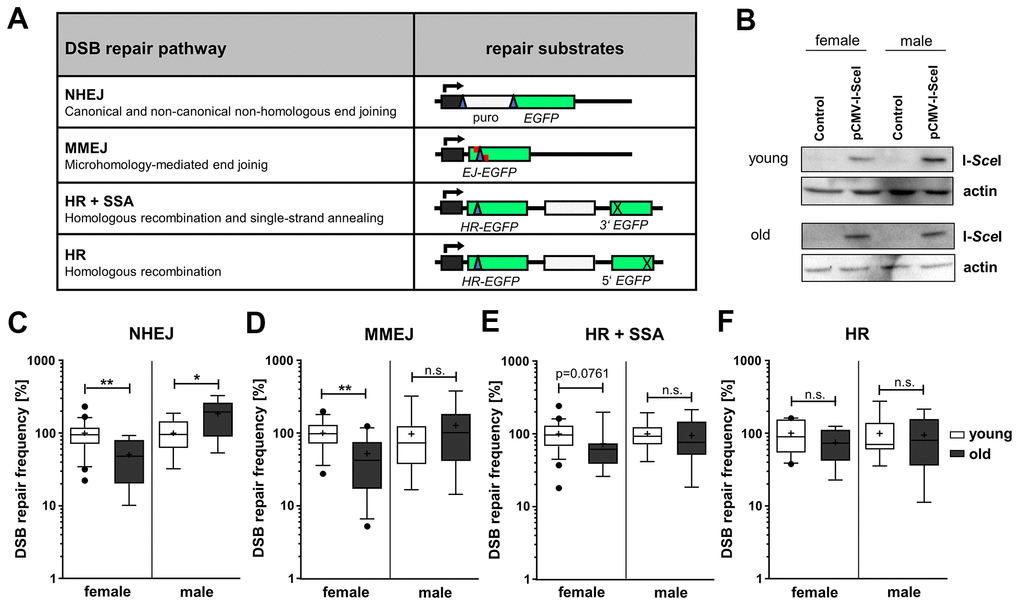
Figure 1.DSB repair pathway activities in PBL from different age groups. (A) DSB repair substrates. To detect NHEJ we used the reporter plasmid EJ5SceGFP, containing two tandem I-SceI cutting sequences flanking a spacer separating the transcriptional promoter from the EGFP coding sequence. For MMEJ we employed substrate EJ-EGFP harboring a mutated EGFP gene, with an I-SceI site flanked by 5 bp sequence repeats. Substrates HR-EGFP/3'EGFP and HR-EGFP/5'EGFP both contain EGFP lacking 4 bp at the position, where the I-SceI site was inserted. For detection of homologous repair (HR+SSA) substrate HR-EGFP/3'EGFP additionally contains 3'EGFP mutated at the start codon, while HR substrate HR-EGFP/5'EGFP contains 3’ truncated 5’EGFP. I-SceI site, blue triangle; cross, inactivating mutation/truncation; green bars, EGFP variants; white bars, spacer sequences; grey bar with kinked arrow, transcriptional promoter. (B) I-SceI protein levels were analyzed by Western Blotting. Shown are representative blots of each age group and gender. (C–F) DSB repair activity measurements. DSB repair frequencies by NHEJ (C), MMEJ (D), HR+SSA (E) and HR (F) are shown in box plots with mean value (cross), median (line) and 95% Cl (whiskers). Data was generated from samples of 7-24 female and 7-15 male donors per age group and normalized to the mean of young donors for each sex group on the experimental day (Supplementary Table 1). The absolute mean DSB repair frequencies for young donors set to 100% were: (C) NHEJ, 8.00 x 10-2 (female) and 2.80x10-2 (male); (D) MMEJ, 0.12x10-2 (female) and 0.13x10-2 (male); (E) HR+SSA, 0.78x10-2 (female) and 0.49x10-2 (male); (F) HR, 0.19x10-2 (female) and 0.18x10-2 (male) (Supplementary Table 1).