



Introduction
Activation of sirtuins has been proved to be positive in age-related diseases [1-5]. However, the role of sirtuin activation in acute diseases, such as inflammatory processes, has not been clearly defined. Stress-activated protein kinases (SAPKs) are essential components for intracellular signalling networks that serve to respond and adapt to extracellular changes. Stress-activated protein kinases play a key role in signal transduction to extracellular insults and have been involved in numerous diseases, including inflammatory processes [6-8]. Exposure of yeast to high osmolarity results in the transient activation of p38-related SAPK, Hog1, the yeast homologue of the p38 SAPK [9, 10]. The transient activation of the SAPK serves to control many aspects of the cell physiology ranging from control of metabolism, cell cycle progression and gene expression which are essential for cellular adaptation to stress [11-15]. Despite the essential role of Hog1 in cell survival upon stress, it has been known for a long time that sustained activation of the SAPK was deleterious for cell growth as it has been shown by other MAPKs [16-18]. Therefore, the dynamics of Hog1 activation are critical for cell fate determination. However, the mechanism by which Hog1 was deleterious for cell growth remained unknown.
The study by Vendrell et al shows that inhibition of cell growth by sustained activation of Hog1 is caused by the induction of apoptosis-like cell death in yeast. Specifically, activation of the Hog1 SAPK inhibits mitochondrial respiration that leads to an increase in ROS levels (Figure 1). This increase in ROS levels correlates with a decrease in mitochondrial respiration and it is prevented when cells are grown in the absence of oxygen. Therefore, the increase in ROS production seems to be caused by electron transport inhibition.
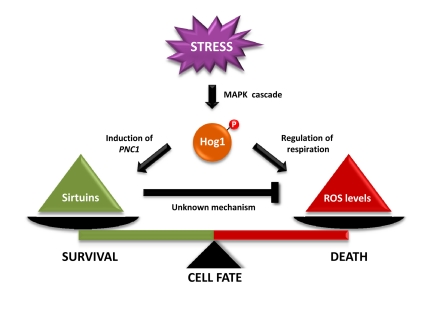
Figure 1. Tentative model that depicts the effect of Hog1 and sirtuins in dictating cell-fate determination. Hog1 inhibits mitochondrial respiration, which results in an increase in reactive oxygen species (ROS) accumulation that leads to cell death. In parallel, Hog1 induces PNC1 expression. Pnc1 activates Sir2, which mediates a decrease in ROS accumulation. Sir2 activation by the stress-activated protein kinase Hog1 relieves the Hog1-induced oxidative stress to prevent apoptotic cell death.
Several apoptotic makers can be detected upon Hog1 activation; such as an increase in SubG1 cells, DNA aggregation, TUNEL assay positive cells and metascaspase activation. Remarkably, deletion of genes encoding apoptosis mediators, such as the Omi yeast homologue (NMA111) or, to a lesser extent, the YCA1 caspase prevented Hog1-induced cell death. Taken together, Hog1-induced cell death is likely to be caused by apoptosis.
Unexpectedly, mutations in the E3 ubiquitin ligase CDC4, as well as the core components of the SCF complex (SKP1 and CDC53) partially suppressed Hog1-induced cell death. SCF mutations resulted in a reduced Msn2/Msn4 degradation that led to an increase in Msn2/Msn4 dependent gene expression (Figure 2).
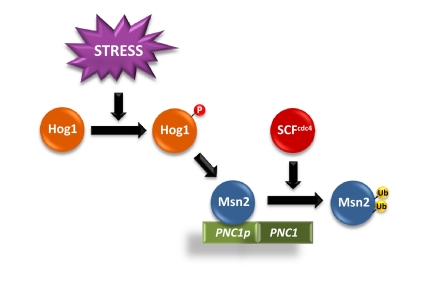
Figure 2. Scheme of Hog1-induced Msn2/Msn4 dependent gene expression upon SCFcdc4 regulation. Activation of Hog1 SAPK induces the activation of Msn2/Msn4-dependent genes, such as PNC1. The E3 SCFcdc4 ubiquitin ligase ubiquitylates the transcription factors to promote their degradation. Partial inactivation of SCFcdc4 decreases Msn2/4 degradation, which results in an increased accumulation of the TFs at the promoters and to an increase in Msn2/Msn4 dependent gene expression.
Interestingly, PNC1, an activator of the Sir2 histone deacetylase, is under the control of Msn2/Msn4 transcription factors [19]. The mutations on SCFcdc4 resulted in an increased association of Msn2/4 at the PNC1 promoter and to an increased PNC1 expression. The relevance of Msn2-mediated PNC1 expression and the role of Sir2 in cell survival are illustrated by the fact that deletion of MSN2/4, PNC1 or SIR2 resulted in cells unable to suppress the apoptosis caused by sustained activation of Hog1. In contrast, over-expression of MSN2, PNC1 or SIR2 prevented Hog1-induced cell death. Therefore, Sir2 activation is important to prevent SAPK-induced apoptotic cell death (Figure 1).
The mechanism by which Sir2 prevents apoptosis in yeast has not been defined. However, whereas Sir2 activation does not alter cellular respiration upon stress, it strongly reduces ROS production and hence, it is likely that this reduction in ROS permits cell survival upon Hog1 activation.
Activation of SAPK signalling is essential for cell adaptation to stress. Strikingly, sustained activation of the pathway unravels a more-complex scenario. Therefore, although a decrease in mitochondrial respiration might be important for adaptation to stress, an extended reduction of respiration leads to excessive ROS formation. To counteract cell damage, Hog1 seems to engage a protective program by inducing PNC1 gene expression, which concomitantly activates Sir2 to balance excessive ROS accumulation. Therefore, cell fate is dictated by the balance between ROS induced by Hog1 SAPK and the protective effects of Sir2.
Discussion
MAPKs, apoptosis and mitochondrial function
Apoptosis is caused by a variety of factors, such as oxidative stress [20]. Induction of apoptosis in response to MAPK activation has been described in several cell types and organisms [17, 21-24]. The p38 MAPK is activated by several stresses as well as oxidative stress [25, 26]. However, it is not known whether p38 is able to cause oxidative stress by altering mitochondrial function. Data from yeast suggests that, directly or indirectly, MAPKs can modulate mitochondrial respiration and ROS production. Then, it would be interesting to test whether, once p38 is activated, the MAPK regulates mitochondrial activity and ROS levels. A putative scenario would be that an increase of ROS could serve to create a positive feedback loop to sustain MAPK activation in the presence of stress.
The mechanism by which Hog1 regulates mitochondrial respiration and ROS levels remains unknown. It cannot be excluded that regulation of metabolism could lead to an altered TCA cycle and/or mitochondrial respiration. Therefore, the characterization of this phenomenon will allow identifying at which level the MAPKs can interact with the mitochondria: regulating mitochondria biogenesis, respiratory chain formation or respiratory chain activity. This would help to know whether the effect of Hog1 MAPK could be conserved in higher eukaryotes.
Sirtuins, apoptosis and lifespan extension
The effect of sirtuins in apoptotic cell death is not clear. Depending on expression of a specific sirtuin, the cell type and the stimuli, sirtuins prevent or induce apoptosis [27-29]. Moreover, it is not clear whether the effect of sirtuin overexpression to extend lifespan in higher eukaryotes, such as worms and flies [30]. In yeast, Sir2 prevents apoptosis mediated by Hog1. There have been several molecular alterations associated to replicative ageing, such as the accumulation of extrachromosomal ribosomal DNA circles (ERCs), the assimetrically distribution of oxidatively damaged proteins and/or altered mitochondria that seems to be, at least partially controlled by Sir2 [31-35]. However, the relationship of these phenomena and the induction of cell death is not yet defined. In yeast, Sir2 effect upon Hog1 activation Net1 (an anchor for Sir2 to rDNA) but not on Sir4 or HM loci [31, 36, 37]. This suggests that the role of Sir2 in Hog1-mediated cell death might be associated to ERCs in contrast to other Sir2 functions.
Recent data on the overexpression of mammalian Sir2 seems to show that Sir2 might not be responsible of lifespan extension albeit important for health in ageing processes [38]. Remarkably, overexpression of Sir2 prevents apoptosis in yeast. It is worth noting that this preventive effect is only observed upon strong overexpression of Sir2. Two-fold overexpression of Sir2 did not show a significant effect on preventing apoptosis upon Hog1 sustained activation. A recent study in mice with moderately overexpression of Sirt1 reports prevention in age-related diseases, but without effects in lifespan [5]. Therefore, the degree of Sir2 expression might be critical to affect some age-related effects that might be associated to the degree of oxidative stress in the cell.
Activation and regulation of apoptosis in an unicellular model
Apoptosis in yeast is a controversial issue. The benefit that could generate programmed cell death in a unicellular organism has been deeply discussed [39]. It is thought that the benefit of programmed cell death in yeast might be associated to a population rather to an individual [40]. Upon stress, the older or damaged individuals could enter into programmed cell death to allow other cells, genetically identical to them, increase their resources to survive.
The determination of cell fate upon stress could result of the balance between cell survival and cell death. Once cells are exposed to a stress, Hog1 SAPK is activated and cells enter into an alarm state. Upon a mild stress, Hog1 SAPK is active for a short period of time. Then, Hog1-induced sirtuin activation might be sufficient to prevent ROS accumulation, and the whole population survives. However, upon severe stress or sustained activation of the SAPK, longer activation of the MAPK Hog1 induces an increase on ROS production beyond a certain threshold. It is likely then that the preventive effect of sirtuin activation might not be sufficient to compensate ROS levels and certain cells of the population would undergo apoptosis and provide more resources to the younger cells to adapt and survive to a severe stress. The fact that both pro and anti-apoptotic pathways are controlled by the same protein, Hog1, emphasize the importance of the balance between the two responses that will determine cell fate.
Future Directions
Vendrell and collaborators have unraveled the ability of Sir2 to prevent ROS formation and cell death upon MAPK activation in yeast. However, the mechanism by which Sir2 prevents ROS formation remains unclear. It would be interesting in the field of sirtuins to define how Sir2 is able to reduce ROS levels and decipher whether Sir2 is modulating mitochondrial activity per se or it is inducing antioxidant defence mechanisms.
It is likely that, as occurs in yeast, the balance in sirtuin activation determines cell fate in higher eukaryotes cells. Recent data show that two-fold overexpression of Sir2 in flies and worms did not extend lifespan at least as much as it was reported before [41-45]. In fact, other previous works showed that, also in yeast, two-fold Sir2 overexpression does not extend lifespan in some backgrounds [46]. Data provided by Vendrell et al show the importance of sirtuin dose and activation in preventing ROS accumulation and apoptosis in yeast. Since only dramatic overexpression of Sir2 is able to prevent apoptosis in yeast, further studies with stronger sirtuin overexpression might uncover the role of sirtuins in higher eukaryotes.
Acknowledgments
We apologize to colleagues in the field for not citing all relevant papers owing to space constraints. We thank to EdeN for constant support. The laboratory of FP and EdeN is supported by grants from the Ministerio de Ciéncia y Innovación, the Consolider Ingenio 2010 programme and FP7 UNICELLSYS grant to F.P, EdeN. F.P. is also supported by the Fundación Marcelino Botín (FMB) and ICREA Acadèmia (Generalitat de Catalunya).
Conflicts of Interest
The authors of this manuscript have no conflict of interest to declare.
References
- 1. Finkel T, Deng CX, Mostoslavsky R. Recent progress in the biology and physiology of sirtuins. Nature. 2009; 460: 587 -591. [PubMed] .
- 2. Kaeberlein M, McVey M, Guarente L. The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes & Dev. 1999; 13: 2570 -2580. [PubMed] .
- 3. Kennedy BK, Gotta M, Sinclair DA, Mills K, McNabb DS, Murthy M, Pak SM, Laroche T, Gasser SM, Guarente L. Redistribution of silencing proteins from telomeres to the nucleolus is associated with extension of life span in S. cerevisiae. Cell. 1997; 89: 381 -391. [PubMed] .
- 4. Kaeberlein M. Lessons on longevity from budding yeast. Nature. 2010; 464: 513 -519. [PubMed] .
- 5. Herranz D, Muñoz-Martin M, Cañamero M, Mulero F, Martinez-Pastor B, Fernandez-Capetillo O, Serrano M. Sirt1 improves healthy aging and protects form metabolic syndrome associated cancer. Nat Commun. 2010; 1: 3 doi: 10.1038/ncomms1001 [PubMed] .
- 6. Wagner EF and Nebreda AR. Signal integration by JNK and p38 MAPK pathways in cancer development. Nature Rev. Cancer. 2009; 9: 537 -549. [PubMed] .
- 7. Kyriakis JM and Avruch J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol. Rev. 2001; 81: 807 -869. [PubMed] .
- 8. Zhu X, Rottkamp CA, Hartzler A, Sun Z, Takeda A, Boux H, Shimohama S, Perry G, Smith MA. Activation of MKK6, an upstream activator of p38, in Alzheimer's disease. J. Neurochem. 2001; 79: 311e318 [PubMed] .
- 9. Sheikh-Hamad D and Gustin MC. MAP kinases and the adaptive response to hypertonicity: functional preservation from yeast to mammals. Am J Physiol Renal Physiol. 2004; 287: F1102 -10. [PubMed] .
- 10. Cuadrado A and Nebreda AR. Mechanisms and functions of p38 MAPK signalling. Biochem J. 2010; 429: 403 -417. [PubMed] .
- 11. Hohmann S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol. Mol. Biol. Rev.. 2002; 66: 300 -372. [PubMed] .
- 12. Chen RE and Thorner J. Function and regulation in MAPK signaling pathways: lessons learned from the yeast Saccharomyces cerevisiae. Biochim. Biophys. Acta. 2007; 1773: 1311 -1340. [PubMed] .
- 13. Clotet J and Posas F. Control of cell cycle in response to osmostress: lessons from yeast. Methods Enzymol. 2007; 428: 63 -76. [PubMed] .
- 14. de Nadal E and Posas F. Multilayered control of gene expression by stress-activated protein kinases. EMBO J. 2010; 29: 4 -13. [PubMed] .
- 15. deNadal E, Ammerer G, Posas F. Controlling gene expression in response to stress. Nat Rev Genet. 2011; 12: 833 -845. [PubMed] .
- 16. Maeda T, Wurgler-Murphy SM, Saito H. A two-component system that regulates an osmosensing MAP kinase cascade in yeast. Nature. 1994; 369: 242 -245. [PubMed] .
- 17. Xia Z, Dickens M, Raingeaud J, Davis RJ, Greenberg ME. Opposing effects of ERK and JNK-p38 MAP kinases on apoptosis. Science. 1995; 270: 1326 -1331. [PubMed] .
- 18. Wang X, McGowan CH, Zhao M, He L, Downey JS, Fearns C, Wang Y, Huang S, Han J. Involvement of the MKK6-p38gamma cascade in gamma-radiation- induced cell cycle arrest. Mol Cell Biol. 2000; 20: 4543 -4552. [PubMed] .
- 19. Medvedik O, Lamming DW, Kim KD, Sinclair DA. MSN2 and MSN4 link calorie restriction and TOR to sirtuin-mediated lifespan extension in Saccharomyces cerevisiae. PLoS Biol. 2007; 5: e261 [PubMed] .
- 20. Wen J, You KR, Lee SY, Song CH, Kim DG. Oxidative stress-mediatedapoptosis. The anticancer effect of the sesquiterpene lactone parthenolide. J Biol Chem. 2002; 277: 38954 -38964. [PubMed] .
- 21. Zarubin T and Han J. Activation and signaling of the p38 MAP kinase pathway. Cell Research. 2005; 15: 11 -18. [PubMed] .
- 22. Grethe S, Ares MP, Anderson T, Porn-Ares MI. p38 MAPK mediates TNF-inducedapoptosisin endothelial cells via phosphorylation and downregulation of Bcl-x(L). Exp Cell Res. 2004; 298: 632 -642. [PubMed] .
- 23. Perfettini JL, Castedo M, Nardacci R, Ciccosanti F, Boya P, Roumier T, Larochette N, Piacentini M, Kroemer G. Essential role of p53 phosphorylation by p38 MAPK in apoptosis induction by the HIV-1 envelope. J Exp Med. 2005; Jan 17 201: 279 -289. [PubMed] .
- 24. Graves JD, Draves KE, Craxton A, Saklatvala J, Krebs EG, Clark EA. Involvement of stress-activated protein kinase and p38 mitogen-activated protein kinase in mIgM-induced apoptosis of human B lymphocytes. Proc Natl Acad Sci U S A. 1996; 93: 13814 -13818. [PubMed] .
- 25. Choi WS, Eom DS, Han BS, Kim WK, Han BH, Choi EJ, Oh TH, Markelonis GJ, Cho JW, Oh YJ. Phosphorylation of p38 MAPK induced by oxidative stress is linked to activation of both caspase-8- and -9-mediated apoptotic pathways in dopaminergic neurons. J Biol Chem. 2004; 279: 20451 -20460. [PubMed] .
- 26. Matos TJ, Duarte CB, Gonçalo M, Lopes MC. Role of oxidative stress in ERK and p38 MAPK activation induced by the chemical sensitizer DNFB in a fetal skin dendritic cell line. Immunol Cell Biol. 2005; 83: 607 -614. [PubMed] .
- 27. Vaziri H, Dessain SK, Eaton E, Imai SI, Frye RA, Pandita TK, Guarente L, Weinberg RA. hSIR2(SIRT1) functions as an NAD-dependent p53 deacetylase. Cell. 2001; 107: 149 -159. [PubMed] .
- 28. Van Meter M, Mao Z, Gorbunova V, Seluanov A. SIRT6 overexpression induces massiveapoptosisin cancer cells but not in normal cells. Cell cycle. 2011; 10: 3153 -3158. [PubMed] .
- 29. Kabra N, Li Z, Chen L, Li B, Zhang X, Wang C, Yeatman T, Coppola D, Chen J. SirT1 is an inhibitor of proliferation and tumor formation in colon cancer. J Biol Chem. 2009; 284: 18210 -18217. [PubMed] .
- 30. Burnett C, Valentini S, Cabreiro F, Goss M, Somogyvári M, Piper MD, Hoddinott M, Sutphin GL, Leko V, McElwee JJ, Vazquez-Manrique RP, Orfila AM, et al. Absence of effects of Sir2 overexpression on lifespan in C. elegans and Drosophila. Nature. 2011; 477: 482 -485. [PubMed] .
- 31. Sinclair DA and Guarente L. Extrachromosomal rDNA circles–a cause of aging in yeast. Cell. 1997; 91: 1033 -1042. [PubMed] .
- 32. Steinkraus KA, Kaeberlein M, Kennedy BK. Replicative aging in yeast: the means to the end. Annu. Rev. Cell Dev. Biol. 2008; 24: 29 -54. [PubMed] .
- 33. Aguilaniu H, Gustafsson L, Rigoulet M, Nystrom T. Asymmetric inheritance of oxidatively damaged proteins during cytokinesis. Science. 2003; 299: 1751 -1753. [PubMed] .
- 34. Erjavec N and Nystrom T. Sir2p-dependent protein segregation gives rise to a superior reactive oxygen species management in the progeny of Saccharomyces cerevisiae. Proc. Natl Acad. Sci. USA. 2007; 104: 10877 -10881. [PubMed] .
- 35. Knorre DA, Kulemzina IA, Sorokin MI, Kochmak SA, Bocharova NA, Sokolov SS, Severin FF. Sir2-dependent daughter-to-mother transport of the damaged proteins in yeast is required to prevent high stress sensitivity of the daughters. Cell Cycle. 2010; Nov 15 9: 4501 -4505. [PubMed] .
- 36. Straight AF, Shou W, Dowd GJ, Turck CW, Deshaies RJ, Johnson AD, Moazed D. Net1, a Sir2-associated nucleolar protein required for rDNA silencing and nucleolar integrity. Cell. 1999; 97: 245 -56. [PubMed] .
- 37. Aparicio OM, Billington BL, Gottschling DE. Modifiers of position effect are shared between telomeric and silent mating-type loci in S. cerevisiae. Cell. 1991; 66: 1279 -87. [PubMed] .
- 38. Herranz D and Serrano M. SIRT1: recent lessons from mouse models. Nat Rev Cancer. 2010; 10: 819 -23. [PubMed] .
- 39. Carmona-Gutierrez D, Eisenberg T, Büttner S, Meisinger C, Kroemer G, Madeo F. Apoptosis in yeast: triggers, pathways, subroutines. Cell Death Differ. 2010; 17: 763 -73. [PubMed] .
- 40. Fabrizio P, Battistella L, Vardavas R, Gattazzo C, Liou LL, Diaspro A, Dossen JW, Gralla EB, Longo VD. Superoxide is a mediator of an altruistic aging program in Saccharomyces cerevisiae. J Cell Biol. 2004; 166: 1055 -67. [PubMed] .
- 41. Bauer JH, Morris SN, Chang C, Flatt T, Wood JG, Helfand SL. dSir2 and Dmp53 interact to mediate aspects of CR-dependent lifespan extension in D. melanogaster. Aging (Albany NY). 2009; 1: 38 -48. [PubMed] .
- 42. Donehower LA. Longevity regulation in flies: a role for p53. Aging. 2009; 1: 6 -8. [PubMed] .
- 43. Parashar V and Rogina B. dSir2 mediates the increased spontaneous physical activity in flies on calorie restriction. Aging. 2009; 1: 529 -41. [PubMed] .
- 44. Bauer JH and Helfand SL. Sir2 and longevity: the p53 connection. Cell Cycle. 2009; 8: 1821 [PubMed] .
- 45. Tissenbaum HA and Guarente L. Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans. Nature. 2001; 410: 227 -30. [PubMed] .
- 46. Kaeberlein M, Kirkland KT, Fields S, Kennedy BK. Sir2-independent life span extension by calorie restriction in yeast. PLoS Biol. 2004; 2: E296 [PubMed] .