21 | Gancao |
| 8 | Quercetin der. | 18 | 46.45 | 0.33 | Gancao |
| 9 | 3’-Hydroxy-4’-O-Methylglabridin | 18 | 43.71 | 0.57 | Gancao |
| 10 | Jaranol | 17 | 50.83 | 0.29 | Gancao |
| 11 | licoisoflavanone | 17 | 52.47 | 0.54 | Gancao |
| 12 | 3,22-Dihydroxy-11-oxo-delta(12)-oleanene -27-alpha-methoxycarbonyl-29-oic acid | 17 | 34.32 | 0.55 | Gancao |
| 13 | Isoglycyrol | 17 | 44.7 | 0.84 | Gancao |
| 14 | kanzonols W | 16 | 50.48 | 0.52 | Gancao |
| 15 | gadelaidic acid | 16 | 30.7 | 0.2 | Gancao |
| 16 | paeoniflorin | 15 | 53.87 | 0.79 | Shaoyao |
| 17 | Calycosin | 15 | 47.75 | 0.24 | Gancao |
| 18 | Glepidotin A | 15 | 44.72 | 0.35 | Gancao |
| 19 | Phaseolinisoflavan | 15 | 32.01 | 0.45 | Gancao |
| 20 | (2S)-7-hydroxy-2-(4-hydroxyphenyl)-8-(3-methylbut-2-enyl)chroman-4-one | 15 | 36.57 | 0.32 | Gancao |
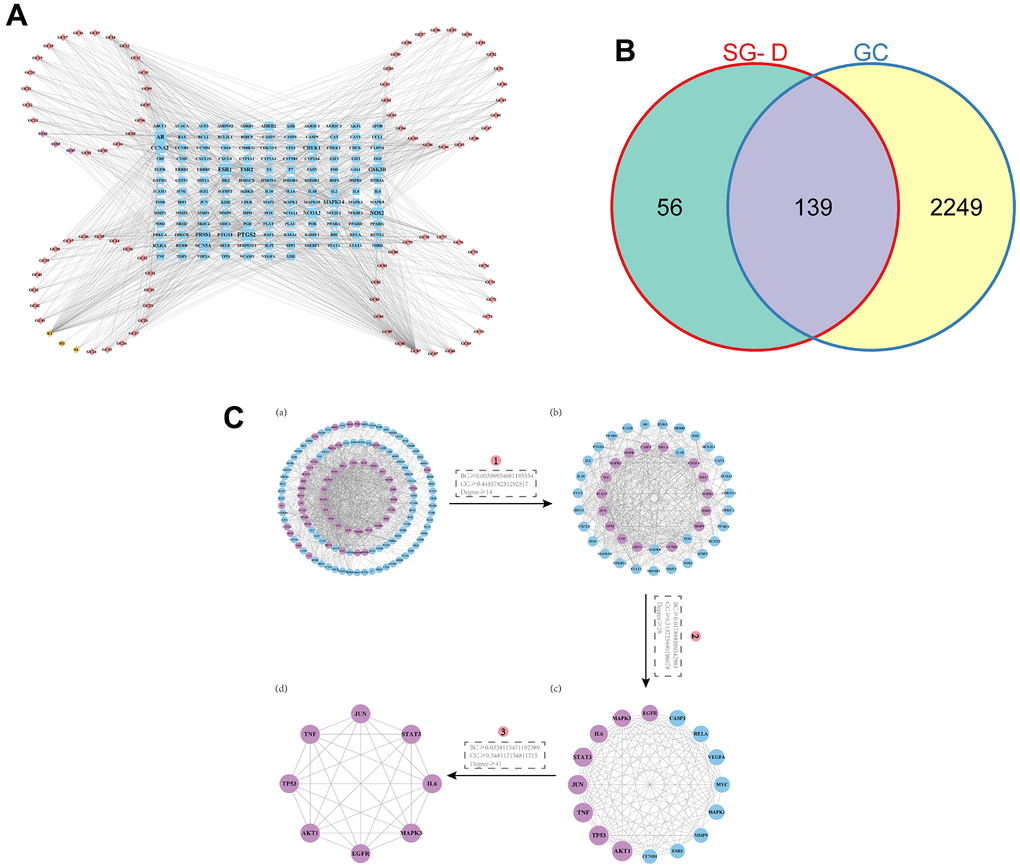
Figure 2. Prediction of active ingredients and core targets. (A) The “component-target” network. The active ingredients in Shaoyao are shown by the purple nodes, those in Gancao are represented by the pink nodes, the common active ingredients of SG-D by yellow nodes, and the common targets by blue node. The more active ingredients are connected to a node as it grows larger. (B) Venn diagram of intersecting targets of SG-D and GC. (C) The screening of core targets in the PPI network, (a) Containing 132 nodes and 1138 edges, purple nodes indicate targets with higher BC, CC, and Degree. (b) Containing 46 nodes and 491 edges. (c) Containing 16 nodes and 103 edges. (d) Containing 8 nodes and 28 edges.
Prediction of potential targets for SG-D in treating GC
2216 targets were obtained after screening and deleting duplicate entries in Genecards database, 321 targets were obtained after deleting duplicate entries in DisGeNET database with the marking criterion ≥ 0.3 and 73 targets were obtained after deleting duplicate entries in DrugBank database. Finally, 2388 GC-related targets were obtained by combining and deleting duplicate entries in the three databases. Then, 195 drug targets and 2388 disease targets were intersected to obtain 139 SG-D targets in the treatment of GC (Figure 2B).
Analysis of the PPI network
A total of 139 intersection targets of SG-D and GC were uploaded into the STRING database, the confidence level was set to “> 0.700” and the free proteins were hidden, 139 nodes and 1138 edges were obtained after screening. Then, the results were imported into Cytoscape 3.9.0 software, and the topology of the PPI network was analyzed with the CytoNCA plug-in under the condition that BC, CC, and Degree were all greater than or equal to the median value, the first screening criteria were BC ≥ 0.00338954681195354, CC ≥ 0.445578231292517, Degree ≥ 14, the second screening criteria were BC ≥ 0.017888109542983, CC ≥ 0.513725490196078, Degree ≥ 29, and the third screening criteria were BC ≥ 0.0338115471192389, CC ≥ 0.548117154811715, Degree ≥ 41. Finally, the core network including 8 nodes and 28 edges were obtained. These targets were AKT1, TP53, TNF, JUN, STAT3, IL6, MAPK3 and EGFR, respectively (Figure 2C). We hypothesized that these targets are the key targets of SG-D in the treatment of GC.
GO functional enrichment analysis and KEGG pathway enrichment analysis
The intersection targets were imported into the Metascape database for GO and KEGG enrichment analysis. The results of GO enrichment analysis showed that the therapeutic effect of SG-D on GC was mainly related to the biological processes such as response to lipopolysaccharide, cell response to lipid, response to inorganic substances, and apoptotic signaling pathway. The cellular components include membrane raft, transcriptional regulatory complex, and receptor complex. The molecular functions include transcription factor binding, protein kinase binding, and protein domain specific binding (Figure 3A). KEGG analysis enriched the top 20 most crucial pathways, and the results suggested that SG-D may exert anti-gastric carcinoma effects through pathways in cancer, MAPK signaling pathway and PI3K/Akt signaling pathway (Figure 3B).
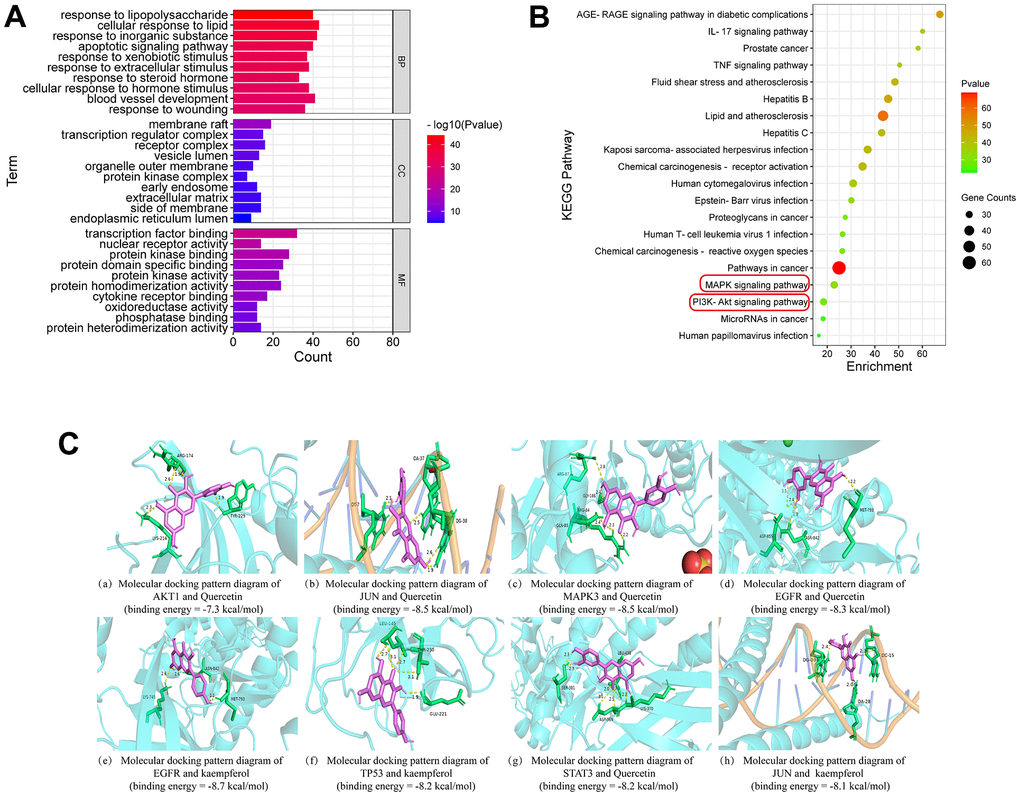
Figure 3. Enrichment analysis and molecular docking. (A) GO enrichment analysis. (B) The bubble diagram of KEGG pathway enrichment analysis. An increase in the value of P is shown by a change in the color of the nodes from green to red, and an increase in the number of genes is indicated by a change in the size of the nodes from tiny to large. (C) Molecular docking patterns of active ingredients and core targets.
Molecular docking results
By using Auto Dock Vina software, the core targets were docked with the key active ingredients one by one to get the binding energy (Table 2). If the binding energy is less than - 4.25 kcal·mol-1, it indicates that the two have certain binding ability; if the binding energy is less than - 5.0 kcal·mol-1, the two have good binding ability; if the binding energy is less than - 7.0 kcal·mol-1, the binding ability of the two is particularly close. The molecular docking results showed that the 8 core targets have excellent binding energy with the key active ingredients, in agreement with network pharmacology results. The molecular docking pattern with higher binding energy are shown in Figure 3C.
Table 2. “Active ingredients-targets” binding energy.
| Uniprot Id | PDB ID | Gene symbol | Binding energy(kcal/mol) | ||
| Quercetin | Kaempferol | Naringenin | |||
| P31749 | 3CQW | AKT1 | -7.3 | -6.4 | -6.2 |
| P01375 | 5M2I | TNF | -7.3 | -7.1 | -7.3 |
| P04637 | 6GGB | TP53 | -8.3 | -8.2 | -7.4 |
| P05412 | 5T01 | JUN | -8.5 | -8.1 | -7.4 |
| P40763 | 6NJS | STAT3 | -8.2 | -8.0 | -6.1 |
| P05231 | 1ALU | IL6 | -6.3 | -6.7 | -6.6 |
| P27361 | 6GES | MAPK3 | -8.5 | -8.0 | -7.3 |
| P00533 | 6P1D | EGFR | -8.3 | -8.7 | -7.7 |
SG-D decreased the viability of AGS cells SG-D
In order to detect the effect of SG-D on the proliferation of AGS cells, we treated human AGS cells with different concentrations of SG-D (2, 4, 6, 8, 10, and 12 g/L) for 24, 48, and 72 h. The results showed that SG-D could significantly decrease the viability of AGS cells in a concentration and time-dependent manner, that is, with the increase of drug concentration and treatment time, its inhibition gradually increased (Figure 4A). We found that cell proliferation was significantly inhibited and cell status was positive at 24 h of drug intervention, therefore we chose 24 h as the time for the follow-up drug intervention. When the concentration of SG-D was 8g/L, the inhibition rate of AGS cells was close to 50% (IC50 = 8.4 g/L for 24 h), so we chose the 8 g/L as the medium concentration and determined the low and high concentration in turn (Figure 4B).
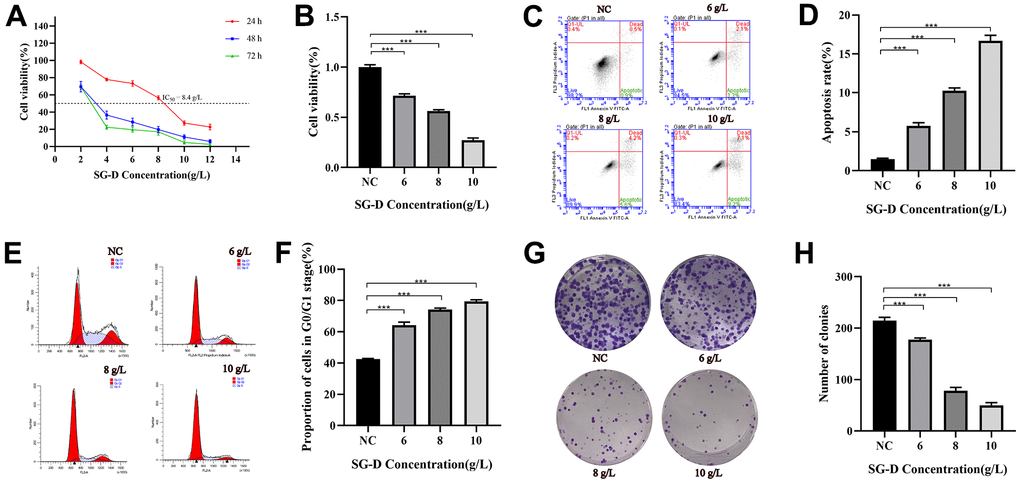
Figure 4. The effect of SG-D on the phenotype of AGS cells. (A) The impact of SG-D on the viability of AGS cells 24, 48, and 72 hours after treatment. (B) The impact of different SG-D concentrations (6, 8, and 10 g/L) on the viability of AGS cells. (C, D) Apoptosis rate. (E, F) Cell cycle distribution. (G, H) The quantity of generated cell clones. All experiments were repeated three times and the data were expressed as mean±SD, *p < 0.05, **p < 0.01, ***p < 0.001.
SG-D induced the apoptosis of AGS cells
The results of apoptosis showed that the apoptosis rates of AGS cells were (5.77 ± 0.40) %, (10.23 ± 0.38) %, (16.67 ± 0.72) % after intervention of low (6 g/L), middle (8 g/L) and high concentrations (10 g/L) of SG-D for 24 h, respectively, all higher than those of the control group (1.50 ± 0.10) % (Figure 4C, 4D).
SG-D arrested AGS cells cycle in the G0/G1 phase
To further investigate the effect of SG-D on GC cells, we used Flow cytometry to examine the effect of SG-D on the cell cycle distribution of GC cells. The results showed that the percentage of AGS cells in G0/G1 phase changed significantly after treatment with different concentrations of SG-D (6, 8, and 10 g/L) for 24 h, which were (64.24 ± 1.97) %, (74.21 ± 0.95) % and (79.38 ± 1.09) %, respectively, they were all higher than the control group (42.50 ± 0.49) % (Figure 4E, 4F).
SG-D inhibited the colony forming ability of AGS cells
In order to detect the effect of SG-D on the colony forming ability of AGS cells, we used SG-D (6, 8, and 10 g/L) to intervene cells. The experimental results showed that the number of cell clones formed in AGS cells was distinctly reduced after treatment with SG-D compared with the control group, and the inhibitory effect was more obvious with the increase of concentration (Figure 4G, 4H).
SG-D inhibited AGS cells proliferation through PI3K/Akt and MAPK signaling pathways
Based on the results of network pharmacology, we found that the PI3K/Akt and MAPK signaling pathways may be the vital pathways through which SG-D exerts its anti-gastric carcinoma effects. Therefore, we used Western blotting to examine PI3K, AKT, p38, JNK, ERK, and their corresponding phosphorylated products. The results showed that compared with the control group, SG-D had no significant effect on the total proteins of PI3K, AKT, p38, JNK, and ERK, but the phosphorylated products were changed to different degrees, SG-D down-regulated the protein expression levels of p-PI3K, p-AKT, p-ERK, and significantly up-regulated the protein expression levels of p-p38 and p-JNK (Figure 5A–5D).
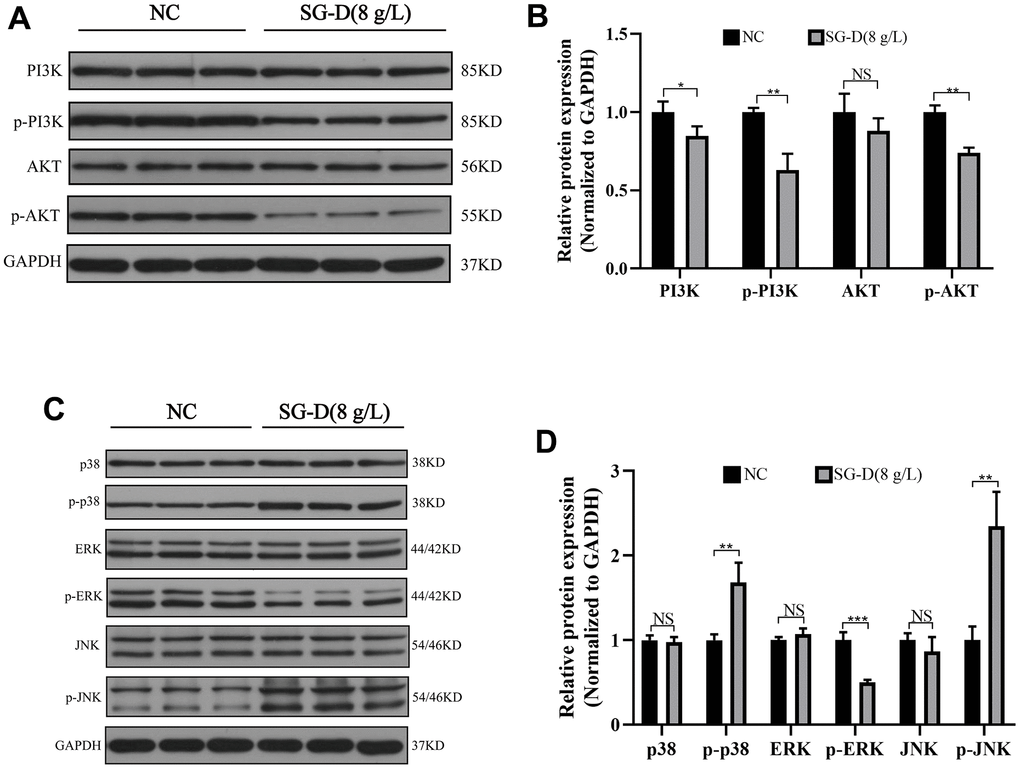
Figure 5. The effect of SG-D on PI3K/Akt and MAPK pathways in AGS cells. (A, B) The impact of SG-D on the expression of proteins connected to the PI3K/Akt signaling pathway in AGS cells. (C, D) The impact of SG-D on the expression of proteins connected to the MAPK signaling pathway in AGS cells. All experiments were repeated three times and the data were expressed as mean±SD, *p < 0.05, **p < 0.01, ***p < 0.001.
Discussion
The occurrence and development of GC is an extremely complex process, which is generally characterized by abnormal cell proliferation, strong infiltration, and metastasis [13], and poses a great threat to human physical and mental health. At present, GC remains one of the leading causes of mortality in humans. Although the treatment of GC has been improved and the mortality has been reduced significantly, the prognosis of patients with GC is usually poor, and the side effects are more [14]. Therefore, it has become a new research direction to search for drugs that have anti-cancer effects and reduce side effects. SG-D, as a classical prescription lasting for thousands of years, has the therapeutic characteristics of “multi-ingredients, multi-targets, and multi-pathways”, which can regulate human body function and harmonize Yin and Yang. It has been widely used in the treatment of various diseases in recent years, but the related studies of GC have not been reported, and its mechanism of action is still unclear. As a result, this study reveals the mechanism of SG-D in the treatment of GC through network pharmacology and experimental verification.
The network pharmacology results showed that quercetin, kaempferol and naringin are the key active ingredients of SG-D in treating GC. Quercetin is a flavonoid compound with various pharmacological effects such as anti-inflammatory, antioxidant, anti-cancer and immunomodulatory effects [15]. Studies have found that quercetin inhibits the proliferation of GC cells by inhibiting HER2 kinase [16], apoptosis can also be induced by regulating the expression of apoptosis-related proteins [17]. Kaempferol is also a flavonoid compound and has anti-inflammatory, antioxidant, and anti-cancer effects [18]. Kaempferol has been shown to be involved in a variety of biological processes, including cell proliferation [19], apoptosis [20], cell cycle [20] and autophagy [21]. Naringenin is a dihydroflavonoid compound that has anti-cancer, anti-inflammatory, and anti-oxidant pharmacological activities [22]. It can down-regulate the protein expression levels of MMP2, MMP9, BCL-2, and Survivin, and up-regulate the protein expression levels of Bax and cleaved Caspase-3 to attenuates cell invasion and migration ability and induce apoptosis, thereby exerting anti-cancer effects [23]. According to the results of PPI network analysis, we screened out several core targets of SG-D in the treatment of GC, such as AKT1, TNF, EGFR, MAPK3, STAT3, IL6, TP53, and JUN. These targets are closely related to cell proliferation, cell cycle, cell apoptosis, and other biological processes, and play a key role in the treatment of GC with SG-D. GO function enrichment analysis revealed that the therapeutic effect of SG-D on GC is mainly related to biological processes and molecular functions such as cell apoptosis and protein kinase binding. KEGG enrichment analysis has shown that the PI3K/Akt and MAPK signaling pathways are essential pathways for SG-D to treat GC. The results of molecular docking also showed that the key ingredients can be tightly bound to the core targets. Therefore, based on the results of network pharmacology, we suggest that SG-D has a therapeutic effect on GC cells by regulating PI3K/Akt and MAPK signaling pathways.
To further clarify the molecular mechanism of SG-D in the treatment of GC, we verified the predicted results by in vitro cell experiments. We used AGS cells to conduct the CCK-8 experiment, and the results showed that the cell viability of AGS cells decreased after 24 h of intervention with SG-D, and the cell viability decreased more significantly with the increase of drug concentration and the extension of time, which indicated that SG-D had a significant inhibitory effect on AGS cells in a concentration-and time-dependent manner. The change in the cell cycle plays an influential role in the occurrence and development of tumors [24]. Our experiment results showed that compared with the control group, the number of cells in G0/G1 phase increased with the increase of SG-D concentration [from (64.24 ± 1.97) % to (79.38 ± 1.09) %], which suggested that SG-D could arrest the AGS cells cycle in G0/G1 phase, thus inhibiting cell proliferation. Apoptosis is a death mechanism tightly regulated by multiple apoptotic genes [25]. Our results showed that compared with the control group, the apoptosis rate increased with the increase of drug concentration [from (1.50 ± 0.10) % to (16.67 ± 0.72) %], which indicated that SG-D could significantly induce apoptosis in AGS cells in a concentration- dependent manner. We also examined the effect of SG-D on the proliferative ability of AGS cells by plate clone formation assay, and the results showed that the number of clonogenic cells was significantly reduced, which indicated that SG-D could significantly inhibit the colony forming ability of AGS cells, and this is consistent with the results of our previous experiments. The PI3K/Akt signaling pathway is closely related to the occurrence and development of tumors, and it is commonly found to be highly expressed in GC. Phosphatidylinositol 3-kinase (PI3K) is an intracellular phosphatidylinositol kinase that can be classified as class I, class II, and class III PI3K depending on the structure and substrate [26]. Relevant studies have shown that class I PI3K plays a crucial role in the occurrence and development of tumors, and participates in cell proliferation, survival, and other processes [27]. Class I PI3K is a heterodimer made up of the regulatory subunit p110 and the catalytic subunit p85 [28]. The activated PI3K catalyzes the phosphorylation of Phosphatidylinositol (4, 5) bisphosphate (PIP2) and conversion to Phosphatidylinositol (3, 4, 5) triphosphate (PIP3) [29]. The AKT1 gene encodes a serine/threonine protein kinase, which consists of AKT1, AKT2, and AKT3 and is an important downstream signaling molecule of PI3K [30]. PIP3 binds to AKT, translocates it to the cell membrane, thus, phosphorylated AKT is activated and subsequently involved in a variety of cellular functions [31]. Mitogen-activated protein kinase (MAPK) is a group of highly conserved protein kinases [32]. The MAPK signaling pathway is one of the hubs of multiple signaling pathways involved in cell proliferation, apoptosis, and inflammation [33]. Previous studies have confirmed that the MAPK signaling pathway is extensively involved in various cellular activities of GC cells [34]. Among the four major MAPK signaling pathways, ERK, JNK, p38/MAPK, and ERK5 [35], ERK1/2 is the most extensively studied and is primarily involved in cell proliferation and differentiation [36]. After extracellular stimulation, ERK1/2 is activated and phosphorylated, and then transferred from the cytoplasm to the nucleus to phosphorylate substrates such as P90 ribosomal S6 kinase (P90rsk), proto-oncogene c-fos and c-jun, thus participating in cell proliferation, survival and other processes [37]. Apoptosis is mainly associated with JNK and p38 [38]. While JNK is activated by various extracellular stimuli such as cytokines and growth factors and then participates in cell activities [39], while p38 is activated by DNA damage and oxidative stress, thus regulating various cellular functions [40]. In this study, we examined the effect of SG-D on the PI3K/Akt and MAPK signaling pathways in AGS cells by Western Blot. The results showed that SG-D down-regulated the protein expression levels of p-PI3K, p-AKT, and p-ERK and up-regulated the protein expression of p-p38 and p-JNK, which indicated that SG-D can inhibit the proliferation of AGS cells by inhibiting the PI3K/Akt signaling pathway and the ERK pathway in the MAPK signaling pathway, and activating the p38 and JNK pathways.
In summary, this study showed that SG-D can induce apoptosis of GC cells, promote cell cycle arrest and reduce cell clone formation ability by regulating PI3K/Akt and MAPK signaling pathways, thus inhibiting the proliferation of GC cells, which could provide a scientific basis for the related research of SG-D in the treatment of GC. However, our study is limited to in vitro cell experiments due to time and funding issues, and additional experiments are needed to support our future research. Therefore, we will continue to explore the link between SG-D and GC in the future with the following study. First, the therapeutic effect of SG-D on GC is investigated by in vitro animal experiments. Second, to explore the effect of SG-D in reducing chemotherapeutic drug sensitivity in combination with chemotherapy drugs. Third, the molecular mechanism of SG-D in GC treatment through gene silencing, co-IP, EMSA, and other methods should be further investigated.
Conclusions
In this study, we investigated the molecular mechanism of SG-D in the treatment of GC through network pharmacology and experimental verification. The results showed that SG-D could inhibit the proliferation of GC cells by regulating PI3K/Akt and MAPK signaling pathways, induced apoptosis and cell cycle arrest, and reduced colony forming ability. Our results confirm the reliability of network pharmacology analysis and provide a strong scientific basis for further research.
Materials and Methods
Screening of active ingredients and corresponding targets for SG-D
The active ingredients of Shaoyao and Gancao were collected by Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform (TCMSP, https://old.tcmsp-e.com/tcmsp.php) [41], and were screened on the conditions of oral bioavailability (OB) ≥ 30% and drug likeness (DL) ≥ 0.18 [42]. The corresponding targets were collected by the TCMSP database, and the results were standardized in UniProt database (UniProtKB, https://www.uniprot.org/), the filter criteria were “Reviewed” and “Human”. Subsequently, the active ingredients of SG-D and their co-action targets were imported into Cytoscape3.9.0 software, and the node degree values were calculated using the Network Analyzer plug-in, then the key active ingredients were screened out and the component-target Network was constructed.
Screening of GC targets and intersecting targets
In the GeneCards database (https://www.genecards.org/) [43], DisGeNET database (https://www.disgenet.org/) [44] and DrugBank database (https://go.drugbank.com/) [45], “Gastricc carcinoma”, “Cancer of the stomach”, “Stomach cancer”, “Gastric cancer” and “Stomach neoplasm” were used as keywords to search for targets related to GC. The target information from the three databases were combined and the duplicated items were deleted to finally obtain the GC targets. Then, using Uniprot database, the targets of SG-D and GC were transformed into the corresponding Gene Symbol and mapped into Venn diagram (http://www.bioinformatics.com.cn/) [46], so as to obtain the intersection targets of SG-D and GC.
Construction of the PPI Network
Imported the intersecting targets into the STRING database (https://cn.string-db.org/), and the filtering criteria was set as “Homo sapiens” and “combined score > 0.700” [47]. The screened data were imported into Cytoscape 3.9.0 software, and the network topology analysis was performed using CytoNCA plug-in under the condition of BetweennessCentrality (BC), ClosenessCentrality (CC), and Degree were all greater than or equal to the median value [48]. The core genes of SG-D in GC treatment were subsequently obtained.
Enrichment analysis of GO and KEGG pathways
The Metascape database (https://metascape.org/) [49] was used for Gene Ontology (GO) analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis of intersection targets, and species were set as “H.sapiens”, the screening threshold was “Min Overlap = 3”, “Pvalue Cutoff < 0.01” and “Min Enrichment = 1.5” [50]. The top 20 items of P value in KEGG enrichment analysis were selected for analysis, and the bubble diagram of the KEGG pathway was generated using bioinformatics (http://www.bioinformatics.com.cn/).
Molecular docking
Using the PubChem database (https://pubchem.ncbi.nlm.nih.gov/) [51], download the key active ingredients of the 2D structures, then import them into Chem3D software to convert and optimize them into 3D structures. Download the 3D structures of core targets from RCSB PDB database (https://www.rcsb.org/), set species as “Homo sapiens”, and imported them into PyMOL software for processing. Then ligands and receptors were processed by AutoDockTools1.5.6 software, and AutoDockVina was used for molecular docking, the results were visualized using PyMOL software [52].
Experimental materials
The human gastric adenocarcinoma AGS cells were purchased from Shanghai Genechem Co., Ltd. (CAT: GCC-ST0003RT, Shanghai, China), the cells have been identified by STR without cross contamination. Fetal bovine serum was purchased from Gemini (CAT: 900108, Shanghai, China). DME/F-12 medium was purchased from Hyclone (CAT: SH30023.01, USA). Cell Counting Kit-8 Kit was purchased from Dojindo (CAT: CK04, Japan). Jiangsu KeyGEN Bio TECH Corp., Ltd. (CAT: KGA512/KGA107, Jiangsu, China) provided the Cell Cycle Detection Kit and the Annexin-V FITC/PI Double Staining Apoptosis Detection Kit. Crystal Violet was purchased from Beijing Biotopped Technology Co., Ltd. (CAT: C6470, Beijing, China). Immunoway provided PI3K, P-PI3K, AKT, p-AKT, p38, p-p38, p-JNK, and GADPH antibodies, while CST provided Erk, p-ERK, and JNK antibodies.
SG-D formula granules came from the Yinchuan Hospital, Affiliated Hospital of Traditional Chinese Medicine of Ningxia Medical University, Shaoyao (Paeonia lactiflora.) and Gancao (Glycyrrhiza uralensis Fisch.) are produced from Inner Mongolia and Xinjiang, China, respectively, and were prepared in a 1:1 ratio [53]. The equivalent formula granules were dissolved in 15 ml ddH2O to prepare SG-D solution (Shaoyao: 1 g granule = 5 g crude drugs; Gancao: 1 g granule = 3 g crude drugs). After centrifugation at 12000 rpm for 10 min, we collected the supernatant and filtered it with a 0.22 μm needle filter. Finally, the solution concentration was calculated as 200 g/L based on the crude drug content.
Cell culture
AGS cells were cultured in DME/F-12 medium containing 10% FBS and 1% penicillin (100 u/ml)/streptomycin (100 μg/ml) and placed in an environment of 37° C and 5 % CO2, cell passage cultivation every 2-3 days.
Cell viability assay
AGS cells were seeded into 96-well plates at the density of 6 × 103 cells per well. The experiment was divided into three groups: the blank control group, the negative control group, and the drug intervention group. The cells were treated with different concentrations of SG-D (2, 4, 6, 8, 10, and 12 g/L) for 24, 48, 72 hours, respectively. Then, 10 μl of CCK-8 solution was added to each well and cultured in the incubator for 2 hours. The optical density (OD) value of each group of cells at 450 nm wavelength was measured with a microplate reader, each group had 3 replicates. Cell viability = [(experimental group absorbance-blank group absorbance)]/[(control group absorbance-blank group absorbance)] × 100%.
Apoptosis assay
The cells were seeded in 6-well plates, and when the cell confluence rate of cells ranged from 70% to 80%, supernatants were taken and continued to be cultured with drug-containing medium, providing 3 replicates per group. After 24 hours, the cells were digested and collected, washed the cells twice with pre-cooled PBS, and 500 μl Binding Buffer, 5 μl Annexin V-FITC and 5 μl Propidium Iodide were added, then reacted in a dark place for 5~15 min and flow cytometry was then used for detection and analysis within 1 hour.
Cell cycle assay
The cells in decent growth conditions were seeded in 6-well plates and cultured for a period of time, after which different concentrations of drug-containing medium were added, and 3 replicates were established in each group. After 24 hours of drug intervention, cells were collected, washed with PBS, and immobilized at 4° C for 2 hours until overnight. After that, the stationary liquid was washed with PBS, and 500 μl of working solution (RNase A:PI = 1:9) was added into each well and reacted in a dark place for 30~60 min at room temperature. Finally, flow cytometry was used to detect the percentage of cells per cycle.
Plate clone formation assay
Based on the cell count results, 500 cells per well were seeded into 6-well plates, and 3 replicates were set up. Conventional culture was carried out for 10-14 days, changing the medium every three days, and culture was terminated when the visible clones were formed. Later, the cells were washed with PBS, and 1 ml 4% paraformaldehyde was added and fixed at 4° C for 30 min, then added 500 μl crystal violet solution and stained for 15 min at room temperature, and then washed the staining fluid, took photos after dried at room temperature, statistical analysis based on clone sizes (Diameter > 1 mm).
Western blot analysis
Cells from each group were collected, and the total protein was extracted by adding the configured lysate (The ratio was 10 μl of phosphatase inhibitor, 1 μl of Protease inhibitor, and 5 μl of 100 mM PMSF to each 1 ml of cold Lysis Buffer). The protein content was determined using the BCA Protein Quantitation Kit, and the protein was then denaturated. At the end of SDS-PAGE, target proteins were transferred to PVDF membranes, sealed with 5% skim milk, washed several times with TBST, and then incubated overnight at 4° C with primary antibodies. The PVDF membrane was then cleaned with TBST several times, the corresponding secondary antibodies were added and incubated at room temperature for 1 h. Finally, ECL luminescent solution and an ultra-sensitive multifunctional imager were used for detection, and gray value analysis of protein bands was performed using ImageJ software (NIH, Bethesda, MD, USA).
Statistical analysis
All data were statistically analyzed with GraphPad prism 8 software and expressed as mean ± SD. Using one-way ANOVA and t-tests analyzed the statistical differences, p < 0.05 indicated a significant difference.
Abbreviations
SG-D: Shaoyao-Gancao Decoction; GC: Gastric carcinoma; TCM: Traditional Chinese Medicine; TCMSP: Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform; PPI: protein-protein interaction; GO: Gene Ontology; KEGG: Kyoto Encyclopedia of Genes and Genomes; CCK-8: Cell Counting Kit-8 kit;.
Author Contributions
Xin Zhou carried out the majority of the experiments, analyzed the data, completed the figure production, and wrote the manuscript. Jiao Min carried out a portion of the experiments, and participated in the production of the figures and the composition of the manuscript. Mengying Che and Yating Yang carried out part of the experiments, and participated in the statistical analysis of the data. Yi Yang, Junfei Zhang, Lei Zhang, and Xiaosha Zheng performed the network pharmacology prediction. Yan Chen and Ling Yuan designed the study. Yi Nan revised and improved the manuscript. All authors have read the manuscript and concur with the findings of the study.
Acknowledgments
We would like to thank the Ningxia Medical University Key Laboratory of Ningxia Minority Medicine Modernization Ministry of Education for providing the experimental platform, and Yingfeng Ma, Xiaoli Du, Tingting Li and Wenjing Liu for their help.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Ethical Statement
This research was given approval by the institutional review board (NXMU-2021-018, NXMU-2021-N001, NXMU-2021-N0063) of Medical University (NXMU).
Funding
This study was supported by Key project of Ningxia Medical University (XZ2021005), Key project of Ningxia Natural Science Foundation (2022AAC02039), Ningxia Natural Science Foundation (2022AAC03144), and Innovative and Entrepreneurship Training Program for college students (S202210752035).
References
- 1. Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021; 71:209–49. https://doi.org/10.3322/caac.21660 [PubMed]
- 2. Joshi SS, Badgwell BD. Current treatment and recent progress in gastric cancer. CA Cancer J Clin. 2021; 71:264–79. https://doi.org/10.3322/caac.21657 [PubMed]
- 3. Correa P. Gastric cancer: overview. Gastroenterol Clin North Am. 2013; 42:211–7. https://doi.org/10.1016/j.gtc.2013.01.002 [PubMed]
- 4. Liang XH, Yu MZ, Shi HL, Wu XJ. [Galangin enhances autophagy by inhibiting NF-κB pathway in gastric cancer MGC-803 cells]. Zhongguo Zhong Yao Za Zhi. 2021; 46:4167–74. https://doi.org/10.19540/j.cnki.cjcmm.20210406.401 [PubMed]
- 5. Xu M, Ren L, Fan J, Huang L, Zhou L, Li X, Ye X. Berberine inhibits gastric cancer development and progression by regulating the JAK2/STAT3 pathway and downregulating IL-6. Life Sci. 2022; 290:120266. https://doi.org/10.1016/j.lfs.2021.120266 [PubMed]
- 6. Li X, Chen W, Miao L, Sun H, Gao X, Guo S, Zhang Y, Yang Y, Guo J. Antitumor Effect of Si-Jun-Zi Decoction on SGC7901 Gastric Cancer Cells by CMTM2 Activation. Evid Based Complement Alternat Med. 2022; 2022:4675815. https://doi.org/10.1155/2022/4675815 [PubMed]
- 7. Zhao AG, Zhao HL, Jin XJ, Yang JK, Tang LD. Effects of Chinese Jianpi herbs on cell apoptosis and related gene expression in human gastric cancer grafted onto nude mice. World J Gastroenterol. 2002; 8:792–6. https://doi.org/10.3748/wjg.v8.i5.792 [PubMed]
- 8. Sun X, Xue D, Zhang K, Jiang F, Li D. Acrid-release and bitter-downbearing therapy and banxia xiexin decoction regulate Wnt/β-catenin pathway, inhibit proliferation and invasion, and induce apoptosis in gastric cancer cells. Am J Transl Res. 2021; 13:6211–20. [PubMed]
- 9. Sun MY, Sun J, Tao J, Yuan YX, Ni ZH, Tang QF. Zuo Jin Wan Reverses DDP Resistance in Gastric Cancer through ROCK/PTEN/PI3K Signaling Pathway. Evid Based Complement Alternat Med. 2018; 2018:4278568. https://doi.org/10.1155/2018/4278568 [PubMed]
- 10. Pan B, Wang Y, Wu C, Jia J, Huang C, Fang S, Liu L. A Mechanism of Action Study on Danggui Sini Decoction to Discover Its Therapeutic Effect on Gastric Cancer. Front Pharmacol. 2021; 11:592903. https://doi.org/10.3389/fphar.2020.592903 [PubMed]
- 11. Bi X, Gong M, Di L. Review on prescription compatibility of shaoyao gancao decoction and reflection on pharmacokinetic compatibility mechanism of traditional Chinese medicine prescription based on in vivo drug interaction of main efficacious components. Evid Based Complement Alternat Med. 2014; 2014:208129. https://doi.org/10.1155/2014/208129 [PubMed]
- 12. Ding Z, Xu F, Sun Q, Li B, Liang N, Chen J, Yu S. Exploring the Mechanism of Action of Herbal Medicine (Gan-Mai-Da-Zao Decoction) for Poststroke Depression Based on Network Pharmacology and Molecular Docking. Evid Based Complement Alternat Med. 2021; 2021:2126967. https://doi.org/10.1155/2021/2126967 [PubMed]
- 13. Wu TT, Lu J, Zheng PQ, Liu SL, Wu J, Sun W, Sun QM, Ma NX, Ding XL, Chen M, Zou X. Yiqi Huayu Jiedu Decoction Inhibits the Invasion and Metastasis of Gastric Cancer Cells through TGF-β/Smad Pathway. Evid Based Complement Alternat Med. 2017; 2017:1871298. https://doi.org/10.1155/2017/1871298 [PubMed] Erratum in: Evid Based Complement Alternat Med. 2021; 2021:4589203. https://doi.org/10.1155/2021/4589203 [PubMed]
- 14. Cats A, Jansen EPM, van Grieken NCT, Sikorska K, Lind P, Nordsmark M, Meershoek-Klein Kranenbarg E, Boot H, Trip AK, Swellengrebel HAM, van Laarhoven HWM, Putter H, van Sandick JW, et al., and CRITICS investigators. Chemotherapy versus chemoradiotherapy after surgery and preoperative chemotherapy for resectable gastric cancer (CRITICS): an international, open-label, randomised phase 3 trial. Lancet Oncol. 2018; 19:616–28. https://doi.org/10.1016/S1470-2045(18)30132-3 [PubMed]
- 15. Rather RA, Bhagat M. Quercetin as an innovative therapeutic tool for cancer chemoprevention: Molecular mechanisms and implications in human health. Cancer Med. 2020; 9:9181–92. https://doi.org/10.1002/cam4.1411 [PubMed]
- 16. Li L, Jin XJ, Li JW, Li CH, Zhou SY, Li JJ, Feng CQ, Liu DL, Liu YQ. Systematic insight into the active constituents and mechanism of Guiqi Baizhu for the treatment of gastric cancer. Cancer Sci. 2021; 112:1772–84. https://doi.org/10.1111/cas.14851 [PubMed]
- 17. Shang HS, Lu HF, Lee CH, Chiang HS, Chu YL, Chen A, Lin YF, Chung JG. Quercetin induced cell apoptosis and altered gene expression in AGS human gastric cancer cells. Environ Toxicol. 2018; 33:1168–81. https://doi.org/10.1002/tox.22623 [PubMed]
- 18. Wang F, Wang L, Qu C, Chen L, Geng Y, Cheng C, Yu S, Wang D, Yang L, Meng Z, Chen Z. Kaempferol induces ROS-dependent apoptosis in pancreatic cancer cells via TGM2-mediated Akt/mTOR signaling. BMC Cancer. 2021; 21:396. https://doi.org/10.1186/s12885-021-08158-z [PubMed]
- 19. Chen AY, Chen YC. A review of the dietary flavonoid, kaempferol on human health and cancer chemoprevention. Food Chem. 2013; 138:2099–107. https://doi.org/10.1016/j.foodchem.2012.11.139 [PubMed]
- 20. Song H, Bao J, Wei Y, Chen Y, Mao X, Li J, Yang Z, Xue Y. Kaempferol inhibits gastric cancer tumor growth: An in vitro and in vivo study. Oncol Rep. 2015; 33:868–74. https://doi.org/10.3892/or.2014.3662 [PubMed]
- 21. Kim TW, Lee SY, Kim M, Cheon C, Ko SG. Kaempferol induces autophagic cell death via IRE1-JNK-CHOP pathway and inhibition of G9a in gastric cancer cells. Cell Death Dis. 2018; 9:875. https://doi.org/10.1038/s41419-018-0930-1 [PubMed]
- 22. Tan M, Zhu J, Pan Y, Chen Z, Liang H, Liu H, Wang H. Synthesis, cytotoxic activity, and DNA binding properties of copper (II) complexes with hesperetin, naringenin, and apigenin. Bioinorg Chem Appl. 2009; 2009:347872. https://doi.org/10.1155/2009/347872 [PubMed]
- 23. Bao L, Liu F, Guo HB, Li Y, Tan BB, Zhang WX, Peng YH. Naringenin inhibits proliferation, migration, and invasion as well as induces apoptosis of gastric cancer SGC7901 cell line by downregulation of AKT pathway. Tumour Biol. 2016; 37:11365–74. https://doi.org/10.1007/s13277-016-5013-2 [PubMed]
- 24. Sun Y, Liu Y, Ma X, Hu H. The Influence of Cell Cycle Regulation on Chemotherapy. Int J Mol Sci. 2021; 22:6923. https://doi.org/10.3390/ijms22136923 [PubMed]
- 25. Ye K, Wei Q, Gong Z, Huang Y, Liu H, Li Y, Peng X. Effect of norcantharidin on the proliferation, apoptosis, and cell cycle of human mesangial cells. Ren Fail. 2017; 39:458–64. https://doi.org/10.1080/0886022X.2017.1308257 [PubMed]
- 26. Vanhaesebroeck B, Guillermet-Guibert J, Graupera M, Bilanges B. The emerging mechanisms of isoform-specific PI3K signalling. Nat Rev Mol Cell Biol. 2010; 11:329–41. https://doi.org/10.1038/nrm2882 [PubMed]
- 27. Janku F. Phosphoinositide 3-kinase (PI3K) pathway inhibitors in solid tumors: From laboratory to patients. Cancer Treat Rev. 2017; 59:93–101. https://doi.org/10.1016/j.ctrv.2017.07.005 [PubMed]
- 28. Faes S, Dormond O. PI3K and AKT: Unfaithful Partners in Cancer. Int J Mol Sci. 2015; 16:21138–52. https://doi.org/10.3390/ijms160921138 [PubMed]
- 29. Guo H, German P, Bai S, Barnes S, Guo W, Qi X, Lou H, Liang J, Jonasch E, Mills GB, Ding Z. The PI3K/AKT Pathway and Renal Cell Carcinoma. J Genet Genomics. 2015; 42:343–53. https://doi.org/10.1016/j.jgg.2015.03.003 [PubMed]
- 30. Fruman DA, Chiu H, Hopkins BD, Bagrodia S, Cantley LC, Abraham RT. The PI3K Pathway in Human Disease. Cell. 2017; 170:605–35. https://doi.org/10.1016/j.cell.2017.07.029 [PubMed]
- 31. Robbins HL, Hague A. The PI3K/Akt Pathway in Tumors of Endocrine Tissues. Front Endocrinol (Lausanne). 2016; 6:188. https://doi.org/10.3389/fendo.2015.00188 [PubMed]
- 32. Jiang T, Xia Y, Lv J, Li B, Li Y, Wang S, Xuan Z, Xie L, Qiu S, He Z, Wang L, Xu Z. A novel protein encoded by circMAPK1 inhibits progression of gastric cancer by suppressing activation of MAPK signaling. Mol Cancer. 2021; 20:66. https://doi.org/10.1186/s12943-021-01358-y [PubMed]
- 33. Song Y, Chang L, Wang X, Tan B, Li J, Zhang J, Zhang F, Zhao L, Liu G, Huo B. Regulatory Mechanism and Experimental Verification of Patchouli Alcohol on Gastric Cancer Cell Based on Network Pharmacology. Front Oncol. 2021; 11:711984. https://doi.org/10.3389/fonc.2021.711984 [PubMed]
- 34. Guo C, Hou J, Ao S, Deng X, Lyu G. HOXC10 up-regulation promotes gastric cancer cell proliferation and metastasis through MAPK pathway. Chin J Cancer Res. 2017; 29:572–80. https://doi.org/10.21147/j.issn.1000-9604.2017.06.12 [PubMed]
- 35. Burotto M, Chiou VL, Lee JM, Kohn EC. The MAPK pathway across different malignancies: a new perspective. Cancer. 2014; 120:3446–56. https://doi.org/10.1002/cncr.28864 [PubMed]
- 36. Sun Y, Liu WZ, Liu T, Feng X, Yang N, Zhou HF. Signaling pathway of MAPK/ERK in cell proliferation, differentiation, migration, senescence and apoptosis. J Recept Signal Transduct Res. 2015; 35:600–4. https://doi.org/10.3109/10799893.2015.1030412 [PubMed]
- 37. Liu F, Yang X, Geng M, Huang M. Targeting ERK, an Achilles’ Heel of the MAPK pathway, in cancer therapy. Acta Pharm Sin B. 2018; 8:552–62. https://doi.org/10.1016/j.apsb.2018.01.008 [PubMed]
- 38. Papa S, Choy PM, Bubici C. The ERK and JNK pathways in the regulation of metabolic reprogramming. Oncogene. 2019; 38:2223–40. https://doi.org/10.1038/s41388-018-0582-8 [PubMed]
- 39. Bubici C, Papa S. JNK signalling in cancer: in need of new, smarter therapeutic targets. Br J Pharmacol. 2014; 171:24–37. https://doi.org/10.1111/bph.12432 [PubMed]
- 40. Pua LJW, Mai CW, Chung FF, Khoo AS, Leong CO, Lim WM, Hii LW. Functional Roles of JNK and p38 MAPK Signaling in Nasopharyngeal Carcinoma. Int J Mol Sci. 2022; 23:1108. https://doi.org/10.3390/ijms23031108 [PubMed]
- 41. Ru J, Li P, Wang J, Zhou W, Li B, Huang C, Li P, Guo Z, Tao W, Yang Y, Xu X, Li Y, Wang Y, Yang L. TCMSP: a database of systems pharmacology for drug discovery from herbal medicines. J Cheminform. 2014; 6:13. https://doi.org/10.1186/1758-2946-6-13 [PubMed]
- 42. Luo Z, Huang J, Li E, He X, Meng Q, Huang X, Shen X, Yan C. An Integrated Pharmacology-Based Strategy to Investigate the Potential Mechanism of Xiebai San in Treating Pediatric Pneumonia. Front Pharmacol. 2022; 13:784729. https://doi.org/10.3389/fphar.2022.784729 [PubMed]
- 43. Fishilevich S, Zimmerman S, Kohn A, Iny Stein T, Olender T, Kolker E, Safran M, Lancet D. Genic insights from integrated human proteomics in GeneCards. Database (Oxford). 2016; 2016:baw030. https://doi.org/10.1093/database/baw030 [PubMed]
- 44. Piñero J, Bravo À, Queralt-Rosinach N, Gutiérrez-Sacristán A, Deu-Pons J, Centeno E, García-García J, Sanz F, Furlong LI. DisGeNET: a comprehensive platform integrating information on human disease-associated genes and variants. Nucleic Acids Res. 2017; 45:D833–9. https://doi.org/10.1093/nar/gkw943 [PubMed]
- 45. Wishart DS, Feunang YD, Guo AC, Lo EJ, Marcu A, Grant JR, Sajed T, Johnson D, Li C, Sayeeda Z, Assempour N, Iynkkaran I, Liu Y, et al. DrugBank 5.0: a major update to the DrugBank database for 2018. Nucleic Acids Res. 2018; 46:D1074–82. https://doi.org/10.1093/nar/gkx1037 [PubMed]
- 46. Zhang MM, Wang D, Lu F, Zhao R, Ye X, He L, Ai L, Wu CJ. Identification of the active substances and mechanisms of ginger for the treatment of colon cancer based on network pharmacology and molecular docking. BioData Min. 2021; 14:1. https://doi.org/10.1186/s13040-020-00232-9 [PubMed]
- 47. Szklarczyk D, Franceschini A, Wyder S, Forslund K, Heller D, Huerta-Cepas J, Simonovic M, Roth A, Santos A, Tsafou KP, Kuhn M, Bork P, Jensen LJ, von Mering C. STRING v10: protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015; 43:D447–52. https://doi.org/10.1093/nar/gku1003 [PubMed]
- 48. Tang Y, Li M, Wang J, Pan Y, Wu FX. CytoNCA: a cytoscape plugin for centrality analysis and evaluation of protein interaction networks. Biosystems. 2015; 127:67–72. https://doi.org/10.1016/j.biosystems.2014.11.005 [PubMed]
- 49. Zhou Y, Zhou B, Pache L, Chang M, Khodabakhshi AH, Tanaseichuk O, Benner C, Chanda SK. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat Commun. 2019; 10:1523. https://doi.org/10.1038/s41467-019-09234-6 [PubMed]
- 50. Chen L, Zhang YH, Wang S, Zhang Y, Huang T, Cai YD. Prediction and analysis of essential genes using the enrichments of gene ontology and KEGG pathways. PLoS One. 2017; 12:e0184129. https://doi.org/10.1371/journal.pone.0184129 [PubMed]
- 51. Kim S, Thiessen PA, Bolton EE, Chen J, Fu G, Gindulyte A, Han L, He J, He S, Shoemaker BA, Wang J, Yu B, Zhang J, Bryant SH. PubChem Substance and Compound databases. Nucleic Acids Res. 2016; 44:D1202–13. https://doi.org/10.1093/nar/gkv951 [PubMed]
- 52. Huang S, Chen Y, Pan L, Fei C, Wang N, Chu F, Peng D, Duan X, Wang Y. Exploration of the Potential Mechanism of Tao Hong Si Wu Decoction for the Treatment of Breast Cancer Based on Network Pharmacology and In Vitro Experimental Verification. Front Oncol. 2021; 11:731522. https://doi.org/10.3389/fonc.2021.731522 [PubMed]
- 53. Chiu YJ, Lin CH, Lee MC, Hsieh-Li HM, Chen CM, Wu YR, Chang KH, Lee-Chen GJ. Formulated Chinese medicine Shaoyao Gancao Tang reduces NLRP1 and NLRP3 in Alzheimer’s disease cell and mouse models for neuroprotection and cognitive improvement. Aging (Albany NY). 2021; 13:15620–37. https://doi.org/10.18632/aging.203125 [PubMed]