



Introduction
Dietary restriction (DR) is the best and nongenetic anti-aging strategy without malnutrition, aiming to intervene on environmental factors and reduce the risk of many age-related diseases [1]. DR has been shown to extend lifespan and improve health with the most consistent non-pharmacological intervention during aging in diverse organisms, such as flies [2], yeast [3], nematodes [4] and rodents [5]. These universal effects suggest that there should be some conserved common potential genetic pathways and biochemical processes by virtue of which different organisms delay aging in response to DR [2]. Extensive studies have been devoted to discover such underlying mechanisms of longevity assurance in DR of model organisms including primates (even human) [6]. Previous studies involving whole-genome gene expression analysis have shown that a number of genes and biological pathways differ between organisms fed on DR and normal-diet [2,7]. A large number of differentially expressed genes (DEGs) are implicated in regulatory mechanism of DR and molecular pathways, such as lipid metabolism, fatty acid metabolism, immune response, and oxidative phosphorylation are identified as being highly responsive to changes in diet composition [8]. Therefore, coding mRNA genes and signaling pathways involved in DR condition have been extensively studied [9]. The long non-coding RNA (lncRNA) is the key regulatory factor which also participates in the modulation of cell senescence and aging-related disease [10]. Pathologies implicating lncRNAs function include cancer [11], neurodegenerative disorders [12], cardiovascular pathologies [13] and metabolic diseases [14]. Some evidences supporting the role of lncRNAs in age-associated molecular processes are emerging recently, for instance, lncRNAs TERC and TERRA are found to be involved in chromosomal instability and short telomeres, which may contribute to premature senescence and aging [15]. While lncRNAs have significant functions and effect on the course of aging, the number of lncRNAs and their functional implications in anti-aging process are not yet well validated during nutritional interventions especially during DR.
RNA-seq techniques have driven the lncRNAs research in the recent years yielding significant insights in various model species [16]. As the classic model organism, RNA-seq data of coding and non-coding RNA, at different developmental stages, tissue samples, and whole-fly samples treated with environmental perturbations of D. melanogaster have been reported [17,18]. Interestingly fruit-fly being the ideal model organism for aging related studies can be used to understand the role of lncRNAs, which have not yet been explored for their role in aging. We analyzed RNA-seq data including mRNA and lncRNAs, from a set of DR and fully fed samples at 7 day and 42 day, respectively. Present data provides a deep understanding of the dynamic expression of lncRNAs in D. melanogaster transcriptome under DR and fully fed conditions. Our analysis revealed a large number of differentially expressed lncRNAs transcript isoforms and their target genes during DR that spanned most of the genome and can provide valuable insights into aging and age-related disease research field.
Results
Dietary restriction extends lifespan
Similar to our previous report [19], the fruit fly females, on DR food had median lifespans extended by 21.88% in comparison to fully fed flies in the present study (Fig. 1a). Fecundity of fruit flies in DR group became lower than in fully fed group (P<0.01, Fig. 1b). Flies at day 7 have been considered as young flies in many previous reports [20,21] and the flies in the present study began to die at day 42. Thus, 7 day and 42 day are the most appropriate time points for carrying out aging specific analysis involving expression patterns of transcriptional dynamics through lncRNA analysis.
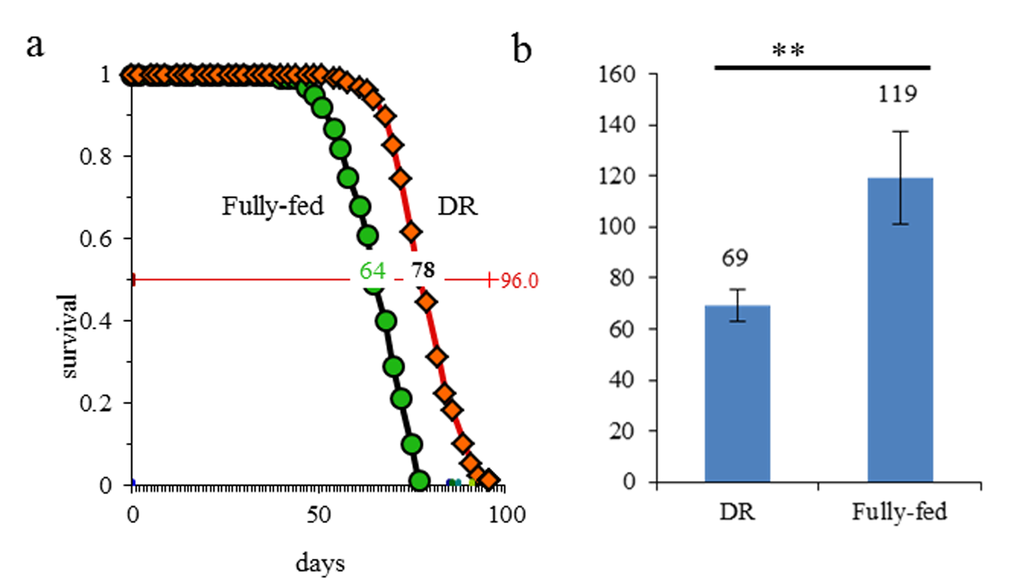
Figure 1. DR extended the lifespan and lowered the egg production in fruit flies. (a) curve of lifespan; (b) the number of eggs produced; **, p≤0.01.
Transcriptome profiles of DR and fully fed flies
A total of 86.78 GB of raw sequence data was produced yielding 7.26 billion strand-specific raw reads (Table S1). Value of Q20, Q30 and GC content were not significantly different among samples. After quality check, 95.79% of clean reads were obtained and were aligned to the D. melanogaster genome from FlyBase (Dmel_Release_6, http://FlyBase.org/), assigning 87.7% reads to the mRNA sequences (Fig. 2a). The read density (log2) was distributed mainly in 2L, 2R, 3L, 3R and X chromosomes (Fig. 2b). FPKM (fragments per kilobase of transcript sequence per millions base pairs sequenced) distribution was not significantly different in expression levels between lncRNAs and mRNA in DR and fully fed flies (P≤0.05, Fig. 2c).
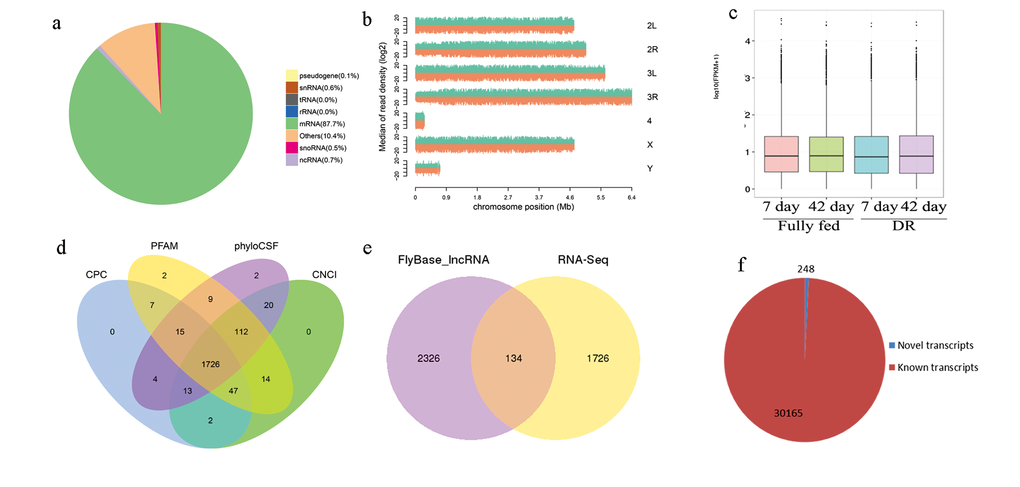
Figure 2. Overall expression profile in RNA-seq. (a) mapping to fruit fly genome; (b) reads density in chromosomes; (c) FPKM distribution of mRNA and lncRNA; (d) 1726 novel lncRNAs were obtained after four programs filtering; e, when compared to 2460 known lncRNAs sequences in FlyBase (Dmel_Release_6), 134 non-redundant known lncRNAs and 1726 non-redundant novel lncRNAs were found; f, When compared to known mRNA sequences in FlyBase, 30165 known and 248 novel transcripts were found.
In total, 3826 redundant lncRNA transcripts and 30413 redundant mRNA transcripts were identified in this study. After BLASTing with FlyBase lncRNA (Dmel_Release_6), these 3826 transcripts corresponded to a total of 134 non-redundant known lncRNAs (Dataset S1) and 1726 non-redundant novel lncRNA (Dataset S2, Fig. 2d and 2e). In those novel lncRNAs, the number of large intergenic noncoding RNA, intronic transcript lncRNAs and antisense lncRNAs were 821, 695 and 210, respectively. A total of 248 non-redundant novel mRNA transcripts (Dataset S3, Fig. 2f) were recognized when 30413 mRNA transcripts were BLASTed with D. melangaster genome from FlyBase, and the remaining 30165 transcripts aligned to 13917 genes.
Differentially expressed genes analysis
Differentially expressed genes (DEGs) had been analyzed between four comparative groups (Dataset S4), including 7 day DR vs 7 day fully fed, 42 day DR vs 42 day fully fed, 7 day DR vs 42 day DR, 7 day fully fed vs 42 day fully fed. In total, 102 differentially expressed lncRNAs (25 known and 77 novels) and 1406 coding genes were identified between these four comparative groups. A total of 719 differentially expressed transcripts were the most abundant as measured at 7 day between DR and fully fed group, including 30 lncRNAs and 689 coding mRNA genes. Also 39 differentially expressed lncRNAs and 571 differentially expressed coding genes were present at 42 day of DR when compared to fully fed group (Fig. 3a). Out of those differentially expressed transcripts, five common differentially expressed lncRNAs and 204 common coding genes appeared at both 7 day and 42 day between DR and fully fed groups (Fig. 3b, 3c). Among five differentially expressed lncRNAs, CR31781 and XLOC_014901 were up-regulated while XLOC_189941 was down-regulated in DR when compared to fully fed group at both time frames. Another two lncRNAs (XLOC_189909 and XLOC_228709) were down-regulated at 7 day and up-regulated at 42 day in DR when compared to fully fed group.
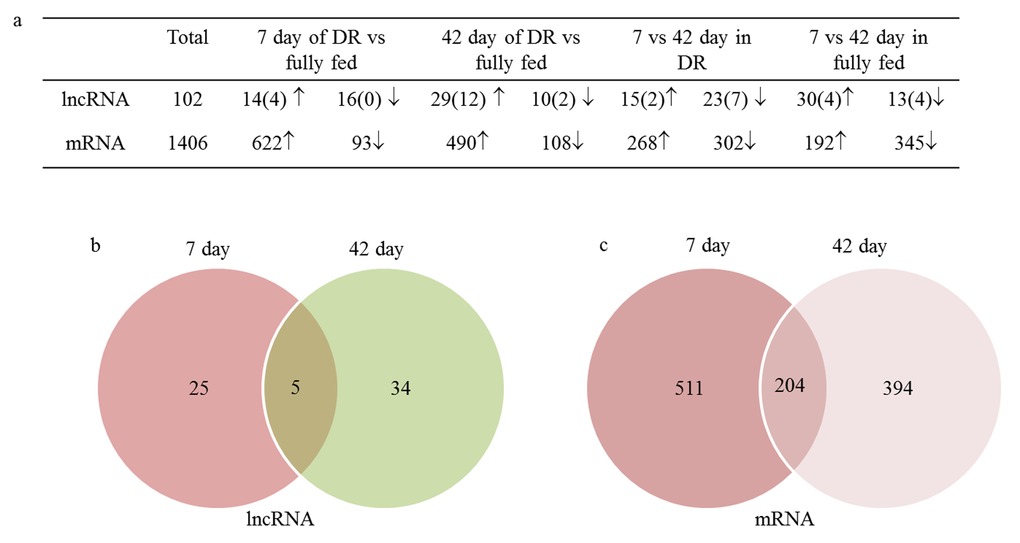
Figure 3. 102 differentially expressed lncRNA and 1406 differentially expressed coding genes were identified in this study. (a) The number of DEGs in different comparative groups (parentheses presented the number of known lncRNA); (b) Common differentially expressed lncRNA in both 7 day and 42 day between DR and fully fed flies; (c) Common differentially expressed mRNA in both 7 day and 42 day between DR and fully fed flies; ↑, upregulation; ↓, downregulation.
In order to validate the RNA-seq analysis finding, we randomly selected 28 differentially expressed lncRNAs and 61 mRNA coding genes, and analyzed their expression using quantitative real-time PCR (qPCR) analysis. All the genes were having expression levels in concordance with the RNA-seq results in different comparison groups with varying levels of significance (P≤0.05) (Dataset S5).
Known lncRNA
When compared to the known lncRNAs existing in FlyBase (Dmel_Release_6), 134 known lncRNAs in our study were detected (Dataset S1). Of 134 annotated lncRNAs, 25 differentially expressed lncRNAs in four different comparisons (7 day DR vs 7 day fully fed, 42 day DR vs 42 day fully fed, 7 day DR vs 42 day DR, 7 day fully fed vs 42 day fully fed). Furthermore, 4 and 14 known lncRNAs displayed differential expression at the 7 day and 42 day when compared between DR and fully fed groups, respectively. Moreover, out of a total of 134 annotated lncRNAs only 11 lncRNAs obtained 22 annotations from GO enrichment. CR31781 was the only one differentially expressed lncRNA which got GO annotation and was further validated by qPCR result. A total of 15 out of 22 annotations belonged to lncRNA CR31781, such as lateral inhibition, cell-cell signaling involved in cell fate commitment, extracellular space, extracellular region part and cellular component, etc.
Novel lncRNA
We predicted the targets of novel lncRNAs, and then analyzed the GO (Dataset S6) and KEGG enrichment (Dataset S7) of targets (Dataset S8 and Dataset S9). In most GO enrichment annotations, 17 non-redundant differentially expressed mRNA genes were the targets of 13 differentially expressed lncRNAs (Table 1). Six differentially expressed coding genes were the target of six differentially expressed lncRNAs and expressed during both 7 day and 42 day in DR when compared to fully fed fruit flies, and were found to be enriched in carboxypeptidase activity, cellular component morphogenesis, heme binding, iron ion binding and serine-type endopeptidase activity. In all the comparison groups, nine differentially expressed target genes related to fruit fly aging, which has resulted in lethal (including CG9733, SPH93, Hsc70-1, Cyp4p3, IscU homolog genes) and long lived (including Socs36E genes) phenotypes under RNAi system.
Table 1. GO annotations of differentially expressed lncRNA and their targets in DR group when compared to fully fed group
| lncRNA(action mode, corrected p-value) | Targets(corrected p-value) | GO annotations | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7 day DR vs 7 day fully fed | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_052958(cis, 0.00715)↓ | CG12374(0.00005)↑ | Carboxypeptidase activity | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_166557(trans, 0.00425)↓ | globin 1(0.0214)↑ | Heme binding/Iron ion binding | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_009418(trans, 0.0322)↑ | CG6048(0.0496)↑ | Serine-type endopeptidase activity | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 42 day DR vs 42 day fully fed | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_001163(trans, 0.0171)↑ | Hsc70-1(0.0045)↑ | Cellular component morphogenesis | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_196039(trans, 0.0028)↑ | Cyp28d2(0.00835)↑ | Heme binding | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_009798(trans, 0.01205)↑ | IscU homolog(0.00395)↑ | Iron ion binding | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7 day DR vs 42 day DR | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_002137(trans, 0.02335)↓ | Hsc70-3(0.0097)↓ | Cell morphogenesis/Cellular component morphogenesis | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG9733(0.00005)↓ | Serine-type endopeptidase activity | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_076307(trans, 0.03315)↓ | Hsp70Bc(0.00005)↓ | Cell morphogenesis/Cellular component morphogenesis/Glycosaminoglycan catabolic process/Glycosaminoglycan metabolic process/Peptidoglycan catabolic process/Peptidoglycan metabolic process/Serine-type endopeptidase activity | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Socs36E(0.00005)↓ | Cell morphogenesis/Cellular component morphogenesis | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CG10041(0.0489)↓ | Serine-type endopeptidase activity | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_000246(trans, 0.01425)↓ | CG15046(0.0405)↓ | Serine-type endopeptidase activity | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_071173(trans, 0.0129)↓ | SPH93(0.00005) ↓ | Serine-type endopeptidase activity | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_066439(cis, 0.0113)↓ | Cyp6w1(0.0103)↓ | Heme binding/Iron ion binding | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_009798(trans, 0.0034)↓ | Cyp4p3(0.01165)↓ CG10041(0.0489)↓ IscU homolog(0.0115)↓ | Heme binding/Iron ion binding Serine-type endopeptidase activity Iron ion binding | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7 day fully fed vs 42 day fully fed | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_052958(cis, 0.0087)↑ | CG12374(0.0001)↑ | Carboxypeptidase activity | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_067962(cis, 0.0097)↑ | PGRP-SC1a(0.00005)↑ | Carboxypeptidase activity/ Glycosaminoglycan catabolic process/ Glycosaminoglycan metabolic process/Peptidoglycan catabolic process | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_151622(cis, 0.00635)↑ | CG33970(0.0346)↓ | Iron ion binding | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Note: ↑, up-regulation; ↓, down-regulation. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
In most significant KEGG pathway enrichment (Dataset S7), 27 differentially expressed lncRNAs and 44 differentially expressed targets were found to be involved in 38 pathways out of which 21 targets were related to fly aging. Most of these were important pathways affecting senescence, such as mTOR, FoxO and Wnt signaling pathway. At 7 day and 42 day, 11 of the 26 genes in the mTOR pathway were present in the RNA-seq data, but, just inac gene was upregulated as the target of lncRNA XLOC_056059 at the 42 day in DR in comparison to 42 day in fully fed. Five lncRNAs and three targets existed in FoxO signaling pathway, and were found upregulated at 42 day in DR when compared to fully fed flies at same day (Table 2). Especially, target genes involved in signaling pathways of biosynthesis of unsaturated fatty acids and fatty acid metabolism were up-regulated in DR group in comparison to fully fed, including CG9743 (as the target of XLOC_201255), CG15531, CG9743, CG15531 and FASN2 at the 7 day, Acsl and CG3961 at the 42 day.
Table 2. Novle lncRNAs and mRNA genes involved in five KEGG pathways during DR
| KEGG pathway | lncRNA (comparable group and correct pvalue, action mode) | mRNA genes (comparable group and correct pvalue) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FoxO signaling pathway | XLOC_000246(A2↑-B2 0.044, A1↓-A2 0.01425, trans) XLOC_076307(A1↑-A2 0.03315, trans) | Gadd45 (A2↑-B2 0.01075,A1↓-A2 0.014) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_000043(A2↑-B2 0.0136, A1↓-A2 0.01345, trans) | Pepck(A2↑-B2 0.00005, A1↓-A2 0.00005) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_221562(A2↑-B2 0.00275, trans) XLOC_001163(A2↑-B2 0.0171, trans) | Egfr(A2↑-B2 0.0204) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Hippo signaling pathway-fly | XLOC_002137(A1↓-A2 0.02335, trans) | Upd2(A1↑-A2 0.0043) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_002137(A1↓-A2 0.02335, trans) | Upd3(A1↓-A2 0.01585) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_186922(A1↓-B1 0.00785, cis) | Zyx(A1↓-B1 0.046) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_000246(A1↓-A2 0.01425, trans) XLOC_000236(A1↓-A2 0.00555, trans) XLOC_009798(A1↓-A2 0.0034, trans) XLOC_000043(A1↓-A2 0.01345, trans) | baz (A1↓-A2 0.03595) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_160411(B1↓-B2 0.0048, trans) | wg(B1↓-B2 0.00905) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Phototransduction-fly | XLOC_092363(A1↓-B1 0.00885, B1↑-B2 0.0075,cis) | Arr2(A1↑-B1 0.00315, B1↑-B2 0.0481) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Protein processing in endoplasmic reticulum | XLOC_076307(A1↓-A2 0.03315, trans) | Hsp70Bc(A1↓-A2 0.00005) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_001163(A2↑-B2 0.0171, trans) | Hsc70-1(A2↑-B2 0.0045) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_072226(A1↓-B1 0.00885, trans) | CG8974(A1↑-B1 0.01995) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_002137(A1↓-A2 0.02335, trans) | Hsc70-3(A1↓-A2 0.02335) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_000043(A2↑-B2 0.0136, A1↓-A2 0.01345, trans) XLOC_009798(A2↑-B2 0.01205, A1↓-A2 0.0034, trans) XLOC_000236(A1↓-A2 0.00555, A2↑-B2 0.00955, trans) | l(2)efl(A1↓-A2 0.0108, A2↓-B2 0.01715) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Metabolism of xenobiotics by cytochrome P450 | XLOC_002137(A1↓-A2 0.02335, trans) | GstD5(A1↓-A2 0.00005) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_000246(A2↑-B2 0.044, A1↓-A2 0.01425, trans) XLOC_002137(A1↓-A2 0.02335, trans) | GstD10 (A2↑-B2 0.0138, A1↓-A2 0.00175) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_201255(A1↑-B1 0.00515, trans) | GstE12 (A1↑-B1 0.0495) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_010702(A1↑-A2 0.0023, trans) | CG17323(A1↑-A2 0.0108) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_002137(A1↓-A2 0.02335, trans) | GstE7 (A1↓-A2 0.00005), CG5999(A1↓-A2 0.00135) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_106174(A1↑-B1 0.02315, trans) | Ugt35b (A1↑-B1 0.00015) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Glutathione metabolism | XLOC_002137(A1↓-A2 0.02335, trans) | GstD10(A1↓-A2 0.00175), GstD5(A1↓-A2 0.00005), GstE7(A1↓-A2 0.00005) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| XLOC_201255(A1↑-B1, trans) | GstE12(A1↑-B1 0.0495) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Note: A1, 7 day in DR; A2, 42 day in DR; B1, 7 day in fully fed; B2, 42 day in fully fed; ↑, up-regulation; ↓, down-regulation. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
In the present study, we have characterized three novel signaling pathways that may be related to aging, which are, hippo signaling pathway-fly, phototransduction-fly and protein processing in endoplasmic reticulum (Table 2). In hippo signaling pathway-fly 11 differentially expressed coding genes were involved and, three and five genes displayed differential expression at the 7 day and 42 day in DR when compared to fully fed flies, respectively, and five coding genes were the targets of seven novel lncRNAs. In phototransduction-fly signaling pathway eight related coding genes were detected and showed differential expression between various comparison groups in DR and fully fed condition. Novel lncRNA XLOC_092363 and respective target Arr2 were involved on phototransduction-fly signaling pathway. Furthermore, in endoplasmic reticulum pathway 15 differentially expressed coding genes were found in protein processing and 11 of these were related to aging and five genes were the targets of 7 novel lncRNAs. Interestingly enough, 9 of 15 differentially expressed coding genes belonged to heat shock protein gene families.
Besides, 17 DEGs and 14 DEGs presented in metabolism of xenobiotics by cytochrome P450 and glutathione metabolism pathway, respectively, which were up-regulated in various comparison groups in DR when compared to fully fed condition (Table 2). Out of these, 10 of 17 DEGs and 11 of 14 DEGs belonged to glutathione S transferase, and there are some common glutathione S transferase genes between both pathways such as GstD5, GstD10, GstD6, GstE1, GstE2, GstE3, GstE4, GstE7 and GstE12. Meanwhile seven targets of five novel lncRNAs and four targets of two novel lncRNAs were found in cytochrome P450 and glutathione metabolism pathway, respectively.
Regulatory elements of lncRNA
The promoter positions in genome of fruit fly were first confirmed according to Drosophila Core Promoter Database, followed by analysis of lncRNA located in 10kb range (90% identification accuracy and 90% coverage, Dataset S10). Forty four novels and one known lncRNA in our study were found in 10 kb range of 38 promoters. Among 45 lncRNAs, 10 lncRNAs and 12 promoters were present at X chromosome, while one lncRNA and one promoter were present at 2L chromosome. Furthermore, 4 lncRNAs and 4 promoters had the overlapping zones, including XLOC_067699 (promoter lcp4, overlap 92bp), XLOC_112986 (promoter primase, overlap 92bp), XLOC_113215 (promoter s15, overlap 92bp) and XLOC_220979 (promoter G6pd, overlap 94bp).
Interestingly enough, two novel lncRNAs (XLOC_092363 and XLOC_166557) differentially expressed at the 7 day in DR group when compared to fully fed group. lncRNA XLOC_092363 was located in promoter hairy in the up-stream of sense strand of 3L chromosome. Five differentially expressed targets of XLOC_092363 were also located in 3L chromosome through cis-acting mode (Fig. 4a). It’s noteworthy that hairy gene as associated promoter and as target of XLOC_092363, differentially expressed at 7 day and 42 day of DR in comparison to fully fed flies. Target arr2 has the same tendency of being differentially expressed with promoter hairy gene. Other three targets Cp16, Cp18 and Cp19 differentially expressed at the 7 day in DR when compared to fully fed group, and their molecular function was being the structural constituent of chorion. In addition, promoter ems sited in the down-stream of XLOC_166557 in sense strand of 3R chromosome is involved in heme binding and iron ion binding of GO annotations (Fig. 4a). The target of XLOC_166557 was globin 1 by cis-acting mode, and XLOC_166557 was found to be down-regulated while globin 1 was up-regulated in DR when compared to fully fed flies. The qPCR results were consistent with RNA-seq data up to certain extent (Dataset S3), and revealed differential expression between DR and fully fed flies (Fig. 4b).
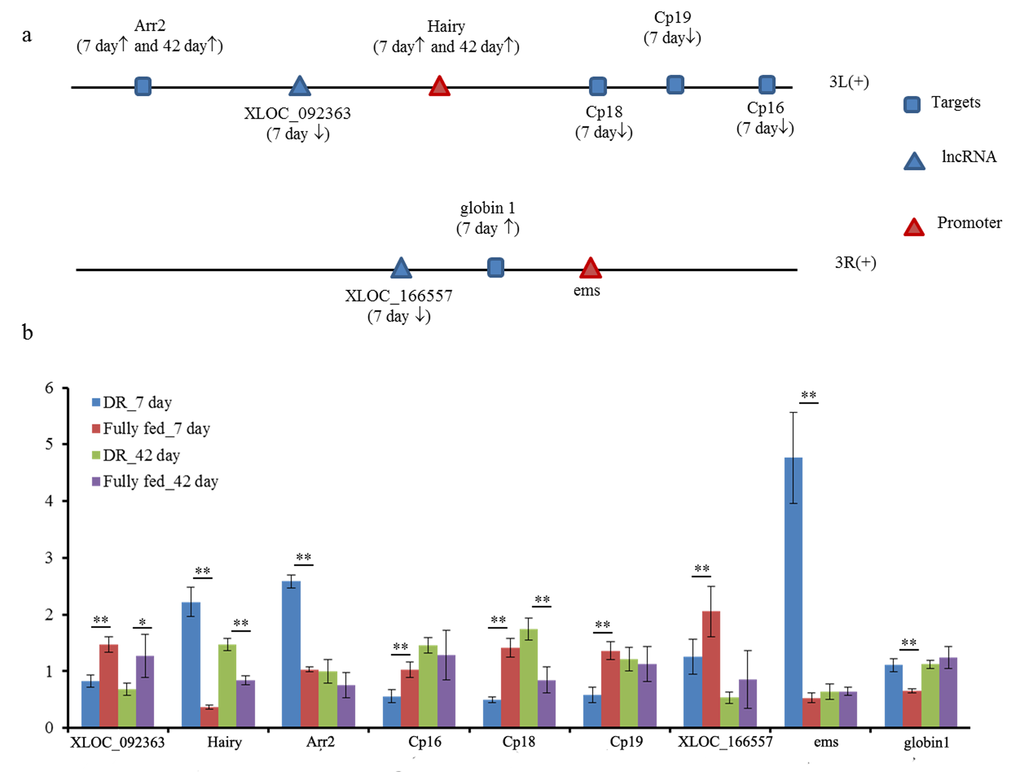
Figure 4. Two promoters associated lncRNA XLOC_092363 and XLOC_166557 in 10 kb range. (a) the positional relationship of two differentially expressed lncRNA and targets in chromosome of fruit fly (↑, up-regulated in DR when compared to fully fed group in RNA-seq data; ↓, down-regulated in DR when compared to fully fed group in RNA-seq profile); (b) expression level of two promoters, lncRNA and their targets between DR and fully fed flies by qPCR. **, P≤0.01; *, P≤0.05.
Tissue specificity of lncRNA
Differentially expressed lncRNA and mRNA in this study were BLASTed with the transcripts from D. melanogaster from cultured cell lines, dissected organ systems and under environmental perturbations and 5 known lncRNAs from the present study showed the tissue specificity (Dataset S11). In five lncRNAs, CR31781 and target CG31810 (cis-acting mode) were upregulated in DR group when compared to fully-fed group, which were mainly expressed in head, carcass, digestive system, central nervous system and under environmental perturbations (including caffeine, paraquat, Cd, Zn, Cu) in fruit fly. Another target CG13284 was seen upregulated at the 42 day when compared between DR and fully-fed groups. The molecular function of CG31810 and CG13284 is described with steroid dehydrogenase activity. Besides, lncRNA CR43641 (up-regulated at the 7 day when compared to fully fed group) along with the differentially expressed targets Dip-B, tal-AA and CG34383 (cis-acting mode), were highly expressed in fat or fatbody. The remaining lncRNA (CR43148, CR40469 and CR44095) and their differentially expressed targets did not have the obviously same tissue specificity.
A total of 20 novel differentially expressed lncRNAs at 7 day under DR condition were randomly selected for analysis in fly head, gut, and fat body using qPCR to determine the tissue specificity (Fig. 5a, primers in Table S2). The results showed that lncRNA XLOC_066439 were at significantly higher expression level in gut compared to head and fat body (P≤0.05). Also the expression levels of XLOC_076307 and XLOC_009418 in head were significantly higher than that of gut and fat body (P≤0.05). XLOC_166557 was mainly expressed in fatbody of fruit flies. Tissue specificity of several lncRNA was consistent with their targets (Table S3), such as XLOC_076307 (Socs36E, head), XLOC_166557 (globin 1, fatbody), XLOC_067962 (PGRP-SC1a, PGRP-SC2 and CG8740, gut and fatbody). On the other hand, 21 differentially expressed lncRNAs were identified in cytoplasm or nucleus of Schneider’s line 2 cells by qPCR (Fig. 5b). Among these lncRNA, XLOC_073604 was specially expressed in S2 cell’s nucleus. LncRNA XLOC_009798, XLOC_010702, XLOC_071213, XLOC_118356, XLOC_009418 and XLOC_076307 were particularly located in S2 cell’s cytoplasm. We also found that lncRNA XLOC_000071, XLOC_056059 and XLOC_196039 were expressed in S2 cell’s nucleus and cytoplasm.
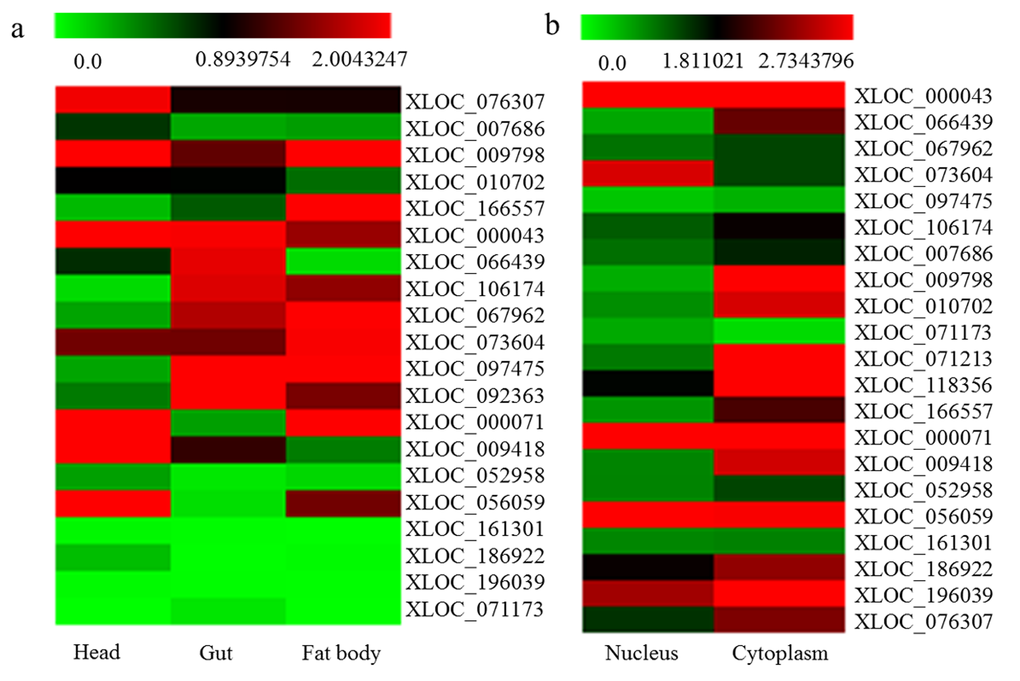
Figure 5. Tissue specificity of differentially expressed lncRNA. (a) the tissue specificity of 20 differentially expressed lncRNA at 7 day under DR condition had been analyzed in gut, head, and fat body of flies; (b) 21 differentially expressed lncRNA whether located in cytoplasm or nucleus of Schneider’s line 2 cell were detected.
Inhibition of a novel lncRNA expression in fly S2 cell
A novel lncRNA, targets among other few, the Gadd45 gene which is involved in FoxO signaling pathway (Fig. 6). In order to understand the role of this novel lncRNA XLOC_076307, we silenced it in fruit fly S2 cells by small interfering RNA (siRNA). The results showed that the expression level of XLOC_076307 and its target Hsp70Bc decreased significantly (P≤0.01), while targets Gadd45 and Socs36E showed significant increases after 72h of transfection, confirming the association between lncRNA and its predicted targets through trans action model.
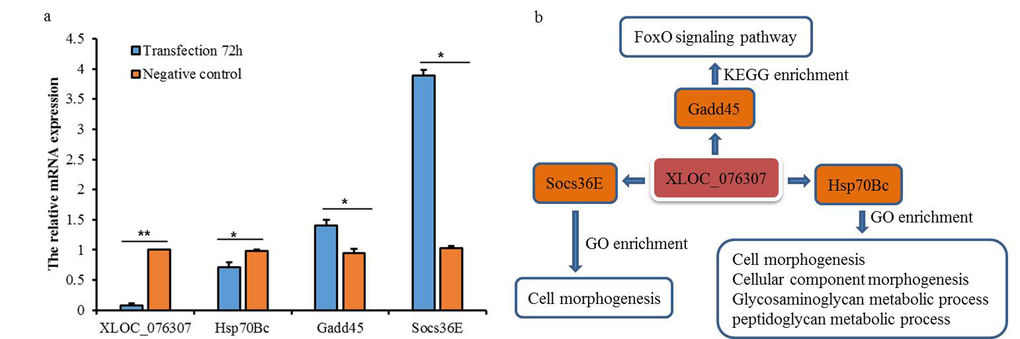
Figure 6. Expression levels of three targets after 72h of transfection XLOC_076307 siRNA and function enrichment. (a) expression level of lncRNA XLOC_076307 was significantly down regulated with target Hsp70Bc down-regulation and Gadd45 and Socs36E up-regulation; (b) function enrichment (including GO and KEGG) of XLOC_076307 and its targets.
Discussion
Long non-coding RNAs make up the majority of the mammalian transcriptome, many of which are emerging as regulators of genome expression/stability and modulators of cell senescence [22,23]. Previous studies have shown that lncRNAs are involved in aging process and age-related diseases [24,25]. Although DR is the best-studied intervention known to delay aging and extend lifespan in evolutionarily distant organisms ranging from fruit flies to mammals in the laboratory, the molecular mechanisms of DR are still elusive. In our study, we analyzed lncRNA and mRNA gene expression patterns of transcriptional dynamics in fruit flies at the 7 day and 42 day during DR and fully-fed conditions, respectively, and identified the differentially expressed lncRNAs and their target genes and revealed their possible regulatory roles in DR process.
A large number of candidate lncRNAs have been identified in Drosophila during different developmental stages, dissected tissue samples, cultured cell lines and under environmental perturbations [17,25,26]. In this study, we found only 134 known lncRNAs when compared to 2460 annotated lncRNAs from FlyBase (Dmel_Release_6). The reason may be the technical differences between ours and previous studies. Also the lncRNAs at the time of DR have not been identified before and thus, the number of novel lncRNAs (1726) was far more than known lncRNA (134) related to DR, demonstrating that DR impact on lncRNAs remains largely unexplored.
LncRNA in known aging pathways
Upon comparison of aging-related signaling pathways with previously published data on fruit flies [26], we found differentially expressed coding genes as the targets of lncRNA which were enriched in signaling pathways, such as mTOR, FoxO and Wnt signaling pathway, biosynthesis of unsaturated fatty acids, and fatty acid metabolism. Especially in FoxO signaling pathway, three (Gadd45, Pepck, Egfr) of seven differentially expressed coding genes were found to be up-regulated at the 42 day of DR when compared to fully fed flies as the targets of novel lncRNA (XLOC_000246, XLOC_076307, XLOC_000043, XLOC_221562 and XLOC_001163). Gadd45 and Egfr had been reported to be involved in Drosophila senescence [27,28]. Adipose tissue is a major source of energy, which declines in old age [23,29]. In our study, DR activates the level of biosynthesis of unsaturated fatty acids and fatty acid metabolism and may rescue the declining stored fat with advancing age to extend healthspan of Drosophila flies. CG9743 (up-regulated at the 7 day of DR versus fully fed flies) as the target of novel differentially expressed lncRNA XLOC_201255 participates in lipid metabolic process [30], such as unsaturated fatty acids and fatty acid metabolism, suggesting a possible involvement in regulation of lipid metabolic process. In this study, 1 gene (inac gene) out of 11 mTOR pathway genes were found to be differentially expressed, and the remaining genes were not significantly different between any of the comparative groups. This result is consistent with what has been previously reported [2]. We presume that the effects of DR on the mTOR pathway genes may occur post-transcriptionally and could not be detected in RNA-seq data. From those results, DR alters the expressing profile of mRNA and lncRNA to activate of the conserved nutrient-signaling pathways.
lncRNA in potential novel aging pathways
Besides known aging-related signaling pathways, we also identified novel pathways which might be related to senescence in DR process. In present study, 11 DEGs were detected which were involved in Hippo pathway and nine of these had been demonstrated to be related to Drosophila aging, such as Upd2, Upd3, Zyx, Pp2A-29B, baz, wg, etc. The Hippo pathway is an evolutionarily conserved and pleiotropic mediator of growth control, cell fate decisions and stem cell identity, cell type specification and differentiation during development [31], and, Hippo pathway components also mediated crosstalk with other pathways, for example Wnt, TGF and BMP pathways [32], and also five targets of seven lncRNAs were present in Hippo pathway in the present study.
Phototransduction-fly was another interesting pathway, and had eight DEGs (27.57% of the total 28 coding genes) upregulated during DR when compared to fully fed group in present study. Transient receptor potential (trp) was the essential element of the transduction cascade [33], which was upregulated at the 7 day and 42 day during DR in comparison to fully fed flies in our study. Furthermore, the differentially expressed Arr2 and inaC genes were the targets of differentially expressed lncRNA XLOC_092363 and XLOC_056059, respectively. These results indicated DR to be the pleiotropic mediator of Drosophila aging for affecting various physiological processes. LncRNA may also take part in regulation of these similar pathways.
lncRNA and its relation with gene promoters
Previous studies have documented the important connection of lncRNA to promoter. Promoter region could be transcribed into lncRNA in quiescent cells to block DHFR gene expression through both direct interactions of the RNA and promoter-specific interference [34]. In our study, we found that 4 lncRNA and 4 promoters had the overlapping zones, which illustrates that these lncRNAs may be transcribed from corresponding promoters.
Another study showed lncRNA HSR1 (heat shock RNA-1) was required for HSF1 (heat-shock transcription factor 1) activation, and the activated HSF1 affect the promoter of related genes to open the transcription in mammalian cells [35]. In this study, we found two differentially expressed lncRNA had the position correlation with promoters. As promoter and target of lncRNA XLOC_092363, hairy gene up-regulation in DR had been proven to be a lethal factor after mutagen treatment [36]. Furthermore, Drosophila sirt1 is required for heterochromatic silencing and euchromatic hairy/E(Spl) bHLH repression in segmentation and sex determination [36], which is a key member of the sirtuin family of proteins and mammalian homolog of the yeast silent information regulator protein that influence lifespan in several model organisms [37]. XLOC_092363 and targets Cp16, Cp18 and Cp19 were down-regulated at 7 day of DR while promoter hairy was up-regulated. Cp16, Cp18 and Cp19 are involved in biological process of multicellular organismal development and eggshell chorion assembly [38]. We also observed a decrease in egg production in DR flies suggesting that promoter hairy may have special correlation with lncRNA XLOC_092363 and downstream targets to affect fecundity in DR flies.
Another lncRNA XLOC_166557 was located in upstream of promoter ems and the expression level of ems was up-regulated at the 7 day during DR when compared to fully fed flies. Its target globin1 located in downstream participated in biological process of oxygen transport [39]. In summary, these results demonstrate that differentially expressed lncRNA and promoters may play a role in DR for extending lifespan in Drosophila.
Tissue specificity of lncRNA
In this study, two differentially expressed annotated lncRNA (CR31781 and CR43641) and its targets have the consistent tissue specificity. CR31781 presented specific expression at the 42 day of DR when compared to fully fed flies, and its molecular function is lateral inhibition in cell fate determination [40]. CR31781 and two targets had the similar tissue specificity in carcass from mated female after 4 days [17]. The previous studies reported the variable expression level of CR31781 (presenting up-regulation) in myopathic lamin mutations and lifespan extension of flies over-expressing a small mitochondrial chaperone Hsp22 [41]. Furthermore, its potential targets CG31810 and CG13284 are involved in steroid metabolic process, had steroid dehydrogenase activity and upregulated in DR. CG13284 also participated in negative regulation of cell proliferation and phagocytosis with relevant senescence [42]. These observations suggest that annotated lncRNA CR31781 and its predicted targets may play an important role in DR, and the specific mechanism of action should be studied in the future research.
Another lncRNA CR43641 and targets Dip-B, tal-AA and CG34383 were highly expressed in Drosophila fat body, a major immune organ, that undergoes immunosenescence and mounts strong systemic inflammation leading to dysregulation of immune deficiency signaling in the midgut of old flies [43]. RNAi system of target CG34383 (Scer\GAL4pnr-MD237) had been a lethal factor before end of pupal stage [40] related to Drosophila aging. Target Dip-B participated in proteolysis process while tal-AA took part in biological process of actin filament organization and morphogenesis of an epithelium [44]. Based on these findings, we would speculate that lncRNA CR43641 and its predicted targets may have the positive function in enhancing the immune system and anti-aging in DR conditions.
lncRNA XLOC_076307 relationship with its targets
We have identified in the Schneider’s line 2 cells, the novel lncRNA XLOC_076307, which was hypothesized to possibly take part in cell morphogenesis, cellular component morphogenesis, glycosaminoglycan metabolic process, peptidoglycan metabolic process, spliceosome and FoxO signaling pathway according to its potential targets (Hsp70Bc, Socs36E and Gadd45), and was found mainly located in flies head and S2 cell cytoplasm. Loss of Hsp70 in Drosophila is pleiotropic, with effects on thermotolerance, recovery from heat shock, neurodegeneration and aging defects [45]. Also the target Hsp70Bc was related to heat shock-mediated polytene chromosome puffing [45]. Another target Socs36E played a critical role in a genetic circuit that establishes a boundary between the motile border cell cluster and its non-invasive epithelial neighbors through STAT attenuation [46]. Furthermore, phenotypes Socs36EScer\UAS.P\T.cCa could reduce death in some flies dying during pharate adult stage [46]. The over-expression of target Gadd45 in the nervous system of D.melanogaster increased the lifespan [47]. Expression level of three targets changed after 72h of transfection of XLOC_076307 siRNA in our study. Thus, we hypothesize that XLOC_076307 may play a role in Drosophila aging.
Materials and Methods
Sample collection and preparation
The wild-type stock (Dahomey) were reared at 25°C on a 12-hour on/off light cycle at 50% humidity. Female flies were sorted in two groups which were given DR and fully fed treatment [48]. Median lifespan was calculated as the mean life span of the longest surviving 50% of the population. Egg count was measured according to Emran et al [19]. Ten vials (100 females) of flies were randomly collected for RNA-seq at the 7 day and 42 day in experimental groups (including DR and fully fed group), respectively. A total of eight samples (two biological replicates at day 7 and 42 for each treatment group) were chosen for paired-end (PE read length of 100bp) sequencing using Hiseq2000.
RNA isolation, library preparation, sequencing and lncRNA prediction
Total RNA was extracted using TRIzol Reagent (Invitrogen, CA), and treated with DNase and purified on an RNAeasy column (Qiagen). The quality, quantity and integrity were measured using standard laboratory procedures. Subsequently, sequencing libraries were generated using the rRNA-depleted RNA by NEBNext® Ultra™ Directional RNA Library Prep Kit for Illumina® (New England Biolabs, USA) following manufacturer’s recommendations and library quality was assessed on the Agilent Bioanalyzer 2100 system. After cluster generation, the libraries were sequenced on an Illumina Hiseq 2000 platform (provided by Novogene Co. China) and 100 bp paired-end reads were generated. After removing low quality reads (QV\20) and trimming the adapters, paired-end clean reads were aligned to the fly reference genome using TopHat v2.0.9 [49].
Cuffcompare software [49] was used to combine the transcripts, which at least existed in two samples. The ≥200bp transcripts were selected and the number of extrons was ≥1. Those transcripts were BLASTed (evalue=1e-10) to FlyBase dataset. The mapped transcripts were directly described as known lncRNAs or mRNA genes. The remaining transcripts were BLASTed (evalue=1e-10) to non-mRNA transcripts, such as rRNA, tRNA, snRNA, snoRNA, pre-miRNA, pseudogenes. And, the similar or same transcripts were filtered out. Then, CNCI (Coding-Non-Coding-Index) (v2) profiles [50], CPC (Coding Potential Calculator) (0.9-r2) [51], Pfam Scan (v1.3) [52], and PhyloCSF (phylogenetic codon substitution frequency) (v20121028) [53] were performed to analyse the coding potential of transcripts Transcripts predicted with coding potential by either/all of the four tools mentioned above, were filtered out, and those without coding potential were our candidate set of novel lncRNAs. The non-coding transcripts were BLASTed to mRNA in FlyBase dataset to screen the lincRNA,intronic lncRNA, anti-sense lncRNA transcripts by Cuffcompare according to class_code (http://cufflinks.cbcb.umd.edu/manual.html#class_codes) [54] and the transcripts with coding potential were the novel mRNA transcripts.
Quantification of gene expression level
Cuffdiff (v2.1.1) [49] was used to calculate FPKMs (expected number of fragments per kilobase of transcript sequence per millions base pairs sequenced) of both lncRNAs and coding genes in each sample. Gene FPKMs were computed by summing up the FPKMs of transcripts in each gene group.
Target gene prediction
LncRNAs have regulatory roles in gene expression at both the transcriptional and post-transcriptional levels in diverse cellular contexts and biological processes. lncRNAs are responsible for nuclear structure integrity, and can regulate the expression of either nearby genes (acting in cis in the nucleus) or genes elsewhere in cells (acting in trans in the nucleus or cytoplasm) by interacting with proteins, RNAs, and DNAs [55,56].Cis acting lncRNAs are those acting on neighboring target genes [55]. We searched coding genes 10k/100k upstream and downstream of lncRNAs and then analyzed their function next. Trans acting targets were identified by expression levels of both lncRNAs and mRNA transcripts [56]. While there were eight samples, we calculated the expressed correlation between lncRNAs and coding genes with custom scripts; otherwise, we clustered the genes from different samples with WGCNA [57] to search common expression modules and then analyzed their function through functional enrichment analysis.
Differential gene expression analysis
Cuffdiff (v2.1.1) [49] was used to calculate FPKMs and p value of both lncRNAs and coding genes in each sample. A differential expression correct p value (using BH orthosis)≤0.05 and fold change>1.5 assigned as differentially expressed in four different comparisons (7 day DR vs fully fed, 42 day DR vs fully fed, 7 day DR vs 42 day DR, 7 day fully fed vs 42 day fully fed). qRT-PCR was used to validate 28 differentially expressed lncRNAs and corresponding 61 targets using eight samples from two biological duplicates including DR and fully fed groups at the 7 day and 42 day, and the relative gene expression level was calculated according to Livak and Schmittgen methods [58]. All custom primers were purchased from Sangon Biotech, and the primers of internal reference and identified transcripts in the qPCR were presented in Table S3. The SYBR green method was used for qRT-PCR using LightMix® kit (Roche, Swiss) following manufacturer’s instructions. Gene expression levels were identified according to the cycle threshold values and values are expressed as 2-△△CT [59]. The relative levels of lncRNAs and targets were normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Differential expression levels were compared by independent-samples multiple comparison methods between four different comparisons.
GO and KEGG enrichment analysis
Gene Ontology (GO) enrichment analysis of differentially expressed genes or lncRNAs target genes was implemented by the GOseq R package, in which gene length bias was corrected. GO terms with corrected P-value less than 0.05 were considered significantly enriched by differentially expressing genes. KOBAS software [60] was performed to test the statistical enrichment of differentially expressed genes or lncRNAs target genes in KEGG pathways (http://www.genome.jp/kegg/).
Evaluation of tissue specificity
In order to analyze tissue specificity, we downloaded RNA-seq data from paired-end sequencing of poly(A)+ RNA in biological duplicates from 29 dissected tissue samples (including the nervous, digestive, reproductive, endocrine, epidermal and muscle organ systems of larvae, pupae and adults), cultured cell lines and under environmental perturbations [17]. The known identified mRNA and lncRNAs in the present study were BLASTed with the above mentioned transcripts dataset generated in the previous study [17]. Then, 18 novel lncRNAs were randomly selected to analyze the tissue specificity of head, gut and fat body of fruit flies at the 7 day during DR process. Furthermore, 21 novel lncRNAs were picked at random from RNA-seq data to analyze the expression level in Drosophila S2 cell nucleus and cytoplasm using qRT-PCR method. Differential expression levels were compared by independent-samples t-test between groups. The heat map was drawn by MEV4 software [61] and heat map visualizations were conducted with TreeView program.
Regulatory elements
In order to find whether these lncRNAs have analogy to regulatory elements, a list of 205 Drosophila melanogaster core promoters were obtained from database (http://labs.biology.ucsd.edu/Kadonaga/DCPD.htm) and were used to blast with upstream 10kb sequences of identified lncRNAs.
Knockdown of lncRNA in Drosophila S2 cells
Drosophila S2 cells were cultured at 25°C in Hyclone SFX-Insect serum free medium (Thermo Scientific, USA) containing 100 U/mL penicillin and 100 µg/mL streptomycin. The lncRNA XLOC_076307 silencer was synthesized by RiboBio (GuangZhou, China). LncRNA silencer with non-specific nucleotide sequences was used as negative control (lncRNA silencer NC). LncRNA XLOC_076307 silencer and NC were transfected into Drosophila S2 cells in 6-well cell culture plates. For each lncRNA silencer, Drosophila S2 cells were transfected in triplicates with a final concentration of 150 nM in each well. The transfection was performed using transfection reagent (Biotool, China). The cells were harvested after 72 hours of transfection, to extract total RNA and expression levels of lncRNA XLOC_076307 and predicted interacting genes (hsp70BC, Gadd45, Socs36E) were measured by qPCR program as the above described method. LncRNA XLOC_076307 silencers oligonucleotide sequences and primers used in qPCR are listed in Table S2.
Data availability
The RNA-seq data used for this analysis are accessible through (SRP073695).
Conclusion
In conclusion, a substantial number of DR-specific lncRNAs and corresponding targets involved in multiple signaling pathways were identified in this study. We have predicted potential functions for novel lncRNA based on target function enrichment and also found important known lncRNA and their targets having the tissue specificity. Moreover, we obtained the result of the tissue specificity of some novel lncRNA in flies and the location of lncRNAs in fly S2 cell. Furthermore we have detected the expression levels of two lncRNA, their targets, and some genes related to aging signaling pathways. As the role of lncRNAs in DR specific lifespan extension have not yet been fully identified and understood, this analysis could serve as the beginning point for exploring the valuable insights into aging and aging-related disease research field for the future studies.
Supplementary Materials
Funding
This work was supported by The National Natural Science Foundation of China (NSFC31471998), the Research Program of Sichuan Bureau of Education (14TD0002), the “Thousand Talents Program” in Sichuan and Key Fund Project of Sichuan Provincial Department of Education (035Z1364).
Conflicts of Interest
The authors declare no competing financial interests.
References
- 1. Metaxakis A, Partridge L. Dietary restriction extends lifespan in wild-derived populations of Drosophila melanogaster.. PLoS One. 2013; 8:e74681. https://doi.org/10.1371/journal.pone.0074681 [PubMed]
- 2. Whitaker R, Gil MP, Ding F, Tatar M, Helfand SL, Neretti N. Dietary switch reveals fast coordinated gene expression changes in Drosophila melanogaster.. Aging (Albany NY). 2014; 6:355–68. https://doi.org/10.18632/aging.100662 [PubMed]
- 3. Fontana L, Partridge L. Promoting health and longevity through diet: from model organisms to humans. Cell. 2015; 161:106–18. https://doi.org/10.1016/j.cell.2015.02.020 [PubMed]
- 4. Heestand BN, Shen Y, Liu W, Magner DB, Storm N, Meharg C, Habermann B, Antebi A. Dietary restriction induced longevity is mediated by nuclear receptor NHR-62 in Caenorhabditis elegans.. PLoS Genet. 2013; 9:e1003651. https://doi.org/10.1371/journal.pgen.1003651 [PubMed]
- 5. Karunadharma PP, Basisty N, Dai DF, Chiao YA, Quarles EK, Hsieh EJ, Crispin D, Bielas JH, Ericson NG, Beyer RP, MacKay VL, MacCoss MJ, Rabinovitch PS. Subacute calorie restriction and rapamycin discordantly alter mouse liver proteome homeostasis and reverse aging effects. Aging Cell. 2015; 14:547–57. https://doi.org/10.1111/acel.12317 [PubMed]
- 6. Cannon MV, Buchner DA, Hester J, Miller H, Sehayek E, Nadeau JH, Serre D. Maternal nutrition induces pervasive gene expression changes but no detectable DNA methylation differences in the liver of adult offspring. PLoS One. 2014; 9:e90335. https://doi.org/10.1371/journal.pone.0090335 [PubMed]
- 7. Testa G, Biasi F, Poli G, Chiarpotto E. Calorie restriction and dietary restriction mimetics: a strategy for improving healthy aging and longevity. Curr Pharm Des. 2014; 20:2950–77. https://doi.org/10.2174/13816128113196660699 [PubMed]
- 8. Renaud HJ, Cui JY, Lu H, Klaassen CD. Effect of diet on expression of genes involved in lipid metabolism, oxidative stress, and inflammation in mouse liver-insights into mechanisms of hepatic steatosis. PLoS One. 2014; 9:e88584. https://doi.org/10.1371/journal.pone.0088584 [PubMed]
- 9. Lee KP. Dietary protein:carbohydrate balance is a critical modulator of lifespan and reproduction in Drosophila melanogaster: a test using a chemically defined diet. J Insect Physiol. 2015; 75:12–19. https://doi.org/10.1016/j.jinsphys.2015.02.007 [PubMed]
- 10. Cui M, Xiao Z, Wang Y, Zheng M, Song T, Cai X, Sun B, Ye L, Zhang X. Long noncoding RNA HULC modulates abnormal lipid metabolism in hepatoma cells through an miR-9-mediated RXRA signaling pathway. Cancer Res. 2015; 75:846–57. https://doi.org/10.1158/0008-5472.CAN-14-1192 [PubMed]
- 11. Pan W, Liu L, Wei J, Ge Y, Zhang J, Chen H, Zhou L, Yuan Q, Zhou C, Yang M. A functional lncRNA HOTAIR genetic variant contributes to gastric cancer susceptibility. Mol Carcinog. 2016; 55:90–96. https://doi.org/10.1002/mc.22261 [PubMed]
- 12. Luo X, Zhu J, Cheng Z, Zhang F, Zhang G, Yuan J, Jin C. Lack of association of a genetic variant in the long intergenic noncoding RNA (linc01080) with Alzheimer’s disease and amnestic mild cognitive impairment in Han Chinese. Int J Neurosci. 2015; 125:419–23. https://doi.org/10.3109/00207454.2014.944616 [PubMed]
- 13. Greco S, Gorospe M, Martelli F. Noncoding RNA in age-related cardiovascular diseases. J Mol Cell Cardiol. 2015; 83:142–55. https://doi.org/10.1016/j.yjmcc.2015.01.011 [PubMed]
- 14. Beltrami C, Angelini TG, Emanueli C. Noncoding RNAs in diabetes vascular complications. J Mol Cell Cardiol. 2015; 89:42–50. https://doi.org/10.1016/j.yjmcc.2014.12.014 [PubMed]
- 15. Porro A, Feuerhahn S, Lingner J. TERRA-reinforced association of LSD1 with MRE11 promotes processing of uncapped telomeres. Cell Reports. 2014; 6:765–76. https://doi.org/10.1016/j.celrep.2014.01.022 [PubMed]
- 16. Bradford JR, Wappett M, Beran G, Logie A, Delpuech O, Brown H, Boros J, Camp NJ, McEwen R, Mazzola AM, D’Cruz C, Barry ST. Whole transcriptome profiling of patient-derived xenograft models as a tool to identify both tumor and stromal specific biomarkers. Oncotarget. 2016; 7:20773–87. [PubMed]
- 17. Brown JB, Boley N, Eisman R, May GE, Stoiber MH, Duff MO, Booth BW, Wen J, Park S, Suzuki AM, Wan KH, Yu C, Zhang D, et al. Diversity and dynamics of the Drosophila transcriptome. Nature. 2014; 512:393–99. https://doi.org/10.1038/nature12962 [PubMed]
- 18. Bunker BD, Nellimoottil TT, Boileau RM, Classen AK, Bilder D. The transcriptional response to tumorigenic polarity loss in Drosophila.. eLife. 2015; 4. https://doi.org/10.7554/eLife.03189 [PubMed]
- 19. Emran S, Yang M, He X, Zandveld J, Piper MD. Target of rapamycin signalling mediates the lifespan-extending effects of dietary restriction by essential amino acid alteration. Aging (Albany NY). 2014; 6:390–98. https://doi.org/10.18632/aging.100665 [PubMed]
- 20. Menger KE, James AM, Cochemé HM, Harbour ME, Chouchani ET, Ding S, Fearnley IM, Partridge L, Murphy MP. Fasting, but Not Aging, Dramatically Alters the Redox Status of Cysteine Residues on Proteins in Drosophila melanogaster.. Cell Reports. 2015; 11:1856–65. https://doi.org/10.1016/j.celrep.2015.05.033 [PubMed]
- 21. Weiss S, Minke B. A new genetic model for calcium induced autophagy and ER-stress in Drosophila photoreceptor cells. Channels (Austin). 2015; 9:14–20. https://doi.org/10.4161/19336950.2014.981439 [PubMed]
- 22. Grammatikakis I, Panda AC, Abdelmohsen K, Gorospe M. Long noncoding RNAs(lncRNAs) and the molecular hallmarks of aging. Aging (Albany NY). 2014; 6:992–1009. https://doi.org/10.18632/aging.100710 [PubMed]
- 23. Kim J, Kim KM, Noh JH, Yoon JH, Abdelmohsen K, Gorospe M. Long noncoding RNAs in diseases of aging. Biochim Biophys Acta. 2016; 209-21. .
- 24. Wu CL, Wang Y, Jin B, Chen H, Xie BS, Mao ZB. Senescence-associated long non-coding RNA (SALNR) delays oncogene-induced senescence through NF90 regulation. J Biol Chem. 2015; 290:30175–92. https://doi.org/10.1074/jbc.M115.661785 [PubMed]
- 25. Kour S, Rath PC. Age-dependent differential expression profile of a novel intergenic long noncoding RNA in rat brain. Int J Dev Neurosci. 2015; 47:286–97. https://doi.org/10.1016/j.ijdevneu.2015.08.008 [PubMed]
- 26. Tupy JL, Bailey AM, Dailey G, Evans-Holm M, Siebel CW, Misra S, Celniker SE, Rubin GM. Identification of putative noncoding polyadenylated transcripts in Drosophila melanogaster.. Proc Natl Acad Sci USA. 2005; 102:5495–500. https://doi.org/10.1073/pnas.0501422102 [PubMed]
- 27. Castanieto A, Johnston MJ, Nystul TG. EGFR signaling promotes self-renewal through the establishment of cell polarity in Drosophila follicle stem cells. eLife. 2014; 3. https://doi.org/10.7554/eLife.04437 [PubMed]
- 28. Bgatova N, Dubatolova T, Omelyanchuk L, Plyusnina E, Shaposhnikov M, Moskalev A. Gadd45 expression correlates with age dependent neurodegeneration in Drosophila melanogaster.. Biogerontology. 2015; 16:53–61. https://doi.org/10.1007/s10522-014-9533-0 [PubMed]
- 29. Kuk JL, Saunders TJ, Davidson LE, Ross R. Age-related changes in total and regional fat distribution. Ageing Res Rev. 2009; 8:339–48. https://doi.org/10.1016/j.arr.2009.06.001 [PubMed]
- 30. Musselman LP, Fink JL, Narzinski K, Ramachandran PV, Hathiramani SS, Cagan RL, Baranski TJ. A high-sugar diet produces obesity and insulin resistance in wild-type Drosophila.. Dis Model Mech. 2011; 4:842–49. https://doi.org/10.1242/dmm.007948 [PubMed]
- 31. Schroeder MC, Halder G. Regulation of the Hippo pathway by cell architecture and mechanical signals. Semin Cell Dev Biol. 2012; 23:803–11. https://doi.org/10.1016/j.semcdb.2012.06.001 [PubMed]
- 32. Zhang L. Control of growth and beyond: a special issue on Hippo signaling. Acta Biochim Biophys Sin (Shanghai). 2015; 47:1. https://doi.org/10.1093/abbs/gmu113 [PubMed]
- 33. Hardie RC, Juusola M. Phototransduction in Drosophila.. Curr Opin Neurobiol. 2015; 34:37–45. https://doi.org/10.1016/j.conb.2015.01.008 [PubMed]
- 34. Martianov I, Ramadass A, Serra Barros A, Chow N, Akoulitchev A. Repression of the human dihydrofolate reductase gene by a non-coding interfering transcript. Nature. 2007; 445:666–70. https://doi.org/10.1038/nature05519 [PubMed]
- 35. Shamovsky I, Ivannikov M, Kandel ES, Gershon D, Nudler E. RNA-mediated response to heat shock in mammalian cells. Nature. 2006; 440:556–60. https://doi.org/10.1038/nature04518 [PubMed]
- 36. Wilusz CJ, Wilusz J. HuR-SIRT: the hairy world of posttranscriptional control. Mol Cell. 2007; 25:485–87. https://doi.org/10.1016/j.molcel.2007.02.001 [PubMed]
- 37. Herskovits AZ, Guarente L. SIRT1 in neurodevelopment and brain senescence. Neuron. 2014; 81:471–83. https://doi.org/10.1016/j.neuron.2014.01.028 [PubMed]
- 38. He F, Wei C, Wu H, Cheung D, Jiao R, Ma J. Fundamental origins and limits for scaling a maternal morphogen gradient. Nat Commun. 2015; 6:6679. https://doi.org/10.1038/ncomms7679 [PubMed]
- 39. Hankeln T, Jaenicke V, Kiger L, Dewilde S, Ungerechts G, Schmidt M, Urban J, Marden MC, Moens L, Burmester T. Characterization of Drosophila hemoglobin. Evidence for hemoglobin-mediated respiration in insects. J Biol Chem. 2002; 277:29012–17. https://doi.org/10.1074/jbc.M204009200 [PubMed]
- 40. Mummery-Widmer JL, Yamazaki M, Stoeger T, Novatchkova M, Bhalerao S, Chen D, Dietzl G, Dickson BJ, Knoblich JA. Genome-wide analysis of Notch signalling in Drosophila by transgenic RNAi. Nature. 2009; 458:987–92. https://doi.org/10.1038/nature07936 [PubMed]
- 41. Kim HJ, Morrow G, Westwood JT, Michaud S, Tanguay RM. Gene expression profiling implicates OXPHOS complexes in lifespan extension of flies over-expressing a small mitochondrial chaperone, Hsp22. Exp Gerontol. 2010; 45:611–20. https://doi.org/10.1016/j.exger.2009.12.012 [PubMed]
- 42. Diaconeasa AG, Rachita M, Stefan-van Staden RI. A new hypothesis of aging. Med Hypotheses. 2015; 84:252–57. https://doi.org/10.1016/j.mehy.2015.01.006 [PubMed]
- 43. Chen H, Zheng X, Zheng Y. Lamin-B in systemic inflammation, tissue homeostasis, and aging. Nucleus. 2015; 6:183–86. https://doi.org/10.1080/19491034.2015.1040212 [PubMed]
- 44. Kondo T, Hashimoto Y, Kato K, Inagaki S, Hayashi S, Kageyama Y. Small peptide regulators of actin-based cell morphogenesis encoded by a polycistronic mRNA. Nat Cell Biol. 2007; 9:660–65. https://doi.org/10.1038/ncb1595 [PubMed]
- 45. Gong WJ, Golic KG. Loss of Hsp70 in Drosophila is pleiotropic, with effects on thermotolerance, recovery from heat shock and neurodegeneration. Genetics. 2006; 172:275–86. https://doi.org/10.1534/genetics.105.048793 [PubMed]
- 46. Monahan AJ, Starz-Gaiano M. Socs36E attenuates STAT signaling to optimize motile cell specification in the Drosophila ovary. Dev Biol. 2013; 379:152–66. https://doi.org/10.1016/j.ydbio.2013.03.022 [PubMed]
- 47. Plyusnina EN, Shaposhnikov MV, Moskalev AA. Increase of Drosophila melanogaster lifespan due to D-GADD45 overexpression in the nervous system. Biogerontology. 2011; 12:211–26. https://doi.org/10.1007/s10522-010-9311-6 [PubMed]
- 48. Bass TM, Grandison RC, Wong R, Martinez P, Partridge L, Piper MD. Optimization of dietary restriction protocols in Drosophila.. J Gerontol A Biol Sci Med Sci. 2007; 62:1071–81. https://doi.org/10.1093/gerona/62.10.1071 [PubMed]
- 49. Ghosh S, Chan CK. Analysis of RNA-Seq Data Using TopHat and Cufflinks. Methods Mol Biol. 2016; 1374:339–61. https://doi.org/10.1007/978-1-4939-3167-5_18 [PubMed]
- 50. Sun L, Luo H, Bu D, Zhao G, Yu K, Zhang C, Liu Y, Chen R, Zhao Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013; 41:e166. https://doi.org/10.1093/nar/gkt646 [PubMed]
- 51. Kong L, Zhang Y, Ye ZQ, Liu XQ, Zhao SQ, Wei L, Gao G. CPC: assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007; 35:W345-9. https://doi.org/10.1093/nar/gkm391 [PubMed]
- 52. Punta M, Coggill PC, Eberhardt RY, Mistry J, Tate J, Boursnell C, Pang N, Forslund K, Ceric G, Clements J, Heger A, Holm L, Sonnhammer EL, et al. The Pfam protein families database. Nucleic Acids Res. 2012; 40:D290–301. https://doi.org/10.1093/nar/gkr1065 [PubMed]
- 53. Lin MF, Jungreis I, Kellis M. PhyloCSF: a comparative genomics method to distinguish protein coding and non-coding regions. Bioinformatics. 2011; 27:i275–82. https://doi.org/10.1093/bioinformatics/btr209 [PubMed]
- 54. Trapnell C, Williams BA, Pertea G, Mortazavi A, Kwan G, van Baren MJ, Salzberg SL, Wold BJ, Pachter L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat Biotechnol. 2010; 28:511–15. https://doi.org/10.1038/nbt.1621 [PubMed]
- 55. Paralkar VR, Taborda CC, Huang P, Yao Y, Kossenkov AV, Prasad R, Luan J, Davies JO, Hughes JR, Hardison RC, Blobel GA, Weiss MJ. Unlinking an lncRNA from Its Associated cis Element. Mol Cell. 2016; 62:104–10. https://doi.org/10.1016/j.molcel.2016.02.029 [PubMed]
- 56. Peschansky VJ, Pastori C, Zeier Z, Wentzel K, Velmeshev D, Magistri M, Silva JP, Wahlestedt C. The long non-coding RNA FMR4 promotes proliferation of human neural precursor cells and epigenetic regulation of gene expression in trans. Mol Cell Neurosci. 2016; 74:49–57. https://doi.org/10.1016/j.mcn.2016.03.008 [PubMed]
- 57. Langfelder P, Horvath S. WGCNA: an R package for weighted correlation network analysis. BMC Bioinformatics. 2008; 9:559. https://doi.org/10.1186/1471-2105-9-559 [PubMed]
- 58. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001; 25:402–08. https://doi.org/10.1006/meth.2001.1262 [PubMed]
- 59. Bernard D, Prasanth KV, Tripathi V, Colasse S, Nakamura T, Xuan Z, Zhang MQ, Sedel F, Jourdren L, Coulpier F, Triller A, Spector DL, Bessis A. A long nuclear-retained non-coding RNA regulates synaptogenesis by modulating gene expression. EMBO J. 2010; 29:3082–93. https://doi.org/10.1038/emboj.2010.199 [PubMed]
- 60. Xie C, Mao X, Huang J, Ding Y, Wu J, Dong S, Kong L, Gao G, Li CY, Wei L. KOBAS 2.0: a web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011 (Suppl ); 39:W316-22. https://doi.org/10.1093/nar/gkr483 [PubMed]
- 61. Ma Y, Song J, Chen B, Xu X, Lin G. SLC7A5 act as a potential leukemic transformation target gene in myelodysplastic syndrome. Oncotarget. 2016; 7:6566–75. [PubMed]