




Introduction
Obesity is a long-term metabolic disease triggered in part by the mutual interaction of genetic, environmental as well as other factors. According to WHO criteria, obesity is commonly described in terms of body mass index (BMI) ≥ 30 kg/m2 [1, 2]. Obesity is associated with an elevated risk of depression, metabolic, anxiety, cardiovascular, and chronic inflammation, as well as some malignant diseases [3]. Studies have shown that obesity is most likely to amplify the prevalence of depression and anxiety disorders [4, 5]. Obesity stems from an expansion of adipose tissue and an imbalance between caloric intake and energy expenditure [6]. Obese patients with higher amounts of visceral fat are at higher risk for serious complications [7]. White adipocytes, which store energy, and thermogenic brown and beige adipocytes, which produce energy, secrete hormones, such as adipokines, lipokines and exosomal microRNAs [8]. The secretion of adipokines and lipokines, such as leptin, adiponectin, monocyte chemoattractant protein-1 (MCP-1), plasminogen activator inhibitor type 1 (PAI-1), retinol Binding Protein 4 (RBP4), visfatin, resistin, apelin, chemerin, palmitoleic acid and lysophosphatidic acid is modified in the presence of adipose tissue dysfunction and may cause a range of obesity-related disorders [9–11]. With a greater comprehension of the functional and molecular targets of adipokines and lipokines, it will hold enormous promise for both new drug treatment strategies and diagnostics.
Depression is a severe psychiatric disorder characterized by persistent low mood, diminished interest, slowed thinking, reduced volitional activity, sleep difficulties or disturbances in appetite, affecting more than 300 million people worldwide and the number is growing [12, 13]. There is a growing body of research recognizing that negative emotions such as anxiety, stress and depression exert a significant negative influence on health and illness. There are also relevant clinical studies that show that obesity suffers from varying degrees of depressive symptoms [14, 15]. Recent scientific evidence suggests that depression and obesity are not independently linked. The two disorders can be interrelated through a vicious, mutually reinforcing cycle of maladaptive physiological adaptations [5]. Lassale et al. demonstrated a bidirectional and complex multifactorial relation between mood disorders and obesity [16]. Data from their meta-analysis revealed that men and women suffering from obesity had a 55% elevated risk of developing depression, while those suffering from depression had a 58% higher risk of developing obesity [16]. There is growing evidence that adipocytokine and lipokine levels are up- or down-regulated in the progression of depression [10, 11, 17]. Furthermore, a large body of evidence suggests that modulating adipocytokine and lipokine levels in models of depression attenuates or promotes depressive-like behaviors [18–20]. Currently, the pathogenesis of depression is not completely clear. There are many hypotheses for depression, such as the monoamine neurotransmitter hypotheses, neurotrophic factor hypotheses (mainly brain-derived neurotrophic factor (BDNF) expression and functional downregulation), neurocircuitry hypotheses, neuroendocrine hypotheses, neuroinflammation hypotheses, gut microbiota hypotheses, neuroplasticity hypotheses, hypothalamic-pituitary-adrenal (HPA) axis hypothesis, etc. [21–26].
Among them, neuroendocrine hypotheses, inflammation, gut microbiota, neuroplasticity and abnormal HPA axis function are currently the most popular directions for investigating the pathogenesis of depression. It has been established nowadays that dysregulation of the innate and adaptive immune system happens in patients with depression and that inflammatory processes are highly correlated with the pathophysiology of depression [27]. Studies have suggested that a sustained, low-grade inflammatory response is a potentially modifiable risk factor for obesity [28, 29]. In addition, there is increasing evidence that the microbial community of the entire gastrointestinal tract (gut microbiota) is related to depressive disorders [30]. Multiple lines of evidence suggest that the gut microbiota is participating in the progression of obesity and related co-morbidities [30–36]. The neuroplasticity hypothesis, suggesting that the antidepressant effect can be regulated by modulating synaptic plasticity in hippocampal neurons and thus affecting structural plasticity in neural networks, is also the hypothesis applied in the study of the rapid antidepressant effect of ketamine [21, 37]. High-fat diet (HFD) feeding is currently a common method for establishing obese animal models [38, 39]. Studies have suggested that HFD leads to persistent elevations in cytokines and chemokines that can cause region-specific neuroplasticity, thereby promoting mood deficits and increased body weight [4]. Neuroendocrine studies have suggested that the HPA axis is relevant to the pathophysiology of depression, and studies have confirmed the overactivity of the HPA axis in patients with major depressive disorder (MDD) [40]. Research shows that the HPA axis may influence the body weight of stressed individuals by regulating cortisol [41]. Taken together, this review offers a well-rounded description of the mechanisms underlying the development of obesity and depression to better understand the interrelationship between the two conditions and to provide more effective treatment approaches.
Shared biological mechanisms of depression and obesity
There is ample reason to assume that depression and obesity are interrelated via a vicious, mutually strengthening cycle of negative physiological adaptations. In this section, we review the mechanisms underlying the co-pathogenesis of obesity and depression, mainly around the inflammation, gut microbiota, gut-brain axis (GBA)/microbiota-gut-brain axis (microbiota-GBA), neuroplasticity and HPA axis abnormalities (Figures 1, 2).
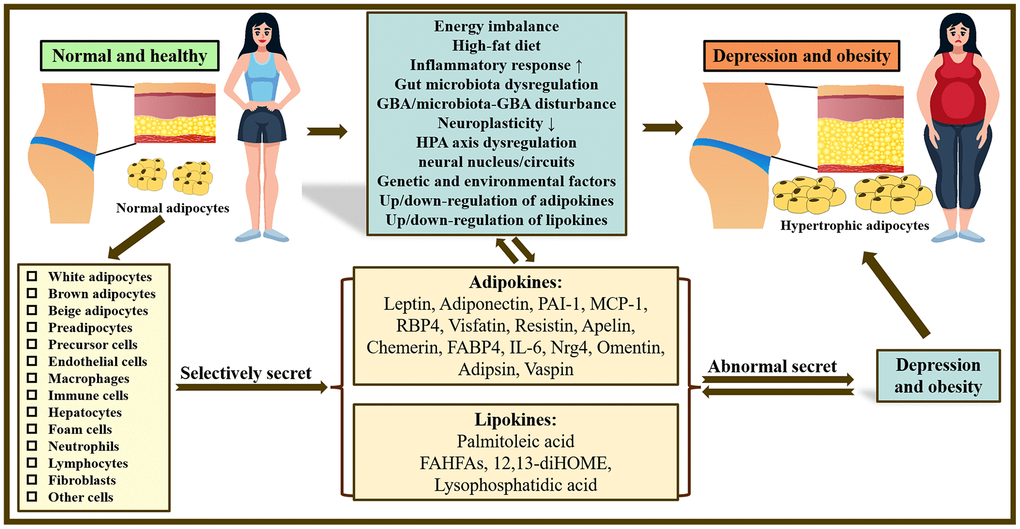
Figure 1. Overview of the mechanisms underlying the co-pathogenesis of depression and obesity: based on adipokines and lipokines. In the body, there are many kinds of adipocytes and different adipocytes can selectively release related adipokines and lipokines. Abnormal secretion of adipokines and lipokines plays a crucial role in obesity and depression and may be related to increased inflammatory response, gut microbiota disturbance, neuroplasticity, dysfunction of HPA as well as GBA/microbiota-GBA disturbance. Note: HPA axis, hypothalamic-pituitary-adrenal axis; GBA, gut-brain-axis; PAI-1, Plasminogen activator inhibitor type 1; MCP-1, Monocyte chemoattractant protein-1; RBP4, Retinol Binding Protein 4; FABP4, fatty acid-binding protein 4; IL-6, Interleukin 6; Nrg4, Neuregulin 4; FAHFAs, fatty acid esters of hydroxy fatty acids; 12,13-diHOME, 12,13-dihydroxy-(9Z)-octadecenoic acid.
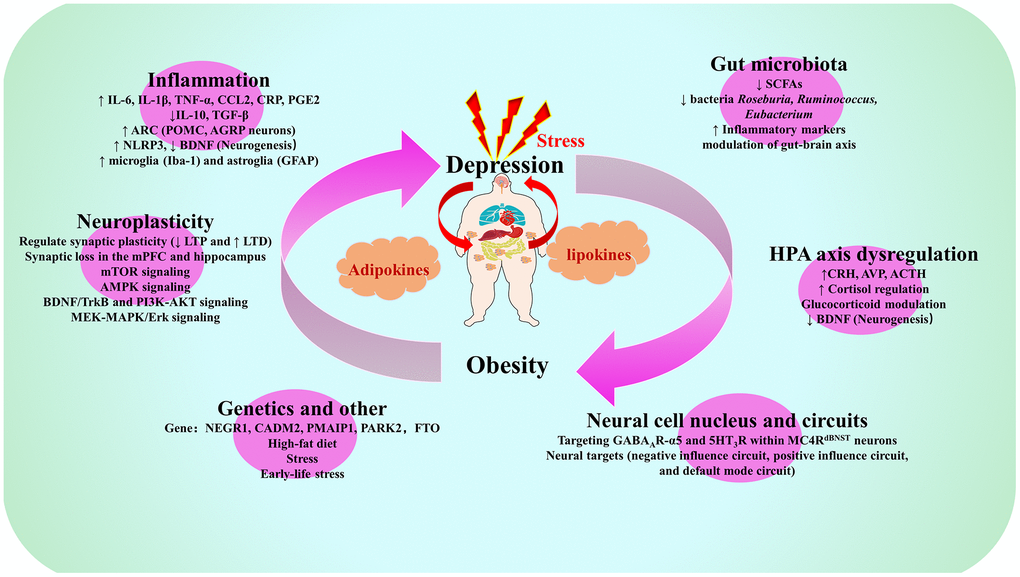
Figure 2. Summary map of common biological mechanisms of depression and obesity. Note: IL-6, Interleukin-6; IL-1β, Interleukin-1β; TNF-α, Tumor necrosis factor-α; CCL2, C-C motif chemokine ligand 2; CRP, C-reactive protein; PGE2, prostaglandin E2; IL-10, Interleukin-10; TGF-β, transforming growth factor-beta; ARC, arcuate nucleus of the hypothalamus; POMC, pro-opiomelanocortin; AGRP, agonist peptide-related peptide; NLRP3, NOD-like receptor family pyrin domain containing 3; BDNF. brain derived neurotrophic factor; Iba-1, ionized calcium binding adapter molecule-1; GFAP, glial fibrillary acidic protein; SCFAs, short-chain fatty acids; LTP long-term potentiation, LTD, long-term depression; mPFC, medial prefrontal cortex; mTOR, mechanistic target of rapamycin; AMPK, AMP-activated protein kinase; TrkB, tyrosine kinase receptor B; PI3K, phosphatidylinositol 3-kinase; AKT, protein kinase B; MEK, MAPK/extracellular signal-regulated kinase; MAPK, mitogen-activated protein kinases; ERK, extracellular signal-regulated kinase; CRH, corticotropin-releasing hormone; AVP, arginine vasopressin; ACTH, adrenocorticotropic hormone; NEGR1, neuronal growth regulator 1; CADM2, Cell adhesion molecule 2; PMAIP1, phorbol-12-myristate-13-acetate-induced protein 1; PARK2, E3 ubiquitin-protein ligase parkin; FTO, Fat mass and obesity-associated gene; GABA, gamma-aminobutyric acid; MC4R, melanocortin 4 receptor; dBNST, bed nucleus of the stria terminus.
The potential role of inflammation in depression and obesity
Overactivation of the immune system is caused by several factors, such as difficult life situations, stress from society, and poor lifestyle habits (smoking, lack of exercise, HFD), all of which cause an increased inflammatory response and thus promote depressive symptoms [42]. Elevated levels of inflammatory markers such as IL-1β, IL-6, IL-2, TNF-α, CRP and PGE2 were found in patients with depression [43–47]. The cytokine hypothesis suggests that in depression, the number of pro-inflammatory cytokines such as IL-6, IL-1β and TNF-α is increased, while the number of anti-inflammatory cytokines such as IL-10 and TGF-β is decreased, tilting the overall immune response toward inflammation [48, 49].
The increase in macroglia peripheral nervous system and central nervous system (CNS) cytokines is coupled with the activation of microglia, the brain’s immunocompetent cells. This, in turn, has been linked to changes in synaptic plasticity, neurogenesis and emotional behavior [50]. A variety of key inflammatory markers also have been implicated in the risk of adverse outcomes of obesity and obesity-related diseases [48]. Adipocytes in abdominal, muscle and liver fat produce pro-inflammatory and chemotactic compounds such as IL-6, IL-1β, TNF-α and C-C motif chemokine ligand 2 (CCL2), as well as inflammatory hormones such as adiponectin and leptin [51, 52]. Overweight and obese individuals have adipocytes and macrophages in their adipose tissue that cause the secretion of cytokines and chemokines which can cross the blood-brain barrier (BBB) as well as stimulate neuroinflammation [53, 54].
Neuroinflammation triggered by obesity has recently been demonstrated to influence multiple brain structures like the hippocampus, cortex, brainstem or amygdala, in addition to being linked to an elevated incidence of central disorders including depression as well as impaired cognitive function [55]. Chronic overnutrition and obesity can lead to chronic low-level inflammation across the body, a state of chronic inflammation known as “meta-inflammation” mediated by macrophages located in the colon, liver, muscle as well as adipose tissue [56]. HFD is associated with the production of cytokines by non-neuronal cells in the hypothalamus that activate diverse inflammatory mediators in the arcuate nucleus of the hypothalamus (ARC), a primary region of the hypothalamus that regulates pro-opiomelanocortin (POMC) and agonist peptide-related peptide (AGRP) neurons, the primary neurons that sense and integrate peripheral metabolic signals and respond accordingly [57, 58]. Therefore, it is important to discuss the role of neuronal and non-neuronal cells in HFD-induced hypothalamic inflammation. The vagus nerve is linked to the visceral organs and the CNS, and the vagal afferent nerve conveys hunger signals to the hypothalamus via the gastric-derived orexigenic peptide ghrelin [59]. Studies have shown that intake of HFD triggers inflammatory responses in the gut, nodal ganglia and hypothalamus within a short period vagal afferent nerves can transmit inflammatory signals of gut origin to the hypothalamus through the nodose ganglion, while ghrelin can prevent inflammation induced by HFD [58, 59]. Ingestion of HFD leads to a lack of integrity of the gut barrier, causing the transfer of macromolecules, such as microbial or pathogen-associated molecular patterns (e.g., lipopolysaccharides) entering the systemic circulation [60]. Nucleotide-binding oligomerization domain–like receptor family pyrin domain-containing 3. (NLRP3) inflammatory vesicles are an essential component of the innate immune system, mediating caspase-1 activation and secretion of the pro-inflammatory cytokine IL-1β/IL-18 in response to microbial infection as well as cellular injury [61, 62]. Li et al. showed that microglia NLRP3 inflammatory vesicles induce neurotoxic astrocyte function by activating the neuroinflammatory caspase-1 pathway as a response to chronic stress [63]. Vandanmagsar et al. showed that NLRP3 inflammatory vesicles sense danger signals associated with obesity and lead to obesity-induced inflammation and insulin resistance [51]. The study by Schachter et al. suggests that reducing obesity-related neuroinflammation may have beneficial effects on depression [30]. Li et al. revealed that HFD-induced diabetes and obesity-related neuroinflammation activated the transcription factor CCAAT/enhancer binding protein beta (C/EBPβ) in hippocampal neurons and that this factor suppressed BDNF expression and caused depression-like behavior in male mice [64]. This suggests that inflammation-activated neuronal C/EBPβ can contribute to HFD-induced depression by decreasing BDNF expression [53].
BDNF is mainly produced by microglia and astrocytes and has neuroprotective as well as anti-inflammatory effects [65, 66]. Obesity creates an environment of chronic inflammation that leads to negative physiological and neurological outcomes such as diabetes, cardiovascular disease, and depression. Although the entire body is involved in metabolic homeostasis, the neuroimmune system has recently emerged as a key regulator of metabolism [67]. Lam et al. found that 12 weeks of HFD feeding not only led to obesity in mice but also to depression-like behaviors and that astrocyte activation, which is closely associated with depression, was also evident in the ventral hippocampus [68]. Four weeks of pioglitazone treatment attenuated HFD-induced glucose metabolic dysfunction, upregulated ventral hippocampal GFAP (glial fibrillary acidic protein), reduced the total process length and number of branching sites of hippocampus CA1 GFAP-immunoreactive astrocytes in the ventral hippocampus, and attenuated the depressive phenotype, suggesting that pioglitazone may be a promising therapeutic agent for metabolic disorders and related depression [68]. Microglia are resident immune cells in the brain that respond directly and indirectly to dietary fat, and the environment in which microglia develop helps them to respond later in life [67]. Numerous studies have shown that chronic peripheral inflammatory states reduce adult hippocampal neurogenesis [69–71]. Peripherally released pro-inflammatory cytokines are implicated in communicating between the peripheral immune system and the brain via the activation of microglia in the brain [71]. Activated microglia may reduce neurogenesis by inhibiting the proliferation of neural stem cells, increasing apoptosis of neural progenitors, and decreasing the survival of newly developing neurons as well as their integration into existing neural circuits [69]. Stress-induced microglia activation and neuroinflammation play a vital role in the pathogenesis of depression [72, 73]. However, obese patients with chronic low-grade inflammation are at increased risk of developing depression [74]. Peroxisome proliferator-activated receptor gamma (PPARγ) is a nuclear transcription factor that moderates microglia polarization and neuroinflammation [72]. Qin et al. showed that chronic unpredictable mild stress (CUMS) was able to induce severe depression-like behavior, neuroinflammation and reduced PPARγ expression in leptin-deficient (ob/ob) mice compared to wild-type/C57BL/6J mice, suggesting that PPARγ mediates an activated phenotype of microglia, which may be related to the susceptibility of stressed ob/ob mice to develop depression [72].
The potential role of gut microbiota in depression and obesity
The degree of diversity and balance of gut microbiota strains is an essential indicator of the overall health status of the organism [75]. Depression and obesity have a high comorbidity in individuals, which may be due to sharing similar risk factors, including disruption of the gut microbiota. The gut microbiota is composed of over 100 trillion microorganisms, including a minimum of 1000 various species of bacteria, and plays a crucial role in the physiological and pathophysiological processes that occur in the body [76, 77]. The gut microbiota evolved together with the host and is an indivisible part of the body. Gut microbes are diverse and affect many processes, including immunity, metabolism as well as the CNS. Microbiota-GBA has emerged as a new candidate for depression [78, 79]. It has been shown that patients with depression, which includes those in remission, have a various gut microbiota composition than healthy controls [80]. Animal models of depression have shown changed gut microbiota compared to controls [81]. In a further meta-analysis, disturbances in the gut were found to be correlated with a decrease in certain anti-inflammatory butyrate-producing bacteria as well as an increased number of pro-inflammatory bacteria in patients with depression and other psychiatric disorders [82]. The fat mass and obesity associated gene (FTO) [83], an RNA demethylase [84, 85], is downregulated in the hippocampus of patients with MDD and in mouse models of depression [86]. Suppression of Fto expression in the mouse hippocampus results in depression-like behaviour in adult mice, whereas overexpression of FTO expression rescues the depression-like phenotype, suggesting that FTO is a modulator of depression-like behavioural mechanisms in mice [86]. Sun et al. showed that Fto deletion resulted in decreased body weight, reduced anxiety and depression-like behavior, and reduced sensitivity to stressful stimuli in Fto+/- mice [36]. In terms of intestinal flora, Fto-deficient mice were characterized by specific anti-inflammatory bacteria, importantly, the behavioral changes in Fto+/- mice were mediated by changes in the gut microbiota [36]. The results on the role of FTO in depression-obesity comorbidity are inconclusive, and further research is needed to deepen knowledge of the genetic basis of this comorbidity [87].
It has been shown that dysregulation of the gut microbiota in mice is associated with several neurobiological traits of depression, such as mild chronic inflammation, abnormal activity of the HPA axis, and reduced adult neurogenesis [88, 89] Recent studies suggest that chronic inflammation resulting from a HFD may exert a central role in the induction of neuroinflammation and depression [30]. Remarkably, the gut microbiota mediates some of the effects of HFD on human physiology and influences host mood and behavior. Pathogens, stressors, and predisposing factors can all contribute to excessive or protracted inflammatory responses (moderators include childhood trauma and obesity). The ensuing illness behaviors (such as pain, and sleep disturbance), depressive symptoms, and unhealthy lifestyle choices (such as a poor diet and sedentary lifestyle) may function as mediation pathways that result in unregulated inflammation and depression. Stress, nutrition, depression, and adverse childhood experiences can all affect the gut microbiome and increase intestinal permeability, which is another route to elevated inflammatory responses [42]. Studies have shown that fruits, vegetables and edible legumes contain high levels of phytochemicals that have anti-inflammatory effects [90, 91]. Camu Camu (CC) has significant antioxidant and anti-inflammatory potential [92]. Treatment of high-fat/high-sucrose (HFHS)-fed mice that are treated with CC prevent weight gain, reduce fat accumulation, attenuate metabolic inflammation and endotoxemia, and mice treated with CC exhibit enhanced glucose tolerance as well as insulin sensitivity [93]. Consuming fruits and vegetables rich in polyphenols may improve cognitive and emotional health by reducing oxidative stress and inflammation [94]. To investigate the effect of the gut microbiota on stress-induced depressive behavior, Chevalier et al. adopted an unpredictable chronic mild stress (UCMS) mouse model of depression as well as fecal microbiota transfer (FMT) from stress donors to naïve mice [95]. Results revealed that microbiota transfer conveyed behavioral symptoms of depression and reduced adult neurogenesis in recipient mice [95]. In addition, the metabolomic analysis demonstrated that FMT mice underwent changed fatty acid metabolism characterized by a deficiency of lipid precursors of endocannabinoid (eCB), which led to the compromised activity of the eCB system of the brain [95]. The eCB system plays its pleiotropic role via multiple neuronal processes. eCB system regulates adult neurogenesis through CB1 receptors expressed by neural progenitor cells [96]. CB1-deficient mice exhibit damage neural progenitor cell proliferation, self-renewal as well as neurosphere production, while CB1 receptor agonists enhance neurogenesis [97, 98].
A randomised controlled trial study by Ren et al. showed that an almond-based low carbohydrate diet (LCD) improved depression and glucose metabolism in patients with type 2 diabetes by modulating gut microbiota and glucagon-like peptide-1 (GLP-1) [99]. Almond-based LCD significantly reversed depressive-like behaviour and glycosylated haemoglobin, while significantly increasing the number of short-chain fatty acid (SCFA)-producing bacteria Roseburia, Ruminococcus and Eubacterium. This suggests that the role of almond-based LCD in improving depression in type 2 diabetic patients may be related to its stimulation of the growth of SCFA-producing bacteria, increased SCFA production and activation of the free fatty acid 2 (FFA2) receptor (known as GPR43), and further maintenance of GLP-1 secretion.
Brain-gut axis plays important role in depression and obesity. The gut microbiota has been proven to interact with various organs, including the brain. Gut microbiota and its metabolites may act directly or indirectly on the brain via vagal stimulation to regulate metabolism, obesity, body homeostasis and energy balance as well as central appetite and food reward signals, which play a vital role in obesity [100]. There is a growing body of data suggesting that the gut microbiota coordinating multiple bodily functions is closely linked to the immune, metabolic and nervous systems and that dysbiosis of the gut dysbiosis disrupts the homeostasis between these systems [101]. The GBA is a bidirectional connection between the gut microbiota and the brain [100]. GBA influences physiological function and behavior through three different pathways (neural pathways, endocrine pathways, and immune pathways) [100, 102]. The neural pathway mainly consists of the enteric nervous system and the vagus nerve; the endocrine pathway mainly affects the neuroendocrine system of the brain, especially the HPA axis as well as the immune pathway [100, 102]. In particular, the signals from a brain influence the motor, sensory and secretory patterns of the gastro-intestinal tract, regulate inflammatory processes and influence the structure of the gut microbiota, and in turn, visceral information from gastro-intestinal features can affect brain function [103]. For example, neuroendocrine hormones (e.g., corticosterone) alter intestinal permeability, barrier function, and communicate with immune cells regarding cytokine secretion, and immune cells release cytokines important in the host responses to inflammation and infection [104]. Some alterations in the gut microbiota can modulate host metabolic pathways and dietary behavior through GBA, leading to obesity [105]. Recently, the role of the microbiota as an important factor in regulating gut-brain signaling has emerged and the concept of the microbiota-GBA has been established [105]. The GBA is proposed as the focus of new scientific and clinical research on its possible sites of systemic therapeutic interventions for depression and obesity [106].
The potential role of neuroplasticity in depression and obesity
Neuroplasticity is characterized as the potential of the brain to undergo neurobiological changes in responding to external stimuli [107]. It is currently accepted that the mammalian brain shows continuous plasticity at all stages of life. The plasticity of neurons enables the CNS to learn newly acquired skills and to engage in ongoing learning and memory. In addition, neuronal plasticity allows the reorganization of neuronal networks in response to environmental stimuli as well as recovery after lesions [108]. Neuronal plasticity can be achieved through neurogenesis, apoptosis, synapse-dependent activity, as well as reorganization of neuronal networks [109]. Stress and depression are usually correlated, and studies have shown that chronic stressful events commonly accelerate depressive episodes in vulnerable individuals. Advanced brain functions are proposed to require synaptic frequency decoding, and changes in synaptic activation frequency may cause an increase or decrease in the long-term efficiency of these synapses, which may lead to long-term potentiation (LTP) or long-term depression (LTD) [110]. Hwang et al. showed that the intensity of synaptic plasticity (LTP and LTD) at Schaffer lateral branch-CA1 synapses in hippocampal isolated slices of obese male mice was lower compared to sex-specific controls [111]. This suggests that, like mice in the depression model, HFD-obese mice also affect hippocampal synaptic plasticity. In neuroscience research, depressive-like behaviors and chronic stress have been related to damage to neuroplasticity, like neuronal atrophy as well as synaptic loss in the medial prefrontal cortex (mPFC) and hippocampus [112]. Neuroplasticity as a downstream mechanism of the effect of novel fast-acting antidepressants such as ketamine has stimulated great interest in the mechanisms of neuroplasticity [113]. Among them, α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) activation, BDNF and mammalian/mechanistic target of rapamycin (mTOR)-mediated signaling, synaptic protein expression and synaptogenesis are framed with a focus on neuroplasticity to explain the potent and sustained depression treatment effects of these compounds [113].
The hypothesis of neuroplasticity in depression suggests that downstream effects of antidepressants, such as increased neurogenesis, contribute to improved cognition and mood [21]. BDNF is a growth factor that modulates neurite growth, functional neuronal connections, synapse formation, as well as synaptic plasticity in the CNS [114]. BNDF signaling was proved to be required for the antidepressant effect of ketamine [115]. Activation of the high-affinity BDNF receptor, tropomyosin receptor kinase B (TrkB), is essential for antidepressant-related behaviors [114]. Numerous reports using depression-like behaviors or different animal models manipulating the expression of BDNF or its receptor TrkB suggest that BDNF/TrkB participates in the pathophysiology of depression as well as the mechanism of action of antidepressant treatments [114, 116]. Enhanced translation and/or release of BDNF and activation of the BDNF receptor target TrkB may lead to further activation of downstream pathways essential in synaptic plasticity. BDNF-mediated activation of the TrkB receptor activates the PI3K/Akt signaling pathway and downstream activation of the MEK-MAPK/Erk signaling pathway [115]. Both pathways promote protein translation by activating the mechanistic target of rapamycin complex 1 (mTORC1) [117]. The acute activation of mTOR and protein translation may trigger sustained changes in synaptic plasticity leading to long-term effects of ketamine. Preclinical studies have shown that sodium-glucose co-transporter 2 inhibitors (SGLT2i) improve cognitive dysfunction, reduce oxidative stress, and neuroinflammation, and improve neuronal plasticity and mitochondrial brain pathways in obese and type 2 diabetes mellitus (T2DM) mice [118]. The mTOR serves as a trophic sensor with critical homeostatic functions in modulating energy metabolism, supporting neuronal growth as well as plasticity [115]. In addition, SGLT2i restored mTOR to its activated physiological state and prevented the onset or progression of neurodegenerative diseases [118].
Neuroplasticity due to chronic sugar intake has been shown to reduce impulse control and thus resistance to high-fat/high-sugar foods, leading to an obesity epidemic [119]. Akhaphong et al. showed that placental mTORC1 is the mechanistic connection between placental function and the programming of obesity as well as insulin resistance in adult offspring [115]. A study by Li et al. found that 8 weeks of HFD-feeding was effective in inducing metabolic disorders, including obesity as well as hyperlipidemia, in mice. Interestingly, the mice also exhibited depressive and anxious behaviors [120]. Li et al. concluded that HFD-feeding inhibited AMPK phosphorylation and induced mTOR phosphorylation [120]. After 28 days of treatment with the mTOR inhibitor rapamycin, autophagy and BDNF levels were elevated [120]. This suggests that improvement of lipid metabolism or enhancement of autophagy via the AMPK/mTOR pathway may be a potential candidate target for the therapy of obesity and depression.
The potential role of HPA axis dysregulation in depression and obesity
The HPA axis, which consists of the hypothalamus, pituitary and adrenal glands, modulates the production of glucocorticoids and is associated with the pathophysiology of psychiatric disorders [40]. During stress conditions, the HPA axis promotes transient physiological adaptations that usually resolve after the stress stimulus is not present. Both psychological and physiological stresses activate the HPA axis, which stimulates the release of corticotrophin-releasing factor (CRF) and arginine vasopressin (AVP) from the hypothalamic paraventricular nucleus (PVN), and the HPA axis is an essential component of the stress response system [40, 121]. AVP activates the locus ceruleus-norepinephrine neuromodulatory system, triggering a “fight or flight” response (regulated by the epinephrine and the norepinephrine), while CRH acting on the pituitary gland, which in response secretes adrenocorticotropic hormone (ACTH) into the bloodstream [40]. Once ACTH arrives in the adrenal glands, it initiates the release of cortisol or corticosterone, an anti-inflammatory hormone, and mediates the physiological behavioral response to stress [40]. In normal conditions, negative feedback from cortisol on CRH and ACTH ensures HPA homeostasis via the activation of glucocorticoid receptors (GRs) as well as mineralocorticoid receptors (MRs) [40]. CRF and AVP both induce the secretion of ACTH from the anterior pituitary gland, which increases the release of ACTH, causing an elevated level of circulating glucocorticoid (GC) and inhibiting the secretion of CRF and AVP from the hypothalamus, forming a reverse feedback circuit [122]. Chronic stress is known to overstimulate the HPA axis and the regulation of corticosterone secretion, which is associated with abdominal obesity [123, 124]. Studies have shown that increased physical activity can normalize corticosterone secretion and thus have a positive impact on physical and mental health. A randomized controlled trial by Lasselin et al. found that immune and behavioral responses to lipopolysaccharide (LPS) differed little between obese and normal-weight subjects, but that cortisol responses to LPS were significantly attenuated in obese individuals and that higher body fat percentage was associated with lower cortisol responses to LPS [125]. This suggests that young and healthy obese individuals do not have increased behavioral sensitivity to cytokines, but have diminished cortisol responses to immune challenge.
Over the last decades, several hypothalamic axis abnormalities associated with stress overreaction have been identified in patients with depression. These include alterations such as excessive CRF secretion in the paraventricular nucleus of the hypothalamus, impairment of negative feedback in the HPA axis, enlarged adrenal glands, hypercortisolism, and reduced inhibition of cortisol by dexamethasone [126]. Neuroendocrine studies have demonstrated HPA axis hyperactivity in patients with MDD. Changes in the HPA axis in depressed patients have been consistently reported, with approximately 30% of depressed patients having higher cortisol levels [127]. It has been shown that over 40-60% of depressed patients experience hypercortisolemia. Increased stress-induced GC secretion may enhance mood and motivation [128]. Microbiota-induced hyperactivity of the HPA axis and inflammation have also been implicated in causing depression [129]. Pro-inflammatory cytokines exert the potential to activate the HPA axis, and patients with depression also have a hyperactive HPA axis. Furthermore, HPA axis dysfunction decreases BDNF expression, inhibits 5-HT synthesis, reduces Glu receptor expression, and even disrupts neuroplasticity as well as neural circuits [122, 130]. Related studies suggest that an excess of pro-inflammatory cytokines inhibits the negative feedback of the HPA axis, growing the permeability of the BBB, reducing 5-HT synthesis, interfering with the glutamatergic system, and ultimately leading to depression [131, 132]. More than half of depressed patients display negative feedback dysfunction of the HPA axis, including chronic increases in circulating GC and ACTH [133]. Chronic stress and stress hormones such as glucocorticoids trigger metabolic changes, including obesity and diabetes.
A growing body of research suggests that chronic stress-related stimulation of the HPA axis and the causing excess glucocorticoid exposure may exert a fundamental role in the development of visceral obesity [134]. FGF21 is a longevity factor that coordinates the interaction between energy metabolism and stress response [135, 136]. FGF21 is a general stress factor that not only alters energy metabolism but also activates multiple rescue processes directly or indirectly by stimulating the secretion of lipocalin and hormones in the HPA axis through the AMPK signaling pathway and several other pathways [137]. Recent studies have shown that FGF21 therapy can relieve many metabolic diseases, such as obesity, type 2 diabetes as well as some cardiovascular diseases. FGF21 can control stress response and metabolism by regulating the function of the somatotropic axis and HPA pathway [135]. Werdermann et al. showed that obese animals show an overactive HPA axis, leading to adrenal hyperplasia [137].
The potential role of the neural cell nucleus and neural circuit in depression and obesity
Although neuroplasticity is important for depression and obesity co-morbidity, there is growing evidence that it is significant to explore the neural mechanisms of depression and obesity co-morbidity at the level of the neural cell nucleus as well as the neural circuit [138]. Xia et al. suggest that melanocortin 4 receptor (MC4R) neurons, located in the bed nucleus of the stria terminus (dBNST), are involved in psychiatric-related body weight regulation by receiving α5-containing GABAA receptor and serotonergic GABAergic projections from hypothalamic AgRP neurons [138]. Overall, targeting GABAAR-α5 and 5HT3R within MC4RdBNST neurons contributed to rescuing HFD-induced anxiety and depression, thereby reducing body weight by simultaneously reducing HFD cravings and enhancing a healthy low-fat diet in mice [138]. In addition, studies have shown that mixtures of zonisamide-granisetron cocktail restore mental normalcy and promote weight loss by targeting the GABAAR-α5 and 5HT3R pathways, respectively, by altering food preferences toward a healthy low-fat diet [138].
Recent studies by Hallihan et al. have found that affective neural circuits and inflammatory markers are associated with symptoms of depression and anxiety in comorbidities of obesity [139]. They used the current feasibility study using functional neuroimaging and biospecimen data to identify whether changes in inflammatory markers, fecal short-chain fatty acids, as well as neural circuit-based targets predicted depression and anxiety outcomes in comorbid obese participants, and preliminary correlation analyses showed significant correlated changes in three inflammatory markers (IL-1RA, IL-6, and TNF-α) and five neural targets (negative influence circuit, positive influence circuit, and default mode circuit) at 2 months [139]. Further, they found that changes in IL-1RA and TNF-α at 2 months, as well as changes in three neural targets (negative influence circuit and positive influence circuit), correlated with changes in depression and anxiety symptoms at 6 months [139]. Perinatal exposure to maternal obesity, metabolic disorders (including diabetes and hypertension), and unhealthy maternal diets have long-term effects on the behavior and physiology of offspring [123]. Evidence from epidemiological studies suggests that maternal obesity and metabolic comorbidities increase the risk of attention deficit hyperactivity disorder (ADHD), autism spectrum disorder (ASD), depression, schizophrenia, eating disorders (food addiction, anorexia nervosa, as well as bulimia nervosa), and cognitive impairment in offspring [140]. In addition, during pregnancy, inflammation in the offspring’s brain impairs the development of neural pathways critical to behavioral regulation, such as the serotonergic, dopaminergic, and melanocortinergic systems [123].
The potential role of genetic association in depression and obesity
Mendelian randomisation (MR) studies have shown that obesity and the development of depression are causally related [141]. To provide a better comprehension of the relationship between obesity and depression, Speed et al. conducted a MR study of the relationship between adiposity, non-adiposity, height and depression, using results from the UK Biobank (n = 332,000) and the Psychiatric Genomics Consortium (n = 480,000) genome-wide association studies [141]. The results of the study found that both adiposity and height (short stature) were causal risk factors for depression, whereas non-adiposity was not [141]. In addition, they note that reducing fat mass will reduce the risk of depression. O’Loughlin et al. performed a two-sample MR using genetic summary statistics (15771 cases and 178777 controls) from the recent Genome-Wide Association Study (GWAS) of depression in East Asian ancestry. i.e., single nucleotide polymorphisms (SNPs) associated with BMI and SNPs associated with waist height ratio (WHR) were selected as genetic instrumental variables and inverse variance weighting (IVW) method to estimate the causal relationship between BMI and WHR and depression [142]. This GWAS data provides the first MR evidence that high obesity is associated with a lower risk of depression in a population of East Asian ancestry living in East Asia. Liao et al. performed genome-wide genotyping of 106,604 unrelated individuals from a Taiwanese biobank to derive polygenic risk scores for BMI and MDD to assess their effects on obesity-associated traits [143]. This study showed that MDD polygenic risk score (PRS) was positively associated with waistline, hipline, waist-hip ratio, body fat percentage, BMI, overweight (BMI ≥ 25), and obesity (BMI ≥ 30) [143]. The results of the meta-analysis of interactions suggest a genetic mechanism for the increased risk of obesity in patients with depression [144]. Rivera et al. showed that depression enhanced the effect of FTO variants on BMI, with a 2.2 increase in BMI per rs9939609 risk allele (A) in depressed patients compared to mentally healthy controls [144]. This suggests that alterations in key biological processes associated with depression may interact with FTO risk alleles to increase BMI or obesity risk.
In a cross-trait meta-analysis, Amare et al. identified 14 genetic loci (including NEGR1, CADM2, PMAIP1, and PARK2) linked to obesity and response to treatment with Selective serotonin reuptake inhibitors (SSRIs) [145]. Weight gain is a side effect of antidepressants and antipsychotics that can cause many comorbidities and reduce life expectancy [146]. The majority of included studies demonstrated a 5% weight gain in individuals treated with antidepressants; however, Quetiapine, Haloperidol, Trifluoperazine, Risperidone, Aripiprazole, Olanzapine, and Clozapine resulted in≥7% weight gain from baseline [146]. Some antidepressants, such as Mirtazapine, show significant levels of weight gain, while others, such as Bupropion, show weight loss effects [147]. Therefore, controlling undesired weight effects is an important consideration in the selection of antidepressants.
Major adipocytokines and lipokines as well as current relevance to obesity and depression
Studies have established that adipose tissue is a dynamic organ that performs several vital physiological processes. It consists of many cell types: e.g., adipocytes, pericytes, preadipocytes, vascular endothelial cells, macrophages and fibroblasts [148]. However, the major cells present in the adipose tissue are the mature adipocytes. Three types of adipocytes are found in mammals and are usually classified according to their color appearance: white, brown and beige/brite/brown adipocytes [149, 150]. White adipocytes are primarily involved in energy storage and mobilization, while brown adipocytes are chiefly involved in nonshivering thermogenesis [149]. Adipose tissue and adipocytes store the energy, produce adipokines and lipokines to regulate the energy supply and are crucial players in the endocrine system. Adipokines and lipokines act on the target tissues to regulate energy supply, lipid metabolism and immune response [151]. Adipokines are derived mainly from adipocytes, even though some members are synthesized by other types of cells as well [152]. These adipocytokines resemble classical cytokines and have pleiotropic functions that influence biological processes throughout the entire organism [152]. It has long been suggested that adipocytokines have a profound protective role in the pathogenesis of diabetes and cardiovascular disease [153]. In recent years, it has been found that adipocytokines also act as an effective modulator of depressive and anxiety-like states, but the mechanism of action of adipocytokines in modulating depressive and anxiety-like behavior is still unclear. Recently, more and more adipocytokines have been recognized, such as leptin, adiponectin, plasminogen Activator I (PAI-1)/visfatin, MCP-1, RBP4, pre-B-cell colony enhancing factor, resistin, apelin, chemerin, aprotinin, vaspin, and others [152]. Leptin and adiponectin are by far the most thoroughly studied adipocytokines. Numerous studies have demonstrated that leptin and adiponectin regulate brain cell proliferation, survival as well as synaptic plasticity via regulation of cellular metabolism and suppression of inflammatory responses [154, 155]. Since leptin and adiponectin play an essential role in brain diseases, these two adipocytokines have been described in detail. In addition, in Table 1, we summarize the representative adipocytokines and their current relevance to obesity and depression.
Table 1. Major adipocytokines and current relevance to obesity and depression.
| NO. | Major adipocytokine | Source | Functions | Relevant to obesity | Relevant to depression | References | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1 | Leptin | white adipocytes, brown adipocytes | regulate energy balance, body weight, metabolism and endocrine function | reduced appetite; elevated energy expenditure; decreased heat loss; reduced insulin secretion, increased by fat mass | leptin levels are higher in patients with moderate to severe depression than in those with mild depression | [20, 162, 168] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2 | Adiponectin | mature white adipocytes, brown adipocytes | enhance insulin sensitivity, decreased inflammation | decreased by obesity, increased insulin sensitivity, decreased gluconeogenesis, increased fatty acid catabolism, | adiponectin works through AdipoR1 receptors on 5-HT neurons to mediate depression-related behaviors in a sex-dependent manner, decreased by oxidative stress, decreased by endoplasmic reticulum stress | [10, 169] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3 | PAI-1 | hepatocytes, endothelial cells, platelets, adipocytes, and cardiac myocytes (in vitro) | the main physiological inhibitor of plasminogen activators/plasmin system; 50-kilodalton glycoprotein of the serine protease inhibitor family; inhibition of the tissue- and urokinase-type plasminogen activator | PAI-1 is frequently elevated in obesity | elevated/ decreased plasma PAI-1 levels in depressed patients; PAI-1 knockout mice are a model of resistance to antidepressants such as SSRIs | [18, 170–173] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4 | MCP-1 | endothelium, fibroblasts, macrophages, monocytes, vascular endothelial cells, smooth muscle cells, astrocytes and microglia | strong monocyte chemotactic active molecule | MCP-1 plays an essential role in obesity-associated monocyte/macrophage infiltration | decreased serum MCP-1 levels in MDD patients | [174, 175] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5 | RBP4 | white and brown adipocytes, liver, adipose tissue | RBP4 is a protein in the lipocalin family and a specific carrier protein of vitamin A in the blood | RBP4 related to systemic insulin resistance, dyslipidemia and obesity | serum RBP4 levels were substantially lower in MDD patients than in controls | [11, 176] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6 | PBEF/ visfatin | high levels in visceral fat, bone marrow, liver tissue and muscle cells, but also a variety of other tissues, including the placenta, kidney, heart and lung. | a pro-inflammatory cytokine that has functions related to cellular metabolism, inflammation and immune regulation. | visfatin reduces apoptosis and necrotic cell death in the CA1 region of the hippocampus of ischemia/reperfusion stroke rats, contributing to a neuroprotective effect | [177–181] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7 | Resistin | white and brown adipocytes, macrophages | presents as trimer/hexamer in plasma, and targets specific receptors TLR4 or Adenylyl CAP1, triggering various intracellular signal transduction pathways to induce vascular inflammation, lipid accumulation, and plaque vulnerability | in rodents, resistin is increased in high-fat/high-carbohydrate-fed, obese states | lower serum resistin levels in MDD patients compared to healthy controls | [182–184] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8 | Apelin (APLN) | adipose tissue also secreted from various tissues in the cardiovascular, digestive, urinary, and CNS | follicle development, regulating glucose and lipid metabolism, modulating insulin secretion, cardiovascular function, blood pressure, angiogenesis, drinking behavior | mediate glucose and lipid metabolism, regulate insulin secretion, plasma apelin concentrations are increased in obesity | Apelin-13 reverses depression-like behavior in CSDS rats model, decreased depressive behavior in sucrose preference and tail suspension tests | [185–188] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9 | Chemerin | white adipose tissue | regulate adipogenesis, insulin sensitivity, and immune response | systemic levels of chemerin are increased in obesity | decreased depressive behavior in forced swim and tail suspension tests, better learning and memory functions | [189–191] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 10 | lipid chaperone FABP4 | highly expressed in white adipocytes, brown adipocytes, macrophages, endothelial cells, | lipid chaperone protein, maintain glucose homeostasis and facilitate communication between energy storage systems and distant organs | long-term involvement of FABP4 in obesity under conditions of immunometabolic stress, regulate metabolic and inflammatory pathways in response to fatty acids | potential role in cell signaling, neuronal development and synaptic function | [192, 193] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 11 | IL-6 | white and brown adipocytes, macrophage, endothelial cells, immune cells | IL-6 is produced rapidly and transiently in response to infection and tissue injury and promotes host defence by stimulating the acute phase response, hematopoiesis and immune response | acutely elevated IL-6 levels aid in fasting or exercise-induced fat mobilization, IL-6-dependent induction of leptin and free fatty acid release from adipocytes | patients with MDD have high levels of various pro-inflammatory cytokines, such as IL-6 | [194–196] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 12 | Nrg4 | activated thermogenic adipose tissue, brown adipocytes, hepatocytes | modulate glucose and lipid metabolism and energy balance | Nrg4 is substantially down-regulated in mouse and human obesity | play a role in neural development and function, Nrg4 is a major novel regulator of dendritic arborizations in the developing cerebral cortex | [197, 198] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 13 | Omentin | omental adipose tissue, macrophage, endothelial cells, the stromal-vascular fraction of visceral adipose tissue | regulate insulin sensitivity, alter inflammatory states | prolonged insulin-glucose infusion in healthy individuals induces a significantly reduced plasma omentin-1 concentration, serum omentin-1 concentrations were significantly lower in overweight and obese subjects than in lean individuals | omentin protects against the decrease in cell viability induced by the pro-inflammatory cytokine TNF-α, omentin promotes the growth and survival of NSCs in vitro by activating the Akt signaling pathway | [199, 200] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 14 | Adipsin | white adipocytes, brown adipocytes | reduced inflammation by chemotaxis, decreased inflammation by clearance of dead cells | decreased by insulin, adipsin serum concentrations are strongly related to obesity, adipsin is downregulated in obesity | mood disorders have lower adipsin levels | [8, 201, 202] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 15 | Vaspin | white and brown adipocytes, preadipocytes, visceral and subcutaneous adipose tissues | play a crucial role in osteogenesis, steroidogenesis, the formation of blood vessels, and food intake, vaspin action on cell apoptosis and proliferation, serine protease inhibitor, improves hyperglycemia | vaspin levels are higher in obese subjects, vaspin mRNA expression was increased in human adipose tissue, vaspin may be a target for the treatment of insulin resistance and inflammation associated with obesity, decreased food intake | elevated vaspin serum concentrations are associated with impaired health levels and leptin serum concentrations | [9, 199, 203, 204] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Note: AdipoR1, adiponectin receptor 1; 5-HT, 5-hydroxytryptamine or serotonin; PAI-1, Plasminogen activator inhibitor type 1; SSRIs, selective serotonin reuptake inhibitors; MCP-1, Monocyte chemoattractant protein-1; MDD, major depressive disorder; RBP4, Retinol Binding Protein 4; PBEF, Pre-B cell colony enhancing factor; TLR4, Toll-Like Receptor 4; CAP1, Cyclase-Associated Protein 1; APLN, Apelin; CNS, central nervous system; CSDS, chronic social defeat stress; FABP4, fatty acid-binding protein 4; IL-6, Interleukin 6; Nrg4, Neuregulin 4; TNF-α, tumor necrosis factor-alpha; NSCs, neural stem cells; AKT, serine-threonine protein kinase. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
In the past, studies on the adipose secretome have mainly focused on polypeptide adipokines. Adipose-derived blood-borne lipids (“lipokines”) are a distinct endocrine factor that has been relatively hotly studied in recent years [156]. Lipokines are fatty acids that influence lipid metabolism and behave like hormones [157]. Interestingly, one of the systemic areas with active lipid metabolism in adipose tissue, which releases diverse lipids into the bloodstream to interact with distant organs [156]. Lipids are also closely related to intracellular fatty acid metabolic pathways and can transfer the intracellular energy of adipocytes to other non-adipose tissues. Unexpectedly, alterations in lipid metabolism might also be connected to the emergence of mental disorders like depression. It has been proposed that increased adipose tissue is linked to chronic inflammation and pro-inflammatory factors that inhibit lipokines production and that chronic inflammation associated with visceral obesity inhibits lipockines production and perpetuates inflammation [158]. There is no doubt that inflammatory dysregulation plays an instrumental role in the pathogenesis of depression, and lipokines may also influence depression via this pathway. Lipokines, mainly including lysophosphatidic acid (LPA) and monounsaturated palmitoleic acid. LPA can interact with the central nervous system and also have endocrine impacts on systemic tissues. In the serum, palmitoleic acid is one of the most prevalent fatty acids. Its circulating levels change according to metabolic conditions. Besides, 12,13-dihydroxy-9Z-octadecenoic acid (12,13-diHOME) is the best-studied lipokines secreted by brown fat, produced by the dihydroxylation of linoleic acid. FAHFAs (fatty acid esters of hydroxy fatty acids) are also lipokine. FAHFAs are secreted by adipocytes into plasma, and in the organism, FAHFA levels in adipose tissue and plasma are associated with insulin sensitivity and a decline in insulin-resistant states. In Table 2, we summarize the representative lipokines and their current relevance to obesity and depression.
Table 2. Major lipokines and current relevance to obesity and depression.
| NO. | Major lipokines | Source | Functions | Relevant to obesity | Relevant to depression | References | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1 | Palmitoleic acid (Palmitoleate) | white adipocytes | decreased inflammation, decreased atherogenesis, increased glucose homeostasis, enhances whole-body insulin sensitivity | PLA (16:1n-7) has hormone-like properties and improves several metabolic parameters that are damaged in obesity | based on fatty acid analysis, the palmitoleic acid was remarkably altered in mice with depressive-like behavior. | [205–207] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2 | DNL FAHFAs | white adipocytes | increased by glucose metabolism, increased by DNL, increased glucose tolerance, increased insulin secretion | dysregulation of DNL is often observed in a variety of metabolic abnormalities, including obesity | expression of de novo fatty acid was enhanced in HFD mice with depression-like behavior | [8, 207, 208] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3 | 12,13-diHOME | brown adipocytes | increased by cold exposure, increased by exercise, increased skeletal muscle fatty acid oxidation, increased fatty acid transport | increasing 12,13- diHOME levels may prevent and treat obesity and metabolic diseases | unknown | [8, 209] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4 | LPA | LPA is found in virtually all biological fluids | influences diverse cellular and organismal processes, including proliferation and growth, survival, development, chemotaxis, vasoregulation, and calcium dynamics | ATX-LPA-LPA1-6 signaling axis in the development of metabolic disorders, including obesity, insulin resistance, as well as damaged glucose homeostasis | LPA management improves depression and anxiety, LPA treatment may regulate the activation of microglia, which plays a critical role in psychiatric disorders such as depression | [210, 211] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Note: DNL, de novo lipogenesis; FAHFAs, fatty acid esters of hydroxy fatty acids; 12,13-diHOME, 12,13-dihydroxy-(9Z)-octadecenoic acid; LPA, lysophosphatidic acid; HFD, high-fat diet; ATX, autotaxin. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The biology of leptin and its relationship to obesity and depression
To date, leptin is by far the finest instance of a successfully moving from discovery to clinical application [9]. The role of leptin in neuroendocrine has been extensively studied. In 1994, Friedman’s team cloned for the first time a hormone-like substance, leptin, secreted by adipocytes and found that it has appetite control and weight reduction effects [159]. The endocrine hormone leptin exerts a pivotal role in modulating food intake and body weight through the action of the hypothalamus [160]. Leptin, a key adipose-derived hormone that regulates eating behavior and body weight, is associated not only with obesity but also with depression. Intraperitoneal injection of leptin into C57BL/6J mice reduced depressive-like behavior in the forced swimming test as well as the tail suspension test [161]. Relevant studies have suggested that leptin levels are higher in patients with moderate to severe depression than in those with mild depression or mild to no depression, and that body mass index (BMI) is higher in patients with moderate to severe depression than in those with mild or mild depression [20]. And after adjusting for multiple factors such as age, gender, and race, leptin levels remained a key predictor of depression [162]. This suggests that leptin may mediate the progression of depression in obese individuals or be a common mechanism causing depression and obesity.
Studies have indicated that the neuroprotective effects of leptin may be related to the leptin/JAK2/STAT3/PGC-1signaling pathway or the leptin-mediated PI3K/Akt/mTOR signaling pathway [163, 164]. Obesity damages leptin-induced regulation of BDNF expression and synaptogenesis, which is thought to be related to the onset of depression. Ginsenoside Rb1 is a major bioactive ingredient of ginseng, and Wu et al. showed that chronic treatment with ginsenoside Rb1 improved central leptin sensitivity, the leptin-JAK2 -STAT3 signaling pathway, as well as the regulation of leptin-induced BDNF expression in the prefrontal cortex of obese mice induced by a HFD [165]. This suggests that supplementation of Rb1 may be a useful way to treat obesity-related psychiatric disorders. Leptin has been proven to affect hippocampal synaptic plasticity [166, 167]. Synaptic plasticity is the most functionally critical form of neuroplasticity, and it exerts an essential role in the neuropathogenesis of several psychiatric disorders. Leptin regulates the efficacy of hippocampal transmission synapses, including LTP and LTD [20]. LTP and LTD are two types of mechanisms that affect impaired cognitive and emotional function in MDD [167]. Under intense, sustained stimulation, increased neuronal firing enhances LTP by strengthening synapses, and LTD causes an activity-dependent decrease in neuronal synaptic efficacy and connectivity [167]. Leptin receptors are widely expressed in various regions of the brain, particularly the hippocampus, which is a crucial region for learning and memory formation as well as emotion regulation. Leptin can cross the BBB and bind to a specific leptin receptor (LepRb) [212]. LepRb deficiency also resulted in memory and cognitive impairment with altered hippocampal synaptic plasticity [213]. It was shown that especially in hippocampal structures, CA1/CA3 regions and dentate gyrus (DG) widely express LepRb mRNA. Several pieces of evidence suggest that leptin is a potent mediator of excitatory synaptic transmission at hippocampal CA1 synapses [213]. The binding of leptin to LepRb isoforms associated with the JAK2-STAT3 signaling pathway causes induction of SOCS3, which terminates JAK2 activity via ubiquitin-mediated degradation of JAK2, thereby terminating the leptin signaling pathway. Enhancing or sustaining the activation of SOCS3, increases obesity-induced leptin concentrations in the blood, leading to leptin resistance. Liu et al. showed that LepRb knockdown-induced depression-like behavior was correlated with STAT3/SOCS3 signaling pathway [19]. LepRb may serve as a novel direction for depression treatment in the future.
The biology of adiponectin and its relationship to obesity and depression
To date, adiponectin is one of the most extensively studied adipokines [214]. Adiponectin, also called 30-kDa adipocyte complement-associated protein (Acrp30), is a type of hormone secreted by adipocytes [215]. The primary role of adiponectin in arcuate nucleus promelanocortin (POMC)-expressing neurons is excitatory, depolarizing neurons, reducing inhibitory synaptic inputs, as well as increasing their responsiveness [216]. The current study suggests that adiponectin directly mediates the cellular activity of arcuate POMC and neuropeptide Y/Agouti-related peptide (NPY/AgRP) neurons [216]. Adiponectin inhibits orexigenic NPY neurons under hypoglycemic conditions and activates orexigenic POMC neurons, thereby attenuating appetite and food intake under fasting or hypoglycemic conditions [166]. In cases of obesity, insulin resistance and type 2 diabetes, blood levels of adiponectin are reduced [215]. Moreover, intracerebroventricular injection of adiponectin -neutralizing antibodies can induce stressful depressive behavior [217]. Adiponectin receptors (AdipoRs) are widely present in the rodent brain, including the hypothalamus, brainstem, prefrontal cortex, as well as hippocampus [155, 217, 218]. While AdipoR1 was highly expressed in the skeletal muscle, AdipoR2 was strongly expressed in the liver [215]. AdipoRs are located on neurons that participated in metabolic regulation in the hypothalamus, including arcuate POMC and NPY/AgRP neurons [216]. Studies have shown that AdipoRs expression is regulated by adiponectin and dopamine signaling pathways [219]. Adiponectin can initiate Notch signaling in the hippocampus by upregulating ADAM10 and Notch1, which are two pivotal molecules in Notch signaling [220]. It has been shown that the removal of AdipoR1 from dopamine neurons could enhance neuronal and anxiogenic responses to suppress stress [219]. The effects of AdipoR1 on neuronal activity and behavior were found to be abolished in dopamine-neuron deficient AdipoR1 mice with VTA infusion of lipocalin [219]. This finding suggests that adiponectin regulates VTA dopamine neuron activity and that AdipoR1 is essential for adipokine-induced inhibition of dopamine neurons directly. Activation of AdipoR1 and AdipoR2 stimulates the activity of AMPK and p38 mitogen-activated protein kinase (p38MAPK), and recently, the p38MAPK signaling pathway was found to regulate adiponectin-induced phosphorylation of the glycogen synthase kinase 3β (GSK-3β) Ser389 inhibitory site [221]. A large body of data suggests that active GSK-3β is associated with increased susceptibility to mood disorders. And inhibition of GSK-3β may be linked to the treatment effects of antidepressants [222]. Thus, adiponectin-induced GSK-3β inhibition may be the basis for the mechanism of antidepressant action of adiponectin. Like LepRs, AdipoRs activate several overlapping signaling cascades, including Janus kinase 2/signal transducer and activator of transcription 3 (JAK2/STAT3), phosphatidylinositol-3-kinase (PI3K), insulin receptor substrate 1/2 (IRS1/2), forkhead box protein O1 (FOXO1) and AMP-activated protein kinase (AMPK) [216, 223]. So far, studies of adiponectin receptors have primarily focused on most aspects of the AMPK signaling pathway regulating metabolism in peripheral tissues [224]. AdipoRs exert a crucial role in the regulation of glucose and fatty acid metabolism via initiating a few signaling cascades that overlap with LepRs [216]. Adiponectin and its adiponectin -receptor interactions are complex processes. In the future, the receptor subtypes and neuronal circuits responsible for the antidepressant-like effects of adiponectin will need to be identified. It was shown that adiponectin expression was reduced in adipose tissue and blood of obese mice, and circulating adiponectin levels were also decreased in obese patients, however, the expression of adiponectin receptors was increased [225–227]. Liu et al. showed that in a chronic social defeat stress (CSDS) model of depression, plasma levels of adiponectin were reduced, which was associated with a reduction in the duration of social interaction [217]. They suggest that reduced adiponectin levels lead to elevated susceptibility to social aversion, pleasure deficit and learned helplessness, as well as to impaired glucocorticoid-mediated negative feedback in the HPA axis [217]. In addition, adiponectin activates neurogenesis in the hippocampus, which may facilitate its antidepressant-like behavioral effects [221].
The biology of palmitoleic acid and its relationship to obesity and depression
Excessive fat accumulation in the body causes adipose tissue dysfunction. This impairment potentially results in elevated release and concentration of circulating free fatty acids, glycerol, hormones, as well as inflammatory cytokines [228]. All these modifications are linked to distinct health problems, such as dyslipidemia, hypertension as well as insulin resistance, collectively known as the “metabolic syndrome” [228]. The ratio of saturated fatty acids to monounsaturated fatty acids is essential in regulating biological membrane fluidity [229]. An imbalance in the ratio of these two may facilitate the development of several diseases, such as diabetes, and cardiovascular disease [229]. Stearoyl-CoA desaturase (SCD) (Δ9 desaturase) and hexadecenoic fatty acids (16:1) are the major monounsaturated fatty acids present in cells and tissues [230, 231] Recently, monounsaturated hexadecenoic fatty acids have been becoming considered biomarkers of health and have vital functions in physiology and pathophysiology [231]. In recent years, palmitoleic acid (cis-9-hexadecenoic acid, 16:1n-7) and its positional isomers 16:1n-9 and 16:1n-10 have gained much attention for their anti-inflammatory properties [231]. PLA (16:1n-7) is the most abundant member of the monounsaturated fatty acids family and the best studied. PLA (16:1n-7) was synthesized from palmitic acid in the presence of SCD-1 and fatty acid desaturase-2 [231]. It can be ingested via the diet and synthesized endogenously from other fatty acids, carbohydrates as well as amino acids [232]. PLA (16:1n-7) is often described as a lipokine capable of regulating a variety of metabolic processes such as increasing insulin sensitivity in muscle, prevention of endoplasmic reticulum stress, β-cell proliferation, and lipogenic activity in white adipocytes [157]. Cao et al. showed that the release of this fatty acid from adipose tissue inhibited steatosis in the liver and modified insulin signaling in muscle [157]. Collectively, this study showed that PLA (16:1n-7) acts as an anti-inflammatory agent in the adipose tissue of mice and contributes to reducing the effects of obesity. This shows that PLA (16:1n-7) inhibits adipocyte cytokine expression, while palmitic acid does not inhibit adipocyte cytokine expression, suggesting that adipocytes are the primary target of PLA (16:1n-7) [157]. Lopes A et al. showed that PLA (16:1n-7) improves systemic insulin sensitivity and glucose uptake into adipose tissue by regulating GLUT-4 and AMPK phosphorylation in HFD-fed mice [233]. Another study also discovered that PLA (16:1n-7) treatment may prevent the enhancement of transcription factors CEBPα and PPARγ in subcutaneous adipocytes in the inguinal groin of HFD-treated mice [234]. PLA (16:1n-7), described as a lipotropic hormone, may play diverse roles according to the organ and disease model under study [207]. According to fatty acid analysis, palmitoleic acid content was significantly altered in mice with depressive-like behavior. Moreover, the expression of acetyl coenzyme a carboxylase (ACC), SCD1, and fatty acid desaturase 1 and 2 (FADS1 and FADS2), which are involved in the fatty acid synthesis, fatty acid desaturation, and arachidonic acid synthesis, was enhanced in HFD mice with depressive-like behavior [207]. Thus, it is hypothesized that HFD-induced disturbances in lipid metabolism speed up the development of depression-like behaviors. Although beneficial effects of PLA have been observed in both in vivo and in vitro studies, there have not been sufficient human intervention studies to fully comprehend the physiological effects of palmitoleic acid [206]. Hence, more human-based studies are required to determine whether PLA has promising therapeutic potential.
The biology of lysophosphatidic acid and its relationship to obesity and depression
LPA, also called monoacyl-sn-glycero-3-phosphate, is a lysophospholipid with a glycerol phosphate head group and a fatty acid moiety. It is not a single chemical entity, but represents a class of biomolecules with different fatty acid chain lengths and saturations [235]. Autotaxin (ATX) is a secreted enzyme that hydrolyzes lysophosphatidylcholine to generate LPA. A growing number of studies suggest that the ATX-LPA axis is involved in obesity and its associated metabolic complications [236, 237]. LPA is a lipid mediator which is produced by adipocytes through specific G-protein-coupled receptors and its synthesis is regulated in obesity. Rancoule et al. showed that reduced adipocyte LPA production was correlated with improved glucose tolerance in HFD-fed obese mice [238]. This suggests that LPA harms glucose homeostasis.
Synaptic signaling is a plastic dynamic process crucial for information processing at the level of brain cells and neuronal networks and is critical for regulating neuronal excitability as well as brain information processing [239]. The LPA-triggered signaling pathway induces rapid and reversible inhibition of excitatory and inhibitory postsynaptic currents. In excitatory synapses, the LPA1/Gαi/o protein/phospholipase C/myosin light chain kinase cascade acts at presynaptic sites [239]. LPA can act as potential partial messengers, modulating synaptic strength to accommodate the prior activity of neurons [239]. LPA primarily mediates its action by activating six known G protein-coupled receptors (GPCRs), and the protein products are named LPA1 to LPA6 [240]. LPA is activating six LPA receptors (LPAR1-6) and modulates different cellular activities, i.e. cell proliferation, cytoprotection as well as wound healing. LPA receptors are classically seven-transmembrane GPCRs activating heterotrimeric G proteins to signal transduction within the cell [235]. It is well documented that LPA and LPA receptor signaling pathways are required for the formation of mature synaptic connections, particularly glutamatergic synapses [241]. As one of the six characteristic G protein-coupled receptors (LPA1-6), the LPA1 receptor through which lysophosphatidic acid serves as an intracellular signaling molecule. Studies have shown that the deletion of LPA1 receptors causes anxiety as well as several behavioral and neurobiological changes that are closely related to depression [242]. Endogenous LPA signaling mediates activity-dependent inhibition primarily through LPA1 in an experimental model of synaptic plasticity [239].
In the hippocampus, genetic deletion of Lpar1 leads to more immature dendritic spines in CA1 pyramidal cells and reduces matrix metalloproteinase 9 (MMP-9), which has been proven to be engaged in regulating synaptic plasticity [243]. In zebrafish in vivo experiments, the Lpar3 gene exerts novel roles in regulating behaviors such as anxiety, social interactions, circadian rhythmic motor activity, and memory retention [244]. Gintonin, an exogenous LPA receptor ligand, was isolated from P. ginseng. Kim et al. suggested that gintonin-enriched fraction may be associated with the relief of depression-related symptoms induced by ginseng extract [245]. All this evidence suggests that LPA plays an essential role in both obesity and depression.
Potential and future treatment of depression and obesity
The multifaceted effects of adipokines and lipokines on neurological and brain health and the dysregulation of adipokine and lipokine secretion may contribute to the co-morbidity of obesity and depression [166]. Exploring the pathogenesis of obesity and depression from the perspective of multiple adipokines and lipokines has beneficial implications for the future treatment of obesity and depression. Adipokines and lipokines such as leptin, adiponectin, PLA, and LPA may be critical targets for the future treatment of obesity and depression. Clinical studies have shown that individuals with POMC or LepRb deficiency typically experience severe obesity, bulimia and comorbidities, which can severely impact the patient’s quality of life and depressive behavior [246].
Diet and obesity have been proven to have a direct effect on mood, and stress-related mental diseases might result in alterations in eating patterns that affect weight [247]. Recent studies suggest that dietary interventions and energy restriction may help prevent depression and anxiety, which would serve as complementary therapies [248]. Ganoderma lucidum is a medicinal mushroom commonly used to improve quality of life, promote health, as well as enhance vitality. Studies have shown that ethanolic extract of Ganoderma lucidum ethanol extract (EEGL) has significant effects on feeding behavioral parameters, depression-like symptoms and locomotor activity in Swiss mice, as reflected by a significant decrease in body weight gain and food intake, a dose-dependent increase in water intake, and a decrease in immobility time in the forced swim test (FST) and the tail suspension test (TST) in Swiss mice [249]. These findings suggest that EEGL can reduce body weight gain and produce antidepressant-like effects.
Unhealthy eating patterns might be connected with an increased risk of depression or anxiety, while healthy eating patterns might reduce that risk. The first human study to show a link between diet and hippocampus volume mirrored findings from previous preclinical investigations in animal models, in which low nutritional food intake and excessive unhealthy food intake were independently linked with decreased left hippocampal volume, respectively [250]. Recent clinical studies have shown that diet can influence the physiological and immune functions of the body and has potential therapeutic strategies. The study found that 83% of participants who adhered to a ketogenic diet experienced significant reductions in fat mass and nearly 50% decreases in self-reported fatigue and depression scores over the study period [251].
Functional brain imaging research confirms that image and verbal cues associated with foods and beverages high in sugar produce higher preferences and higher emotional activation, making it harder for overweight people to resist eating unhealthy foods [252]. Several variables, including inflammation, oxidative stress, and insulin resistance, have been postulated to contribute to diet-induced brain damage, all of which can be affected by dietary consumption and are related to the onset of depression [48]. Maintaining a balanced diet with anti-inflammatory properties may aid in the prevention of depressive symptoms, particularly in men, smokers, and those who are inactive [253].
In observational studies, adherence to a healthy diet, particularly a traditional Mediterranean diet or avoidance of a pro-inflammatory diet, appears to have a protective effect against depression [16]. A population-based cohort study showed that people who adhered to a Mediterranean diet during midlife had a lower risk of depression later in life [16, 254]. Although the relationship between obesity and depression is known, there is little research on the clinical benefits of nutritional therapy for obese patients. A clinical study evaluating the effects of a traditional Brazilian diet (DieTBra) and extra virgin olive oil (EVOO) on anxiety and depressive symptoms in severely obese participants showed that both DieTBra and olive oil interventions were effective in reducing anxiety and depressive symptoms in severely obese adults and that these interventions could be combined with clinical protocols for the treatment of anxiety and depressive symptoms in severely obese patients [255].
Current evidence suggests that diet quality is a modifiable risk factor for affective disorders, however, further research is needed to investigate the impact of dietary patterns and weight loss on improving psychological symptoms. Rodriguez-Lozada et al. randomly assigned overweight and obese participants (n=305) to two low-calorie diets with different macronutrient distributions: a moderately high protein diet and a low-fat diet for 16 weeks to assess the effects of prescribed energy restriction on anxiety and depressive symptoms in overweight and obese participants, as well as some baseline potential predictive value of psychological characteristics for weight loss [256]. The nutritional intervention demonstrated beneficial effects of weight loss on trait anxiety scores in women, depression scores in all populations, and especially in women and subjects following a low-fat diet. In addition, weight loss can be predicted by anxiety status at baseline, which occurs primarily in women and those on a low-fat diet, and this trial suggests that weight loss triggers improvements in psychological traits, while anxiety symptoms predict those volunteers who benefit most from a balanced calorie restriction intervention, which would help to personalize precise nutrition [256]. In addition, a randomized clinical trial study by Hariri et al. demonstrated the beneficial effects of sumac (Rhus coriaria L.) supplementation with a calorie-restricted diet on anthropometric indices, oxidative stress, and inflammation in overweight or obese women with depression [257].
Probiotics have been demonstrated to have antidepressant responses and anti-inflammatory effects. Borges et al. conducted a systematic review and meta-analysis of overweight or obese patients to determine the effects of prebiotics on blood biomarkers of obesity, depression, and anxiety (including ACTH, cortisol, leptin, ghrelin, thyroid stimulating hormone (TSH), parathyroid hormone (PTH), vitamin D, and BDNF) [258]. Prebiotics were found to possibly facilitate the regulation of blood concentrations of ghrelin and CRP in overweight or obese individuals. A double-blind, randomized, placebo-controlled trial was conducted in obese men (n = 45) and women (n = 60), consisting of a 12-week weight loss period based on moderate energy restriction and a 12-week weight maintenance period [259]. Each subject consumed two capsules per day of either a placebo or probiotic supplement Lactobacillus rhamnosus CGMCC1.3724 (LPR) [259]. During both phases of the program, LPR supplementation increased weight loss in women, which was associated with an increased desire to eat on an empty stomach. In addition, the LPR female group showed a more pronounced decrease in food cravings as well as a decrease in Beck Depression Scale scores [259]. Significant benefits of LPR on fasting satiety and cognitive restraint were also observed in men [259]. These clinical observations support the hypothesis that the gut-brain axis may influence appetite control and related behaviors in obesity management. Hulkkonen et al. studied the efficacy of probiotics and/or fish oil in improving prenatal and postnatal depression and anxiety symptoms and found that diet quality was negatively associated with depressive symptoms in early pregnancy and 6 months postpartum and with anxiety symptoms in early pregnancy [260].
Probiotics, prebiotics, and mushroom extracts may be used to simultaneously prevent and treat obesity and depression, and further research is required to optimally use these substances in humans [30]. Furthermore, the using dietary interventions may demonstrate to be an appealing and cost-effective alternative or adjunctive treatment for the clinical management of these conditions. Prospective investigations of the connection between diet and mental health ought to use clearer definitions to define diet and include or control for significant confounding factors. Regulation of gut microbiota might be a new strategy for the treatment of both neuroinflammation and depression [30]. Biochemical markers are positively associated with the development and severity of obesity, depression as well as anxiety, and are regulated by changes in the composition of the gut microbiota [258]. The gut microbiota is a potentially important regulating pathway between diet and brain health. Altered gut microbiota may be a therapeutic strategy for depression and obesity. FMT is currently the most effective gut microbiota intervention [261]. FMT may be a promising therapeutic choice for neurological disorders, however, the available evidence remains sparse, with a limited number of human studies conducted or ongoing, and for some diseases, only animal trials have been conducted. To further elucidate the role of FMT in neurological disorders, large-scale double-blind randomized controlled trials are required [261].
With the growing insight into the mechanisms underlying the complex interactions between diet and gut microbiota and their effects on depression, specific dietary patterns that aid in the prevention of anxiety and mood disorders may be identified [248]. Furthermore, preclinical studies have shown that exercise can improve various types of behaviors, such as depression and anxiety [262–264]. Morgan et al. showed that aerobic exercise had significant beneficial effects on depression-like, anxiety-like and cognitive-like behaviors during the healthy adult lifespan of C57BL/6 mice [265]. A combined treatment for both obesity and depression improves weight and mood risk factors more than treatment for each disease alone [266]. In a randomized controlled trial of 63 overweight/obese participants, Pilates and aerobic training were found to improve depression, anxiety levels and quality of life in overweight and obese people [267]. Recent clinical studies have shown that virtual reality exercise programs have a positive impact on BMI, depression levels, exercise enjoyment and exercise immersion in overweight middle-aged women. This is an effective home exercise program for people undergoing obesity management [268]. Developing tailored treatments according to a personalized medical approach to the biology of obesity-depression co-morbidities may ultimately benefit the patients involved. In Figure 3, we summarized the potential and possible future treatment options for obesity and depression (Figure 3).
Figure 3. Map of potential and possible future treatment options for depression and obesity comorbidity. LTP, long-term potentiation; LTD, long-term depression; HPA axis, hypothalamo-pituitary-adrenal axis.
Overview
Acute/chronic inflammation, gut microbiota imbalance, gut-brain dysfunction, diminished neuroplasticity and HPA axis dysfunction are common mechanisms in the pathogenesis of depression, and several of these effects often co-occur in depression and also affect obesity at the same time. These factors would potentially provide a biologically based multi-level description of obesity and depression. The characterisation of the mechanisms of action of adipokines and lipokines and the identification of molecular targets of adipokines and lipokines will provide new ideas for this study of the co-morbidities of obesity and depression. Further study of adipokines and lipokines and their interaction with the brain may provide new therapeutic targets for the treatment of depression. Probiotics, herbal extracts and mushroom extracts may be used to prevent and treat the co-morbidities of obesity and depression. Modulation of the gut microbiota may be a novel strategy for the treatment of neuroinflammatory and depressive disorders. FMT may regain clinical attention as a therapy to restore gut flora for the clinical treatment of obesity and depression. The selection and development of treatments tailored to their biology, according to a personalised medicine approach, may ultimately benefit patients with depression and obesity.
Author Contributions
XYF and FYZ drafted the manuscript. YCW, WX and QQL made critical revisions. RJC made substantial revisions to the logic of the article. WY completed the conception, design, editing and acquisition of funding. All authors approved the final version of the manuscript for submission.
Conflicts of Interest
The authors declare no conflicts of interest.
Funding
This work was supported by grants from the Natural Science Foundation of China (Grant No.81971276), Jilin Science and Technology Agency (20220203124SF, 20220204031YY, 20210204028YY, YDZJ202301ZYTS079, 20210204006YY, YDZJ202301ZYTS058) and Jilin University Bethune Program Project (2022B35).
References
- 1. Obesity: preventing and managing the global epidemic. Report of a WHO consultation. World Health Organ Tech Rep Ser. 2000; 894. [PubMed]
- 2. Petrelli F, Cortellini A, Indini A, Tomasello G, Ghidini M, Nigro O, Salati M, Dottorini L, Iaculli A, Varricchio A, Rampulla V, Barni S, Cabiddu M, et al. Association of Obesity With Survival Outcomes in Patients With Cancer: A Systematic Review and Meta-analysis. JAMA Netw Open. 2021; 4:e213520. https://doi.org/10.1001/jamanetworkopen.2021.3520 [PubMed]
- 3. Piché ME, Tchernof A, Després JP. Obesity Phenotypes, Diabetes, and Cardiovascular Diseases. Circ Res. 2020; 126:1477–500. https://doi.org/10.1161/CIRCRESAHA.120.316101 [PubMed]
- 4. Fulton S, Décarie-Spain L, Fioramonti X, Guiard B, Nakajima S. The menace of obesity to depression and anxiety prevalence. Trends Endocrinol Metab. 2022; 33:18–35. https://doi.org/10.1016/j.tem.2021.10.005 [PubMed]
- 5. Milaneschi Y, Simmons WK, van Rossum EF, Penninx BW. Depression and obesity: evidence of shared biological mechanisms. Mol Psychiatry. 2019; 24:18–33. https://doi.org/10.1038/s41380-018-0017-5 [PubMed]
- 6. Conway B, Rene A. Obesity as a disease: no lightweight matter. Obes Rev. 2004; 5:145–51. https://doi.org/10.1111/j.1467-789X.2004.00144.x [PubMed]
- 7. Weber-Hamann B, Werner M, Hentschel F, Bindeballe N, Lederbogen F, Deuschle M, Heuser I. Metabolic changes in elderly patients with major depression: evidence for increased accumulation of visceral fat at follow-up. Psychoneuroendocrinology. 2006; 31:347–54. https://doi.org/10.1016/j.psyneuen.2005.08.014 [PubMed]
- 8. Scheja L, Heeren J. The endocrine function of adipose tissues in health and cardiometabolic disease. Nat Rev Endocrinol. 2019; 15:507–24. https://doi.org/10.1038/s41574-019-0230-6 [PubMed]
- 9. Fasshauer M, Blüher M. Adipokines in health and disease. Trends Pharmacol Sci. 2015; 36:461–70. https://doi.org/10.1016/j.tips.2015.04.014 [PubMed]
- 10. Funcke JB, Scherer PE. Beyond adiponectin and leptin: adipose tissue-derived mediators of inter-organ communication. J Lipid Res. 2019; 60:1648–84. https://doi.org/10.1194/jlr.R094060 [PubMed]
- 11. Yao Q, Li Y. Study of decreased serum levels of retinol binding protein 4 in major depressive disorder. J Psychiatr Res. 2020; 129:24–30. https://doi.org/10.1016/j.jpsychires.2020.05.030 [PubMed]
- 12. Patel V, Chisholm D, Parikh R, Charlson FJ, Degenhardt L, Dua T, Ferrari AJ, Hyman S, Laxminarayan R, Levin C, Lund C, Medina Mora ME, Petersen I, et al, and DCP MNS Author Group. Addressing the burden of mental, neurological, and substance use disorders: key messages from Disease Control Priorities, 3rd edition. Lancet. 2016; 387:1672–85. https://doi.org/10.1016/S0140-6736(15)00390-6 [PubMed]
- 13. Herrman H, Kieling C, McGorry P, Horton R, Sargent J, Patel V. Reducing the global burden of depression: a Lancet-World Psychiatric Association Commission. Lancet. 2019; 393:e42–3. https://doi.org/10.1016/S0140-6736(18)32408-5 [PubMed]
- 14. Bergantin LB. The clinical link between depression and obesity: Role of Ca2+/cAMP signalling. Psychiatry Res. 2020; 291:113167. https://doi.org/10.1016/j.psychres.2020.113167 [PubMed]
- 15. Lv N, Kringle EA, Ma J. Integrated Behavioral Interventions for Adults with Comorbid Obesity and Depression: a Systematic Review. Curr Diab Rep. 2022; 22:157–68. https://doi.org/10.1007/s11892-022-01458-z [PubMed]
- 16. Lassale C, Batty GD, Baghdadli A, Jacka F, Sánchez-Villegas A, Kivimäki M, Akbaraly T. Healthy dietary indices and risk of depressive outcomes: a systematic review and meta-analysis of observational studies. Mol Psychiatry. 2019; 24:965–86. https://doi.org/10.1038/s41380-018-0237-8 [PubMed]
- 17. Ji L, Zhang L, Liang Z, Zhong S, Liu X, Liu Z, Poon WS, Song Y, Chen B, Wang R. Role of omentin-1 in susceptibility to anxiety and depression like behaviors. Mol Cell Endocrinol. 2023:111990. [Epub ahead of print]. https://doi.org/10.1016/j.mce.2023.111990 [PubMed]
- 18. Party H, Dujarrier C, Hébert M, Lenoir S, Martinez de Lizarrondo S, Delépée R, Fauchon C, Bouton MC, Obiang P, Godefroy O, Save E, Lecardeur L, Chabry J, et al. Plasminogen Activator Inhibitor-1 (PAI-1) deficiency predisposes to depression and resistance to treatments. Acta Neuropathol Commun. 2019; 7:153. https://doi.org/10.1186/s40478-019-0807-2 [PubMed]
- 19. Liu W, Liu J, Xia J, Xue X, Wang H, Qi Z, Ji L. Leptin receptor knockout-induced depression-like behaviors and attenuated antidepressant effects of exercise are associated with STAT3/SOCS3 signaling. Brain Behav Immun. 2017; 61:297–305. https://doi.org/10.1016/j.bbi.2017.01.001 [PubMed]
- 20. Ge T, Fan J, Yang W, Cui R, Li B. Leptin in depression: a potential therapeutic target. Cell Death Dis. 2018; 9:1096. https://doi.org/10.1038/s41419-018-1129-1 [PubMed]
- 21. Tartt AN, Mariani MB, Hen R, Mann JJ, Boldrini M. Dysregulation of adult hippocampal neuroplasticity in major depression: pathogenesis and therapeutic implications. Mol Psychiatry. 2022; 27:2689–99. https://doi.org/10.1038/s41380-022-01520-y [PubMed]
- 22. Gerhard DM, Wohleb ES, Duman RS. Emerging treatment mechanisms for depression: focus on glutamate and synaptic plasticity. Drug Discov Today. 2016; 21:454–64. https://doi.org/10.1016/j.drudis.2016.01.016 [PubMed]
- 23. Beurel E, Toups M, Nemeroff CB. The Bidirectional Relationship of Depression and Inflammation: Double Trouble. Neuron. 2020; 107:234–56. https://doi.org/10.1016/j.neuron.2020.06.002 [PubMed]
- 24. Boku S, Nakagawa S, Toda H, Hishimoto A. Neural basis of major depressive disorder: Beyond monoamine hypothesis. Psychiatry Clin Neurosci. 2018; 72:3–12. https://doi.org/10.1111/pcn.12604 [PubMed]
- 25. Krishnan V, Nestler EJ. The molecular neurobiology of depression. Nature. 2008; 455:894–902. https://doi.org/10.1038/nature07455 [PubMed]
- 26. Fries GR, Saldana VA, Finnstein J, Rein T. Molecular pathways of major depressive disorder converge on the synapse. Mol Psychiatry. 2023; 28:284–97. https://doi.org/10.1038/s41380-022-01806-1 [PubMed]
- 27. Cox AJ, West NP, Cripps AW. Obesity, inflammation, and the gut microbiota. Lancet Diabetes Endocrinol. 2015; 3:207–15. https://doi.org/10.1016/S2213-8587(14)70134-2 [PubMed]
- 28. Ying W, Fu W, Lee YS, Olefsky JM. The role of macrophages in obesity-associated islet inflammation and β-cell abnormalities. Nat Rev Endocrinol. 2020; 16:81–90. https://doi.org/10.1038/s41574-019-0286-3 [PubMed]
- 29. Plovier H, Everard A, Druart C, Depommier C, Van Hul M, Geurts L, Chilloux J, Ottman N, Duparc T, Lichtenstein L, Myridakis A, Delzenne NM, Klievink J, et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat Med. 2017; 23:107–13. https://doi.org/10.1038/nm.4236 [PubMed]
- 30. Schachter J, Martel J, Lin CS, Chang CJ, Wu TR, Lu CC, Ko YF, Lai HC, Ojcius DM, Young JD. Effects of obesity on depression: A role for inflammation and the gut microbiota. Brain Behav Immun. 2018; 69:1–8. https://doi.org/10.1016/j.bbi.2017.08.026 [PubMed]
- 31. Koopman M, El Aidy S, and MIDtrauma consortium. Depressed gut? The microbiota-diet-inflammation trialogue in depression. Curr Opin Psychiatry. 2017; 30:369–77. https://doi.org/10.1097/YCO.0000000000000350 [PubMed]
- 32. Kumar A, Pramanik J, Goyal N, Chauhan D, Sivamaruthi BS, Prajapati BG, Chaiyasut C. Gut Microbiota in Anxiety and Depression: Unveiling the Relationships and Management Options. Pharmaceuticals (Basel). 2023; 16:565. https://doi.org/10.3390/ph16040565 [PubMed]
- 33. Takeuchi T, Kameyama K, Miyauchi E, Nakanishi Y, Kanaya T, Fujii T, Kato T, Sasaki T, Tachibana N, Negishi H, Matsui M, Ohno H. Fatty acid overproduction by gut commensal microbiota exacerbates obesity. Cell Metab. 2023; 35:361–75.e9. https://doi.org/10.1016/j.cmet.2022.12.013 [PubMed]
- 34. Liu Z, Li L, Ma S, Ye J, Zhang H, Li Y, Sair AT, Pan J, Liu X, Li X, Yan S, Liu X. High-Dietary Fiber Intake Alleviates Antenatal Obesity-Induced Postpartum Depression: Roles of Gut Microbiota and Microbial Metabolite Short-chain Fatty Acid Involved. J Agric Food Chem. 2020; 68:13697–710. https://doi.org/10.1021/acs.jafc.0c04290 [PubMed]
- 35. Marx W, Lane M, Hockey M, Aslam H, Berk M, Walder K, Borsini A, Firth J, Pariante CM, Berding K, Cryan JF, Clarke G, Craig JM, et al. Diet and depression: exploring the biological mechanisms of action. Mol Psychiatry. 2021; 26:134–50. https://doi.org/10.1038/s41380-020-00925-x [PubMed]
- 36. Sun L, Ma L, Zhang H, Cao Y, Wang C, Hou N, Huang N, von Deneen KM, Zhao C, Shi Y, Pan Y, Wang M, Ji G, Nie Y. Fto Deficiency Reduces Anxiety- and Depression-Like Behaviors in Mice via Alterations in Gut Microbiota. Theranostics. 2019; 9:721–33. https://doi.org/10.7150/thno.31562 [PubMed]
- 37. Duman RS, Aghajanian GK, Sanacora G, Krystal JH. Synaptic plasticity and depression: new insights from stress and rapid-acting antidepressants. Nat Med. 2016; 22:238–49. https://doi.org/10.1038/nm.4050 [PubMed]
- 38. Ma J, Hu W, Zhang D, Xie J, Duan C, Liu Y, Wang Y, Xu X, Cheng K, Jin B, Zhang Y, Zhuang R. CD226 knockout alleviates high-fat diet induced obesity by suppressing proinflammatory macrophage phenotype. J Transl Med. 2021; 19:477. https://doi.org/10.1186/s12967-021-03150-4 [PubMed]
- 39. Lee ES, Kwon MH, Kim HM, Woo HB, Ahn CM, Chung CH. Curcumin analog CUR5-8 ameliorates nonalcoholic fatty liver disease in mice with high-fat diet-induced obesity. Metabolism. 2020; 103:154015. https://doi.org/10.1016/j.metabol.2019.154015 [PubMed]
- 40. Almeida FB, Pinna G, Barros HMT. The Role of HPA Axis and Allopregnanolone on the Neurobiology of Major Depressive Disorders and PTSD. Int J Mol Sci. 2021; 22:5495. https://doi.org/10.3390/ijms22115495 [PubMed]
- 41. Dallman MF. Stress-induced obesity and the emotional nervous system. Trends Endocrinol Metab. 2010; 21:159–65. https://doi.org/10.1016/j.tem.2009.10.004 [PubMed]
- 42. Kiecolt-Glaser JK, Derry HM, Fagundes CP. Inflammation: depression fans the flames and feasts on the heat. Am J Psychiatry. 2015; 172:1075–91. https://doi.org/10.1176/appi.ajp.2015.15020152 [PubMed]
- 43. Wichers MC, Kenis G, Koek GH, Robaeys G, Nicolson NA, Maes M. Interferon-alpha-induced depressive symptoms are related to changes in the cytokine network but not to cortisol. J Psychosom Res. 2007; 62:207–14. https://doi.org/10.1016/j.jpsychores.2006.09.007 [PubMed]
- 44. Lu Y, Liu BP, Tan CT, Pan F, Larbi A, Ng TP. Lifetime pathogen burden, inflammatory markers, and depression in community-dwelling older adults. Brain Behav Immun. 2022; 102:124–34. https://doi.org/10.1016/j.bbi.2022.02.020 [PubMed]
- 45. Kappelmann N, Arloth J, Georgakis MK, Czamara D, Rost N, Ligthart S, Khandaker GM, Binder EB. Dissecting the Association Between Inflammation, Metabolic Dysregulation, and Specific Depressive Symptoms: A Genetic Correlation and 2-Sample Mendelian Randomization Study. JAMA Psychiatry. 2021; 78:161–70. https://doi.org/10.1001/jamapsychiatry.2020.3436 [PubMed]
- 46. Zhou Y, Wang C, Lan X, Li H, Chao Z, Ning Y. Plasma inflammatory cytokines and treatment-resistant depression with comorbid pain: improvement by ketamine. J Neuroinflammation. 2021; 18:200. https://doi.org/10.1186/s12974-021-02245-5 [PubMed]
- 47. Carboni L, McCarthy DJ, Delafont B, Filosi M, Ivanchenko E, Ratti E, Learned SM, Alexander R, Domenici E. Biomarkers for response in major depression: comparing paroxetine and venlafaxine from two randomised placebo-controlled clinical studies. Transl Psychiatry. 2019; 9:182. https://doi.org/10.1038/s41398-019-0521-7 [PubMed]
- 48. Slavich GM, Irwin MR. From stress to inflammation and major depressive disorder: a social signal transduction theory of depression. Psychol Bull. 2014; 140:774–815. https://doi.org/10.1037/a0035302 [PubMed]
- 49. Zhao F, Li B, Yang W, Ge T, Cui R. Brain-immune interaction mechanisms: Implications for cognitive dysfunction in psychiatric disorders. Cell Prolif. 2022; 55:e13295. https://doi.org/10.1111/cpr.13295 [PubMed]
- 50. Delpech JC, Madore C, Nadjar A, Joffre C, Wohleb ES, Layé S. Microglia in neuronal plasticity: Influence of stress. Neuropharmacology. 2015; 96:19–28. https://doi.org/10.1016/j.neuropharm.2014.12.034 [PubMed]
- 51. Vandanmagsar B, Youm YH, Ravussin A, Galgani JE, Stadler K, Mynatt RL, Ravussin E, Stephens JM, Dixit VD. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat Med. 2011; 17:179–88. https://doi.org/10.1038/nm.2279 [PubMed]
- 52. Marques M, Perre S, Aertgeerts A, Derde S, Güiza F, Casaer MP, Hermans G, Van den Berghe G, Langouche L. Critical illness induces nutrient-independent adipogenesis and accumulation of alternatively activated tissue macrophages. Crit Care. 2013; 17:R193. https://doi.org/10.1186/cc12887 [PubMed]
- 53. Gómez-Apo E, Mondragón-Maya A, Ferrari-Díaz M, Silva-Pereyra J. Structural Brain Changes Associated with Overweight and Obesity. J Obes. 2021; 2021:6613385. https://doi.org/10.1155/2021/6613385 [PubMed]
- 54. Caputa G, Matsushita M, Sanin DE, Kabat AM, Edwards-Hicks J, Grzes KM, Pohlmeyer R, Stanczak MA, Castoldi A, Cupovic J, Forde AJ, Apostolova P, Seidl M, et al. Intracellular infection and immune system cues rewire adipocytes to acquire immune function. Cell Metab. 2022; 34:747–60.e6. https://doi.org/10.1016/j.cmet.2022.04.008 [PubMed]
- 55. Guillemot-Legris O, Muccioli GG. Obesity-Induced Neuroinflammation: Beyond the Hypothalamus. Trends Neurosci. 2017; 40:237–53. https://doi.org/10.1016/j.tins.2017.02.005 [PubMed]
- 56. Li C, Xu MM, Wang K, Adler AJ, Vella AT, Zhou B. Macrophage polarization and meta-inflammation. Transl Res. 2018; 191:29–44. https://doi.org/10.1016/j.trsl.2017.10.004 [PubMed]
- 57. Ullah R, Rauf N, Nabi G, Yi S, Yu-Dong Z, Fu J. Mechanistic insight into high-fat diet-induced metabolic inflammation in the arcuate nucleus of the hypothalamus. Biomed Pharmacother. 2021; 142:112012. https://doi.org/10.1016/j.biopha.2021.112012 [PubMed]
- 58. Jais A, Paeger L, Sotelo-Hitschfeld T, Bremser S, Prinzensteiner M, Klemm P, Mykytiuk V, Widdershooven PJM, Vesting AJ, Grzelka K, Minère M, Cremer AL, Xu J, et al. PNOCARC Neurons Promote Hyperphagia and Obesity upon High-Fat-Diet Feeding. Neuron. 2020; 106:1009–25.e10. https://doi.org/10.1016/j.neuron.2020.03.022 [PubMed]
- 59. Waise TMZ, Toshinai K, Naznin F, NamKoong C, Md Moin AS, Sakoda H, Nakazato M. One-day high-fat diet induces inflammation in the nodose ganglion and hypothalamus of mice. Biochem Biophys Res Commun. 2015; 464:1157–62. https://doi.org/10.1016/j.bbrc.2015.07.097 [PubMed]
- 60. Lama A, Pirozzi C, Severi I, Morgese MG, Senzacqua M, Annunziata C, Comella F, Del Piano F, Schiavone S, Petrosino S, Mollica MP, Diano S, Trabace L, et al. Palmitoylethanolamide dampens neuroinflammation and anxiety-like behavior in obese mice. Brain Behav Immun. 2022; 102:110–23. https://doi.org/10.1016/j.bbi.2022.02.008 [PubMed]
- 61. Kelley N, Jeltema D, Duan Y, He Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int J Mol Sci. 2019; 20:3328. https://doi.org/10.3390/ijms20133328 [PubMed]
- 62. Hou J, Li AL, Xiong WQ, Chen R. Hsa Circ 001839 Promoted Inflammation in Renal Ischemia-Reperfusion Injury Through NLRP3 by miR-432-3p. Nephron. 2021; 145:540–52. https://doi.org/10.1159/000515279 [PubMed]
- 63. Li S, Fang Y, Zhang Y, Song M, Zhang X, Ding X, Yao H, Chen M, Sun Y, Ding J, Wang Q, Lu M, Wu G, Hu G. Microglial NLRP3 inflammasome activates neurotoxic astrocytes in depression-like mice. Cell Rep. 2022; 41:111532. https://doi.org/10.1016/j.celrep.2022.111532 [PubMed]
- 64. Li Y, Chen H, Wang J, Wang J, Niu X, Wang C, Qin D, Li F, Wang Y, Xiong J, Liu S, Huang L, Zhang X, et al. Inflammation-activated C/EBPβ mediates high-fat diet-induced depression-like behaviors in mice. Front Mol Neurosci. 2022; 15:1068164. https://doi.org/10.3389/fnmol.2022.1068164 [PubMed]
- 65. Pignataro P, Dicarlo M, Zerlotin R, Zecca C, Dell’Abate MT, Buccoliero C, Logroscino G, Colucci S, Grano M. FNDC5/Irisin System in Neuroinflammation and Neurodegenerative Diseases: Update and Novel Perspective. Int J Mol Sci. 2021; 22:1605. https://doi.org/10.3390/ijms22041605 [PubMed]
- 66. Parkhurst CN, Yang G, Ninan I, Savas JN, Yates JR 3rd, Lafaille JJ, Hempstead BL, Littman DR, Gan WB. Microglia promote learning-dependent synapse formation through brain-derived neurotrophic factor. Cell. 2013; 155:1596–609. https://doi.org/10.1016/j.cell.2013.11.030 [PubMed]
- 67. Ceasrine AM, Bilbo SD. Dietary fat: a potent microglial influencer. Trends Endocrinol Metab. 2022; 33:196–205. https://doi.org/10.1016/j.tem.2021.12.005 [PubMed]
- 68. Lam YY, Tsai SF, Chen PC, Kuo YM, Chen YW. Pioglitazone rescues high-fat diet-induced depression-like phenotypes and hippocampal astrocytic deficits in mice. Biomed Pharmacother. 2021; 140:111734. https://doi.org/10.1016/j.biopha.2021.111734 [PubMed]
- 69. Chesnokova V, Pechnick RN, Wawrowsky K. Chronic peripheral inflammation, hippocampal neurogenesis, and behavior. Brain Behav Immun. 2016; 58:1–8. https://doi.org/10.1016/j.bbi.2016.01.017 [PubMed]
- 70. Green HF, Nolan YM. Inflammation and the developing brain: consequences for hippocampal neurogenesis and behavior. Neurosci Biobehav Rev. 2014; 40:20–34. https://doi.org/10.1016/j.neubiorev.2014.01.004 [PubMed]
- 71. Borsini A, Nicolaou A, Camacho-Muñoz D, Kendall AC, Di Benedetto MG, Giacobbe J, Su KP, Pariante CM. Omega-3 polyunsaturated fatty acids protect against inflammation through production of LOX and CYP450 lipid mediators: relevance for major depression and for human hippocampal neurogenesis. Mol Psychiatry. 2021; 26:6773–88. https://doi.org/10.1038/s41380-021-01160-8 [PubMed]
- 72. Qin X, Wang W, Wu H, Liu D, Wang R, Xu J, Jiang H, Pan F. PPARγ-mediated microglial activation phenotype is involved in depressive-like behaviors and neuroinflammation in stressed C57BL/6J and ob/ob mice. Psychoneuroendocrinology. 2020; 117:104674. https://doi.org/10.1016/j.psyneuen.2020.104674 [PubMed]
- 73. Bassett B, Subramaniyam S, Fan Y, Varney S, Pan H, Carneiro AM, Chung CY. Minocycline alleviates depression-like symptoms by rescuing decrease in neurogenesis in dorsal hippocampus via blocking microglia activation/phagocytosis. Brain Behav Immun. 2021; 91:519–30. https://doi.org/10.1016/j.bbi.2020.11.009 [PubMed]
- 74. Chu K, Cadar D, Iob E, Frank P. Excess body weight and specific types of depressive symptoms: Is there a mediating role of systemic low-grade inflammation? Brain Behav Immun. 2023; 108:233–44. https://doi.org/10.1016/j.bbi.2022.11.016 [PubMed]
- 75. Fava F, Rizzetto L, Tuohy KM. Gut microbiota and health: connecting actors across the metabolic system. Proc Nutr Soc. 2019; 78:177–88. https://doi.org/10.1017/S0029665118002719 [PubMed]
- 76. Guarner F, Malagelada JR. Gut flora in health and disease. Lancet. 2003; 361:512–9. https://doi.org/10.1016/S0140-6736(03)12489-0 [PubMed]
- 77. Fan Y, Pedersen O. Gut microbiota in human metabolic health and disease. Nat Rev Microbiol. 2021; 19:55–71. https://doi.org/10.1038/s41579-020-0433-9 [PubMed]
- 78. Agirman G, Yu KB, Hsiao EY. Signaling inflammation across the gut-brain axis. Science. 2021; 374:1087–92. https://doi.org/10.1126/science.abi6087 [PubMed]
- 79. Foster JA, McVey Neufeld KA. Gut-brain axis: how the microbiome influences anxiety and depression. Trends Neurosci. 2013; 36:305–12. https://doi.org/10.1016/j.tins.2013.01.005 [PubMed]
- 80. Jiang H, Ling Z, Zhang Y, Mao H, Ma Z, Yin Y, Wang W, Tang W, Tan Z, Shi J, Li L, Ruan B. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav Immun. 2015; 48:186–94. https://doi.org/10.1016/j.bbi.2015.03.016 [PubMed]
- 81. Park AJ, Collins J, Blennerhassett PA, Ghia JE, Verdu EF, Bercik P, Collins SM. Altered colonic function and microbiota profile in a mouse model of chronic depression. Neurogastroenterol Motil. 2013; 25:733–e575. https://doi.org/10.1111/nmo.12153 [PubMed]
- 82. Nikolova VL, Smith MRB, Hall LJ, Cleare AJ, Stone JM, Young AH. Perturbations in Gut Microbiota Composition in Psychiatric Disorders: A Review and Meta-analysis. JAMA Psychiatry. 2021; 78:1343–54. https://doi.org/10.1001/jamapsychiatry.2021.2573 [PubMed]
- 83. Fawcett KA, Barroso I. The genetics of obesity: FTO leads the way. Trends Genet. 2010; 26:266–74. https://doi.org/10.1016/j.tig.2010.02.006 [PubMed]
- 84. Jia G, Fu Y, Zhao X, Dai Q, Zheng G, Yang Y, Yi C, Lindahl T, Pan T, Yang YG, He C. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat Chem Biol. 2011; 7:885–7. https://doi.org/10.1038/nchembio.687 [PubMed]
- 85. Peng S, Xiao W, Ju D, Sun B, Hou N, Liu Q, Wang Y, Zhao H, Gao C, Zhang S, Cao R, Li P, Huang H, et al. Identification of entacapone as a chemical inhibitor of FTO mediating metabolic regulation through FOXO1. Sci Transl Med. 2019; 11:eaau7116. https://doi.org/10.1126/scitranslmed.aau7116 [PubMed]
- 86. Liu S, Xiu J, Zhu C, Meng K, Li C, Han R, Du T, Li L, Xu L, Liu R, Zhu W, Shen Y, Xu Q. Fat mass and obesity-associated protein regulates RNA methylation associated with depression-like behavior in mice. Nat Commun. 2021; 12:6937. https://doi.org/10.1038/s41467-021-27044-7 [PubMed]
- 87. Zarza-Rebollo JA, Molina E, Rivera M. The role of the FTO gene in the relationship between depression and obesity. A systematic review. Neurosci Biobehav Rev. 2021; 127:630–7. https://doi.org/10.1016/j.neubiorev.2021.05.013 [PubMed]
- 88. Hodes GE, Kana V, Menard C, Merad M, Russo SJ. Neuroimmune mechanisms of depression. Nat Neurosci. 2015; 18:1386–93. https://doi.org/10.1038/nn.4113 [PubMed]
- 89. Dean J, Keshavan M. The neurobiology of depression: An integrated view. Asian J Psychiatr. 2017; 27:101–11. https://doi.org/10.1016/j.ajp.2017.01.025 [PubMed]
- 90. Zhu F, Du B, Xu B. Anti-inflammatory effects of phytochemicals from fruits, vegetables, and food legumes: A review. Crit Rev Food Sci Nutr. 2018; 58:1260–70. https://doi.org/10.1080/10408398.2016.1251390 [PubMed]
- 91. Huang Q, Liu H, Suzuki K, Ma S, Liu C. Linking What We Eat to Our Mood: A Review of Diet, Dietary Antioxidants, and Depression. Antioxidants (Basel). 2019; 8:376. https://doi.org/10.3390/antiox8090376 [PubMed]
- 92. Bataglion GA, da Silva FMA, Eberlin MN, Koolen HHF. Determination of the phenolic composition from Brazilian tropical fruits by UHPLC-MS/MS. Food Chem. 2015; 180:280–7. https://doi.org/10.1016/j.foodchem.2015.02.059 [PubMed]
- 93. Anhê FF, Nachbar RT, Varin TV, Trottier J, Dudonné S, Le Barz M, Feutry P, Pilon G, Barbier O, Desjardins Y, Roy D, Marette A. Treatment with camu camu (Myrciaria dubia) prevents obesity by altering the gut microbiota and increasing energy expenditure in diet-induced obese mice. Gut. 2019; 68:453–64. https://doi.org/10.1136/gutjnl-2017-315565 [PubMed]
- 94. Spencer SJ, Korosi A, Layé S, Shukitt-Hale B, Barrientos RM. Food for thought: how nutrition impacts cognition and emotion. NPJ Sci Food. 2017; 1:7. https://doi.org/10.1038/s41538-017-0008-y [PubMed]
- 95. Chevalier G, Siopi E, Guenin-Macé L, Pascal M, Laval T, Rifflet A, Boneca IG, Demangel C, Colsch B, Pruvost A, Chu-Van E, Messager A, Leulier F, et al. Effect of gut microbiota on depressive-like behaviors in mice is mediated by the endocannabinoid system. Nat Commun. 2020; 11:6363. https://doi.org/10.1038/s41467-020-19931-2 [PubMed]
- 96. Rueda D, Navarro B, Martinez-Serrano A, Guzman M, Galve-Roperh I. The endocannabinoid anandamide inhibits neuronal progenitor cell differentiation through attenuation of the Rap1/B-Raf/ERK pathway. J Biol Chem. 2002; 277:46645–50. https://doi.org/10.1074/jbc.M206590200 [PubMed]
- 97. Aguado T, Monory K, Palazuelos J, Stella N, Cravatt B, Lutz B, Marsicano G, Kokaia Z, Guzmán M, Galve-Roperh I. The endocannabinoid system drives neural progenitor proliferation. FASEB J. 2005; 19:1704–6. https://doi.org/10.1096/fj.05-3995fje [PubMed]
- 98. Prenderville JA, Kelly ÁM, Downer EJ. The role of cannabinoids in adult neurogenesis. Br J Pharmacol. 2015; 172:3950–63. https://doi.org/10.1111/bph.13186 [PubMed]
- 99. Ren M, Zhang H, Qi J, Hu A, Jiang Q, Hou Y, Feng Q, Ojo O, Wang X. An Almond-Based Low Carbohydrate Diet Improves Depression and Glycometabolism in Patients with Type 2 Diabetes through Modulating Gut Microbiota and GLP-1: A Randomized Controlled Trial. Nutrients. 2020; 12:3036. https://doi.org/10.3390/nu12103036 [PubMed]
- 100. Asadi A, Shadab Mehr N, Mohamadi MH, Shokri F, Heidary M, Sadeghifard N, Khoshnood S. Obesity and gut-microbiota-brain axis: A narrative review. J Clin Lab Anal. 2022; 36:e24420. https://doi.org/10.1002/jcla.24420 [PubMed]
- 101. Farzi A, Hassan AM, Zenz G, Holzer P. Diabesity and mood disorders: Multiple links through the microbiota-gut-brain axis. Mol Aspects Med. 2019; 66:80–93. https://doi.org/10.1016/j.mam.2018.11.003 [PubMed]
- 102. Niccolai E, Boem F, Russo E, Amedei A. The Gut-Brain Axis in the Neuropsychological Disease Model of Obesity: A Classical Movie Revised by the Emerging Director “Microbiome”. Nutrients. 2019; 11:156. https://doi.org/10.3390/nu11010156 [PubMed]
- 103. Mayer EA. Gut feelings: the emerging biology of gut-brain communication. Nat Rev Neurosci. 2011; 12:453–66. https://doi.org/10.1038/nrn3071 [PubMed]
- 104. Cryan JF, Dinan TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci. 2012; 13:701–12. https://doi.org/10.1038/nrn3346 [PubMed]
- 105. Margolis KG, Cryan JF, Mayer EA. The Microbiota-Gut-Brain Axis: From Motility to Mood. Gastroenterology. 2021; 160:1486–501. https://doi.org/10.1053/j.gastro.2020.10.066 [PubMed]
- 106. Boem F, Amedei A. Healthy axis: Towards an integrated view of the gut-brain health. World J Gastroenterol. 2019; 25:3838–41. https://doi.org/10.3748/wjg.v25.i29.3838 [PubMed]
- 107. Dahmen B, Puetz VB, Scharke W, von Polier GG, Herpertz-Dahlmann B, Konrad K. Effects of Early-Life Adversity on Hippocampal Structures and Associated HPA Axis Functions. Dev Neurosci. 2018; 40:13–22. https://doi.org/10.1159/000484238 [PubMed]
- 108. Leuner B, Gould E. Structural plasticity and hippocampal function. Annu Rev Psychol. 2010; 61:111–40. https://doi.org/10.1146/annurev.psych.093008.100359 [PubMed]
- 109. Johnston MV. Plasticity in the developing brain: implications for rehabilitation. Dev Disabil Res Rev. 2009; 15:94–101. https://doi.org/10.1002/ddrr.64 [PubMed]
- 110. Cook SG, Buonarati OR, Coultrap SJ, Bayer KU. CaMKII holoenzyme mechanisms that govern the LTP versus LTD decision. Sci Adv. 2021; 7:eabe2300. https://doi.org/10.1126/sciadv.abe2300 [PubMed]
- 111. Hwang LL, Wang CH, Li TL, Chang SD, Lin LC, Chen CP, Chen CT, Liang KC, Ho IK, Yang WS, Chiou LC. Sex differences in high-fat diet-induced obesity, metabolic alterations and learning, and synaptic plasticity deficits in mice. Obesity (Silver Spring). 2010; 18:463–9. https://doi.org/10.1038/oby.2009.273 [PubMed]
- 112. Price RB, Duman R. Neuroplasticity in cognitive and psychological mechanisms of depression: an integrative model. Mol Psychiatry. 2020; 25:530–43. https://doi.org/10.1038/s41380-019-0615-x [PubMed]
- 113. Aleksandrova LR, Phillips AG. Neuroplasticity as a convergent mechanism of ketamine and classical psychedelics. Trends Pharmacol Sci. 2021; 42:929–42. https://doi.org/10.1016/j.tips.2021.08.003 [PubMed]
- 114. Castrén E, Kojima M. Brain-derived neurotrophic factor in mood disorders and antidepressant treatments. Neurobiol Dis. 2017; 97:119–26. https://doi.org/10.1016/j.nbd.2016.07.010 [PubMed]
- 115. Zanos P, Gould TD. Mechanisms of ketamine action as an antidepressant. Mol Psychiatry. 2018; 23:801–11. https://doi.org/10.1038/mp.2017.255 [PubMed]
- 116. Rantamäki T, Hendolin P, Kankaanpää A, Mijatovic J, Piepponen P, Domenici E, Chao MV, Männistö PT, Castrén E. Pharmacologically diverse antidepressants rapidly activate brain-derived neurotrophic factor receptor TrkB and induce phospholipase-Cgamma signaling pathways in mouse brain. Neuropsychopharmacology. 2007; 32:2152–62. https://doi.org/10.1038/sj.npp.1301345 [PubMed]
- 117. Yoshii A, Constantine-Paton M. Postsynaptic BDNF-TrkB signaling in synapse maturation, plasticity, and disease. Dev Neurobiol. 2010; 70:304–22. https://doi.org/10.1002/dneu.20765 [PubMed]
- 118. Rizzo MR, Di Meo I, Polito R, Auriemma MC, Gambardella A, di Mauro G, Capuano A, Paolisso G. Cognitive impairment and type 2 diabetes mellitus: Focus of SGLT2 inhibitors treatment. Pharmacol Res. 2022; 176:106062. https://doi.org/10.1016/j.phrs.2022.106062 [PubMed]
- 119. Jacques A, Chaaya N, Beecher K, Ali SA, Belmer A, Bartlett S. The impact of sugar consumption on stress driven, emotional and addictive behaviors. Neurosci Biobehav Rev. 2019; 103:178–99. https://doi.org/10.1016/j.neubiorev.2019.05.021 [PubMed]
- 120. Li Y, Cheng Y, Zhou Y, Du H, Zhang C, Zhao Z, Chen Y, Zhou Z, Mei J, Wu W, Chen M. High fat diet-induced obesity leads to depressive and anxiety-like behaviors in mice via AMPK/mTOR-mediated autophagy. Exp Neurol. 2022; 348:113949. https://doi.org/10.1016/j.expneurol.2021.113949 [PubMed]
- 121. Juruena MF, Cleare AJ, Pariante CM. O eixo hipotálamo-pituitária-adrenal, a função dos receptores de glicocorticóides e sua importância na depressão [The hypothalamic pituitary adrenal axis, glucocorticoid receptor function and relevance to depression]. Braz J Psychiatry. 2004; 26:189–201. Portuguese. https://doi.org/10.1590/s1516-44462004000300009 [PubMed] Erratum in: Rev Bras Psiquiatr. 2004; 26:2 p following 300.
- 122. Kunugi H, Hori H, Adachi N, Numakawa T. Interface between hypothalamic-pituitary-adrenal axis and brain-derived neurotrophic factor in depression. Psychiatry Clin Neurosci. 2010; 64:447–59. https://doi.org/10.1111/j.1440-1819.2010.02135.x [PubMed]
- 123. Rivera HM, Christiansen KJ, Sullivan EL. The role of maternal obesity in the risk of neuropsychiatric disorders. Front Neurosci. 2015; 9:194. https://doi.org/10.3389/fnins.2015.00194 [PubMed]
- 124. Stomby A, Otten J, Ryberg M, Andrew R, Walker BR, Olsson T. Diet-induced weight loss alters hepatic glucocorticoid metabolism in type 2 diabetes mellitus. Eur J Endocrinol. 2020; 182:447–57. https://doi.org/10.1530/EJE-19-0901 [PubMed]
- 125. Lasselin J, Benson S, Hebebrand J, Boy K, Weskamp V, Handke A, Hasenberg T, Remy M, Föcker M, Unteroberdörster M, Brinkhoff A, Engler H, Schedlowski M. Immunological and behavioral responses to in vivo lipopolysaccharide administration in young and healthy obese and normal-weight humans. Brain Behav Immun. 2020; 88:283–93. https://doi.org/10.1016/j.bbi.2020.05.071 [PubMed]
- 126. Pruessner M, Hellhammer DH, Pruessner JC, Lupien SJ. Self-reported depressive symptoms and stress levels in healthy young men: associations with the cortisol response to awakening. Psychosom Med. 2003; 65:92–9. https://doi.org/10.1097/01.psy.0000040950.22044.10 [PubMed]
- 127. Stetler C, Miller GE. Depression and hypothalamic-pituitary-adrenal activation: a quantitative summary of four decades of research. Psychosom Med. 2011; 73:114–26. https://doi.org/10.1097/PSY.0b013e31820ad12b [PubMed]
- 128. Dallman MF, Pecoraro NC, La Fleur SE, Warne JP, Ginsberg AB, Akana SF, Laugero KC, Houshyar H, Strack AM, Bhatnagar S, Bell ME. Glucocorticoids, chronic stress, and obesity. Prog Brain Res. 2006; 153:75–105. https://doi.org/10.1016/S0079-6123(06)53004-3 [PubMed]
- 129. Varghese FP, Brown ES. The Hypothalamic-Pituitary-Adrenal Axis in Major Depressive Disorder: A Brief Primer for Primary Care Physicians. Prim Care Companion J Clin Psychiatry. 2001; 3:151–5. https://doi.org/10.4088/pcc.v03n0401 [PubMed]
- 130. Nasca C, Bigio B, Zelli D, Nicoletti F, McEwen BS. Mind the gap: glucocorticoids modulate hippocampal glutamate tone underlying individual differences in stress susceptibility. Mol Psychiatry. 2015; 20:755–63. https://doi.org/10.1038/mp.2014.96 [PubMed]
- 131. Haroon E, Miller AH. Inflammation Effects on Brain Glutamate in Depression: Mechanistic Considerations and Treatment Implications. Curr Top Behav Neurosci. 2017; 31:173–98. https://doi.org/10.1007/7854_2016_40 [PubMed]
- 132. Leonard BE. Inflammation and depression: a causal or coincidental link to the pathophysiology? Acta Neuropsychiatr. 2018; 30:1–16. https://doi.org/10.1017/neu.2016.69 [PubMed]
- 133. Rook GA, Lowry CA. The hygiene hypothesis and psychiatric disorders. Trends Immunol. 2008; 29:150–8. https://doi.org/10.1016/j.it.2008.01.002 [PubMed]
- 134. Adam TC, Epel ES. Stress, eating and the reward system. Physiol Behav. 2007; 91:449–58. https://doi.org/10.1016/j.physbeh.2007.04.011 [PubMed]
- 135. Salminen A, Kaarniranta K, Kauppinen A. Regulation of longevity by FGF21: Interaction between energy metabolism and stress responses. Ageing Res Rev. 2017; 37:79–93. https://doi.org/10.1016/j.arr.2017.05.004 [PubMed]
- 136. Itoh N. Hormone-like (endocrine) Fgfs: their evolutionary history and roles in development, metabolism, and disease. Cell Tissue Res. 2010; 342:1–11. https://doi.org/10.1007/s00441-010-1024-2 [PubMed]
- 137. Werdermann M, Berger I, Scriba LD, Santambrogio A, Schlinkert P, Brendel H, Morawietz H, Schedl A, Peitzsch M, King AJF, Andoniadou CL, Bornstein SR, Steenblock C. Insulin and obesity transform hypothalamic-pituitary-adrenal axis stemness and function in a hyperactive state. Mol Metab. 2021; 43:101112. https://doi.org/10.1016/j.molmet.2020.101112 [PubMed]
- 138. Xia G, Han Y, Meng F, He Y, Srisai D, Farias M, Dang M, Palmiter RD, Xu Y, Wu Q. Reciprocal control of obesity and anxiety-depressive disorder via a GABA and serotonin neural circuit. Mol Psychiatry. 2021; 26:2837–53. https://doi.org/10.1038/s41380-021-01053-w [PubMed]
- 139. Hallihan H, Tsai P, Lv N, Xiao L, Peñalver Bernabé B, Wu Y, Pandey GN, Williams LM, Ajilore OA, Ma J. Affective neural circuits and inflammatory markers linked to depression and anxiety symptoms in patients with comorbid obesity. J Psychiatr Res. 2023; 160:9–18. https://doi.org/10.1016/j.jpsychires.2023.01.044 [PubMed]
- 140. Kong L, Chen X, Gissler M, Lavebratt C. Relationship of prenatal maternal obesity and diabetes to offspring neurodevelopmental and psychiatric disorders: a narrative review. Int J Obes (Lond). 2020; 44:1981–2000. https://doi.org/10.1038/s41366-020-0609-4 [PubMed]
- 141. Speed MS, Jefsen OH, Børglum AD, Speed D, Østergaard SD. Investigating the association between body fat and depression via Mendelian randomization. Transl Psychiatry. 2019; 9:184. https://doi.org/10.1038/s41398-019-0516-4 [PubMed]
- 142. O’Loughlin J, Casanova F, Fairhurst-Hunter Z, Hughes A, Bowden J, Watkins ER, Freathy RM, Millwood IY, Lin K, Chen Z, Li L, Lv J, Walters RG, et al, and China Kadoorie Biobank Collaborative Group. Mendelian randomisation study of body composition and depression in people of East Asian ancestry highlights potential setting-specific causality. BMC Med. 2023; 21:37. https://doi.org/10.1186/s12916-023-02735-8 [PubMed]
- 143. Liao SF, Su CY, Su MH, Chen CY, Chen CY, Lin YF, Pan YJ, Hsiao PC, Chen PC, Huang YT, Wu CS, Wang SH. Associations of polygenic risks, depression, and obesity-related traits in Taiwan Biobank. J Affect Disord. 2023; 320:397–403. https://doi.org/10.1016/j.jad.2022.09.149 [PubMed]
- 144. Rivera M, Locke AE, Corre T, Czamara D, Wolf C, Ching-Lopez A, Milaneschi Y, Kloiber S, Cohen-Woods S, Rucker J, Aitchison KJ, Bergmann S, Boomsma DI, et al. Interaction between the FTO gene, body mass index and depression: meta-analysis of 13701 individuals. Br J Psychiatry. 2017; 211:70–6. https://doi.org/10.1192/bjp.bp.116.183475 [PubMed] Erratum in: Br J Psychiatry. 2017; 211:401. https://doi.org/10.1192/bjp.211.6.401 [PubMed]
- 145. Amare AT, Schubert KO, Tekola-Ayele F, Hsu YH, Sangkuhl K, Jenkins G, Whaley RM, Barman P, Batzler A, Altman RB, Arolt V, Brockmöller J, Chen CH, et al. The association of obesity and coronary artery disease genes with response to SSRIs treatment in major depression. J Neural Transm (Vienna). 2019; 126:35–45. https://doi.org/10.1007/s00702-018-01966-x [PubMed]
- 146. Alonso-Pedrero L, Bes-Rastrollo M, Marti A. Effects of antidepressant and antipsychotic use on weight gain: A systematic review. Obes Rev. 2019; 20:1680–90. https://doi.org/10.1111/obr.12934 [PubMed]
- 147. Gill H, Gill B, El-Halabi S, Chen-Li D, Lipsitz O, Rosenblat JD, Van Rheenen TE, Rodrigues NB, Mansur RB, Majeed A, Lui LMW, Nasri F, Lee Y, Mcintyre RS. Antidepressant Medications and Weight Change: A Narrative Review. Obesity (Silver Spring). 2020; 28:2064–72. https://doi.org/10.1002/oby.22969 [PubMed]
- 148. Géloën A, Roy PE, Bukowiecki LJ. Regression of white adipose tissue in diabetic rats. Am J Physiol. 1989; 257:E547–53. https://doi.org/10.1152/ajpendo.1989.257.4.E547 [PubMed]
- 149. Cedikova M, Kripnerová M, Dvorakova J, Pitule P, Grundmanova M, Babuska V, Mullerova D, Kuncova J. Mitochondria in White, Brown, and Beige Adipocytes. Stem Cells Int. 2016; 2016:6067349. https://doi.org/10.1155/2016/6067349 [PubMed]
- 150. Warner A, Mittag J. Brown fat and vascular heat dissipation: The new cautionary tail. Adipocyte. 2014; 3:221–3. https://doi.org/10.4161/adip.28815 [PubMed]
- 151. Han S, Sun HM, Hwang KC, Kim SW. Adipose-Derived Stromal Vascular Fraction Cells: Update on Clinical Utility and Efficacy. Crit Rev Eukaryot Gene Expr. 2015; 25:145–52. https://doi.org/10.1615/critreveukaryotgeneexpr.2015013057 [PubMed]
- 152. Wieser V, Moschen AR, Tilg H. Adipocytokines and hepatocellular carcinoma. Dig Dis. 2012; 30:508–13. https://doi.org/10.1159/000341702 [PubMed]
- 153. Koerner A, Kratzsch J, Kiess W. Adipocytokines: leptin--the classical, resistin--the controversical, adiponectin--the promising, and more to come. Best Pract Res Clin Endocrinol Metab. 2005; 19:525–46. https://doi.org/10.1016/j.beem.2005.07.008 [PubMed]
- 154. Garza JC, Guo M, Zhang W, Lu XY. Leptin increases adult hippocampal neurogenesis in vivo and in vitro. J Biol Chem. 2008; 283:18238–47. https://doi.org/10.1074/jbc.M800053200 [PubMed]
- 155. Song J, Kang SM, Kim E, Kim CH, Song HT, Lee JE. Adiponectin receptor-mediated signaling ameliorates cerebral cell damage and regulates the neurogenesis of neural stem cells at high glucose concentrations: an in vivo and in vitro study. Cell Death Dis. 2015; 6:e1844. https://doi.org/10.1038/cddis.2015.220 [PubMed]
- 156. Li VL, Kim JT, Long JZ. Adipose Tissue Lipokines: Recent Progress and Future Directions. Diabetes. 2020; 69:2541–8. https://doi.org/10.2337/dbi20-0012 [PubMed]
- 157. Cao H, Gerhold K, Mayers JR, Wiest MM, Watkins SM, Hotamisligil GS. Identification of a lipokine, a lipid hormone linking adipose tissue to systemic metabolism. Cell. 2008; 134:933–44. https://doi.org/10.1016/j.cell.2008.07.048 [PubMed]
- 158. Fantuzzi G. Adiponectin and inflammation: consensus and controversy. J Allergy Clin Immunol. 2008; 121:326–30. https://doi.org/10.1016/j.jaci.2007.10.018 [PubMed]
- 159. Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM. Positional cloning of the mouse obese gene and its human homologue. Nature. 1994; 372:425–32. https://doi.org/10.1038/372425a0 [PubMed]
- 160. Friedman JM. The function of leptin in nutrition, weight, and physiology. Nutr Rev. 2002; 60:S1–14. https://doi.org/10.1301/002966402320634878 [PubMed]
- 161. Liu J, Garza JC, Bronner J, Kim CS, Zhang W, Lu XY. Acute administration of leptin produces anxiolytic-like effects: a comparison with fluoxetine. Psychopharmacology (Berl). 2010; 207:535–45. https://doi.org/10.1007/s00213-009-1684-3 [PubMed]
- 162. Morris AA, Ahmed Y, Stoyanova N, Hooper WC, De Staerke C, Gibbons G, Quyyumi A, Vaccarino V. The association between depression and leptin is mediated by adiposity. Psychosom Med. 2012; 74:483–8. https://doi.org/10.1097/PSY.0b013e31824f5de0 [PubMed]
- 163. Zhang W, Jin Y, Wang D, Cui J. Neuroprotective effects of leptin on cerebral ischemia through JAK2/STAT3/PGC-1-mediated mitochondrial function modulation. Brain Res Bull. 2020; 156:118–30. https://doi.org/10.1016/j.brainresbull.2020.01.002 [PubMed]
- 164. Wen R, Hu S, Xiao Q, Han C, Gan C, Gou H, Liu H, Li L, Xu H, He H, Wang J. Leptin exerts proliferative and anti-apoptotic effects on goose granulosa cells through the PI3K/Akt/mTOR signaling pathway. J Steroid Biochem Mol Biol. 2015; 149:70–9. https://doi.org/10.1016/j.jsbmb.2015.01.001 [PubMed]
- 165. Wu Y, Huang XF, Bell C, Yu Y. Ginsenoside Rb1 improves leptin sensitivity in the prefrontal cortex in obese mice. CNS Neurosci Ther. 2018; 24:98–107. https://doi.org/10.1111/cns.12776 [PubMed]
- 166. Lee TH, Cheng KK, Hoo RL, Siu PM, Yau SY. The Novel Perspectives of Adipokines on Brain Health. Int J Mol Sci. 2019; 20:5638. https://doi.org/10.3390/ijms20225638 [PubMed]
- 167. Harvey J, Solovyova N, Irving A. Leptin and its role in hippocampal synaptic plasticity. Prog Lipid Res. 2006; 45:369–78. https://doi.org/10.1016/j.plipres.2006.03.001 [PubMed]
- 168. Paz-Filho G, Mastronardi CA, Licinio J. Leptin treatment: facts and expectations. Metabolism. 2015; 64:146–56. https://doi.org/10.1016/j.metabol.2014.07.014 [PubMed]
- 169. Li C, Meng F, Garza JC, Liu J, Lei Y, Kirov SA, Guo M, Lu XY. Modulation of depression-related behaviors by adiponectin AdipoR1 receptors in 5-HT neurons. Mol Psychiatry. 2021; 26:4205–20. https://doi.org/10.1038/s41380-020-0649-0 [PubMed]
- 170. Lee SH, Shin C, Ko YH, Lee MS, Park MH, Pae CU, Yoon HK, Han C. Plasminogen Activator Inhibitor-1: Potential Inflammatory Marker in Late-life Depression. Clin Psychopharmacol Neurosci. 2023; 21:147–61. https://doi.org/10.9758/cpn.2023.21.1.147 [PubMed]
- 171. Sillen M, Declerck PJ. A Narrative Review on Plasminogen Activator Inhibitor-1 and Its (Patho)Physiological Role: To Target or Not to Target? Int J Mol Sci. 2021; 22:2721. https://doi.org/10.3390/ijms22052721 [PubMed]
- 172. Macfelda K, Weiss TW, Kaun C, Breuss JM, Zorn G, Oberndorfer U, Voegele-Kadletz M, Huber-Beckmann R, Ullrich R, Binder BR, Losert UM, Maurer G, Pacher R, et al. Plasminogen activator inhibitor 1 expression is regulated by the inflammatory mediators interleukin-1alpha, tumor necrosis factor-alpha, transforming growth factor-beta and oncostatin M in human cardiac myocytes. J Mol Cell Cardiol. 2002; 34:1681–91. https://doi.org/10.1006/jmcc.2002.2117 [PubMed]
- 173. Coudriet GM, Stoops J, Orr AV, Bhushan B, Koral K, Lee S, Previte DM, Dong HH, Michalopoulos GK, Mars WM, Piganelli JD. A Noncanonical Role for Plasminogen Activator Inhibitor Type 1 in Obesity-Induced Diabetes. Am J Pathol. 2019; 189:1413–22. https://doi.org/10.1016/j.ajpath.2019.04.004 [PubMed]
- 174. Lorkiewicz P, Waszkiewicz N. Biomarkers of Post-COVID Depression. J Clin Med. 2021; 10:4142. https://doi.org/10.3390/jcm10184142 [PubMed]
- 175. Proma MA, Daria S, Nahar Z, Ashraful Islam SM, Bhuiyan MA, Islam MR. Monocyte chemoattractant protein-1 levels are associated with major depressive disorder. J Basic Clin Physiol Pharmacol. 2022; 33:735–41. https://doi.org/10.1515/jbcpp-2021-0132 [PubMed]
- 176. Zhou Z, Chen H, Ju H, Sun M. Circulating retinol binding protein 4 levels in nonalcoholic fatty liver disease: a systematic review and meta-analysis. Lipids Health Dis. 2017; 16:180. https://doi.org/10.1186/s12944-017-0566-7 [PubMed]
- 177. Sun Z, Lei H, Zhang Z. Pre-B cell colony enhancing factor (PBEF), a cytokine with multiple physiological functions. Cytokine Growth Factor Rev. 2013; 24:433–42. https://doi.org/10.1016/j.cytogfr.2013.05.006 [PubMed]
- 178. Luk T, Malam Z, Marshall JC. Pre-B cell colony-enhancing factor (PBEF)/visfatin: a novel mediator of innate immunity. J Leukoc Biol. 2008; 83:804–16. https://doi.org/10.1189/jlb.0807581 [PubMed]
- 179. Samal B, Sun Y, Stearns G, Xie C, Suggs S, McNiece I. Cloning and characterization of the cDNA encoding a novel human pre-B-cell colony-enhancing factor. Mol Cell Biol. 1994; 14:1431–7. https://doi.org/10.1128/mcb.14.2.1431-1437.1994 [PubMed]
- 180. Pilz S, Mangge H, Obermayer-Pietsch B, März W. Visfatin/pre-B-cell colony-enhancing factor: a protein with various suggested functions. J Endocrinol Invest. 2007; 30:138–44. https://doi.org/10.1007/BF03347412 [PubMed]
- 181. Jing Z, Xing J, Chen X, Stetler RA, Weng Z, Gan Y, Zhang F, Gao Y, Chen J, Leak RK, Cao G. Neuronal NAMPT is released after cerebral ischemia and protects against white matter injury. J Cereb Blood Flow Metab. 2014; 34:1613–21. https://doi.org/10.1038/jcbfm.2014.119 [PubMed]
- 182. Carvalho AF, Rocha DQC, McIntyre RS, Mesquita LM, Köhler CA, Hyphantis TN, Sales PMG, Machado-Vieira R, Berk M. Adipokines as emerging depression biomarkers: a systematic review and meta-analysis. J Psychiatr Res. 2014; 59:28–37. https://doi.org/10.1016/j.jpsychires.2014.08.002 [PubMed]
- 183. Zhou L, Li JY, He PP, Yu XH, Tang CK. Resistin: Potential biomarker and therapeutic target in atherosclerosis. Clin Chim Acta. 2021; 512:84–91. https://doi.org/10.1016/j.cca.2020.11.010 [PubMed]
- 184. Huang X, Yang Z. Resistin’s, obesity and insulin resistance: the continuing disconnect between rodents and humans. J Endocrinol Invest. 2016; 39:607–15. https://doi.org/10.1007/s40618-015-0408-2 [PubMed]
- 185. Yang F, Bai Y, Jiang Y. Effects of Apelin on RAW264.7 cells under both normal and hypoxic conditions. Peptides. 2015; 69:133–43. https://doi.org/10.1016/j.peptides.2015.04.025 [PubMed]
- 186. Liu Q, Jiang J, Shi Y, Mo Z, Li M. Apelin/Apelin receptor: A new therapeutic target in Polycystic Ovary Syndrome. Life Sci. 2020; 260:118310. https://doi.org/10.1016/j.lfs.2020.118310 [PubMed]
- 187. Tian SW, Xu F, Gui SJ. Apelin-13 reverses memory impairment and depression-like behavior in chronic social defeat stressed rats. Peptides. 2018; 108:1–6. https://doi.org/10.1016/j.peptides.2018.08.009 [PubMed]
- 188. Dai TT, Wang B, Xiao ZY, You Y, Tian SW. Apelin-13 Upregulates BDNF Against Chronic Stress-induced Depression-like Phenotypes by Ameliorating HPA Axis and Hippocampal Glucocorticoid Receptor Dysfunctions. Neuroscience. 2018; 390:151–9. https://doi.org/10.1016/j.neuroscience.2018.08.018 [PubMed]
- 189. Buechler C, Feder S, Haberl EM, Aslanidis C. Chemerin Isoforms and Activity in Obesity. Int J Mol Sci. 2019; 20:1128. https://doi.org/10.3390/ijms20051128 [PubMed]
- 190. Zhang Y, Xu N, Ding Y, Doycheva DM, Zhang Y, Li Q, Flores J, Haghighiabyaneh M, Tang J, Zhang JH. Chemerin reverses neurological impairments and ameliorates neuronal apoptosis through ChemR23/CAMKK2/AMPK pathway in neonatal hypoxic-ischemic encephalopathy. Cell Death Dis. 2019; 10:97. https://doi.org/10.1038/s41419-019-1374-y [PubMed]
- 191. Deyama S, Shimoda K, Suzuki H, Ishikawa Y, Ishimura K, Fukuda H, Hitora-Imamura N, Ide S, Satoh M, Kaneda K, Shuto S, Minami M. Resolvin E1/E2 ameliorate lipopolysaccharide-induced depression-like behaviors via ChemR23. Psychopharmacology (Berl). 2018; 235:329–36. https://doi.org/10.1007/s00213-017-4774-7 [PubMed]
- 192. Prentice KJ, Saksi J, Hotamisligil GS. Adipokine FABP4 integrates energy stores and counterregulatory metabolic responses. J Lipid Res. 2019; 60:734–40. https://doi.org/10.1194/jlr.S091793 [PubMed]
- 193. Pinacho R, Villalmanzo N, Meana JJ, Ferrer I, Berengueras A, Haro JM, Villén J, Ramos B. Altered CSNK1E, FABP4 and NEFH protein levels in the dorsolateral prefrontal cortex in schizophrenia. Schizophr Res. 2016; 177:88–97. https://doi.org/10.1016/j.schres.2016.04.050 [PubMed]
- 194. Tanaka T, Narazaki M, Kishimoto T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb Perspect Biol. 2014; 6:a016295. https://doi.org/10.1101/cshperspect.a016295 [PubMed]
- 195. Wueest S, Konrad D. The controversial role of IL-6 in adipose tissue on obesity-induced dysregulation of glucose metabolism. Am J Physiol Endocrinol Metab. 2020; 319:E607–13. https://doi.org/10.1152/ajpendo.00306.2020 [PubMed]
- 196. Roohi E, Jaafari N, Hashemian F. On inflammatory hypothesis of depression: what is the role of IL-6 in the middle of the chaos? J Neuroinflammation. 2021; 18:45. https://doi.org/10.1186/s12974-021-02100-7 [PubMed]
- 197. Paramo B, Wyatt S, Davies AM. An essential role for neuregulin-4 in the growth and elaboration of developing neocortical pyramidal dendrites. Exp Neurol. 2018; 302:85–92. https://doi.org/10.1016/j.expneurol.2018.01.002 [PubMed]
- 198. Tutunchi H, Ostadrahimi A, Hosseinzadeh-Attar MJ, Miryan M, Mobasseri M, Ebrahimi-Mameghani M. A systematic review of the association of neuregulin 4, a brown fat-enriched secreted factor, with obesity and related metabolic disturbances. Obes Rev. 2020; 21:e12952. https://doi.org/10.1111/obr.12952 [PubMed]
- 199. Escoté X, Gómez-Zorita S, López-Yoldi M, Milton-Laskibar I, Fernández-Quintela A, Martínez JA, Moreno-Aliaga MJ, Portillo MP. Role of Omentin, Vaspin, Cardiotrophin-1, TWEAK and NOV/CCN3 in Obesity and Diabetes Development. Int J Mol Sci. 2017; 18:1770. https://doi.org/10.3390/ijms18081770 [PubMed]
- 200. Zhao LR, Du YJ, Chen L, Liu ZG, Jia XY, Pan YH, Liu JF, Liu B. Omentin-1 promotes the growth of neural stem cells via activation of Akt signaling. Mol Med Rep. 2015; 11:1859–64. https://doi.org/10.3892/mmr.2014.2937 [PubMed]
- 201. Liu L, Chan M, Yu L, Wang W, Qiang L. Adipsin deficiency does not impact atherosclerosis development in Ldlr-/- mice. Am J Physiol Endocrinol Metab. 2021; 320:E87–92. https://doi.org/10.1152/ajpendo.00440.2020 [PubMed]
- 202. Bond DJ, Andreazza AC, Hughes J, Dhanoa T, Torres IJ, Kozicky JM, Young LT, Lam RW, Yatham LN. A Longitudinal Study of the Relationships Between Mood Symptoms, Body Mass Index, and Serum Adipokines in Bipolar Disorder. J Clin Psychiatry. 2017; 78:441–8. https://doi.org/10.4088/JCP.15m10189 [PubMed]
- 203. Blüher M. Vaspin in obesity and diabetes: pathophysiological and clinical significance. Endocrine. 2012; 41:176–82. https://doi.org/10.1007/s12020-011-9572-0 [PubMed]
- 204. Heiker JT. Vaspin (serpinA12) in obesity, insulin resistance, and inflammation. J Pept Sci. 2014; 20:299–306. https://doi.org/10.1002/psc.2621 [PubMed]
- 205. Nunes EA, Rafacho A. Implications of Palmitoleic Acid (Palmitoleate) On Glucose Homeostasis, Insulin Resistance and Diabetes. Curr Drug Targets. 2017; 18:619–28. https://doi.org/10.2174/1389450117666151209120345 [PubMed]
- 206. de Souza CO, Vannice GK, Rosa Neto JC, Calder PC. Is Palmitoleic Acid a Plausible Nonpharmacological Strategy to Prevent or Control Chronic Metabolic and Inflammatory Disorders? Mol Nutr Food Res. 2018; 62. https://doi.org/10.1002/mnfr.201700504 [PubMed]
- 207. Yu H, Qin X, Yu Z, Chen Y, Tang L, Shan W. Effects of high-fat diet on the formation of depressive-like behavior in mice. Food Funct. 2021; 12:6416–31. https://doi.org/10.1039/d1fo00044f [PubMed]
- 208. Ameer F, Scandiuzzi L, Hasnain S, Kalbacher H, Zaidi N. De novo lipogenesis in health and disease. Metabolism. 2014; 63:895–902. https://doi.org/10.1016/j.metabol.2014.04.003 [PubMed]
- 209. Macêdo APA, Muñoz VR, Cintra DE, Pauli JR. 12,13-diHOME as a new therapeutic target for metabolic diseases. Life Sci. 2022; 290:120229. https://doi.org/10.1016/j.lfs.2021.120229 [PubMed]
- 210. D’Souza K, Paramel GV, Kienesberger PC. Lysophosphatidic Acid Signaling in Obesity and Insulin Resistance. Nutrients. 2018; 10:399. https://doi.org/10.3390/nu10040399 [PubMed]
- 211. Nagata W, Koizumi A, Nakagawa K, Takahashi S, Gotoh M, Satoh Y, Ishizuka T. Treatment with lysophosphatidic acid prevents microglial activation and depression-like behaviours in a murine model of neuropsychiatric systemic lupus erythematosus. Clin Exp Immunol. 2023; 212:81–92. https://doi.org/10.1093/cei/uxad010 [PubMed]
- 212. Tartaglia LA, Dembski M, Weng X, Deng N, Culpepper J, Devos R, Richards GJ, Campfield LA, Clark FT, Deeds J, Muir C, Sanker S, Moriarty A, et al. Identification and expression cloning of a leptin receptor, OB-R. Cell. 1995; 83:1263–71. https://doi.org/10.1016/0092-8674(95)90151-5 [PubMed]
- 213. Shanley LJ, Irving AJ, Harvey J. Leptin enhances NMDA receptor function and modulates hippocampal synaptic plasticity. J Neurosci. 2001; 21:RC186. https://doi.org/10.1523/JNEUROSCI.21-24-j0001.2001 [PubMed]
- 214. Straub LG, Scherer PE. Metabolic Messengers: Adiponectin. Nat Metab. 2019; 1:334–9. https://doi.org/10.1038/s42255-019-0041-z [PubMed]
- 215. Yamauchi T, Kamon J, Ito Y, Tsuchida A, Yokomizo T, Kita S, Sugiyama T, Miyagishi M, Hara K, Tsunoda M, Murakami K, Ohteki T, Uchida S, et al. Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. Nature. 2003; 423:762–9. https://doi.org/10.1038/nature01705 [PubMed]
- 216. Sun J, Gao Y, Yao T, Huang Y, He Z, Kong X, Yu KJ, Wang RT, Guo H, Yan J, Chang Y, Chen H, Scherer PE, et al. Adiponectin potentiates the acute effects of leptin in arcuate Pomc neurons. Mol Metab. 2016; 5:882–91. https://doi.org/10.1016/j.molmet.2016.08.007 [PubMed]
- 217. Liu J, Guo M, Zhang D, Cheng SY, Liu M, Ding J, Scherer PE, Liu F, Lu XY. Adiponectin is critical in determining susceptibility to depressive behaviors and has antidepressant-like activity. Proc Natl Acad Sci USA. 2012; 109:12248–53. https://doi.org/10.1073/pnas.1202835109 [PubMed]
- 218. Várhelyi ZP, Kálmán J, Oláh Z, Ivitz EV, Fodor EK, Sántha M, Datki ZL, Pákáski M. Adiponectin Receptors Are Less Sensitive to Stress in a Transgenic Mouse Model of Alzheimer’s Disease. Front Neurosci. 2017; 11:199. https://doi.org/10.3389/fnins.2017.00199 [PubMed]
- 219. Sun F, Lei Y, You J, Li C, Sun L, Garza J, Zhang D, Guo M, Scherer PE, Lodge D, Lu XY. Adiponectin modulates ventral tegmental area dopamine neuron activity and anxiety-related behavior through AdipoR1. Mol Psychiatry. 2019; 24:126–44. https://doi.org/10.1038/s41380-018-0102-9 [PubMed]
- 220. You J, Sun L, Wang J, Sun F, Wang W, Wang D, Fan X, Liu D, Xu Z, Qiu C, Chen J, Yan H, Liu B. Role of Adiponectin-Notch pathway in cognitive dysfunction associated with depression and in the therapeutic effect of physical exercise. Aging Cell. 2021; 20:e13387. https://doi.org/10.1111/acel.13387 [PubMed]
- 221. Zhang D, Guo M, Zhang W, Lu XY. Adiponectin stimulates proliferation of adult hippocampal neural stem/progenitor cells through activation of p38 mitogen-activated protein kinase (p38MAPK)/glycogen synthase kinase 3β (GSK-3β)/β-catenin signaling cascade. J Biol Chem. 2011; 286:44913–20. https://doi.org/10.1074/jbc.M111.310052 [PubMed]
- 222. Beurel E, Song L, Jope RS. Inhibition of glycogen synthase kinase-3 is necessary for the rapid antidepressant effect of ketamine in mice. Mol Psychiatry. 2011; 16:1068–70. https://doi.org/10.1038/mp.2011.47 [PubMed]
- 223. Coope A, Milanski M, Araújo EP, Tambascia M, Saad MJA, Geloneze B, Velloso LA. AdipoR1 mediates the anorexigenic and insulin/leptin-like actions of adiponectin in the hypothalamus. FEBS Lett. 2008; 582:1471–6. https://doi.org/10.1016/j.febslet.2008.03.037 [PubMed]
- 224. Huypens P, Moens K, Heimberg H, Ling Z, Pipeleers D, Van de Casteele M. Adiponectin-mediated stimulation of AMP-activated protein kinase (AMPK) in pancreatic beta cells. Life Sci. 2005; 77:1273–82. https://doi.org/10.1016/j.lfs.2005.03.008 [PubMed]
- 225. Guo C, Ricchiuti V, Lian BQ, Yao TM, Coutinho P, Romero JR, Li J, Williams GH, Adler GK. Mineralocorticoid receptor blockade reverses obesity-related changes in expression of adiponectin, peroxisome proliferator-activated receptor-gamma, and proinflammatory adipokines. Circulation. 2008; 117:2253–61. https://doi.org/10.1161/CIRCULATIONAHA.107.748640 [PubMed]
- 226. Arita Y, Kihara S, Ouchi N, Takahashi M, Maeda K, Miyagawa J, Hotta K, Shimomura I, Nakamura T, Miyaoka K, Kuriyama H, Nishida M, Yamashita S, et al. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem Biophys Res Commun. 1999; 257:79–83. https://doi.org/10.1006/bbrc.1999.0255 [PubMed]
- 227. Guenther M, James R, Marks J, Zhao S, Szabo A, Kidambi S. Adiposity distribution influences circulating adiponectin levels. Transl Res. 2014; 164:270–7. https://doi.org/10.1016/j.trsl.2014.04.008 [PubMed]
- 228. Alberti KGMM, Eckel RH, Grundy SM, Zimmet PZ, Cleeman JI, Donato KA, Fruchart JC, James WPT, Loria CM, Smith SC
Jr , and International Diabetes Federation Task Force on Epidemiology and Prevention, and Hational Heart, Lung, and Blood Institute, and American Heart Association, and World Heart Federation, and International Atherosclerosis Society, and International Association for the Study of Obesity. Harmonizing the metabolic syndrome: a joint interim statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation. 2009; 120:1640–5. https://doi.org/10.1161/CIRCULATIONAHA.109.192644 [PubMed] - 229. Klawitter J, Bek S, Zakaria M, Zeng C, Hornberger A, Gilbert R, Shokati T, Klawitter J, Christians U, Boernsen KO. Fatty acid desaturation index in human plasma: comparison of different analytical methodologies for the evaluation of diet effects. Anal Bioanal Chem. 2014; 406:6399–408. https://doi.org/10.1007/s00216-014-8020-4 [PubMed]
- 230. Ntambi JM, Miyazaki M. Regulation of stearoyl-CoA desaturases and role in metabolism. Prog Lipid Res. 2004; 43:91–104. https://doi.org/10.1016/s0163-7827(03)00039-0 [PubMed]
- 231. Bermúdez MA, Pereira L, Fraile C, Valerio L, Balboa MA, Balsinde J. Roles of Palmitoleic Acid and Its Positional Isomers, Hypogeic and Sapienic Acids, in Inflammation, Metabolic Diseases and Cancer. Cells. 2022; 11:2146. https://doi.org/10.3390/cells11142146 [PubMed]
- 232. Carta G, Murru E, Banni S, Manca C. Palmitic Acid: Physiological Role, Metabolism and Nutritional Implications. Front Physiol. 2017; 8:902. https://doi.org/10.3389/fphys.2017.00902 [PubMed]
- 233. Bolsoni-Lopes A, Festuccia WT, Chimin P, Farias TSM, Torres-Leal FL, Cruz MM, Andrade PB, Hirabara SM, Lima FB, Alonso-Vale MIC. Palmitoleic acid (n-7) increases white adipocytes GLUT4 content and glucose uptake in association with AMPK activation. Lipids Health Dis. 2014; 13:199. https://doi.org/10.1186/1476-511X-13-199 [PubMed]
- 234. Cruz MM, Simão JJ, de Sá RDCC, Farias TSM, da Silva VS, Abdala F, Antraco VJ, Armelin-Correa L, Alonso-Vale MIC. Palmitoleic Acid Decreases Non-alcoholic Hepatic Steatosis and Increases Lipogenesis and Fatty Acid Oxidation in Adipose Tissue From Obese Mice. Front Endocrinol (Lausanne). 2020; 11:537061. https://doi.org/10.3389/fendo.2020.537061 [PubMed]
- 235. Birgbauer E. Lysophosphatidic Acid Signalling in Nervous System Development and Function. Neuromolecular Med. 2021; 23:68–85. https://doi.org/10.1007/s12017-020-08630-2 [PubMed]
- 236. Wunsch E, Krawczyk M, Milkiewicz M, Trottier J, Barbier O, Neurath MF, Lammert F, Kremer AE, Milkiewicz P. Serum Autotaxin is a Marker of the Severity of Liver Injury and Overall Survival in Patients with Cholestatic Liver Diseases. Sci Rep. 2016; 6:30847. https://doi.org/10.1038/srep30847 [PubMed]
- 237. Kaffe E, Katsifa A, Xylourgidis N, Ninou I, Zannikou M, Harokopos V, Foka P, Dimitriadis A, Evangelou K, Moulas AN, Georgopoulou U, Gorgoulis VG, Dalekos GN, Aidinis V. Hepatocyte autotaxin expression promotes liver fibrosis and cancer. Hepatology. 2017; 65:1369–83. https://doi.org/10.1002/hep.28973 [PubMed]
- 238. Rancoule C, Attané C, Grès S, Fournel A, Dusaulcy R, Bertrand C, Vinel C, Tréguer K, Prentki M, Valet P, Saulnier-Blache JS. Lysophosphatidic acid impairs glucose homeostasis and inhibits insulin secretion in high-fat diet obese mice. Diabetologia. 2013; 56:1394–402. https://doi.org/10.1007/s00125-013-2891-3 [PubMed]
- 239. García-Morales V, Montero F, González-Forero D, Rodríguez-Bey G, Gómez-Pérez L, Medialdea-Wandossell MJ, Domínguez-Vías G, García-Verdugo JM, Moreno-López B. Membrane-derived phospholipids control synaptic neurotransmission and plasticity. PLoS Biol. 2015; 13:e1002153. https://doi.org/10.1371/journal.pbio.1002153 [PubMed]
- 240. Kihara Y, Maceyka M, Spiegel S, Chun J. Lysophospholipid receptor nomenclature review: IUPHAR Review 8. Br J Pharmacol. 2014; 171:3575–94. https://doi.org/10.1111/bph.12678 [PubMed]
- 241. Roza C, Campos-Sandoval JA, Gómez-García MC, Peñalver A, Márquez J. Lysophosphatidic Acid and Glutamatergic Transmission. Front Mol Neurosci. 2019; 12:138. https://doi.org/10.3389/fnmol.2019.00138 [PubMed]
- 242. Moreno-Fernández RD, Pérez-Martín M, Castilla-Ortega E, Rosell Del Valle C, García-Fernández MI, Chun J, Estivill-Torrús G, Rodríguez de Fonseca F, Santín LJ, Pedraza C. maLPA1-null mice as an endophenotype of anxious depression. Transl Psychiatry. 2017; 7:e1077. https://doi.org/10.1038/tp.2017.24 [PubMed]
- 243. Peñalver A, Campos-Sandoval JA, Blanco E, Cardona C, Castilla L, Martín-Rufián M, Estivill-Torrús G, Sánchez-Varo R, Alonso FJ, Pérez-Hernández M, Colado MI, Gutiérrez A, de Fonseca FR, Márquez J. Glutaminase and MMP-9 Downregulation in Cortex and Hippocampus of LPA1 Receptor Null Mice Correlate with Altered Dendritic Spine Plasticity. Front Mol Neurosci. 2017; 10:278. https://doi.org/10.3389/fnmol.2017.00278 [PubMed]
- 244. Lin YN, Audira G, Malhotra N, Ngoc Anh NT, Siregar P, Lu JH, Lee H, Hsiao CD. A Novel Function of the Lysophosphatidic Acid Receptor 3 (LPAR3) Gene in Zebrafish on Modulating Anxiety, Circadian Rhythm Locomotor Activity, and Short-Term Memory. Int J Mol Sci. 2020; 21:2837. https://doi.org/10.3390/ijms21082837 [PubMed]
- 245. Kim HJ, Park SD, Lee RM, Lee BH, Choi SH, Hwang SH, Rhim H, Kim HC, Nah SY. Gintonin attenuates depressive-like behaviors associated with alcohol withdrawal in mice. J Affect Disord. 2017; 215:23–9. https://doi.org/10.1016/j.jad.2017.03.026 [PubMed]
- 246. Kühnen P, Wabitsch M, von Schnurbein J, Chirila C, Mallya UG, Callahan P, Gnanasakthy A, Poitou C, Krabusch PM, Stewart M, Clément K. Quality of life outcomes in two phase 3 trials of setmelanotide in patients with obesity due to LEPR or POMC deficiency. Orphanet J Rare Dis. 2022; 17:38. https://doi.org/10.1186/s13023-022-02186-z [PubMed]
- 247. Bremner JD, Moazzami K, Wittbrodt MT, Nye JA, Lima BB, Gillespie CF, Rapaport MH, Pearce BD, Shah AJ, Vaccarino V. Diet, Stress and Mental Health. Nutrients. 2020; 12:2428. https://doi.org/10.3390/nu12082428 [PubMed]
- 248. Bear TLK, Dalziel JE, Coad J, Roy NC, Butts CA, Gopal PK. The Role of the Gut Microbiota in Dietary Interventions for Depression and Anxiety. Adv Nutr. 2020; 11:890–907. https://doi.org/10.1093/advances/nmaa016 [PubMed]
- 249. Ezurike PU, Odunola E, Oke TA, Bakre AG, Olumide O, Odetoye O, Alege AM, Abiodun OO. Ganoderma lucidum ethanol extract promotes weight loss and improves depressive-like behaviors in male and female Swiss mice. Physiol Behav. 2023; 265:114155. https://doi.org/10.1016/j.physbeh.2023.114155 [PubMed]
- 250. Jacka FN, Cherbuin N, Anstey KJ, Sachdev P, Butterworth P. Western diet is associated with a smaller hippocampus: a longitudinal investigation. BMC Med. 2015; 13:215. https://doi.org/10.1186/s12916-015-0461-x [PubMed]
- 251. Brenton JN, Lehner-Gulotta D, Woolbright E, Banwell B, Bergqvist AGC, Chen S, Coleman R, Conaway M, Goldman MD. Phase II study of ketogenic diets in relapsing multiple sclerosis: safety, tolerability and potential clinical benefits. J Neurol Neurosurg Psychiatry. 2022; 93:637–44. https://doi.org/10.1136/jnnp-2022-329074 [PubMed]
- 252. Pursey KM, Stanwell P, Callister RJ, Brain K, Collins CE, Burrows TL. Neural responses to visual food cues according to weight status: a systematic review of functional magnetic resonance imaging studies. Front Nutr. 2014; 1:7. https://doi.org/10.3389/fnut.2014.00007 [PubMed]
- 253. Adjibade M, Andreeva VA, Lemogne C, Touvier M, Shivappa N, Hébert JR, Wirth MD, Hercberg S, Galan P, Julia C, Assmann KE, Kesse-Guyot E. The Inflammatory Potential of the Diet Is Associated with Depressive Symptoms in Different Subgroups of the General Population. J Nutr. 2017; 147:879–87. https://doi.org/10.3945/jn.116.245167 [PubMed]
- 254. Yin W, Löf M, Chen R, Hultman CM, Fang F, Sandin S. Mediterranean diet and depression: a population-based cohort study. Int J Behav Nutr Phys Act. 2021; 18:153. https://doi.org/10.1186/s12966-021-01227-3 [PubMed]
- 255. Canheta ABdS, Santos ASEAdS, Souza JDd, Silveira EA. Traditional Brazilian diet and extra virgin olive oil reduce symptoms of anxiety and depression in individuals with severe obesity: Randomized clinical trial. Clin Nutr. 2021; 40:404–11. https://doi.org/10.1016/j.clnu.2020.05.046 [PubMed]
- 256. Rodriguez-Lozada C, Cuervo M, Cuevas-Sierra A, Goni L, Riezu-Boj JI, Navas-Carretero S, Milagro FI, Martinez JA. Changes in Anxiety and Depression Traits Induced by Energy Restriction: Predictive Value of the Baseline Status. Nutrients. 2019; 11:1206. https://doi.org/10.3390/nu11061206 [PubMed]
- 257. Hariri N, Darafshi Ghahroudi S, Jahangiri S, Borumandnia N, Narmaki E, Saidpour A. The beneficial effects of sumac (Rhus coriaria L.) supplementation along with restricted calorie diet on anthropometric indices, oxidative stress, and inflammation in overweight or obese women with depression: A randomized clinical trial. Phytother Res. 2020; 34:3041–51. https://doi.org/10.1002/ptr.6737 [PubMed]
- 258. da Silva Borges D, Fernandes R, Thives Mello A, da Silva Fontoura E, Soares Dos Santos AR, Santos de Moraes Trindade EB. Prebiotics may reduce serum concentrations of C-reactive protein and ghrelin in overweight and obese adults: a systematic review and meta-analysis. Nutr Rev. 2020; 78:235–48. https://doi.org/10.1093/nutrit/nuz045 [PubMed]
- 259. Sanchez M, Darimont C, Panahi S, Drapeau V, Marette A, Taylor VH, Doré J, Tremblay A. Effects of a Diet-Based Weight-Reducing Program with Probiotic Supplementation on Satiety Efficiency, Eating Behaviour Traits, and Psychosocial Behaviours in Obese Individuals. Nutrients. 2017; 9:284. https://doi.org/10.3390/nu9030284 [PubMed]
- 260. Hulkkonen P, Kataja EL, Vahlberg T, Koivuniemi E, Houttu N, Pellonperä O, Mokkala K, Karlsson H, Laitinen K. The efficacy of probiotics and/or n-3 long-chain polyunsaturated fatty acids intervention on maternal prenatal and postnatal depressive and anxiety symptoms among overweight and obese women. J Affect Disord. 2021; 289:21–30. https://doi.org/10.1016/j.jad.2021.04.006 [PubMed]
- 261. Vendrik KEW, Ooijevaar RE, de Jong PRC, Laman JD, van Oosten BW, van Hilten JJ, Ducarmon QR, Keller JJ, Kuijper EJ, Contarino MF. Fecal Microbiota Transplantation in Neurological Disorders. Front Cell Infect Microbiol. 2020; 10:98. https://doi.org/10.3389/fcimb.2020.00098 [PubMed]
- 262. Su CH, Chuang HC, Hong CJ. Physical exercise prevents mice from L-Kynurenine-induced depression-like behavior. Asian J Psychiatr. 2020; 48:101894. https://doi.org/10.1016/j.ajp.2019.101894 [PubMed]
- 263. Liu W, Xue X, Xia J, Liu J, Qi Z. Swimming exercise reverses CUMS-induced changes in depression-like behaviors and hippocampal plasticity-related proteins. J Affect Disord. 2018; 227:126–35. https://doi.org/10.1016/j.jad.2017.10.019 [PubMed]
- 264. Ruilian L, Honglin Q, Jun X, Jianxin L, Qingyun B, Yilin C, Haifeng M. H2S-mediated aerobic exercise antagonizes the hippocampal inflammatory response in CUMS-depressed mice. J Affect Disord. 2021; 283:410–9. https://doi.org/10.1016/j.jad.2021.02.005 [PubMed]
- 265. Morgan JA, Singhal G, Corrigan F, Jaehne EJ, Jawahar MC, Baune BT. The effects of aerobic exercise on depression-like, anxiety-like, and cognition-like behaviours over the healthy adult lifespan of C57BL/6 mice. Behav Brain Res. 2018; 337:193–203. https://doi.org/10.1016/j.bbr.2017.09.022 [PubMed]
- 266. Faulconbridge LF, Driscoll CFB, Hopkins CM, Bailer Benforado B, Bishop-Gilyard C, Carvajal R, Berkowitz RI, DeRubeis R, Wadden TA. Combined Treatment for Obesity and Depression: A Pilot Study. Obesity (Silver Spring). 2018; 26:1144–52. https://doi.org/10.1002/oby.22209 [PubMed]
- 267. Vancini RL, Rayes ABR, Lira CABd, Sarro KJ, Andrade MS. Pilates and aerobic training improve levels of depression, anxiety and quality of life in overweight and obese individuals. Arq Neuropsiquiatr. 2017; 75:850–7. https://doi.org/10.1590/0004-282X20170149 [PubMed]
- 268. Seo EY, Kim YS, Lee YJ, Hur MH. Virtual Reality Exercise Program Effects on Body Mass Index, Depression, Exercise Fun and Exercise Immersion in Overweight Middle-Aged Women: A Randomized Controlled Trial. Int J Environ Res Public Health. 2023; 20:900. https://doi.org/10.3390/ijerph20020900 [PubMed]