



Introduction
Pancreatic cancer (PC) is the worst prognosis malignancy within the category of digestive system tumors, known for its aggressive nature and significant mortality rate [1, 2]. While significant advancements have been made in the management of pancreatic cancer, a majority of patients experience postoperative recurrence due to local infiltration or extensive metastasis [3, 4]. Due to the advanced stage of PC in the majority of diagnosed patients, it is not feasible to perform a comprehensive radical operation for PC patients. Hence, there exists a significant requirement to discover efficacious treatment options and preventive strategies against metastasis and recurrence, aiming at enhancing the overall management of PC.
MicroRNAs (miRNAs), the regulators mentioned are highly regarded as crucial factors in tumor development and possess significant biomarker and diagnostic potential [5–7]. Recently, a growing body of research has revealed that the role of miRNAs as tumor suppressor genes is evident in various types of tumors. The malignant progression of PC cells is regulated by miR-939-5p through targeting ARHGAP4 in PC [8]. MiR-211 has been confirmed to be associated with pancreatic cancer [9]. MiRNAs play a pivotal role in orchestrating cancer development by modulating key cellular processes, including apoptosis, cell proliferation, migration, and invasion [10–12]. MiR-211-5p is a highly conserved microRNA, exhibits remarkable conservation across species, which is involved in the process of tumorigenesis and development in different cancers [13–15]. MiR-211-5p was recently characterized as a tumor suppressor in prostate cancer [16]. Nevertheless, the mechanisms underlying miR-211-5p in PC remains to be investigated.
Bone morphogenetic protein 2 (BMP2), belongs to the transforming growth factors-β (TGF-β) subfamily, was initially discovered for its ability to stimulate osteogenesis and facilitate the growth of bone tissue [17, 18]. The aberrant expression of BMP2 has been demonstrated to participate in the biological process of outside the skeletal system, including regulating the proliferation, differentiation, processes of cancer cells [19]. The most recent research results have revealed that the promotion of epithelial mesenchymal transition (EMT) and enhancement of tumor cells’ metastatic capability are facilitated by BMP2 [20]. Moreover, evidence has demonstrated that BMP2 is upregulated in many cancers, and is related to the progression and growth of malignant tumors including non-small-cell lung cancer [21], osteosarcoma [22], gastric cancer [23], and nasopharyngeal carcinoma [24]. The complete elucidation of the regulatory mechanism of BMP2 in PC necessitates further investigation.
The preliminary results of this investigation revealed a significant reduction in the levels of miR-211-5p observed in both PC specimens and cells. Furthermore, when the levels of miR-211-5p was increased, it inhibited the proliferative ability of PC, as evidenced through the CCK-8 assay. The transwell assay revealed a notable decrease in the migratory capacity of PC cells upon upregulation of miR-211-5p expression. Additionally, according to the luciferase reporter assay, it was observed that BMP2 expression is directly suppressed by miR-211-5p specific binding to the 3’-UTR region of BMP2. Further rescue experiments demonstrate that the proliferation and migration of PC are suppressed by miR-211-5p through its targeting of BMP2. The function of the MiR-211-5p/BMP2 axis in PC development highlights its promising target for therapeutic interventions of PC.
Materials and Methods
Patient samples
All PC samples and their corresponding adjacent samples were obtained from patients who underwent surgeries at the Second Affiliated Hospital of Nanchang University between January 2015 and January 2017. All samples were stored at a temperature of -80° C after removal. The Ethics Committee of the second affiliated Hospital of Nanchang University granted approval for our research.
Cell culture
The human normal pancreatic cell line (HPDE6-C7) and PC cell lines (PANC-1, AsPC-1, BxPC-3, CFPAC-3, SW1990) were purchased from the esteemed Shanghai Institute of Cells under the auspices of the Chinese Academy of Sciences. All cells were cultured in DMEM (Gibco; Thermo Fisher Scientific, USA) with 10% FBS (Gibco; Thermo Fisher Scientific, USA) in a 37° C with 5% CO2. MiR-NC mimics and miR-211-5p mimics were synthesized by Ribo Bio (Guangzhou, China). Small interfering RNAs (siRNAs) specific for BMP2 (siBMP2) and pcDNA 3.1(+)-BMP2 were used as described previously [18].
The prediction of miRNA targets
The potential gene targeted by miR-211-5p was predicted using TargetScan (https://www.Targetscan.org/). Additionally, it was employed to explore the alleged interaction site between them.
Cell counting kit-8 (CCK-8) assay
PC cells were harvested 48 hours post-transfection and evenly distributed at a seeding density of 2000 cells per well. Following that, PC cells were different time intervals: 24h, 48h, 72h, and 96h. Subsequently, each well was supplemented with a volume of CCK-8 solution (Dojindo Molecular Technologies, Kumamoto, Japan) measuring only 10 μL and incubated for a duration of 2 hours. The optical density (OD) value of each hole was measured by a microplate reader set to an absorption wavelength of 490 nm.
Transwell assay
The transwell chamber was seeded with PC cells at a density of 1x104 cells per group. Following 24 hours, subependymal cells were fixed using formaldehyde, and a staining solution consisting of 0.2% crystal violet was applied.
Western blotting
As mentioned earlier, western blotting analysis used as previously described [19]. The antibodies examined in this investigation encompass the BMP2 rabbit antibody (ab53146, 1:1000, Abcam, USA), mouse antibody GAPDH (ab8245, 1:3000, Abcam, USA).
Quantitative real-time PCR (qRT-PCR)
The RNA extraction process involved the utilization of the RNAiso Plus reagent (Takara, Japan) on either tissue samples or cultured cells. Subsequently, reverse transcription was performed on the extracted RNA to generate complementary DNA (cDNA), which was subsequently employed for polymerase chain reaction (PCR) amplification. The primers were as follows: miR-211-5p (F): 5’-ACACTCCAGCTGGGCAAGTAGCATCAACTA-3’, miR-211-5p (R): 5’-TGGTGTCGTGGAGTCG-3’, U6 (F): 5’-CTCGCTTCGGCAGCACA-3’ and U6 (R): 5’-AACGCTTCACGAATTTGCGT-3’, BMP2 (F): 5’-GGTATGCTCCTGCCGCCTGCA-3’ and BMP2 (R): 5’-ATCAGCATAGGCCGGTGCAA-3’, GAPDH (F): 5’-CAAGGTCATCCATGACAACTTTG-3’, and GAPDH (R): 5’-GTCCACCACCCTGTTGCTGTAG-3’. The ABI Step One software (Applied Biosystems, Foster City, CA, USA) was for data analysis, while the 2-ΔΔCt method was employed to analyze the relative expression levels of mRNA.
Dual-luciferase reporter system
The pmirGLO vector 3’-UTR was utilized to construct the wild-type (Wt) and mutant (Mut) BMP2, which were synthesized by Shanghai GenePharma, China. The cells were transfected with miR-NC mimics or miR-211-5p-mimics at a concentration of 20 nM. Following a 48-hour incubation period, the cell luciferase activity was assessed using the dual-luciferase reporting system from Promega (Madison, WI, USA). The experimental procedure followed the previously described methodology [25].
Xenograft experiments
The in vivo xenograft model was established using PC cells that were transfected with either miR-NC mimic or miR-211-5p mimic. These cells were then randomly divided into two groups. Depending on the grouping, a dose of 5×106 miR-NC mimics or miR-211-5p mimics suspended in a mixture of 0.1 mL PBS and 0.1 mL Matrigel (BD Biosciences, San Jose, CA, USA) was subcutaneously injected into the right axilla of each mouse.
Statistical analysis
The analysis of the data was conducted using SPSS 22.0 software (Chicago, IL, USA), and all conclusions were based on this examination. GraphPad Prism 8.0. The significance of differences between two and multiple groups was assessed through One-way ANOVA and Student’s t-test. p < 0.05 was considered statistically significant.
Consent for publication
All authors agree for publication.
Results
The expression levels of miR-211-5p exhibited a significant reduction in both pancreatic cancer (PC) tissues and cells
The suppressive impact of miR-211-5p on neoplastic cell proliferation has been extensively documented across various human malignancies. To investigate the function of miR-211-5p in pancreatic cancer (PC), we first examined the levels of miR-211-5p between PC tissues and their adjacent tissues. qRT-PCR assay indicated that miR-211-5p was highly expressed in corresponding adjacent tissues compared to PC tissues (Figure 1A). The clinicopathological correlation analysis revealed a significant association between the levels of miR-211-5p and TNM stage (Table 1, p = 0.023). In addition, a significant correlation was observed between the survival of patients with pancreatic cancer and the levels of miR-211-5p, as determined by Kaplan-Meier survival analysis. (Figure 1B, p = 0.0002). Subsequently, the analysis was conducted on the levels of miR-211-5p in PC cells. Our findings indicated a decrease in the levels of miR-211-5p in ASPC-1, BXPC-3, and PANC-1 cells when compared to their normal cell (Figure 1C). Our findings showed that miR-211-5p served as a promising prognostic biomarker for individuals diagnosed with PC.
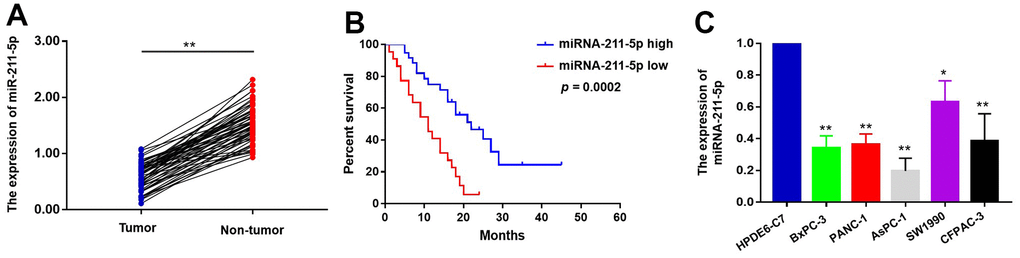
Figure 1. The expression levels of miR-211-5p in PC tissues and cells. (A) The expression of miR-211-5p was detected by qRT-PCR between PC tissues and corresponding adjacent tissues (n=58, p < 0.001, Wilcoxon signed-rank test). GAPDH was used as a loading control. Tumor represented PC tissues and non-tumor represented corresponding adjacent tissues. (B) Kaplan–Meier survival curve of the relationship between miR-211-5p high and low expression groups and the prognosis of PC patients (n=58, p=0.0002, Log-rank test). (C) The expression of miR-211-5p in HPDEC, PANC-1, AsPC-1, SW1990, BxPC-3 and CFPAC-1 cells analyzed by qRT-PCR. Data represent the mean ± SD of triplicate experiments and were statistically analyzed with Student’s t-test, *p < 0.05, ** p < 0.01.
Table 1. The relationship between miR-211-5p expression and clinicopathological characteristics in 58 PC patients.
| Characteristics | No. of patients | miR-211-5p expression | p-value | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| High n=37 | Low n=21 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Age (years) | 0.601 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| <60 | 25 | 15 | 10 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ≥60 | 33 | 22 | 11 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gender | 0.702 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Male | 24 | 16 | 8 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Female | 34 | 21 | 13 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Lymph node metastasis | 0.732 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Negative | 33 | 23 | 14 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Positive | 25 | 14 | 7 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TNM stage | 0.023 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| I and II | 36 | 27 | 9 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| III and IV | 22 | 10 | 12 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Bold values are statistically significant, p < 0.05. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The in vitro experiments demonstrated that the upregulation of miR-211-5p resulted in a decrease in the proliferation and metastatic potential of PC cells
The observed expression pattern and potential clinical significance of miR-211-5p indicate its possible inhibitory role in PC. To explore the impact of miR-211-5p on PC cells, we introduced miR-NC mimics and miR-211-5p mimics into PC cells through transfection. The qRT-PCR assay demonstrated a notable increase in the levels of miR-211-5p in both AsPC-1 and BxPC-3 cells (Figure 2A). Moreover, the proliferation ability of PC cells was assessed using the CCK-8 assay. Our findings indicated that the enhanced the levels of miR-211-5p exerted a substantial suppressive effect on cellular proliferation in both AsPC-1 and BxPC-3 cell lines (Figure 2B, 2C). Similarly, transwell assay demonstrated that cell metastasis was inhibited by the upregulation of miR-211-5p in AsPC-1 and BxPC-3 cells (Figure 2D, 2E). Our findings suggested that miR-211-5p exerted a suppressive effect on the cellular proliferation and metastasis of PC.
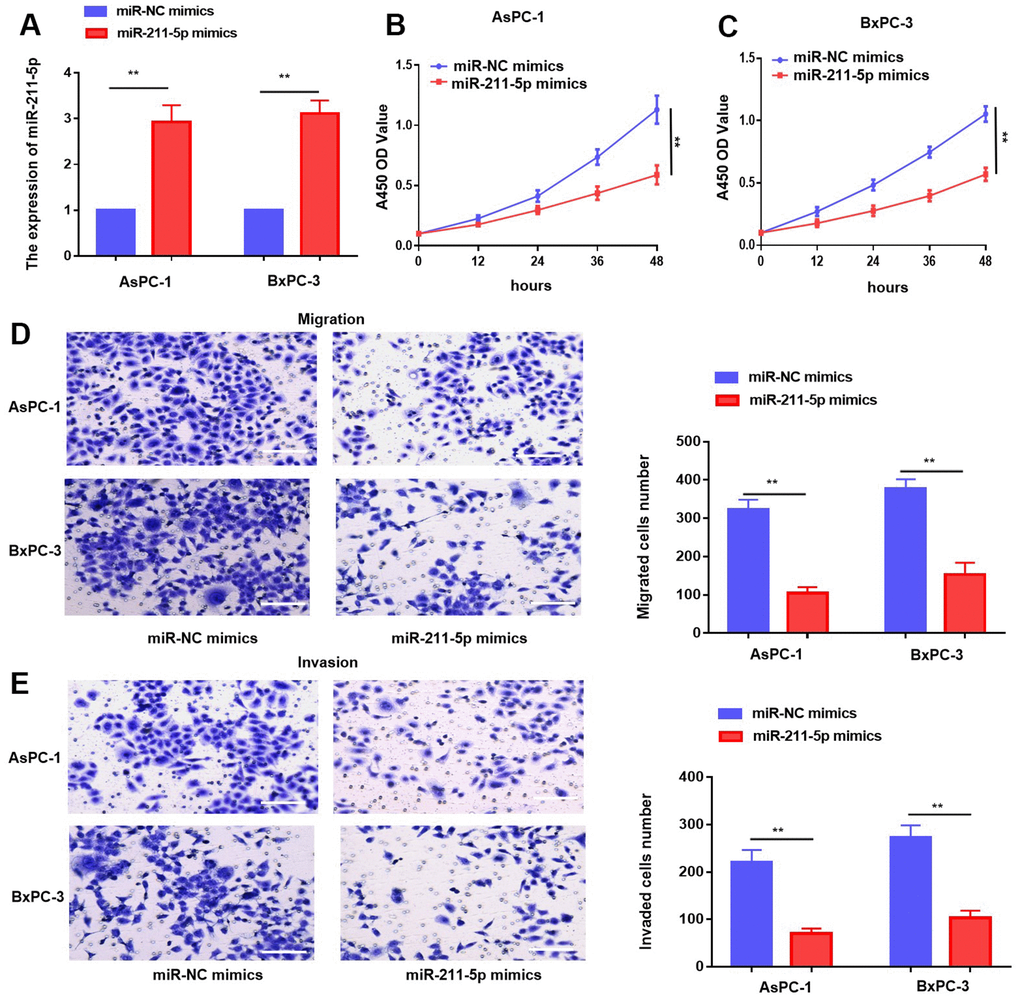
Figure 2. Overexpression of miR-211-5p inhibited cell proliferation and metastasis in PC in vitro. (A) miR-211-5p expression was analyzed in AsPC-1 and BxPC-3 cells transient transfected with miR-NC mimics, and miR-211-5p mimics by qRT-PCR. (B, C) CCK-8 analysis with miR-211-5p mimics or miR-NC mimics in AsPC-1 and BxPC-3. (D, E) Transwell assay for cell migration with miR-211-5p mimics or miR-NC mimics in AsPC-1 and BxPC-3. **p < 0.01, scale bar, 100 μm.
In vivo, the upregulation of miR-211-5p resulted in the suppression of tumor development
To evaluate the function of miR-211-5p on PC in vivo, we injected intravenous injections of PC cells that were transfected with miR-NC mimics and miR-211-5p mimics to BALB/c nude mice (male, 5 weeks). The size of the tumors injected with cells transfected with miR-211-5p mimics was found to be significantly smaller compared to those in the group of mice treated with miR-NC mimics, as indicated by the study (Figure 3A). Moreover, the miR-211-5p mimics group exhibited significantly reduced tumor weight and volume compared to the miR-NC mimics group (Figure 3B, 3C). Altogether, the significant inhibition of in vivo PC growth was observed upon the upregulation of miR-211-5p.
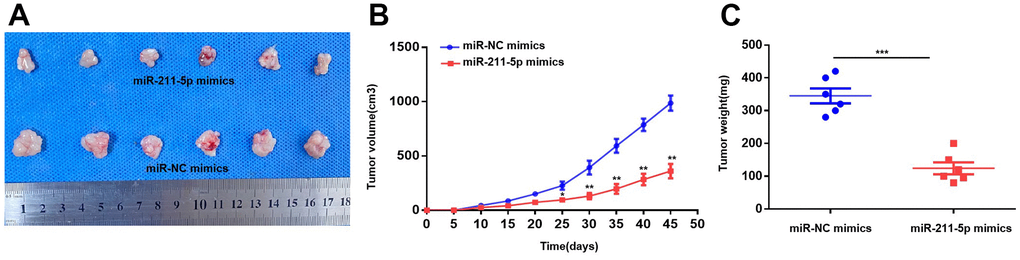
Figure 3. Overexpression of miR-211-5p inhibits tumor proliferation in vivo. (A) The images of tumors the miR-NC mimics and miR-211-5p mimics groups. (B) Comparison of tumor volume of the two groups every 5 days. (C) Comparison of tumor weight of the two groups 45 days after the subcutaneous injection. *p < 0.05, **p < 0.01.
BMP2 was identified as a direct target of miR-211-5p
To explore the mechanism responsible for the inhibitory function of miR-211p, we employed the online platform TargetScan to predict BMP2 as a potential target of miR-211-5p (Figure 4A). Notably, the experiment involving the dual luciferase reporter assay was performed, and as expected, the miR-211-5p-mimics demonstrated inhibition of relative luciferase activity in AsPC-1 and BxPC-3 cells by targeting the BMP2 3’ UTR-WT (Figure 4B). Furthermore, our findings revealed that the upregulation of miR-211-5p decreased the levels of BMP2 mRNA and protein in AsPC-1 and BxPC-3 cells (Figure 4C, 4D). To further explore their correlation in clinical specimens, we evaluated the expression levels of BMP2 in 58 sets of tumor and non-tumor pancreatic cancer tissues. In line with prior discoveries, the expression of BMP2 in PC samples was found to be significantly elevated compared to non-tumor PC tissues (Figure 4E). Moreover, there was a negative correlation observed between the levels of BMP2 and miR-211-5p in PC samples (Figure 4F, r=-0.3112, p = 0.0174). Furthermore, the GEPIA (Gene Expression Profiling Interactive Analysis) findings revealed a significant association between increased BMP2 levels and decreased survival rates among PC patients, as indicated by the Kaplan-Meier survival analysis (Figure 4F, p = 0.041). This evidence verified that BMP2 was a direct target of miR-211-5p in PC.
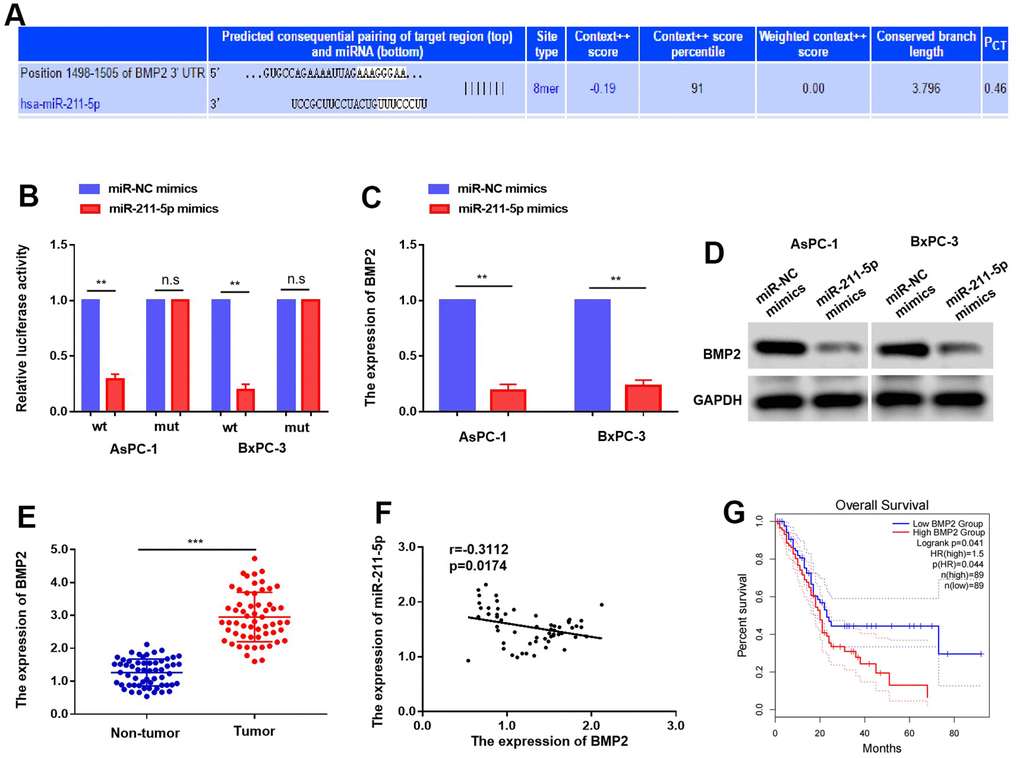
Figure 4. Targeting of BMP2 by miR-211-5p in PC. (A) BMP2 was predicted as a potential targets of miR-211-5p. (B) The luciferase activity of BMP2 WT or MUT’3 level of BMP2 affected with miR-211-5p mimics in AsPC-1 and BxPC-3. (C, D) the expression mRNA and protein of BMP2 with miR-211-5p mimics in AsPC-1 and BxPC-3 cells tested by qRT-PCR and western blot. (E) The expression of BMP2 mRNA is upregulated in PC tissues. (F) The relationship of BMP2 and miR-211-5p in PC. (r = −0.3112, p = 0.0174). (G) The overall survival time of PC patients with different expression of BMP2.
Upregulation of BMP2 enhances the proliferative and metastatic potential of PC cells
Given the well-established oncogenic role of BMP2, our study aims to validate its functional significance in PC cells. The efficiency of BMP2 siRNAs in suppressing gene expression was evaluated in AsPC-1 and BxPC-3 cells through transfection, followed by analysis using qRT-PCR and western blotting [17]. The transfection of si-BMP2 resulted in a notable decrease in the levels of BMP2 in AsPC-1 and BxPC-3 cells (Figure 5A, 5B). Moreover, we further investigated the effects of si-BMP2 on the proliferative and metastatic potential of PC. The outcomes obtained from the CCK-8 assay revealed a notable reduction in proliferation rates for both AsPC-1 and BxPC-3 cell lines transfection with si-BMP2 (Figure 5C, 5D). Furthermore, the transwell assay revealed a reduction in migration activity of AsPC-1 and BxPC-3 cells following si-BMP2 transfection when compared to the control group. (Figure 5E, 5F). The combined results of our investigation suggest that the suppression of BMP2 activity leads to a decrease in both growth and metastasis.
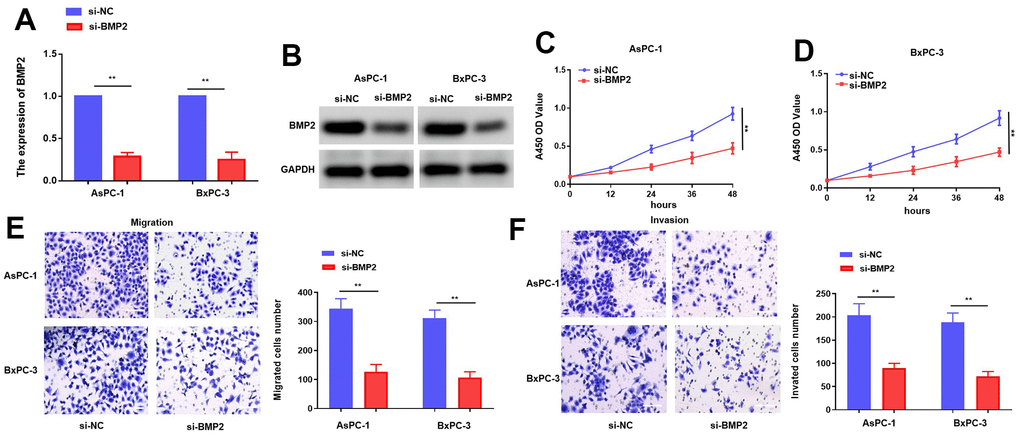
Figure 5. BMP2 inhibits PC cells proliferation and metastasis while it is knocking down. (A, B) The mRNA and protein of BMP2 affected by si-BMP2 siRNAs in PC cells. (C, D) CCK-8 assay with si-BMP2. (E, F) Transwell assay with si-BMP2 in PC cells. *p < 0.05 and **p < 0.01, scale bar, 100 μm.
MiR-211-5p inhibits the progression of PC via BMP2
To elucidate the pivotal role of BMP2 in orchestrating regulation of miR-211-5p, we transfected miR-211-5p mimics or miR-NC mimics into PC cells together with pcDNA 3.1(+)–BMP2 through transfection. The results obtained from qRT-PCR and western blotting revealed a significant reduction in the abundance of BMP2 caused by miR-211-5p, which was partly reversed by BMP2 (Figure 6A, 6B). The CCK-8 assay revealed that the inhibitory impact of miR-211-5p on cellular proliferation in AsPC-1 and BxPC-3 cells was counteracted by BMP2 overexpression (Figure 6C). Additionally, the transwell assay also revealed that BMP2 upregulation attenuated the inhibitory effect of miR-211-5p on cellular metastasis in AsPC-1 and BxPC-3 cells (Figure 6D, 6E). Our findings suggest that the modulation of BMP2 plays a significant role in mediating the regulatory effects of miR-211-5p on the proliferative and metastatic of PC cells.
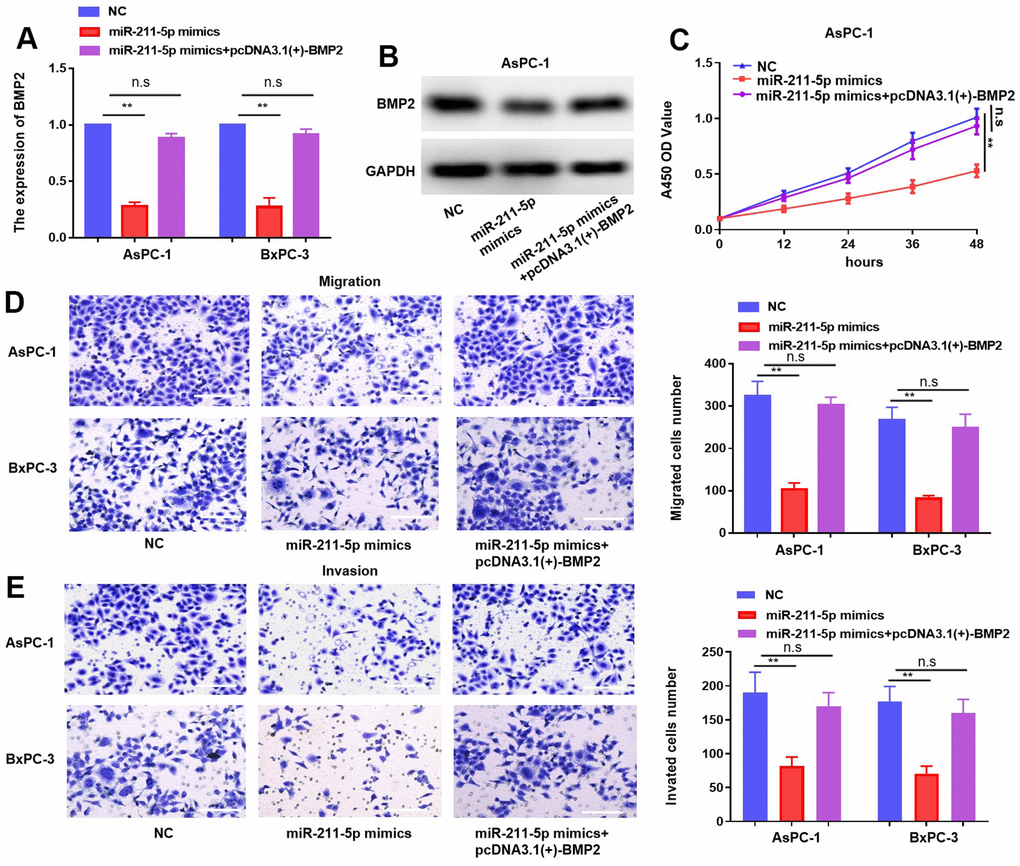
Figure 6. MiR-211-5p effect rescued by the overexpression of BMP2 in PC. (A, B) Expression of mRNA and protein of BMP2 in PC cells under transfection of miR-211-5p mimics and plasmid of pcDNA3.1(+)-BMP2. (C) Cell proliferation in PC cells under transfection of miR-211-5p mimics and pcDNA3.1(+)-BMP2 plasmid. (D, E) Transwell assay for PC cells migration under transfection of miR-211-5p mimics and pcDNA3.1(+)-BMP2. **p < 0.01, scale bar, 100 μm.
Discussion
Accumulating evidences has showed that miRNAs serve as a crucial role as regulators in the progression of oncogenesis and are recently extensively studied in the development of PC [26, 27]. Dysregulation of miRNAs plays a role in the progression of PC by initiating cell growth signaling and promoting the upregulation of genes associated with metastasis [28, 29]. Firstly, it is recommended that studies of miR-211-5p in other cancers could be sufficiently discussed. However, there has been a scarcity of relevant research investigating the functional of miR-211-5p in PC. Our investigation unveiled a significant reduction in the levels of miR-211-5p in PC, its presence was discovered to be associated with a favorable prognosis among individuals diagnosed with PC. Wang et al. it was discovered that bladder cancer exhibited a notable decrease in miR-211-5p, which had the ability to suppress bladder metastasis through its targeting of HDAC9 [30]. The results obtained from their investigation, in conjunction with our own clinical observations, revealed that miR-211-5p possessed the capability to impede the advancement of PC. Furthermore, in function assays, we noticed a notable suppression of the proliferative and metastatic abilities of PC cells when miR-211-5p was overexpressed. Our findings have revealed the significant role played by miR-211-5p in the progression of PC.
To further elucidate its underlying molecular mechanisms, we then study the potential functional mechanisms of miR-211-5p in PC cells by investigating the downstream targets. Based on the comprehensive analysis conducted using bioinformatics tool TargetScan and incorporating findings from previous studies [31]. We placed emphasis on bone morphogenetic protein 2 (BMP2), which has been found to regulate bone and cartilage formation [32, 33]. Although the relationship between BMP2 and various tumors, as well as the underlying mechanism of action, has been previously described, the impact of BMP2 on PC development remains elusive [34, 35]. The bioinformatics database analysis revealed BMP2 as the potential gene targeted by miR-211-5p. Further investigation using luciferase gene detection technology confirmed the specific binding of miR-211-5p to a target site located on the 3’untranslated region (3’UTR) of BMP2. Moreover, the mRNA and protein expression of BMP2 was detected in cells transfected with miR-211-5p mimics, confirming that miR-211-5p specifically targets BMP2 gene. The regulation of BMP2 was found to be influenced by miR-211-5p at the post-transcriptional level. In the present study, we discovered that BMP2 was a target of miR-211-5p.
In summary, our results indicate that the functionality of BMP2 in PC cells is directly affected by miR-211-5p through targeted regulation. Interestingly, we found that their expression levels in PC tissues were negatively correlated. Reduced expression of miR-211-5p was associated with a decrease in survival time among PC patients. Furthermore, rescue experiments demonstrated that the progression of PC is facilitated by miR-211-5p specifically targeting BMP2. These findings suggest that targeting miR-211-5p could be a promising approach for future therapeutic interventions in PC.
Conclusions
Our research results indicate that the inhibitory effect of pancreatic cancer cell proliferative and metastatic is significantly influenced through miR-211-5p, which interacts with BMP2. This discovery suggests that miR-211-5p may be a prognostic biomarker for PC. However, further investigation was needed to found other downstream targets regulated by miR-211-5p and explored its regulatory mechanism in cancer.
Author Contributions
J.W designed the research. D.L and C.L drafted the manuscript. C.L and Y.X prepared the figures. K.L and S.F analyzed the data. D.L and C.L performed the experiments of this study. D.L, C.L and J.D revised the manuscript. D.L, C.L and J.W provided the funding. All authors read and approved the final paper.
Acknowledgments
All authors are particularly grateful for the experimental equipment provided by Jiangxi key Laboratory of Molecular Medicine in the second affiliated Hospital of Nanchang University.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Ethical Statement and Consent
Our research was performed in accordance with the Declaration of Helsinki. The Ethics Committee of the Second Affiliated Hospital of Nanchang University approved the use of human participants, human data and human tissues in this study. The written informed consent was obtained from all patients. The Animal Research Committee of the Experimental Animal Science Center of Nanchang University approved the use of animals in this study.
Funding
This work was supported by grants from the National Natural Science Foundation of China (Nos. 82060435), the Project of the Jiangxi Provincial Department of Science and Technology (Nos. 20202BAB206052, 20232BAB216079), the First Affiliated Hospital of Nanchang University, Jiangxi Province, Young Talents Research and Cultivation Fund (Nos. YFYPY202253), and the Project of the Jiangxi Provincial Innovation Fund for Graduate Students (Nos. YC2021-B041).
References
- 1. Mizrahi JD, Surana R, Valle JW, Shroff RT. Pancreatic cancer. Lancet. 2020; 395:2008–20. https://doi.org/10.1016/S0140-6736(20)30974-0 [PubMed]
- 2. Pereira SP, Oldfield L, Ney A, Hart PA, Keane MG, Pandol SJ, Li D, Greenhalf W, Jeon CY, Koay EJ, Almario CV, Halloran C, Lennon AM, Costello E. Early detection of pancreatic cancer. Lancet Gastroenterol Hepatol. 2020; 5:698–710. https://doi.org/10.1016/S2468-1253(19)30416-9 [PubMed]
- 3. Ying H, Dey P, Yao W, Kimmelman AC, Draetta GF, Maitra A, DePinho RA. Genetics and biology of pancreatic ductal adenocarcinoma. Genes Dev. 2016; 30:355–85. https://doi.org/10.1101/gad.275776.115 [PubMed]
- 4. Allen PJ, Kuk D, Castillo CF, Basturk O, Wolfgang CL, Cameron JL, Lillemoe KD, Ferrone CR, Morales-Oyarvide V, He J, Weiss MJ, Hruban RH, Gönen M, et al. Multi-institutional Validation Study of the American Joint Commission on Cancer (8th Edition) Changes for T and N Staging in Patients With Pancreatic Adenocarcinoma. Ann Surg. 2017; 265:185–91. https://doi.org/10.1097/SLA.0000000000001763 [PubMed]
- 5. Chatterjee N, Rana S, Espinosa-Diez C, Anand S. MicroRNAs in Cancer: challenges and opportunities in early detection, disease monitoring, and therapeutic agents. Curr Pathobiol Rep. 2017; 5:35–42. https://doi.org/10.1007/s40139-017-0123-0 [PubMed]
- 6. Ryan BM. microRNAs in Cancer Susceptibility. Adv Cancer Res. 2017; 135:151–71. https://doi.org/10.1016/bs.acr.2017.06.004 [PubMed]
- 7. Wang J, Chen J, Sen S. MicroRNA as Biomarkers and Diagnostics. J Cell Physiol. 2016; 231:25–30. https://doi.org/10.1002/jcp.25056 [PubMed]
- 8. Shen Y, Chen G, Gao H, Li Y, Zhuang L, Meng Z, Liu L. miR-939-5p Contributes to the Migration and Invasion of Pancreatic Cancer by Targeting ARHGAP4. Onco Targets Ther. 2020; 13:389–99. https://doi.org/10.2147/OTT.S227644 [PubMed]
- 9. Han X, Saiyin H, Zhao J, Fang Y, Rong Y, Shi C, Lou W, Kuang T. Overexpression of miR-135b-5p promotes unfavorable clinical characteristics and poor prognosis via the repression of SFRP4 in pancreatic cancer. Oncotarget. 2017; 8:62195–207. https://doi.org/10.18632/oncotarget.19150 [PubMed]
- 10. Fan Y, Li Y, Zhu Y, Dai G, Wu D, Gao Z, Zhang L, Xu D. miR-301b-3p Regulates Breast Cancer Cell Proliferation, Migration, and Invasion by Targeting NR3C2. J Oncol. 2021; 2021:8810517. https://doi.org/10.1155/2021/8810517 [PubMed]
- 11. Liu H, Ma X, Niu N, Zhao J, Lu C, Yang F, Qi W. MIR-301b-3p Promotes Lung Adenocarcinoma Cell Proliferation, Migration and Invasion by Targeting DLC1. Technol Cancer Res Treat. 2021; 20:1533033821990036. https://doi.org/10.1177/1533033821990036 [PubMed]
- 12. Lulli V, Buccarelli M, Martini M, Signore M, Biffoni M, Giannetti S, Morgante L, Marziali G, Ilari R, Pagliuca A, Larocca LM, De Maria R, Pallini R, Ricci-Vitiani L. miR-135b suppresses tumorigenesis in glioblastoma stem-like cells impairing proliferation, migration and self-renewal. Oncotarget. 2015; 6:37241–56. https://doi.org/10.18632/oncotarget.5925 [PubMed]
- 13. Chen LL, Zhang ZJ, Yi ZB, Li JJ. MicroRNA-211-5p suppresses tumour cell proliferation, invasion, migration and metastasis in triple-negative breast cancer by directly targeting SETBP1. Br J Cancer. 2017; 117:78–88. https://doi.org/10.1038/bjc.2017.150 [PubMed]
- 14. Song D, Yang K, Wang W, Tian R, Wang H, Wang K. MicroRNA-211-5p promotes apoptosis and inhibits the migration of osteosarcoma cells by targeting proline-rich protein PRR11. Biochem Cell Biol. 2020; 98:258–66. https://doi.org/10.1139/bcb-2018-0380 [PubMed]
- 15. Wang H, Shi X, Guo Z, Zhao F, He W, Kang M, Lv Z. microRNA-211-5p predicts the progression of postmenopausal osteoporosis and attenuates osteogenesis by targeting dual specific phosphatase 6. Bioengineered. 2022; 13:5709–23. https://doi.org/10.1080/21655979.2021.2017626 [PubMed]
- 16. Qin X, Zhang J, Lin Y, Sun XM, Zhang JN, Cheng ZQ. Identification of MiR-211-5p as a tumor suppressor by targeting ACSL4 in Hepatocellular Carcinoma. J Transl Med. 2020; 18:326. https://doi.org/10.1186/s12967-020-02494-7 [PubMed]
- 17. Zuo WH, Zeng P, Chen X, Lu YJ, Li A, Wu JB. Promotive effects of bone morphogenetic protein 2 on angiogenesis in hepatocarcinoma via multiple signal pathways. Sci Rep. 2016; 6:37499. https://doi.org/10.1038/srep37499 [PubMed]
- 18. Wu JB, Fu HQ, Huang LZ, Liu AW, Zhang JX. Effects of siRNA-targeting BMP-2 on the abilities of migration and invasion of human liver cancer SMMC7721 cells and its mechanism. Cancer Gene Ther. 2011; 18:20–5. https://doi.org/10.1038/cgt.2010.55 [PubMed]
- 19. Fei ZH, Yao CY, Yang XL, Huang XE, Ma SL. Serum BMP-2 up-regulation as an indicator of poor survival in advanced non-small cell lung cancer patients. Asian Pac J Cancer Prev. 2013; 14:5293–9. https://doi.org/10.7314/apjcp.2013.14.9.5293 [PubMed]
- 20. Hsu YL, Huang MS, Yang CJ, Hung JY, Wu LY, Kuo PL. Lung tumor-associated osteoblast-derived bone morphogenetic protein-2 increased epithelial-to-mesenchymal transition of cancer by Runx2/Snail signaling pathway. J Biol Chem. 2011; 286:37335–46. https://doi.org/10.1074/jbc.M111.256156 [PubMed]
- 21. Choi YJ, Kim ST, Park KH, Oh SC, Seo JH, Shin SW, Kim JS, Kim YH. The serum bone morphogenetic protein-2 level in non-small-cell lung cancer patients. Med Oncol. 2012; 29:582–8. https://doi.org/10.1007/s12032-011-9852-9 [PubMed]
- 22. Sari AS, Demirçay E, Öztürk A, Terzi A, Karaöz E. The promising effects of BMP2 transfected mesenchymal stem cells on human osteosarcoma. Turk J Biol. 2021; 45:301–13. https://doi.org/10.3906/biy-2101-50 [PubMed]
- 23. Chen Z, Yuan L, Li X, Yu J, Xu Z. BMP2 inhibits cell proliferation by downregulating EZH2 in gastric cancer. Cell Cycle. 2022; 21:2298–308. https://doi.org/10.1080/15384101.2022.2092819 [PubMed]
- 24. Wang MH, Zhou XM, Zhang MY, Shi L, Xiao RW, Zeng LS, Yang XZ, Zheng XF, Wang HY, Mai SJ. BMP2 promotes proliferation and invasion of nasopharyngeal carcinoma cells via mTORC1 pathway. Aging (Albany NY). 2017; 9:1326–40. https://doi.org/10.18632/aging.101230 [PubMed]
- 25. Luo C, Lin K, Hu C, Zhu X, Zhu J, Zhu Z. LINC01094 promotes pancreatic cancer progression by sponging miR-577 to regulate LIN28B expression and the PI3K/AKT pathway. Mol Ther Nucleic Acids. 2021; 26:523–35. https://doi.org/10.1016/j.omtn.2021.08.024 [PubMed]
- 26. Ambros V. The functions of animal microRNAs. Nature. 2004; 431:350–5. https://doi.org/10.1038/nature02871 [PubMed]
- 27. Thomson JM, Newman M, Parker JS, Morin-Kensicki EM, Wright T, Hammond SM. Extensive post-transcriptional regulation of microRNAs and its implications for cancer. Genes Dev. 2006; 20:2202–7. https://doi.org/10.1101/gad.1444406 [PubMed]
- 28. Daoud AZ, Mulholland EJ, Cole G, McCarthy HO. MicroRNAs in Pancreatic Cancer: biomarkers, prognostic, and therapeutic modulators. BMC Cancer. 2019; 19:1130. https://doi.org/10.1186/s12885-019-6284-y [PubMed]
- 29. Wang W, Lou W, Ding B, Yang B, Lu H, Kong Q, Fan W. A novel mRNA-miRNA-lncRNA competing endogenous RNA triple sub-network associated with prognosis of pancreatic cancer. Aging (Albany NY). 2019; 11:2610–27. https://doi.org/10.18632/aging.101933 [PubMed]
- 30. Wang W, Liu Z, Zhang X, Liu J, Gui J, Cui M, Li Y. miR-211-5p is down-regulated and a prognostic marker in bladder cancer. J Gene Med. 2020; 22:e3270. https://doi.org/10.1002/jgm.3270 [PubMed]
- 31. Yang S, He P, Wang J, Schetter A, Tang W, Funamizu N, Yanaga K, Uwagawa T, Satoskar AR, Gaedcke J, Bernhardt M, Ghadimi BM, Gaida MM, et al. A Novel MIF Signaling Pathway Drives the Malignant Character of Pancreatic Cancer by Targeting NR3C2. Cancer Res. 2016; 76:3838–50. https://doi.org/10.1158/0008-5472.CAN-15-2841 [PubMed]
- 32. Mukai T, Otsuka F, Otani H, Yamashita M, Takasugi K, Inagaki K, Yamamura M, Makino H. TNF-alpha inhibits BMP-induced osteoblast differentiation through activating SAPK/JNK signaling. Biochem Biophys Res Commun. 2007; 356:1004–10. https://doi.org/10.1016/j.bbrc.2007.03.099 [PubMed]
- 33. Yamashita M, Otsuka F, Mukai T, Otani H, Inagaki K, Miyoshi T, Goto J, Yamamura M, Makino H. Simvastatin antagonizes tumor necrosis factor-alpha inhibition of bone morphogenetic proteins-2-induced osteoblast differentiation by regulating Smad signaling and Ras/Rho-mitogen-activated protein kinase pathway. J Endocrinol. 2008; 196:601–13. https://doi.org/10.1677/JOE-07-0532 [PubMed]
- 34. Matsumoto Y, Otsuka F, Takano M, Mukai T, Yamanaka R, Takeda M, Miyoshi T, Inagaki K, Sada KE, Makino H. Estrogen and glucocorticoid regulate osteoblast differentiation through the interaction of bone morphogenetic protein-2 and tumor necrosis factor-alpha in C2C12 cells. Mol Cell Endocrinol. 2010; 325:118–27. https://doi.org/10.1016/j.mce.2010.05.004 [PubMed]
- 35. Huang RL, Yuan Y, Tu J, Zou GM, Li Q. Opposing TNF-α/IL-1β- and BMP-2-activated MAPK signaling pathways converge on Runx2 to regulate BMP-2-induced osteoblastic differentiation. Cell Death Dis. 2014; 5:e1187. https://doi.org/10.1038/cddis.2014.101 [PubMed]