



Introduction
Aging is a complex process associated with various molecular modifications in cells including epigenetic transitions that create changes in gene expression. Many studies have indicated DNA methylation (DNAmet) is an important component of aging. In addition to inherited, genetically driven DNAmet changes, a broad spectrum of environmental factors can influence the methylation status of many sites making the human DNA methylome a sensitive marker set of epigenetic drift rather than a stable imprint [1–3]. Therefore, age predicted with epigenetic markers better reflects the biological age than the chronological age of an individual and such differences in aging rates may potentially be linked to various clinical and environmental factors.
With the advent of high-throughput sequencing technologies epigenome-wide association studies became available and led to successful identification of age-correlated DNAmet changes [1,2,4–8]. These discoveries were associated with development of age prediction models that have allowed unprecedented accuracy for age estimation regimes. Epigenetic age prediction methods have been mostly developed based on DNA extracted from blood but other DNA sources have also been investigated [9]. The studies of age-related DNAmet markers have also revealed that age acceleration defined as the discrepancy between methylation age and chronological age, may be a reliable indicator of increased disease and mortality risk [10–13]. Since age acceleration can be a biologically meaningful biomarker, knowledge of its progression may find practical application in many disciplines including sports medicine and forensics [14,15].
It is widely accepted that people who engage in moderate physical activity will benefit from better mood and cognition, lower levels of cardiovascular disease and a slower rate of aging [16,17]. Indeed, the bulk of research has reported longer life expectancy and lower mortality risk for elite athletes compared to the general population and such results are particularly consistent for groups of endurance athletes. Results from several large meta-analyses are the most informative [18–20]. Investigating the epigenome of elite athletes provides a potentially informative way to study how life-style can affect the functioning of the human genome. Moreover, it has been shown that DNAmet in various CpG sites may undergo diverse regulation [21]. Thus, the tracking of individual epigenetic markers may provide better insight into the molecular processes that underlie aging by disclosing the cellular activity and potential interactions the key genes are involved in. The possibility to examine more closely the molecular processes associated with aging from the advent of epigenomic analysis and epigenetic age prediction, permits more objective exploration of the role of physical activity in a healthy lifestyle.
Our study aimed to examine whether intense physical exercise can alter DNAmet profiles and lead to changes in the rate of aging. To do this we studied the DNAmet profile of Polish professional athletes at 5 CpG sites in five different genes that we had previously selected and developed into a DNAm-based age prediction calculator [22]. The study intended to distinguish markers following a regular clock-like pattern of methylation change from those sites more prone to environmental factors associated with the life of top-level athletes.
Results
DNA methylation status at the 5 CpG sites in five age related genes was analyzed in 176 athletes, in comparison to 128 healthy control individuals from the general population matched by age (P=0.760) and gender (P=0.949). Elite athletes comprised young individuals with a mean age of 24.1 and was 112 males (63.6%) and 64 females (36.4%); as outlined in Table 1. The group of power athletes consisted of 57 males (60.6%) and 37 females (39.4%) and had a mean age of 23 years. The group of endurance athletes comprised 55 males (67.1%) and 27 females (32.9%) with a mean age of 25.2 years. The control group had a mean age of 24.2 and was 81 males (63.3%) and 47 females (36.7%). Age distribution graphs for athletes and controls by gender are provided in Supplementary Figure 1. The mean age of males in the group of athletes was higher (24.9) compared to females (22.7); and similarly in the control group with a mean age of 25.4 for males and 22.2 for females.
Table 1. DNA methylation status of single age-related CpG sites measured in Polish professional athletes compared to age- and gender-matched controls from the general population
| Locus CpG site | Mean % of DNAmet | P value | Mean % of DNAmet | P value | Mean % of DNAmet | P value | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| All athletes | Controls | Endurance | Controls | Power | Controls | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ELOVL2 C7 | 60.89 | 61.09 | 0.791 | 61.94 | 61.09 | 0.342 | 59.98 | 61.09 | 0.196 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MIR29B2C C1 | 79.43 | 80.54 | 0.113 | 80.04 | 80.54 | 0.542 | 78.89 | 80.54 | 0.058 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FHL2 C2 | 36.37 | 35.87 | 0.375 | 36.41 | 35.87 | 0.453 | 36.33 | 35.87 | 0.465 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TRIM59 C7 | 31.43 | 28.19 | 1.894x10-7 | 30.56 | 28.19 | 0.004 | 32.18 | 28.19 | 2.946x10-7 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KLF14 C1 | 5.16 | 4.12 | 8.974x10-10 | 5.23 | 4.12 | 1.866x10-7 | 5.10 | 4.12 | 4.987x10-7 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| . Significant values are marked in bold. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
DNA methylation status
The study revealed significantly altered DNA methylation patterns between the whole group of athletes and controls in TRIM59 (P=1.894x10-7) and KLF14 (P=8.974x10-10), as well as separately for endurance and power athletes (Table 1). Although the results were significant for both endurance and power athletes, a more marked difference was noted in TRIM59 C7 for the power athletes (P=2.946x10-7). In KLF14 C1, hypermethylation was observed for power and endurance athletes with a similar level of significance of P=4.987x10-7 and P=1.866x10-7, respectively (Table 1). Analyses performed separately for females and males confirmed TRIM59 and KLF14 markers to be significantly hypermethylated in athletes of both genders. (Supplementary Table 1).
Age prediction and age acceleration
Table 2 shows that our established multivariate 5-CpG ANN model predicted the whole group of elite athletes with a lower accuracy of MAE = 3.3, compared to healthy controls of similar mean age and age distribution that had a MAE = 2.4, P = 3.815x10-4. When the two athlete groups were analysed separately, a significant decrease of age prediction accuracy from MAE=2.4 to MAE = 3.7 was only found for power athletes (P=5.727x10-5). Analyses performed separately for females and males confirmed reduced accuracy of age prediction in the group of power athletes vs. controls in both genders (P=0.013 and P=0.002, for females and males respectively); outlined in Supplementary Table 2.
Table 2. Age prediction accuracy measured by MAE using the 5-CpG sites model published in [22], modified by using ANN statistical analysis.
| Model | Compared groups | N | MAE | Std. Deviation | P value | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5 CpG model [22] | Athletes | 175 | 3.273 | 2.620 | 3.815x10-4 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Controls | 128 | 2.361 | 1.797 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Endurance | 82 | 2.776 | 2.391 | 0.180 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Controls | 128 | 2.361 | 1.797 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Power | 93 | 3.712 | 2.745 | 5.727x10-5 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Controls | 128 | 2.361 | 1.797 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| One sample was removed from prediction analysis because of missing data in MIR29B2C C1. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Age acceleration from chronological and methylation age divergence (determined using the 5-CpG model) was further calculated and tested for its association with intense physical exercise. This analysis showed the status of elite athletes is significantly associated with age acceleration (P=1.503x10-7) and was significant for both groups (1.051x10-9 and 0.004 for power and endurance athletes, respectively; Table 3). Analysis performed at the level of single CpG predictors showed that significant association of the status of elite athletes with age acceleration arose from increased DNA methylation of the CpGs at TRIM59 and KLF14. The level of significance of association between the status of power athletes and age acceleration was highest when just TRIM59 was used as a predictor of age (P=1.239x10-10 comparing to P=0.003 obtained for endurance athletes), but at a comparable level in KLF14 in both groups (P=3.345x10-7 and P=2.116x10-8, for endurance and power groups, respectively). The type of sport competition performed was also found to be significantly associated with age acceleration (P=0.003) indicating the increased aging of athletes engaged in power sports compared to endurance sports (Table 3).
Table 3. Association between extensive physical exercise and age acceleration in Polish athletes.
| Models | Age acceleration (5 CpGs model) | Age acceleration (KLF14 C1) | Age acceleration (TRIM59 C7) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Effect size* | SE | P value | Effect size* | SE | P value | Effect size* | SE | P value | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Athletes vs. Controls | 2.156 | 0.401 | 1.503x10-7 | 5.559 | 0.845 | 2.111x10-10 | 4.629 | 0.834 | 6.270x10-8 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Endurance vs. Controls | 1.329 | 0.452 | 0.004 | 5.478 | 1.039 | 3.345x10-7 | 2.647 | 0.867 | 0.003 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Power vs. Controls | 2.885 | 0.452 | 1.051x10-9 | 5.630 | 0.968 | 2.116x10-8 | 6.194 | 0.917 | 1.239x10-10 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Power vs. Endurance | 1.665 | 0.558 | 0.003 | -0.052 | 1.169 | 0.965 | 3.455 | 1.272 | 0.007 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| *Effect sizes are the beta coefficients from linear regression models adjusted for age (years) and gender and additionally for training experience (years) when the type of the competition (power vs. endurance) was analysed. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Predicted age versus chronological age in endurance and power athletes plus the group of controls is presented on Figure 1, which shows predictions based on the 5-CpGs model and TRIM59 and KLF14 separately. Figure 2 shows that age prediction based on CpG site C1 in KLF14 indicated athletes to be an average 5.5 years older than healthy controls (P=1.766x10-9), endurance athletes to be 5.9 years older than controls from the general population (P=2.511x10-7) and power athletes to be 5.1 years older than healthy controls (P=8.443x10-7). Age prediction performed with DNA methylation values from CpG site C7 in TRIM59 showed athletes to be an average 4.5 years older than healthy controls (P=9.405x10-8), power athletes to be 5.6 years older than healthy controls (P=2.105x10-7) while endurance athletes were predicted to be 3.3 years older than healthy controls (P=0.003).
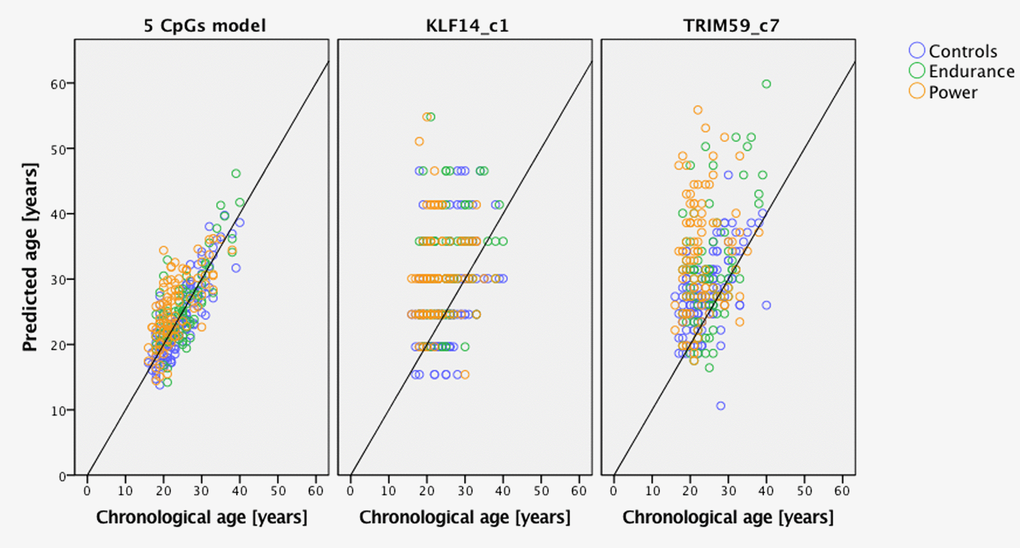
Figure 1. Predicted age vs. chronological age of power and endurance athletes compared to age- and gender-matched controls. Predictions based on the 5 CpGs model and separately KLF14 c1 and TRIM59 c7.
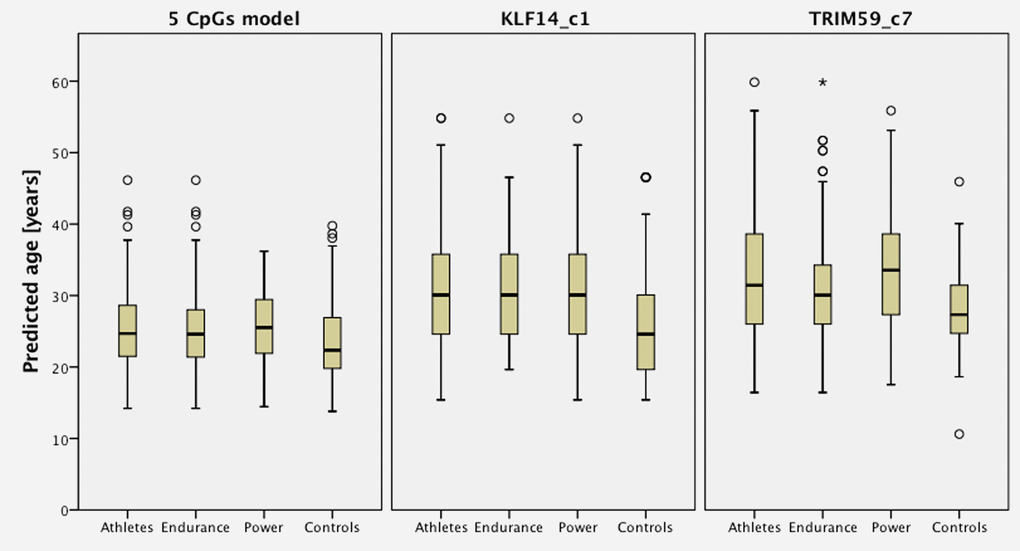
Figure 2. Predicted age of athletes comparing with age- and gender- matched controls based on predictors from 5 CpGs model, KLF14 and TRIM59.
Discussion
The impact of physical exercises on epigenetic age
Very intensive physical exercise as a routine part of elite athlete training has been shown to have favorable effects on lifespan longevity and is associated with a lowered risk of cardiovascular diseases and cancer in later life [19,20,24]. Importantly, individuals with active lifestyles but free from the stress of extreme exercise regimes benefit from regular physical activity by reducing their risk of all forms of mortality, and in proportion to the intensity of the training [25]. Better insight into the molecular processes behind the aging of elite athletes is now possible thanks to recent advances in epigenetics and the efficiency of DNAmet age prediction systems in providing reliable indicators of the aging rate. Our study indicates accelerated aging of the whole group of elite athletes, with power athletes shown to have higher levels of age acceleration compared to endurance athletes. The observed acceleration of aging may potentially have significant implications, as it has been shown that a difference between the chronological age of an individual and the age measured by DNA methylation is an indicator of a person’s overall condition and is predictive of mortality in later life.
The meaning of epigenetic age
The epigenetic clock CpGs defined as correlated with age consistently between individuals and included in various age prediction models, reveal some degree of inter-individual variability [26]. There is an on-going debate about the real meaning of age measured by DNAmet status. The picture arising from recent research indicates that various markers included in age estimation models may be regulated in a different way. Particular age-correlated CpGs could be liable to moderation by inherited factors and the environment [2]. It has been shown that the epigenetic clock developed by Horvath can also measure age in non-proliferative tissues but at the same time there is evidence that hypermethylation of one of the most informative age predictors – ELOVL2, is associated with cell replications and thus indicates a mitotic age [21]. Therefore, it is conceivable that some DNAmet markers may be more sensitive to the environment than others and consequently they can provide a better measure of epigenetic drift. Consequently, it is unsurprising that not all age predictors are informative in terms of increased mortality risk. The study of Lin et al selected 37 CpGs indicative of life expectancy from their full set of 99 markers. Additionally, they tested alternative age prediction models and found that the marker sets proposed by Horvath and Hannum contained mortality sensitive CpG sites [12,1,2]. Two out of five markers investigated here, namely TRIM59 and KLF14, are among the suggested life expectancy-sensitive markers from the Hannum model. TRIM59 belongs to the ten most informative markers from the age predictors included in that age prediction model [12]. Notably, an independent study has demonstrated that 5-year higher DNA methylation age predicted by the Hannum model was associated with a 21% increased mortality risk [11]. Overall, multiple studies have reported an 11% to 35% increase in mortality risk for individuals with a 5-year higher DNA methylation age [11,12,27]. Recently, Zhang et al presented a list of life expectancy DNA methylation markers which are independent from the epigenetic clock, providing further evidence that aging related markers can be different from markers predicting chronological age [13].
Conclusions
Our study findings indicate various age predictors can show detectable differences in sensitivity to intense physical activity. The study revealed accelerated hypermethylation of TRIM59 and KLF14 in elite athletes suggesting their higher epigenetic age, which is contrary to epidemiological data showing longer life duration of athletes. However, the two genes are involved in anti-tumor and anti-inflammatory activities, respectively, which in turn could affect better life-expectancy. The results obtained may reveal a complex influence of intense physical training on the aging process. We speculate that the resulting diminished expression of these genes may potentially launch signaling networks that contribute to an observed lower risk for elite athletes to develop cardiovascular disease and cancer.
Investigation of an extended list of age-related markers and detailed analyses of the relevance of individual CpGs for biological aging will aid in the understanding of the role of DNA methylation in human aging and an assessment of the relevance of DNA methylation as an indicator of the condition of an individual.
Materials and Methods
Samples and analysis of DNA methylation
The study was approved by the Ethics Committee of the Józef Piłsudski University of Physical Education in Warsaw (SKE 01-50/2012) and the institutional ethical review board of the Ethics Committee of Investigation in Galicia, Spain (CAEI: 2013/543). Written informed consent was obtained from the elite athletes and counterpart controls from the general population forming the study subjects. Peripheral blood was collected in EDTA tubes from 176 elite athletes in the years 2010-2014. Chronological age at the time of blood collection was used for all the calculations in this study. Top-level professionals (including medallists of Olympic Games and World championships) in power and endurance sports, 100 athletics, 21 speed skaters, 28 swimmers and 27 rowers were enrolled in the study. 128 blood samples from the general population matched by age and gender, were recruited for the control group (Table 4). DNA was extracted using standard methods and quantified using Qubit dsDNA High Sensitivity Assay Kit (Thermo Fisher). From 500 ng to 2 μg of DNA was treated with sodium bisulfite using the EpiTect 96 Bisulfite Kit (Qiagen, Hilden, Germany), following manufacturer’s guidelines. Specific CpG sites were analysed using previously described primers and methods [22]. Details of the analysed sites are presented in Table 5.
Table 4. Characteristics of the Polish elite athletes and controls used in the study.
| Sport discipline | N [%] | The specifics of competition | Male N [%] | Mean Age ± SD | |||||
| Power N [%] | Endurance N [%] | All | Power N [%] | Endurance N [%] | All | Power N [%] | Endurance N [%] | ||
| Athletics | 100 [56.8] | 70 [70.0] | 30 [30.0] | 61 [61.0] | 42 [60.0] | 19 [63.3] | 24.8 ±5.3 | 23.8±4.9 | 27.2±5.3 |
| Speed skating | 21 [11.9] | 7 [33.3] | 14 [66.7] | 17 [81.0] | 6 [85.7] | 11 [78.6] | 21.5 ± 4.1 | 19.3±1.4 | 22.6±4.6 |
| Swimming | 28 [15.9] | 17 [60.7] | 11 [39.3] | 16 [57.1] | 9 [52.9] | 7 [63.6] | 22.0 ± 5.0 | 21.4±3.8 | 23±6.5 |
| Rowing | 27 [15.4] | 0 [0.0] | 27 [100.0] | 18 [66.7] | - | 18 [66.7] | 25.4 ± 4.2 | - | 25.4±4.2 |
| Total athletes | 176 [100] | 94 [53.4] | 82 [46.6] | 112 [63.6] | 57 [60.6] | 55 [67.1] | 24.1 ± 5.1 | 23.0±4.8 | 25.2±5.3 |
| Controls | 128 [100] | - | - | 81 [63.3] | - | - | 24.2 ± 5.0 | - | - |
Table 5. The studied markers and CpG sites.
| Gene locus | CpG site | Chromosome | Chromosome location (GRCH38) |
| ELOVL2 | C7 | Chr6 | 11044634 |
| MIR29B2C | C1 | Chr1 | 207823681 |
| TRIM59 | C7 | Chr3 | 160450199 |
| KLF14 | C1 | Chr7 | 130734355 |
| FHL2 | C2 | Chr2 | 105399288 |
Statistical analyses and prediction of DNA methylation age
Analyses were performed using PS IMAGO 4 (IBM SPSS Statistics 24). DNA methylation was measured in the two groups of 176 elite athletes plus 128 healthy counterparts of the age/gender matched control group at 5 CpGs included in the age prediction model (Table 5). Methylation data was analysed using independent sample Student’s t test and the balanced distribution of age in the athlete and control groups confirmed with nonparametric Kolmogorov-Smirnov test. Age predictions were made with the method developed by Zbieć-Piekarska et al. [22] after remodelling with the artificial neural networks (ANN) approach applied in the study of Spólnicka et al. [23]. The training set of 305 individuals from the above previous studies was used for the age predictions of the current study. The predicted age of athletes was compared with their true chronological age to calculate mean absolute error (MAE). Independent sample Student’s t test was used to compare mean predicted age and MAE. Age acceleration (AA) - a measure of the discrepancy between methylation age and chronological age - was designated in the form of residuals calculated from linear regression analysis; where predicted age was treated as the dependent variable and chronological age as the independent variable as described in [14]. Association analysis of individual levels of intense training with age acceleration was tested using linear regression, controlling for age and gender and in some analyses, for the number of years spent in training.
Supplementary Materials
Acknowledgements
The authors wish to thank all participants of the study.
Conflicts of Interest
Authors declare no conflicts of interest.
Funding
The study was supported by grants from the National Centre for Research and Development in Poland, NEXT no. DOB-BIO7/17/01/2015 and from National Science Centre no. 2012/07/B/NZ7/04261. EP is supported by the Foundation for Polish Science (FNP) programme START 2017.
References
- 1. Horvath S. DNA methylation age of human tissues and cell types. Genome Biol. 2013; 14:R115. https://doi.org/10.1186/gb-2013-14-10-r115 [PubMed]
- 2. Hannum G, Guinney J, Zhao L, Zhang L, Hughes G, Sadda S, Klotzle B, Bibikova M, Fan JB, Gao Y, Deconde R, Chen M, Rajapakse I, et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol Cell. 2013; 49:359–67. https://doi.org/10.1016/j.molcel.2012.10.016 [PubMed]
- 3. Fraga MF, Ballestar E, Paz MF, Ropero S, Setien F, Ballestar ML, Heine-Suñer D, Cigudosa JC, Urioste M, Benitez J, Boix-Chornet M, Sanchez-Aguilera A, Ling C, et al. Epigenetic differences arise during the lifetime of monozygotic twins. Proc Natl Acad Sci USA. 2005; 102:10604–09. https://doi.org/10.1073/pnas.0500398102 [PubMed]
- 4. Bocklandt S, Lin W, Sehl ME, Sánchez FJ, Sinsheimer JS, Horvath S, Vilain E. Epigenetic predictor of age. PLoS One. 2011; 6:e14821. https://doi.org/10.1371/journal.pone.0014821 [PubMed]
- 5. Garagnani P, Bacalini MG, Pirazzini C, Gori D, Giuliani C, Mari D, Di Blasio AM, Gentilini D, Vitale G, Collino S, Rezzi S, Castellani G, Capri M, et al. Methylation of ELOVL2 gene as a new epigenetic marker of age. Aging Cell. 2012; 11:1132–34. https://doi.org/10.1111/acel.12005 [PubMed]
- 6. Florath I, Butterbach K, Müller H, Bewerunge-Hudler M, Brenner H. Cross-sectional and longitudinal changes in DNA methylation with age: an epigenome-wide analysis revealing over 60 novel age-associated CpG sites. Hum Mol Genet. 2014; 23:1186–201. https://doi.org/10.1093/hmg/ddt531 [PubMed]
- 7. Weidner CI, Lin Q, Koch CM, Eisele L, Beier F, Ziegler P, Bauerschlag DO, Jöckel KH, Erbel R, Mühleisen TW, Zenke M, Brümmendorf TH, Wagner W. Aging of blood can be tracked by DNA methylation changes at just three CpG sites. Genome Biol. 2014; 15:R24. https://doi.org/10.1186/gb-2014-15-2-r24 [PubMed]
- 8. Lee HY, Jung SE, Oh YN, Choi A, Yang WI, Shin KJ. Epigenetic age signatures in the forensically relevant body fluid of semen: a preliminary study. Forensic Sci Int Genet. 2015; 19:28–34. https://doi.org/10.1016/j.fsigen.2015.05.014 [PubMed]
- 9. Freire-Aradas A, Phillips C, Lareu MV. Forensic individual age estimation with DNA: from initial approaches to methylation tests. Forensic Sci Rev. 2017; 29:121–44. [PubMed]
- 10. Perna L, Zhang Y, Mons U, Holleczek B, Saum KU, Brenner H. Epigenetic age acceleration predicts cancer, cardiovascular, and all-cause mortality in a German case cohort. Clin Epigenetics. 2016; 8:64. https://doi.org/10.1186/s13148-016-0228-z [PubMed]
- 11. Marioni RE, Shah S, McRae AF, Chen BH, Colicino E, Harris SE, Gibson J, Henders AK, Redmond P, Cox SR, Pattie A, Corley J, Murphy L, et al. DNA methylation age of blood predicts all-cause mortality in later life. Genome Biol. 2015; 16:25. https://doi.org/10.1186/s13059-015-0584-6 [PubMed]
- 12. Lin Q, Weidner CI, Costa IG, Marioni RE, Ferreira MR, Deary IJ, Wagner W. DNA methylation levels at individual age-associated CpG sites can be indicative for life expectancy. Aging (Albany NY). 2016; 8:394–401. https://doi.org/10.18632/aging.100908 [PubMed]
- 13. Zhang Y, Wilson R, Heiss J, Breitling LP, Saum KU, Schöttker B, Holleczek B, Waldenberger M, Peters A, Brenner H. DNA methylation signatures in peripheral blood strongly predict all-cause mortality. Nat Commun. 2017; 8:14617. https://doi.org/10.1038/ncomms14617 [PubMed]
- 14. Gao X, Zhang Y, Breitling LP, Brenner H. Relationship of tobacco smoking and smoking-related DNA methylation with epigenetic age acceleration. Oncotarget. 2016; 7:46878–89. https://doi.org/10.18632/oncotarget.9795 [PubMed]
- 15. Freire-Aradas A, Phillips C, Mosquera-Miguel A, Girón-Santamaría L, Gómez-Tato A, Casares de Cal M, Álvarez-Dios J, Ansede-Bermejo J, Torres-Español M, Schneider PM, Pośpiech E, Branicki W, Carracedo Á, Lareu MV. Development of a methylation marker set for forensic age estimation using analysis of public methylation data and the Agena Bioscience EpiTYPER system. Forensic Sci Int Genet. 2016; 24:65–74. https://doi.org/10.1016/j.fsigen.2016.06.005 [PubMed]
- 16. Lee IM, Paffenbarger RS
Jr . Associations of light, moderate, and vigorous intensity physical activity with longevity. The Harvard Alumni Health Study. Am J Epidemiol. 2000; 151:293–99. https://doi.org/10.1093/oxfordjournals.aje.a010205 [PubMed] - 17. Gajewski AK, Poznańska A. Mortality of top athletes, actors and clergy in Poland: 1924-2000 follow-up study of the long term effect of physical activity. Eur J Epidemiol. 2008; 23:335–40. https://doi.org/10.1007/s10654-008-9237-3 [PubMed]
- 18. Teramoto M, Bungum TJ. Mortality and longevity of elite athletes. J Sci Med Sport. 2010; 13:410–16. https://doi.org/10.1016/j.jsams.2009.04.010 [PubMed]
- 19. Garatachea N, Santos-Lozano A, Sanchis-Gomar F, Fiuza-Luces C, Pareja-Galeano H, Emanuele E, Lucia A. Elite athletes live longer than the general population: a meta-analysis. Mayo Clin Proc. 2014; 89:1195–200. https://doi.org/10.1016/j.mayocp.2014.06.004 [PubMed]
- 20. Lemez S, Baker J. Do Elite Athletes Live Longer? A Systematic Review of Mortality and Longevity in Elite Athletes. Sports Med Open. 2015; 1:16. https://doi.org/10.1186/s40798-015-0024-x [PubMed]
- 21. Bacalini MG, Deelen J, Pirazzini C, De Cecco M, Giuliani C, Lanzarini C, Ravaioli F, Marasco E, van Heemst D, Suchiman HE, Slieker R, Giampieri E, Recchioni R, et al. Systemic Age-Associated DNA Hypermethylation of ELOVL2 Gene: In Vivo and In Vitro Evidences of a Cell Replication Process. J Gerontol A Biol Sci Med Sci. 2017; 72:1015–23. https://doi.org/10.1093/gerona/glw185 [PubMed]
- 22. Zbieć-Piekarska R, Spólnicka M, Kupiec T, Parys-Proszek A, Makowska Ż, Pałeczka A, Kucharczyk K, Płoski R, Branicki W. Development of a forensically useful age prediction method based on DNA methylation analysis. Forensic Sci Int Genet. 2015; 17:173–79. https://doi.org/10.1016/j.fsigen.2015.05.001 [PubMed]
- 23. Spólnicka M, Pośpiech E, Pepłońska B, Zbieć-Piekarska R, Makowska Ż, Pięta A, Karłowska-Pik J, Ziemkiewicz B, Wężyk M, Gasperowicz P, Bednarczuk T, Barcikowska M, Żekanowski C, et al. DNA methylation in ELOVL2 and C1orf132 correctly predicted chronological age of individuals from three disease groups. Int J Legal Med. 2018; 132:1–11 https://doi.org/10.1007/s00414-017-1636-0 [PubMed]
- 24. Lin Y, Gajewski A, Poznańska A. Examining mortality risk and rate of ageing among Polish Olympic athletes: a survival follow-up from 1924 to 2012. BMJ Open. 2016; 6:e010965. https://doi.org/10.1136/bmjopen-2015-010965 [PubMed]
- 25. Löllgen H, Böckenhoff A, Knapp G. Physical activity and all-cause mortality: an updated meta-analysis with different intensity categories. Int J Sports Med. 2009; 30:213–24. https://doi.org/10.1055/s-0028-1128150 [PubMed]
- 26. Jones MJ, Goodman SJ, Kobor MS. DNA methylation and healthy human aging. Aging Cell. 2015; 14:924–32. https://doi.org/10.1111/acel.12349 [PubMed]
- 27. Christiansen L, Lenart A, Tan Q, Vaupel JW, Aviv A, McGue M, Christensen K. DNA methylation age is associated with mortality in a longitudinal Danish twin study. Aging Cell. 2016; 15:149–54. https://doi.org/10.1111/acel.12421 [PubMed]
- 28. Cho S, Jung SE, Hong SR, Lee EH, Lee JH, Lee SD, Lee HY. Independent validation of DNA-based approaches for age prediction in blood. Forensic Sci Int Genet. 2017; 29:250–56. https://doi.org/10.1016/j.fsigen.2017.04.020 [PubMed]
- 29. Naue J, Hoefsloot HC, Mook OR, Rijlaarsdam-Hoekstra L, van der Zwalm MC, Henneman P, Kloosterman AD, Verschure PJ. Chronological age prediction based on DNA methylation: massive parallel sequencing and random forest regression. Forensic Sci Int Genet. 2017; 31:19–28. https://doi.org/10.1016/j.fsigen.2017.07.015 [PubMed]
- 30. Valiyeva F, Jiang F, Elmaadawi A, Moussa M, Yee SP, Raptis L, Izawa JI, Yang BB, Greenberg NM, Wang F, Xuan JW. Characterization of the oncogenic activity of the novel TRIM59 gene in mouse cancer models. Mol Cancer Ther. 2011; 10:1229–40. https://doi.org/10.1158/1535-7163.MCT-11-0077 [PubMed]
- 31. Shamma A, Takegami Y, Miki T, Kitajima S, Noda M, Obara T, Okamoto T, Takahashi C. Rb Regulates DNA damage response and cellular senescence through E2F-dependent suppression of N-ras isoprenylation. Cancer Cell. 2009; 15:255–69. https://doi.org/10.1016/j.ccr.2009.03.001 [PubMed]
- 32. Zhan W, Han T, Zhang C, Xie C, Gan M, Deng K, Fu M, Wang JB. TRIM59 Promotes the Proliferation and Migration of Non-Small Cell Lung Cancer Cells by Upregulating Cell Cycle Related Proteins. PLoS One. 2015; 10:e0142596. https://doi.org/10.1371/journal.pone.0142596 [PubMed]
- 33. Aierken G, Seyiti A, Alifu M, Kuerban G. Knockdown of Tripartite-59 (TRIM59) Inhibits Cellular Proliferation and Migration in Human Cervical Cancer Cells. Oncol Res. 2017; 25:381–88. https://doi.org/10.3727/096504016X14741511303522 [PubMed]
- 34. Hu SH, Zhao MJ, Wang WX, Xu CW, Wang GD. TRIM59 is a key regulator of growth and migration inrenal cell carcinoma. Cell Mol Biol. 2017; 63:68–74. https://doi.org/10.14715/cmb/2017.63.5.13 [PubMed]
- 35. Wu W, Chen J, Wu J, Lin J, Yang S, Yu H. Knockdown of tripartite motif-59 inhibits the malignant processes in human colorectal cancer cells. Oncol Rep. 2017; 38:2480–88. https://doi.org/10.3892/or.2017.5896 [PubMed]
- 36. Baker DJ, Alimirah F, van Deursen JM, Campisi J, Hildesheim J. Oncogenic senescence: a multi-functional perspective. Oncotarget. 2017; 8:27661–72. https://doi.org/10.18632/oncotarget.15742 [PubMed]
- 37. Tyner SD, Venkatachalam S, Choi J, Jones S, Ghebranious N, Igelmann H, Lu X, Soron G, Cooper B, Brayton C, Park SH, Thompson T, Karsenty G, et al. p53 mutant mice that display early ageing-associated phenotypes. Nature. 2002; 415:45–53. https://doi.org/10.1038/415045a [PubMed]
- 38. Raj D, Yin Z, Breur M, Doorduin J, Holtman IR, Olah M, Mantingh-Otter IJ, Van Dam D, De Deyn PP, den Dunnen W, Eggen BJ, Amor S, Boddeke E. Increased White Matter Inflammation in Aging- and Alzheimer’s Disease Brain. Front Mol Neurosci. 2017; 10:206. https://doi.org/10.3389/fnmol.2017.00206 [PubMed]
- 39. Fröhlich E, Wahl R. Thyroid Autoimmunity: Role of Anti-thyroid Antibodies in Thyroid and Extra-Thyroidal Diseases. Front Immunol. 2017; 8:521. https://doi.org/10.3389/fimmu.2017.00521 [PubMed]
- 40. Wei X, Yang R, Wang C, Jian X, Li L, Liu H, Yang G, Li Z. A novel role for the Krüppel-like factor 14 on macrophage inflammatory response and atherosclerosis development. Cardiovasc Pathol. 2017; 27:1–8. https://doi.org/10.1016/j.carpath.2016.11.003 [PubMed]
- 41. Xie W, Li L, Zheng XL, Yin WD, Tang CK. The role of Krüppel-like factor 14 in the pathogenesis of atherosclerosis. Atherosclerosis. 2017; 263:352–60. https://doi.org/10.1016/j.atherosclerosis.2017.06.011 [PubMed]