



Introduction
Renal cell carcinoma (RCC), originating from the renal epithelial cells, is a common malignant solid tumor of urinary system, accounting for 80-90% of all kidney malignancies [1, 2]. RCC tumors are extremely heterogenous with multiple histological subtypes, among which clear cell renal cell carcinoma (ccRCC) is the most frequent one, making up nearly 70% of all kidney malignancies [3]. With the ability to invade renal sinus and extend to the renal vein, ccRCC is characterized by high metastatic potential, which causes approximately 30% of ccRCC patients possessing metastatic disorders at the first time of diagnosis [4]. Therefore, it makes sense to uncover the molecular mechanism of ccRCC metastasis.
Circular RNAs (circRNAs) are a class of novel, endogenous and noncoding RNAs with the feature of covalently closed loops [5, 6]. CircRNAs are more stable than their linear counterparts as they have no 5’-cap and 3’-polyadenylated tail structure [7, 8]. During the past years, increasing lines of evidence have found that circRNAs play key roles in initiation and progression of multiple human malignancies, including breast cancer [9, 10], papillary thyroid carcinoma [11, 12], hepatocellular carcinoma [13, 14] and colorectal cancer [15, 16]. Intriguingly, several circRNAs have also been reported to participate in biological process of ccRCC metastasis. For example, circ-TNPO3 hindered ccRCC metastasis by binding to IGF2BP2 [17]; circular RNA ITCH suppressed ccRCC metastasis by regulating miR-106b-5p/PDCD4 axis [18]; circ-AKT3 inhibited ccRCC metastasis through changing miR-296-3p/E-cadherin signals [19]. However, to date, our knowledge of circRNAs in metastasis of ccRCC is still inadequate and needs to be further studied.
In this work, we firstly screened out and validated candidate ccRCC metastasis-related circRNAs by analyzing two GEO datasets (GSE100186 and GSE137836). Then, the structural pattern of potential circRNA and its parental gene were obtained. Subsequently, the binding miRNAs of potential circRNA and the downstream target genes of potential circRNA/miRNA axis were predicted and analyzed by using a series of online databases or tools. At the end, a potential circRNA/miRNA/mRNA triple regulatory axis involved in the process of ccRCC metastasis was successfully constructed.
Materials and Methods
Inclusion of datasets
In this study, to explore the role and mechanism of circRNA in malignant metastasis of ccRCC, the potential datasets from the NCBI GEO database (http://www.ncbi.nlm.nih.gov/geo/) were included. The included datasets should meet the following criteria: (1) the included datasets should study the circRNA expression profile in ccRCC and/or normal tissues; (2) the datasets regarding cell lines should be excluded; (3) the datasets focusing on the circRNA expression in metastasis of ccRCC should also be included. At the end, only two datasets, consisting of GSE100186 and GSE137836, met the above selection criteria. Both GSE100186 and GSE137836 were based on the platform of GPL21825 074301 Arraystar Human CircRNA microarray V2. GSE100186 contained 4 ccRCC and 4 matched non-tumor samples and GSE137836 contained 3 primary tumor tissues and 3 ccRCC metastatic tumor tissues from 3 ccRCC patients.
Differential expression analysis
As previously described [11], the online tool GEO2R (http://www.ncbi.nlm.nih.gov/geo/geo2r/) from the NCBI GEO database (http://www.ncbi.nlm.nih.gov/geo/) was employed to obtain differentially expressed circRNAs between ccRCC and normal tissues or metastatic ccRCC by performing differential expression after conducting sample data normalization.
Intersection analysis
VENNY 2.1 (http://bioinfogp.cnb.csic.es/tools/venny) tool was introduced to conduct intersection analysis to acquire the potential circRNAs that were commonly appeared in GSE100186 and GSE137836. Moreover, this tool was also used to obtain the common binding miRNAs of hsa_circ_0037858 from CSCD and starBase databases.
Expression validation for circRNAs
The expression levels of three potential circRNAs related to malignant metastasis of ccRCC were further validated by extracting their expression data from GSE100186 and GSE137836, after which t-test was employed to calculate the expression difference of them in normal, ccRCC and metastatic ccRCC tissues.
circBase analysis
CircBase (http://www.circbase.org/) [20], a database providing scripts to obtain known and novel circRNAs in sequencing data, was introduced to acquire the genome location and parental gene of hsa_circ_0037858.
Cancer-specific CircRNA database (CSCD) analysis
CSCD (https://gb.whu.edu.cn/CSCD) [21], a cancer-specific circRNA database for predicting the potential MREs, RBPs and open reading frame of circRNAs, was utilized to generate the structural pattern and forecast the binding miRNAs of hsa_circ_0037858.
miRNA prediction
In addition to CSCD as mentioned above, another database, namely starBase (http://starbase.sysu.edu.cn/) [22], was also employed to predict the possible miRNAs that could potentially bind to hsa_circ_0037858. Only the binding miRNAs that were commonly appeared in both CSCD and starBase databases were included for subsequent analysis.
Expression determination for miRNAs
The expression levels of binding miRNAs of hsa_circ_0037858 in ccRCC were determined using starBase (http://starbase.sysu.edu.cn/) [22]. Only P-value < 0.05 was considered as statistically significant.
CancerMIRNome analysis
CancerMIRNome (http://bioinfo.jialab-ucr.org/CancerMIRNome) [23] is an integrated analysis and visualization database for miRNome profiles of human cancer, which was introduced to validate the expression of 4 potential binding miRNAs of hsa_circ_0037858 in ccRCC. Furthermore, the database was also used to assess the diagnostic values of them in ccRCC.
Target gene prediction
The possible target genes of miR-5000-3p were predicted by usage of a series of online target gene prediction tools, including PITA, RNA22, miRmap, microT, miRanda, PicTar and TargetScan. Only the predicted target genes that were commonly appeared in more than 2 tools were included for subsequent analysis.
STRING analysis
STRING database (https://string-db.org/) is an online tool aiming to integrate all known and predicted associations between proteins, containing physical interactions and functional associations [24]. The database was used to construct a protein-protein interaction (PPI) regulatory network of target genes of miR-5000-3p.
Identification of hub genes
The hub genes among the target genes of miR-5000-3p were identified with the help of STRING database and Cytoscape software. At the beginning, the interaction gene pairs among the target genes were downloaded from STRING database. Then, these gene pairs were re-entered into Cytoscape software. Finally, the node degree of individual gene was calculated by Cytohubba. According to the node degree, the top 20 hub genes were screened out.
Kaplan-Meier plotter analysis
Kaplan-Meier plotter database (http://kmplot.com/) is capable to access the relationship between the expression levels of all genes (mRNA, miRNA, protein) and survival in more than 20 tumor types, including ccRCC [25]. In this study, this database was utilized to evaluate the prognostic values of the top 20 hub genes in ccRCC.
Correlation analysis
Expression correlation between miRNAs and target genes could provide key clues to identify potential miRNA-target gene pairs based on the negative role of miRNAs in regulating corresponding target genes. Thus, correlation analysis was employed for determining the expression relationship between miR-5000-3p and the top 20 hub genes in ccRCC using starBase database (http://starbase.sysu.edu.cn/) as previously described [26].
Cell lines, cell culture and cell transfection
Human ccRCC cell lines ACHN and Caki-1 were purchased from the Chinese Academy of Sciences (Shanghai, China) and were cultured in DMEM supplemented with 10% FBS under a humidified atmosphere of 5% CO2 at 37° C. circRNA overexpressed plasmid, miRNA mimic and corresponding negative controls were transfected into ccRCC cells by usage of Lipofectamine™ 3000 based on the manufacturer’s instruction.
RNA isolation and qRT-PCR analysis
RNAiso plus Reagent was used to extract the total RNAs from cell lines, after which RNAs were reversely-transcribed into cDNA. Next, PCR was performed in triplicates using a Roche LightCycler480 II Real-Time PCR Detection System by SYBR Premix Ex Taq (TaKaRa, RR420A). Finally, FMR1 expression (relative to GAPDH) was calculated by the method of 2-ddCt.
In vitro migration assay
The in vitro migration assay was conducted by 24-well transwell chambers (Corning, USA). The transfected cells were re-cultured with serum-free medium for 24 hours, after which cells were suspended in 0.2 ml serum-free medium and added into the upper chamber and medium containing 10% FBS was added into the lower chamber following 24-hour culture. Subsequently, the cells on the lower surface of the membrane were fixed and stained by 100% methanol and 0.1% crystal violet, respectively. Finally, the cells were counted under a microscope.
Statistical analysis
Most of the statistical analyses in this study were automatically calculated by the online databases and tools as mentioned above. Expression determination between two groups was analyzed by GraphPad Prism software (Version 7). P-value < 0.05 or logrank P-value < 0.05 was considered as statistically significant.
Availability of data and materials
All data and materials have been provided in the manuscript. Please do not hesitate to contact the corresponding author.
Results
Identification of hsa_circ_0037858 as a potential circRNA related to metastasis of ccRCC
To intend to explore the role and mechanism of circRNA in malignant metastasis of ccRCC, two GEO datasets, consisting of GSE100186 and GSE137836, were included in this study. By using GEO2R tool, data normalization and differential expression analysis for GSE100186 and GSE137836 were performed (Figure 1). A total of 4477 significant differentially expressed circRNAs (DECs) were identified in ccRCC compared with normal controls in GSE100186, consisting of 2281 upregulated and 2196 downregulated DECs (Figure 1B). 1017 DECs were significantly differentially expressed between ccRCC and metastatic ccRCC tissues in GSE137836, containing 567 upregulated and 450 downregulated DECs (Figure 1C). Next, intersection analysis for the two datasets was performed (Figure 2A, 2B). The result suggested that three downregulated circRNAs were commonly appeared in both GSE100186 and GSE137836, including ASCRP3006810, ASCRP3009648 and ASCRP3012612. To improve the analytic accuracy, the expression data of the three potential circRNAs from GSE100186 and GSE137836 were downloaded and re-analyzed (Figure 2C–2G). The results showed that only ASCRP3006810 (hsa_circ_0037858) was markedly downregulated in ccRCC compared with normal controls (Figure 2C) as well as decreased in metastatic ccRCC compared with primary cancer tissues (Figure 2D). All these findings suggested that hsa_circ_0037858 might be a potential tumor suppressive circRNA in metastasis of ccRCC.
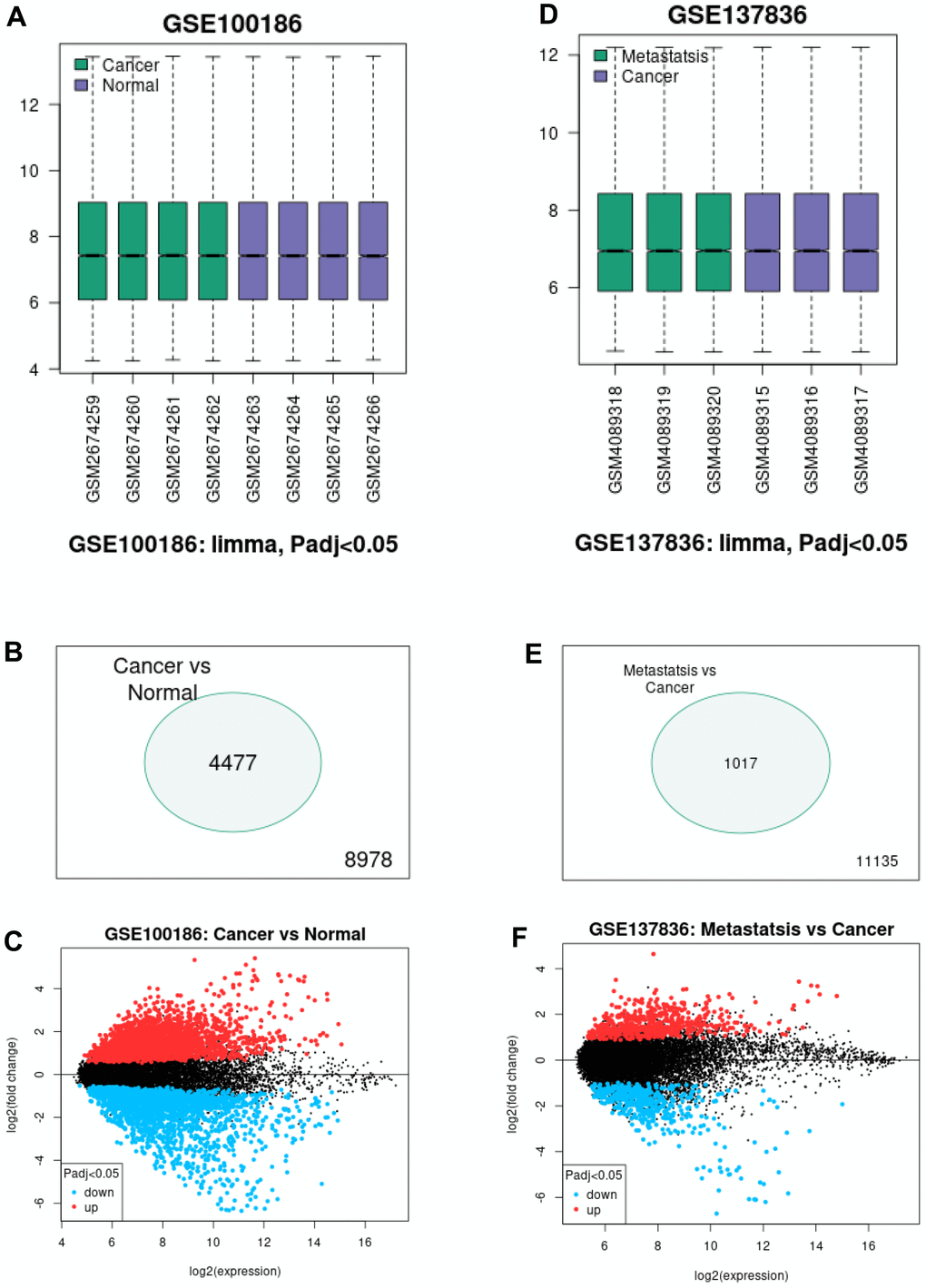
Figure 1. Screening of differentially expressed circRNAs (DECs) associated with malignant metastasis of ccRCC. (A) The normalization of samples in GSE100186 analyzed by GEO2R online tool. (B) The significant DECs between ccRCC cancer tissues and normal tissues. (C) The volcano plot of the DECs between ccRCC cancer tissues and normal tissues in GSE100186. (D) The normalization of samples in GSE137836 analyzed by GEO2R online tool. (E) The significant DECs between ccRCC cancer tissues and metastatic cancer tissues. (F) The volcano plot of the DECs between ccRCC cancer tissues and metastatic cancer tissues in GSE137836.
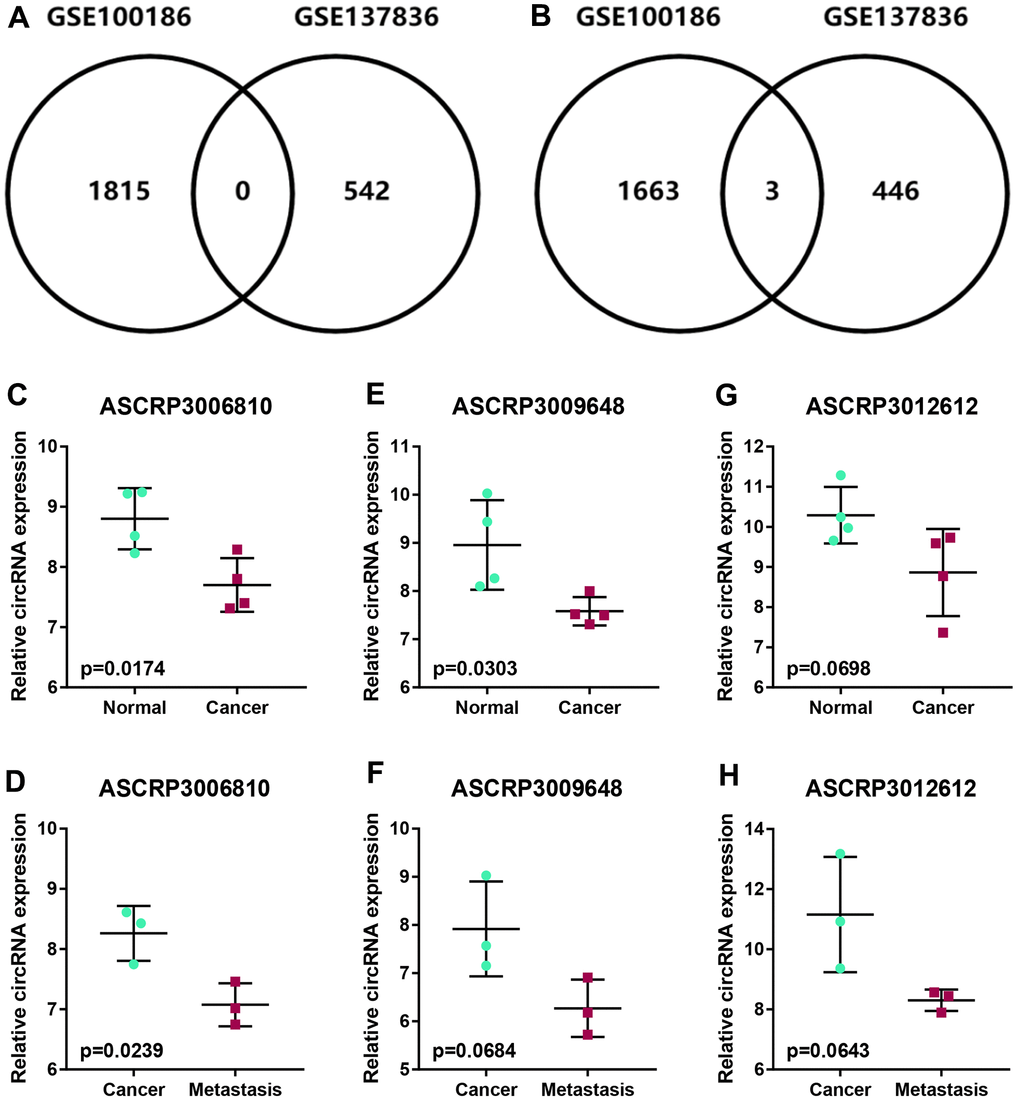
Figure 2. Identification of potential circRNAs associated with metastasis of ccRCC. (A) The intersection analysis for the significant upregulated circRNAs with |log2FC| > 1 in GSE100186 and GSE137836. (B) The intersection analysis for the significant downregulated circRNAs with |log2FC| > 1 in GSE100186 and GSE137836. (C, D) The expression levels of ASCRP3006810 in normal, cancer and metastatic cancer tissues (ccRCC). (E, F) The expression levels of ASCRP3009648 in normal, cancer and metastatic cancer tissues (ccRCC). (G, H) The expression levels of ASCRP3012612 in normal, cancer and metastatic cancer tissues (ccRCC).
The structural characteristics of hsa_circ_0037858
To further understand hsa_circ_0037858, circBase database was employed. The genome position of hsa_circ_0037858 was chr16: 11647388-11650591, and the parental gene of hsa_circ_0037858 was LITAF. Next, Cancer-Specific CircRNA Database (CSCD) database was utilized to generate the structural pattern of hsa_circ_0037858. As shown in Figure 3, hsa_circ_0037858 was generated by exons 13, 14, 15, 16 and 17. Intriguingly, hsa_circ_0037858 possessed several microRNA response elements, RNA binding protein binding domain and open reading frame, representing three potential action mechanisms of circRNA.
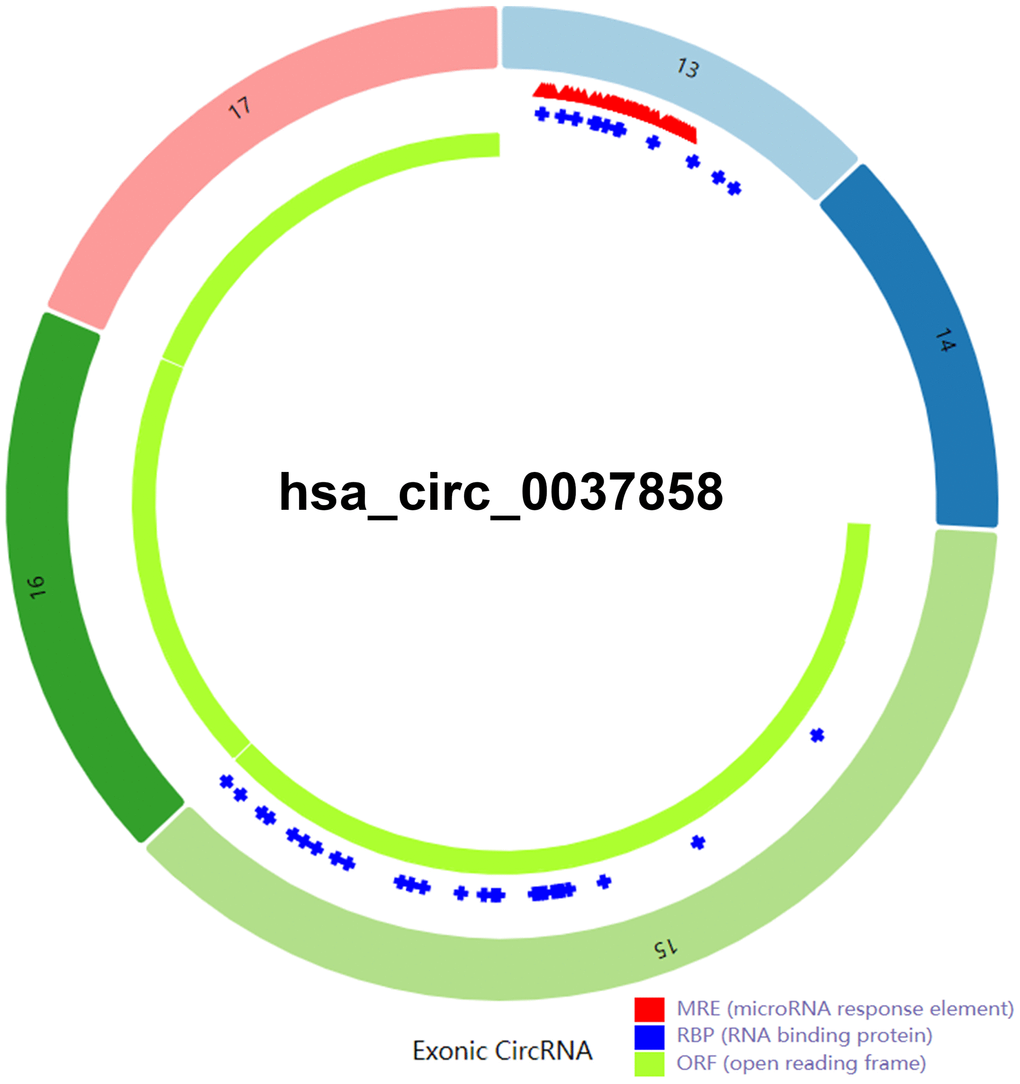
Figure 3. The structural pattern of exonic hsa_circ_0037858, consisting of exon 13, 14, 15, 16 and 17.
Identification of 4 potential binding miRNAs of hsa_circ_0037858
It has been widely acknowledged that miRNA binding is the main classic action mechanism of circRNA. Two databases, consisting of CSCD and starBase, were introduced to predict the binding miRNAs of hsa_circ_0037858. A total of 98 and 16 miRNAs of hsa_circ_0037858 were forecasted in CSCD and starBase, respectively. To better visualization, hsa_circ_0037858-miRNA regulatory networks were constructed as presented in Figure 4A, 4B. Subsequently, intersection analysis for miRNAs of hsa_circ_0037858 from CSCD and starBase databases were performed (Figure 4C). The results indicated that 4 miRNAs, consisting of miR-3064-5p, miR-6504-5p, miR-345-5p and miR-5000-3p, were commonly appeared in CSCD and starBase databases (Figure 4D). Taken together, miR-3064-5p, miR-6504-5p, miR-345-5p and miR-5000-3p might be the most potential binding miRNAs of hsa_circ_0037858.
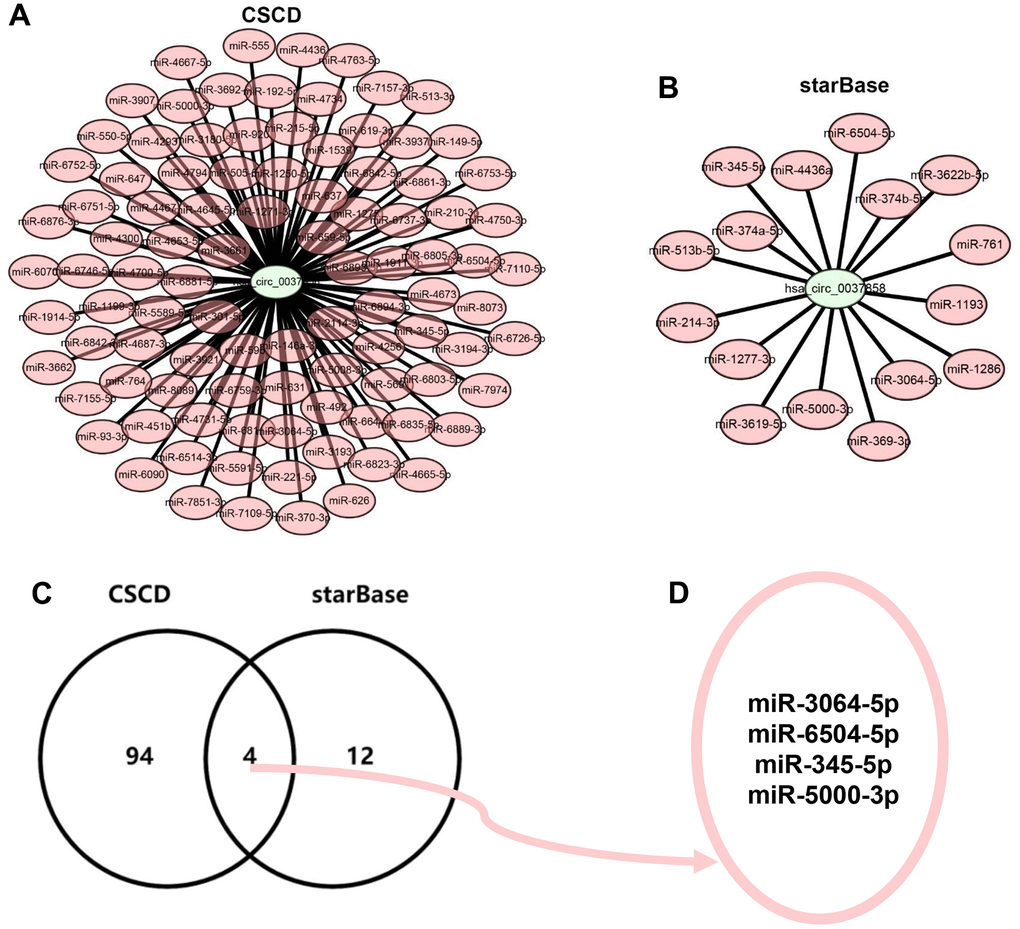
Figure 4. Identification of potential binding miRNAs of hsa_circ_0037858. (A) The binding miRNAs of hsa_circ_0037858 predicted by CSCD database. (B) The binding miRNAs of hsa_circ_0037858 predicted by starBase database. (C) The intersection analysis for the binding miRNAs predicted by CSCD and starBase databases. (D) miR-3064-5p, miR-6504-5p, miR-345-5p and miR-5000-3p are commonly appeared in CSCD and starBase databases.
Analysis for the 4 potential binding miRNAs of hsa_circ_0037858 in ccRCC
Expression analysis for the four potential binding miRNAs of hsa_circ_0037858 in ccRCC was conducted using starBase database. As shown in Figure 5A–5D, among the 4 miRNAs, miR-345-5p was significantly downregulated and miR-5000-3p were markedly upregulated in ccRCC when compared with normal controls. Next, CancerMIRNome database was used to validate the expression levels of miR-3064-5p, miR-6504-5p, miR-345-5p and miR-5000-3p in ccRCC (Figure 5E–5H). Identical with the analytic result from starBase, miR-345-5p and miR-5000-3p were statistically downregulated and upregulated in ccRCC when compared with normal controls, respectively. Subsequently, ROC curve analysis by CancerMIRNome database was employed to assess the diagnostic values of miR-3064-5p, miR-6504-5p, miR-345-5p and miR-5000-3p in ccRCC (Figure 5I–5L). The results showed that only 2 of 4 miRNAs (miR-345-5p and miR-5000-3p) possessed the obvious abilities to distinguish ccRCC tissues from normal tissues. All these findings suggested that miR-5000-3p might be the most potential binding miRNA of downregulated hsa_circ_0037858 in ccRCC.
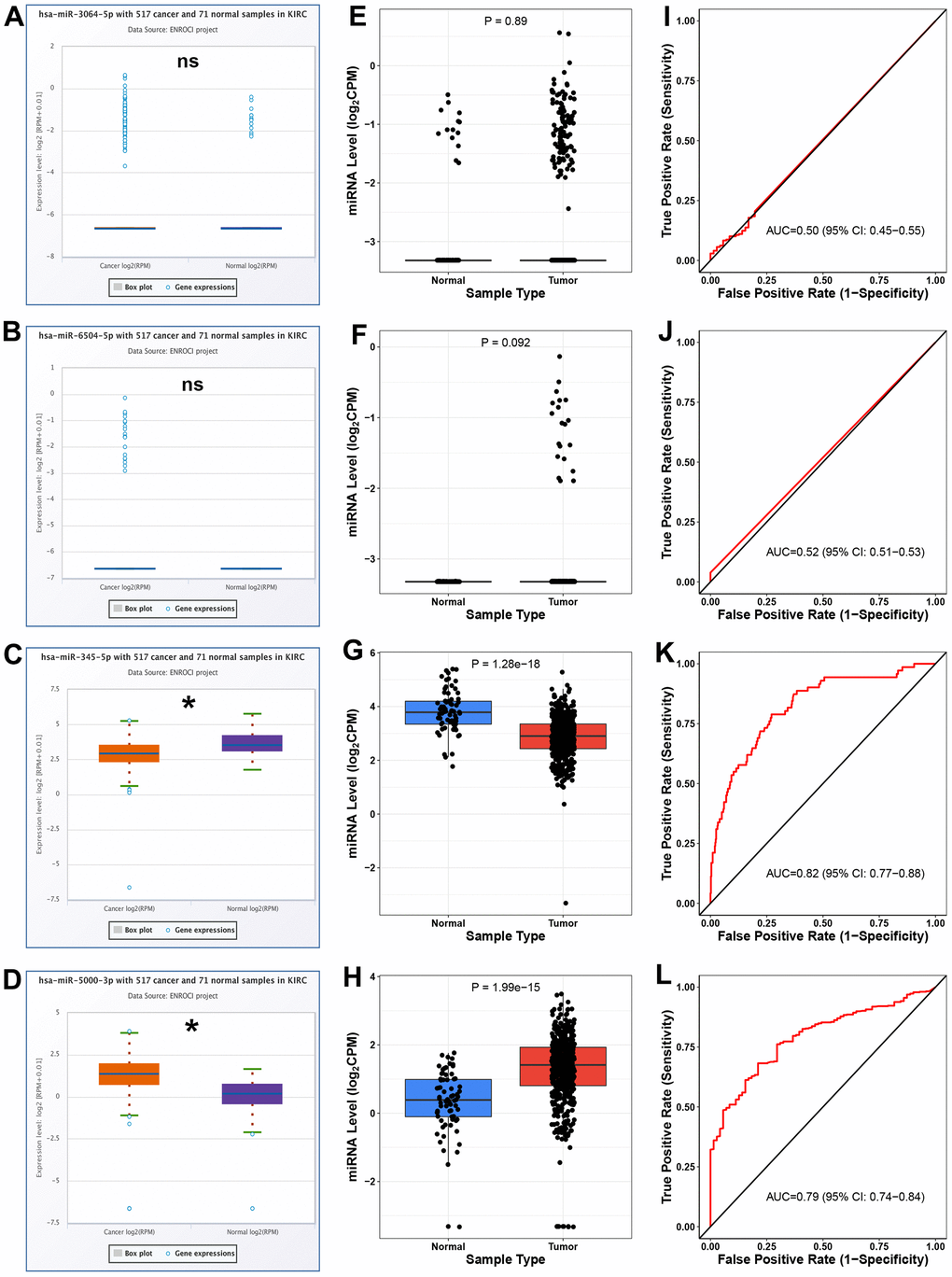
Figure 5. Analysis for the expression and diagnostic values of 4 potential miRNAs of hsa_circ_0037858 in ccRCC. The expression levels of miR-3064-5p (A), miR-6504-5p (B), miR-345-5p (C) and miR-5000-3p (D) in ccRCC compared with normal controls analyzed by starBase. The expression levels of miR-3064-5p (E), miR-6504-5p (F), miR-345-5p (G) and miR-5000-3p (H) in ccRCC compared with normal controls analyzed by CancerMIRNome. The ROC curves of miR-3064-5p (I), miR-6504-5p (J), miR-345-5p (K) and miR-5000-3p (L) in ccRCC analyzed by CancerMIRNome. nsP>0.05; *P<0.05.
Protein-protein interaction (PPI) analysis for the target genes of miR-5000-3p
The target genes of miR-5000-3p were predicted by using a series of target gene prediction programs. Consequently, a total of 327 target genes were found to potentially bind to miR-5000-3p. To understand the molecular interaction of these target genes, STRING database was employed. As presented in Figure 6A, a PPI regulatory network of miR-5000-3p’s target genes was constructed. Next, the interaction gene pairs from STRING database were re-entered into Cytoscape software. Using CytoHubba, the top 20 hub genes among all the target genes of miR-5000-3p were identified by calculating node degree, and a sub-PPI network of the top 20 hub genes were re-established as shown in Figure 6B. As presented in Figure 6C, according to node degree, MYC, RHOA, NCL, FMR1 and AGO1 ranked as the top 5 hub genes, which might act in metastasis of ccRCC.
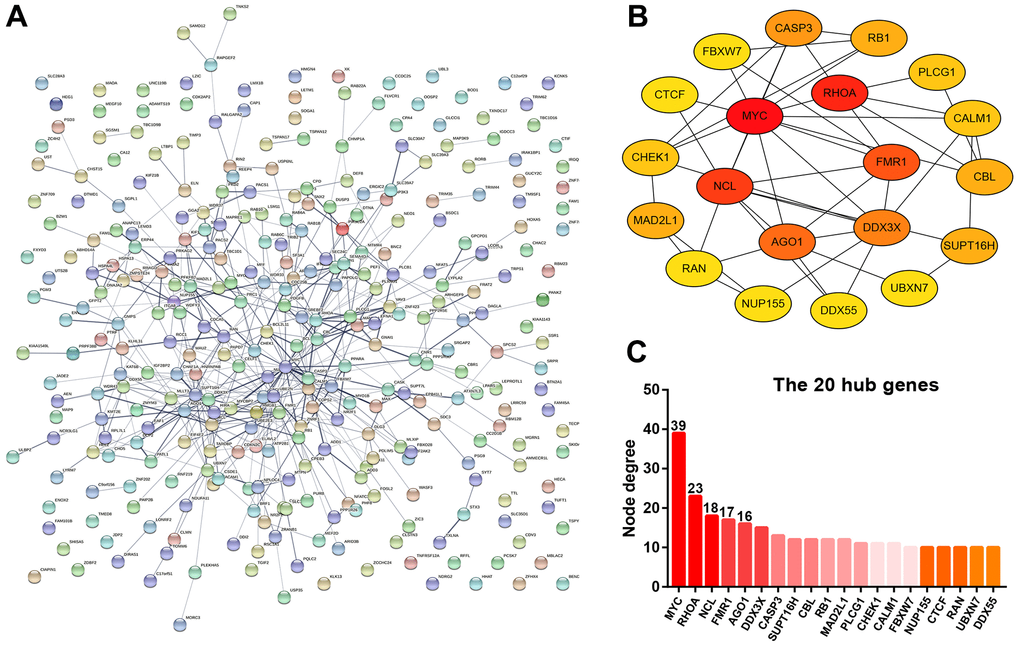
Figure 6. Protein-protein interaction (PPI) analysis for the target genes of miR-5000-3p. (A) The PPI network of target genes of miR-5000-3p established by STRING database. (B) The sub-PPI network of the top 20 hub genes among the target genes of miR-5000-3p constructed by Cytoscape. (C) The presentation of the top 20 hub genes and their corresponding node degree.
Analysis for the top 20 hub genes in ccRCC
Expression, survival and correlation analyses for the top 20 hub genes in ccRCC were successively performed (Figure 7). Expression analysis revealed that 11 genes (MYC, CASP3, CBL, RB1, MAD2L1, PLCG1, CHEK1, FBXW7, NUP155, RAN and DDX55) and 7 genes (RHOA, FMR1, SUPT16H, CALM1, CTCF and UBXN7) were significantly upregulated and downregulated in ccRCC when compared with normal controls, respectively. Survival analysis showed that 8 genes (RHOA, CASP3, MAD2L1, PLCG1, CHEK1, FBXW7, RAN and DDX55) and 11 genes (NCL, FMR1, AGO1, DDX3X, SUPT16H, CBL, RB1, CALM1, NUP155, CTCF and UBXN7) indicated poor and good prognosis in ccRCC patients, respectively. Correlation analysis demonstrated that only two genes (FMR1 and CASP3) were significantly negatively associated with miR-5000-3p in ccRCC. Taken together, FMR1 might be the most potential downstream gene of hsa_circ_0037858/miR-5000-3p axis related to metastasis of ccRCC.
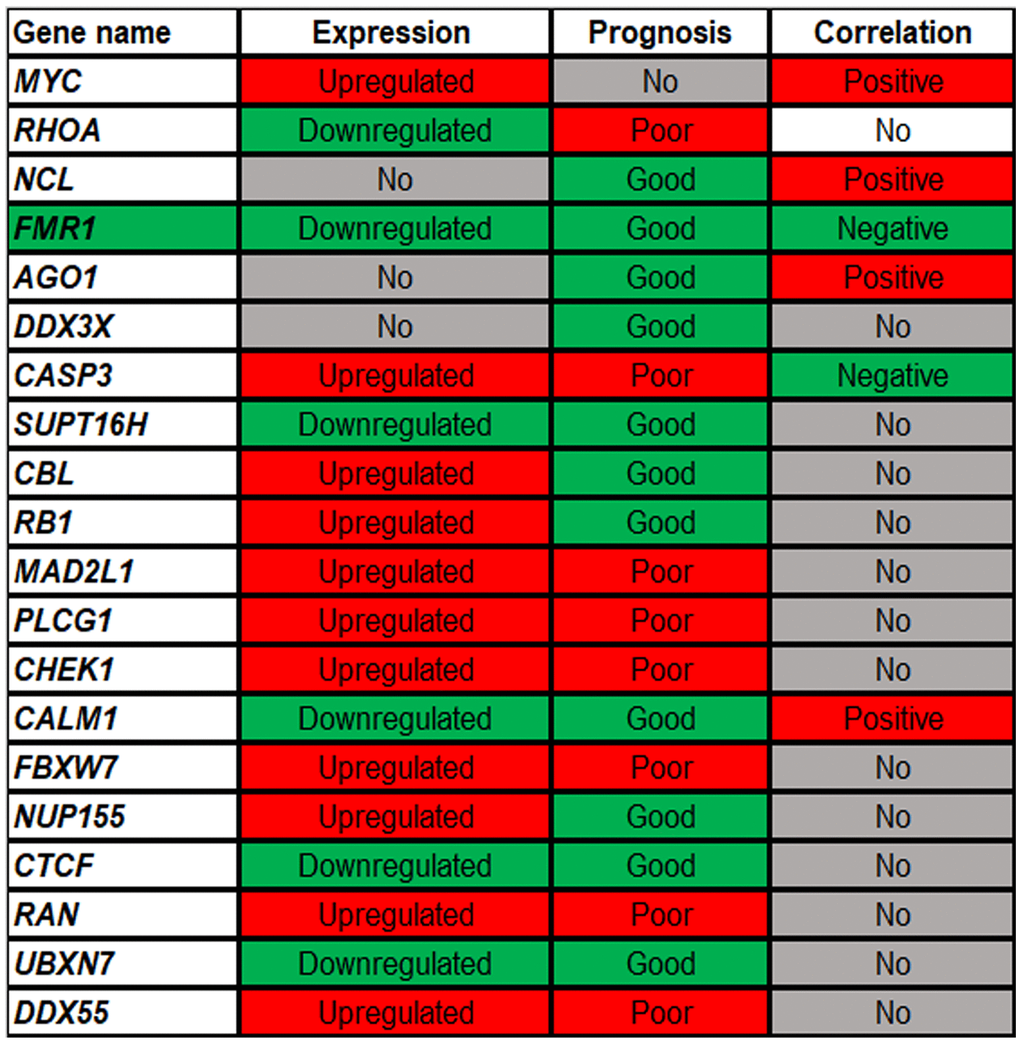
Figure 7. The expression, prognosis and correlation landscape of the top 20 hub genes of the target genes of miR-5000-3p in ccRCC. “Upregulated” and “Downregulated” mean that the expression of genes is significantly increased and decreased in ccRCC, respectively; “poor” and “good” represent the prognostic values of genes in ccRCC. “positive” and “negative” suggest the correlation relationship of gene-miRNA pairs in ccRCC.
Establishment of a potential hsa_circ_0037858/miR-5000-3p/FMR1 axis related to metastasis of ccRCC
To preliminarily validate the identified circRNA/miRNA/mRNA axis, qRT-PCR assay was employed. As presented in Figure 8A, 8B, overexpression of hsa_circ_0037858 could significantly increase FMR1 expression in both ACHN and Caki-1 cells. However, this upregulation could be markedly reversed after miR-5000-3p overexpression in ccRCC. Functional assay showed that increased expression of hsa_circ_0037858 significantly inhibited in vitro metastasis of ACHN and Caki-1 cells (Figure 8C–8F). Moreover, upregulation of miR-5000-3p could weaken hsa_circ_0037858-mediated inhibition of in vitro metastasis of ccRCC cells. Taken all these findings into consideration, hsa_circ_0037858/miR-5000-3p/FMR1 axis might be a potential pathway involved in suppression of ccRCC metastasis as shown in Figure 8G.
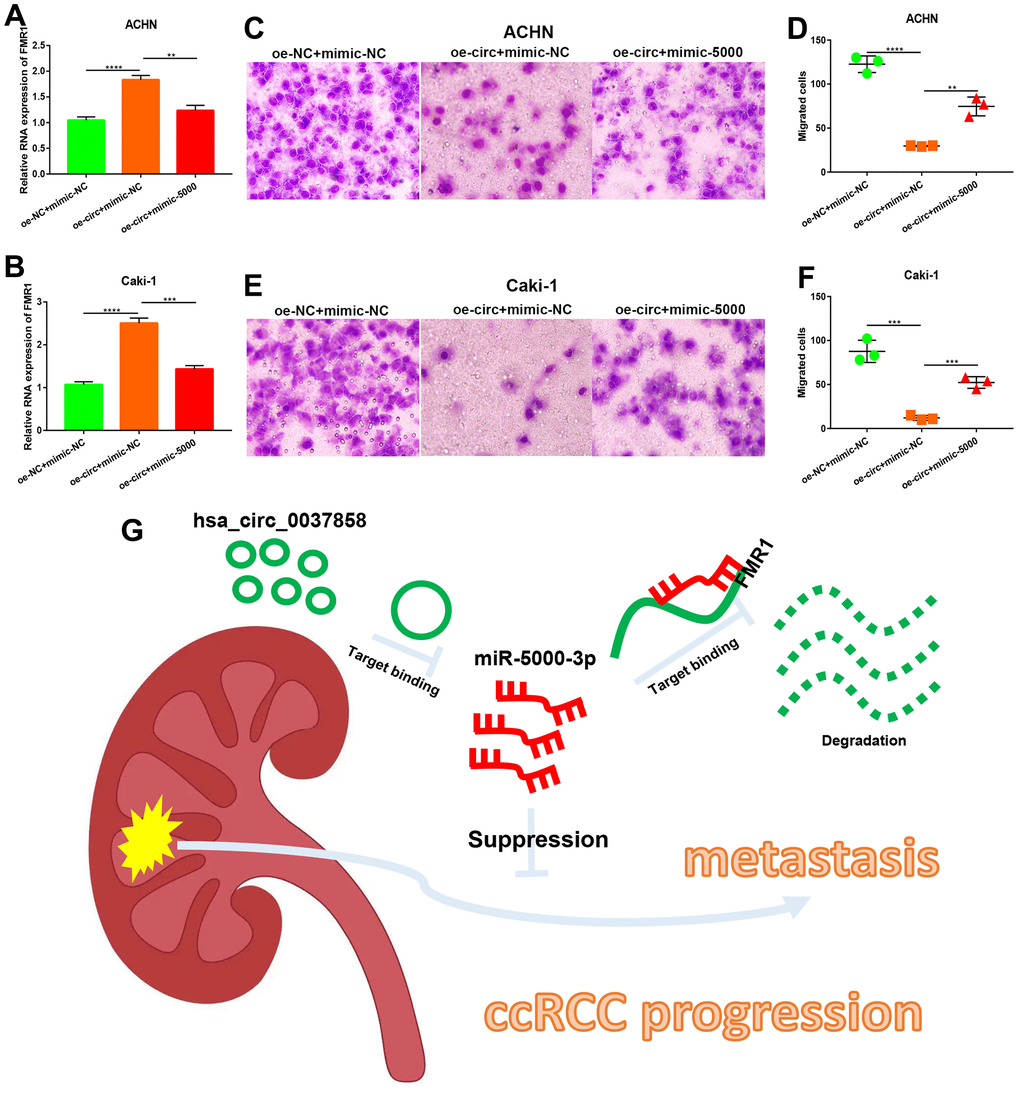
Figure 8. Identification of a potential hsa_circ_0037858/miR-5000-3p/FMR1 axis contributing to metastasis of ccRCC. The regulatory role of hsa_circ_0037858/miR-5000-5p in FMR1 expression in ACHN (A) or Caki-1 (B). The inhibition of hsa_circ_0037858 in in vitro migration could be reversed by miR-5000-3p overexpression in ACHN (C, D) and Caki-1 (E, F) cell lines (G) The action model of hsa_circ_0037858/miR-5000-3p/FMR1 axis in metastasis of ccRCC. **P<0.01; ***P<0.001; ****P<0.0001.
Discussion
ccRCC is the most frequent histological subtype among all kidney malignancies, which is notorious for its high metastatic potential. Recently, numerous studies have reported the critical role of circRNA in cancer initiation and progression. However, the knowledge of circRNA in ccRCC metastasis is still inadequate and needs to be further explored.
At the beginning of this study, the circRNAs that might be associated with metastasis of ccRCC were screened out by combination of two GEO datasets, consisting of GSE100186 and GSE137836. After performing expression validation, hsa_circ_0037858 that was significantly downregulated in ccRCC was selected as the most potential circRNA involved in ccRCC metastasis. A previous study showed that hsa_circ_0037858 might function as a key regulator of progression of lumbar disc degeneration [27]. However, to date, its role in cancer, including ccRCC, has not been reported.
It has been widely acknowledged that miRNAs are involved in circRNA’s biological effects [28–30]. Intriguingly, CSCD database [21] analysis revealed that hsa_circ_0037858 possessed potential microRNA response elements. After performing binding miRNA prediction, miR-3064-5p, miR-6504-5p, miR-345-5p and miR-5000-3p were selected as candidate binding miRNAs of hsa_circ_0037858.
Based on competing endogenous RNA hypothesis [31], the binding miRNA of downregulated hsa_circ_0037858 should be upregulated in ccRCC. Consequently, among the 4 miRNAs, only miR-5000-3p expression was markedly increased in ccRCC compared with normal controls. Several studies have confirmed the oncogenic role of miR-5000-3p in human malignancies. For example, Zhuang et al. suggested that miR-5000-3p facilitated oxaliplatin resistance by targeting USP49 in colorectal cancer [32]; Chen et al. indicated that overexpression of miR-5000-3p enhanced proliferation and migration of laryngocarcinoma [33]. Furthermore, ROC curve analysis demonstrated that miR-5000-3p possessed significant diagnostic value in ccRCC.
Previous studies have showed that miRNAs exert their roles by negatively modulating target genes’ expression and function [34–36]. Therefore, the downstream target genes of miR-5000-3p were predicted using a series of target gene prediction programs. PPI network analysis showed close connection among these target genes, and MYC, RHOA, NCL, FMR1 and AGO1 were identified as the top 5 hub genes. Intriguingly, after performing expression, survival and correlation analysis, FMR1 was considered as the most potential downstream target gene of hsa_circ_0037858/miR-5000-3p axis in ccRCC metastasis. To date, the function of FMR1 in ccRCC has not been uncovered and needs to be further studied.
Our preliminary experimental validation showed that overexpression of miR-5000-3p could reverse hsa_circ_0037858-mediated FMR1 upregulation in ccRCC cells. Moreover, after increased expression of miR-5000-3p, hsa_circ_0037858-induced suppression of in vitro metastasis of ccRCC cells was also markedly weakened. All these findings elucidated a potential role of hsa_circ_0037858/miR-5000-3p/FMR1 axis in inhibition of ccRCC metastasis. However, there are still some limitations in the current work: (1) in vivo assays (including animal) were not conducted to confirm these findings; (2) clinical samples of metastatic ccRCC were not used to validate circRNA expression. Thus, our team and other group should further conduct much more basic experiments and large clinical trials to confirm the current findings in the future.
Author Contributions
NYY: designed this work, performed analysis and wrote this manuscript. GWL: performed some analysis. The authors have read and approved the final version of the manuscript.
Acknowledgement
We wish to thank all the help from Program of Innovative Cancer Therapeutics, College of Medicine, Zhejiang University.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
No funding was provided for this study.
References
- 1. Cao H, Sun Z, Wu J, Hao C, Wang W. Metastatic Clear Cell Renal Cell Carcinoma to Pancreas and Distant Organs 24 Years After Radical Nephrectomy: A Case Report and Literature Review. Front Surg. 2022; 9:894272. https://doi.org/10.3389/fsurg.2022.894272 [PubMed]
- 2. Linehan WM, Ricketts CJ. The Cancer Genome Atlas of renal cell carcinoma: findings and clinical implications. Nat Rev Urol. 2019; 16:539–52. https://doi.org/10.1038/s41585-019-0211-5 [PubMed]
- 3. Jonasch E, Gao J, Rathmell WK. Renal cell carcinoma. BMJ. 2014; 349:g4797. https://doi.org/10.1136/bmj.g4797 [PubMed]
- 4. Zhang F, Hou T, Chen L, Xiong M, Zhou M, Kazobinka G, Zhao J, Han X. Comprehensive analysis of lower mitochondrial complex I expression is associated with cell metastasis of clear cell renal cell carcinoma. Transl Cancer Res. 2022; 11:1488–502. https://doi.org/10.21037/tcr-22-242 [PubMed]
- 5. Urdánoz-Casado A, de Gordoa JS, Robles M, Roldan M, Zelaya MV, Blanco-Luquin I, Mendioroz M. Profile of TREM2-Derived circRNA and mRNA Variants in the Entorhinal Cortex of Alzheimer's Disease Patients. Int J Mol Sci. 2022; 23:7682. https://doi.org/10.3390/ijms23147682 [PubMed]
- 6. Sun Y, Zhang Y, Ye Z, Wang Y, Lao Y, Zhang J, Fang M, He J, Yin H, Yan W, Jin W. circRNA-miRNA Complex Participates in the Apoptosis of Myocardial Cells in Myocardial Ischemia/Reperfusion Injury. Discov Med. 2022. [PubMed]
- 7. Lu Y, Li Z, Lin C, Zhang J, Shen Z. Translation role of circRNAs in cancers. J Clin Lab Anal. 2021; 35:e23866. https://doi.org/10.1002/jcla.23866 [PubMed]
- 8. Zheng H, Huang S, Wei G, Sun Y, Li C, Si X, Chen Y, Tang Z, Li X, Chen Y, Liao W, Liao Y, Bin J. CircRNA Samd4 induces cardiac repair after myocardial infarction by blocking mitochondria-derived ROS output. Mol Ther. 2022; 30:3477–98. https://doi.org/10.1016/j.ymthe.2022.06.016 [PubMed]
- 9. Liu X, Song J, Kang Y, Wang Y, Chen A. CircPDSS1 promotes the proliferation, invasion, migration, and EMT of breast cancer cell via regulating miR-320c/CKAP5 axis. Cancer Cell Int. 2022; 22:238. https://doi.org/10.1186/s12935-022-02657-0 [PubMed]
- 10. Shao G, Fan X, Zhang P, Liu X, Huang L, Ji S. Circ_0004676 exacerbates triple-negative breast cancer progression through regulation of the miR-377-3p/E2F6/PNO1 axis. Cell Biol Toxicol. 2022. [Epub ahead of print]. https://doi.org/10.1007/s10565-022-09704-6 [PubMed]
- 11. Lou W, Ding B, Wang J, Xu Y. The Involvement of the hsa_circ_0088494-miR-876-3p-CTNNB1/CCND1 Axis in Carcinogenesis and Progression of Papillary Thyroid Carcinoma. Front Cell Dev Biol. 2020; 8:605940. https://doi.org/10.3389/fcell.2020.605940 [PubMed]
- 12. Fu L, Huo J, Fitrat H, Kong Y, Zhang L, Shang C, Li G, Ji F, Fu X, Qiu X. CircNRIP1 Exerts Oncogenic Functions in Papillary Thyroid Carcinoma by Sponging miR-653-5p and Regulating PBX3 Expression. J Oncol. 2022; 2022:2081501. https://doi.org/10.1155/2022/2081501 [PubMed]
- 13. Ding B, Fan W, Lou W. hsa_circ_0001955 Enhances In Vitro Proliferation, Migration, and Invasion of HCC Cells through miR-145-5p/NRAS Axis. Mol Ther Nucleic Acids. 2020; 22:445–55. https://doi.org/10.1016/j.omtn.2020.09.007 [PubMed]
- 14. Liu L, Gu M, Ma J, Wang Y, Li M, Wang H, Yin X, Li X. CircGPR137B/miR-4739/FTO feedback loop suppresses tumorigenesis and metastasis of hepatocellular carcinoma. Mol Cancer. 2022; 21:149. https://doi.org/10.1186/s12943-022-01619-4 [PubMed]
- 15. Ding B, Yao M, Fan W, Lou W. Whole-transcriptome analysis reveals a potential hsa_circ_0001955/hsa_circ_0000977-mediated miRNA-mRNA regulatory sub-network in colorectal cancer. Aging (Albany NY). 2020; 12:5259–79. https://doi.org/10.18632/aging.102945 [PubMed]
- 16. Wang S, Cao J, Pei L. Knockdown of circ_0004585 enhances the chemosensitivity of colorectal cancer cells to 5-fluorouracil via the miR-874-3p/CCND1 axis. Histol Histopathol. 2023; 38:99–112. https://doi.org/10.14670/HH-18-502 [PubMed]
- 17. Pan X, Huang B, Ma Q, Ren J, Liu Y, Wang C, Zhang D, Fu J, Ran L, Yu T, Li H, Wang X, Yang F, et al. Circular RNA circ-TNPO3 inhibits clear cell renal cell carcinoma metastasis by binding to IGF2BP2 and destabilizing SERPINH1 mRNA. Clin Transl Med. 2022; 12:e994. https://doi.org/10.1002/ctm2.994 [PubMed]
- 18. Gao P, Huang Y, Hou Y, Li Q, Wang H. Circular RNA ITCH Is a Tumor Suppressor in Clear Cell Renal Cell Carcinoma Metastasis through miR-106b-5p/PDCD4 Axis. J Immunol Res. 2021; 2021:5524344. https://doi.org/10.1155/2021/5524344 [PubMed]
- 19. Xue D, Wang H, Chen Y, Shen D, Lu J, Wang M, Zebibula A, Xu L, Wu H, Li G, Xia L. Circ-AKT3 inhibits clear cell renal cell carcinoma metastasis via altering miR-296-3p/E-cadherin signals. Mol Cancer. 2019; 18:151. https://doi.org/10.1186/s12943-019-1072-5 [PubMed]
- 20. Glažar P, Papavasileiou P, Rajewsky N. circBase: a database for circular RNAs. RNA. 2014; 20:1666–70. https://doi.org/10.1261/rna.043687.113 [PubMed]
- 21. Xia S, Feng J, Chen K, Ma Y, Gong J, Cai F, Jin Y, Gao Y, Xia L, Chang H, Wei L, Han L, He C. CSCD: a database for cancer-specific circular RNAs. Nucleic Acids Res. 2018; 46:D925–9. https://doi.org/10.1093/nar/gkx863 [PubMed]
- 22. Li JH, Liu S, Zhou H, Qu LH, Yang JH. starBase v2.0: decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014; 42:D92–7. https://doi.org/10.1093/nar/gkt1248 [PubMed]
- 23. Li R, Qu H, Wang S, Chater JM, Wang X, Cui Y, Yu L, Zhou R, Jia Q, Traband R, Wang M, Xie W, Yuan D, et al. CancerMIRNome: an interactive analysis and visualization database for miRNome profiles of human cancer. Nucleic Acids Res. 2022; 50:D1139–46. https://doi.org/10.1093/nar/gkab784 [PubMed]
- 24. Szklarczyk D, Gable AL, Nastou KC, Lyon D, Kirsch R, Pyysalo S, Doncheva NT, Legeay M, Fang T, Bork P, Jensen LJ, von Mering C. The STRING database in 2021: customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021; 49:D605–12. https://doi.org/10.1093/nar/gkaa1074 [PubMed]
- 25. Lánczky A, Győrffy B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J Med Internet Res. 2021; 23:e27633. https://doi.org/10.2196/27633 [PubMed]
- 26. Wang S, Jin J, Chen J, Lou W. MUC14-Related ncRNA-mRNA Network in Breast Cancer. Genes (Basel). 2021; 12:1677. https://doi.org/10.3390/genes12111677 [PubMed]
- 27. Zhang YH, Song J, Shen L, Shao J. Systematic identification of lncRNAs and circRNAs-associated ceRNA networks in human lumbar disc degeneration. Biotech Histochem. 2019; 94:606–16. https://doi.org/10.1080/10520295.2019.1622782 [PubMed]
- 28. Fan J, Xia X, Fan Z. Hsa_circ_0129047 regulates the miR-375/ACVRL1 axis to attenuate the progression of lung adenocarcinoma. J Clin Lab Anal. 2022; 36:e24591. https://doi.org/10.1002/jcla.24591 [PubMed]
- 29. Luo JT, Wang YF, Wang Y, Wang CL, Liu RY, Zhang Z. A Circular RNA, hsa_circ_0018180 (circPARD3), Triggers Glycolysis and Promotes Malignancy of Head and Neck Squamous Cell Carcinoma Through the miR-5194/ENO1 Axis. Biochem Genet. 2023; 61:316–35. https://doi.org/10.1007/s10528-022-10253-0 [PubMed]
- 30. Lei TX, He DJ, Cao J, Lv WG. CircWDR26 regulates endometrial carcinoma progression via miR-212-3p-mediated typing genes MSH2. Eur J Med Res. 2022; 27:135. https://doi.org/10.1186/s40001-022-00755-3 [PubMed]
- 31. Salmena L, Poliseno L, Tay Y, Kats L, Pandolfi PP. A ceRNA hypothesis: the Rosetta Stone of a hidden RNA language? Cell. 2011; 146:353–8. https://doi.org/10.1016/j.cell.2011.07.014 [PubMed]
- 32. Zhuang YY, Zhong W, Xia ZS, Lin SZ, Chan MC, Jiang K, Li WF, Xu XY. miR-5000-3p confers oxaliplatin resistance by targeting ubiquitin-specific peptidase 49 in colorectal cancer. Cell Death Discov. 2021; 7:129. https://doi.org/10.1038/s41420-021-00494-0 [PubMed]
- 33. Chen H, Ali M, Ruben A, Stelmakh D, Pak M. E2F6-Mediated Downregulation of MIR22HG Facilitates the Progression of Laryngocarcinoma by Targeting the miR-5000-3p/FBXW7 Axis. Mol Cell Biol. 2020; 40:e00496–19. https://doi.org/10.1128/MCB.00496-19 [PubMed]
- 34. Lou W, Chen J, Ding B, Fan W. XIAP, commonly targeted by tumor suppressive miR-3607-5p and miR-3607-3p, promotes proliferation and inhibits apoptosis in hepatocellular carcinoma. Genomics. 2021; 113:933–45. https://doi.org/10.1016/j.ygeno.2021.02.003 [PubMed]
- 35. Zheng H, Ding B, Xue K, Yu J, Lou W. Construction of a lncRNA/pseudogene-hsa-miR-30d-5p-GJA1 regulatory network related to metastasis of pancreatic cancer. Genomics. 2021; 113:1742–53. https://doi.org/10.1016/j.ygeno.2021.04.013 [PubMed]
- 36. Gao X, Wu H, Li Y, Zhang L, Song M, Fu X, Chen R, Ding S, Zeng J, Li J, Liu P. Dancing in local space: rolling hoop orbital amplification combined with local cascade nanozyme catalytic system to achieve ultra-sensitive detection of exosomal miRNA. J Nanobiotechnology. 2022; 20:357. https://doi.org/10.1186/s12951-022-01568-6 [PubMed]