



Introduction
Aging is a multifactorial process characterized by a widespread loss of homeostasis, leading to gradual decrease in functional capacity at all levels of biological organization which, reduced resistance to environmental stresses and exponential increase in probability of death [1]. Evolutionarily conserved stress- and nutrient-responsive signaling pathways (such as growth hormone/IGF-1, NAD+/sirtuins, PI3K/AKT/mTOR, AMPK/mTOR and JNK) regulate homeostasis and longevity by coordinating metabolic functions, cell growth and proliferation, and stress responses throughout the organism [1,2]. A number of genetic, pharmacological or environmental interventions in activities of these pathways are known to induce beneficial pleiotropic effects on cell and extend lifespan in model organisms [3].
Recent experimental studies highlighted a potential role of hydrogen sulfide (H2S) in the regulation of lifespan and aging. Endogenous H2S is the product of metabolism of sulphur containing amino acids, methionine and cysteine in evolutionarily conserved transsulfuration pathway (TSP) [4,5]. The TSP is activated in response to such anti-aging intervention as dietary restriction (DR), and H2S apparently mediates the pleiotropic DR benefits including longevity and stress resistance in different models, from yeast, worms and flies to mice [4,5]. H2S affects aging by sulfhydration of target proteins and influencing cellular signaling pathways [6]. H2S influences aging-related processes such as cellular bioenergetics, autophagy, inflammation, oxidative stress, proliferation and differentiation of stem cell, cellular senescence, cell death and cellular metabolism [6–10].
In animals and particular in Drosophila melanogaster, H2S is produced by two key enzymes of the TSP, cystathionine β-synthase (CBS) and cystathionine γ-lyase (CSE) [4,11]. The lifespan extension under DR in Drosophila was associated with increased CBS activity [4]. Overexpression of CBS homologs in nematodes and flies increased lifespan independent of diet, while RNAi-mediated knocking down of the same genes or inhibition of the CSE in Drosophila using propargylglycine limited or abrogated DR-mediated lifespan extension [4,12]. Mice with loss-of-function mutations in CBS and CSE genes demonstrated increased plasma homocysteine level, growth retardation and significantly reduced lifespan (about 5 and 12 weeks, respectively) [13,14]. Exposing to H2S-containing atmosphere has been shown to induce thermotolerance and increase longevity in Caenorhabditis elegans [15]. Chronic treatment of C. elegans with an exogenous H2S (via the slow-releasing donor GYY4137) extended median survival and increased tolerance towards oxidative and endoplasmic reticulum stress [16]. Thus, both exogenous and endogenous H2S are important for stress resistance, longevity and health of the model organisms. However, effects of the overexpression of the genes controlling endogenous H2S synthesis on the stress resistance, lifespan and locomotor activity are not investigated fully. In the present work, we for the first time investigated whether ubiquitous transgenic overexpression of CSE (coding mitochondria- and cytoplasm-located forms of CSE) affect the resistance to various stress factors (oxidative stress, hyperthermia and combination of arid and food-free conditions), longevity and locomotor activity in Drosophilamelanogaster and compared obtained results with the effects of pro-longevity gene CBS overexpression.
Results
Overexpression of transgenes
Using FlyBase search [17] we identified 3 genes responsible for H2S production in Drosophila, CG12264 coding for mitochondria-located enzyme with CSE activity [18], CG5345 that codes the cytoplasm-located orthologue of CSE [11] and CG1753 that is the single homolog of CBS [4]. Using cDNA of these genes 3 UAS lines were generated (Table S1).
In their previous report, Kabil [4] showed that ubiquitous (driven by tubulin-GAL4) or neuron-specific (driven by Elav-GAL4) transgenic overexpression of CBS enhanced Drosophila longevity, while CBS overexpression in abdominal fat body (driven by GeneSwitch S1-106) and in gut (driven by GeneSwitch TIGS-2) had no effect on lifespan [4]. To further study the role of genes affecting H2S production in longevity of Drosophila, we tested whether ubiquitous overexpression of CBS and CSE under the control of da-GAL4 driver also increase resistance to stress factors, prolong lifespan and improve locomotor performance.
The expression levels of transgenes were examined by qRT-PCR assays before further analysis (Figure S1). We found 2.4-, 6.1- and 1.9-fold induction (p<0.05) of CSE[LD22661], CSE[LD22255], CBS[LD21426], respectively compared with their isogenic UAS controls (Figure S1, Table S3). The level of transgene expression was found to depend on the UAS transgene.
Lifespan
The activation of constitutive CSE[LD22661] overexpression leaded to prolongation by 15.6% relative to da-GAL4 parental control line (p<0.05). At the same time, we observed the decline of median and maximum lifespan in males with CSE[LD22661] overexpression compared with maternal control line UAS-CSE[LD22661] by 24.4% (p<0.0001) and 3.1% (p<0.01), respectively (Table 1, Figure 1A). The increase in median lifespan by 11.6% (p<0.05) was caused by upregulation of CSE[LD22255] relative to da-GAL4 parental control line, but the lifespan differences in comparison with the UAS-CSE[LD22255] maternal control line were not statistically significant (Table 1, Figure 1B). Thus, overexpression of both forms of the CSE gene did not lead to an increase in lifespan relative to the both paternal and maternal control lines. As we know the effect of CSE overexpression on the longevity was not previously studied. As it was established by Kabil [4] pharmacological inhibition of the CSE using propargylglycine reversed lifespan extension induced by DR, and there was no effect of CSE inhibition on fully fed flies [4]. Overexpression of CSE in Drosophila SCA3-model restored protein persulfidation, decreased oxidative stress, dampened the immune response and improved SCA3-associated tissue damage and neurodegeneration [19]. Despite these published beneficial effects on aging-related processes, we did not observe the positive effect of CSE overexpression on the lifespan.
Table 1. The effects of CSE and CBS overexpression on lifespan.
| Genotype | Variant | M (days) | dM (%) | Log-Rank Test (p) | 90% (days) | d90% (%) | Wang-Allison Test (p) | MRDT (days) | dMRDT (%) | n | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| da-GAL4>UAS-CSE[LD22661] | overexpression | 45 | 65 | 9 | 270 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| da-GAL4 | control (parental) | 38 | -15.6 | p<0.05 (0.0305) | 63 | -3.1 | p>0.05 (0.051) | 13.6 | 51.1 | 289 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| UAS-CSE[LD22661] | control (maternal) | 56 | 24.4 | p<0.0001 (4.59e-08) | 67 | 3.1 | p<0.01 (0.009) | 7.5 | -16.7 | 243 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| da-GAL4>UAS-CSE[LD22255] | overexpression | 43 | 61 | 12 | 281 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| da-GAL4 | control (parental) | 38 | -11.6 | p<0.05 (0.0305) | 63 | 3.3 | p>0.05 (0.051) | 13.6 | 13.3 | 289 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| UAS-CSE[LD22255] | control (maternal) | 44 | 2.3 | p>0.05 (0.196) | 67 | 9.8 | p>0.05 (0.799) | 9.6 | -20 | 270 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| da-GAL4>UAS-CBS[LD21426] | overexpression | 53 | 72 | 11.2 | 278 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| da-GAL4 | control (parental) | 38 | -28.3 | p<0.05 (0.0305) | 63 | -12.5 | p>0.05 (0.051) | 13.6 | 21.4 | 289 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| UAS-CBS[LD21426] | control (maternal) | 45 | -15.1 | p>0.05 (0.0707) | 67 | -6.9 | p<0.05 (0.013) | 13.1 | 17 | 299 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| M - median lifespan; 90% - age of 90% mortality (maximum lifespan); MRDT - mortality rate doubling time; dM, d90%, dMRDT - differences between median lifespan, age of 90% mortality, and MRDT of control and experimental flies, respectively; n - number of flies. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
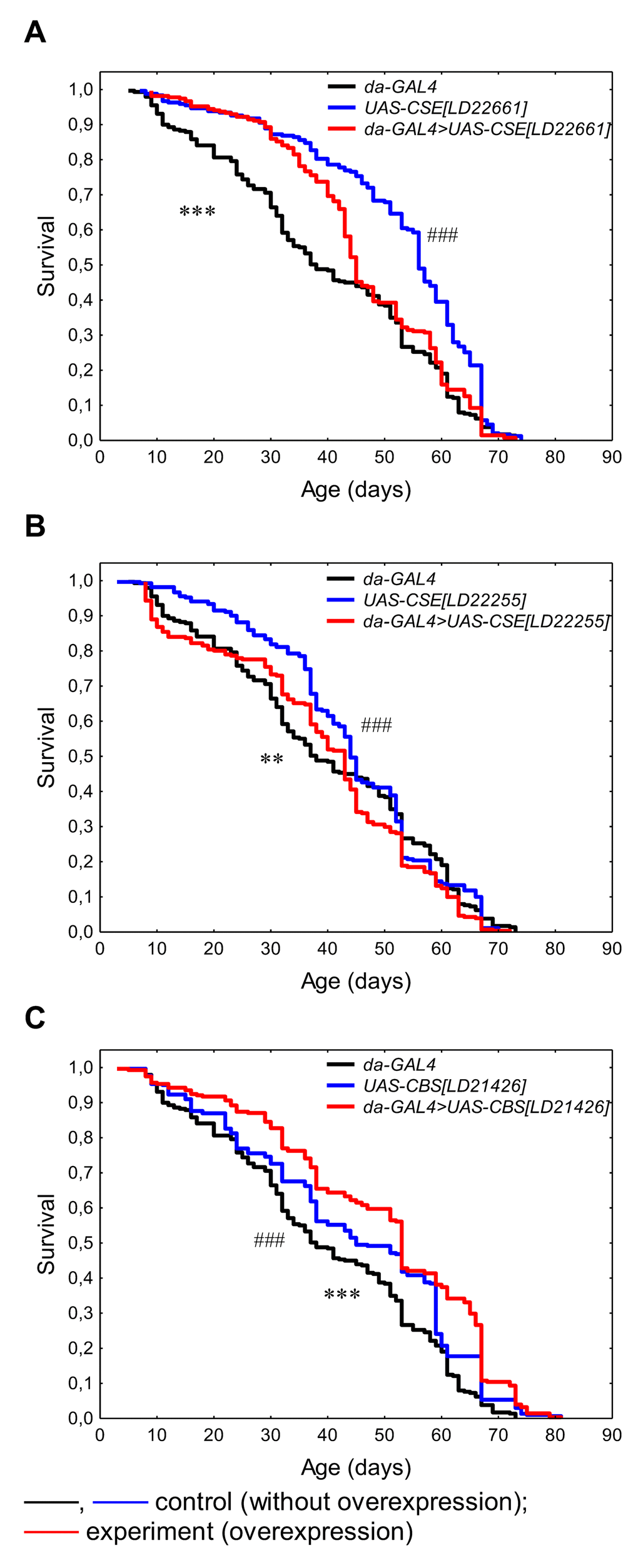
Figure 1. The effects of constitutive ubiquitous overexpression of CSE[LD22661] (A), CSE[LD22255] (B), CBS[LD21426] (C) on lifespan. *p<0.05, **p<0.01, ***p<0.001, da-GAL4>UAS vs da-GAL4; #p<0.05, ##p<0.01, ###p<0.001, da-GAL4>UAS vs UAS, Kolmogorov-Smirnov test.
The CBS[LD21426] overexpression increased median (by 15.1-28.3%, p<0.05) and maximum (by 6.9-12.5%, p<0.05) lifespan relative to UAS-CBS[LD21426] maternal and da-GAL4 parental control lines, respectively (Table 1, Figure 1C). In addition, CBS[LD21426] overexpression decreased MRDT (by 17% and 21.4%, respectively) suggesting that geroprotective effect of CBS in associated with prolongation of healthspan. The observed lifespan extending effect of CBS ubiquitous overexpression is consistent with the results of Kabil [4].
Stress resistance
We also analyzed the resistance to different kinds of stress factors of flies with overexpression of the CBS and CSE genes and the control lines. The survival curves and times of 25%, 50%, 75% and 90% mortality in flies under conditions of paraquat (oxidative stress), hyperthermia, and combination of arid and food-free conditions were estimated (Table S4, Figure 2). The CSE[LD22661] overexpression leaded to marginal decrease (p<0.05) of resistance to paraquat (Figure 2A), while CSE[LD22255] and CBS[LD21426] overexpression had no effect (p>0.05) on resistance to oxidative stress (Figures 2B and C). According to published data exogenous and endogenous H2S has been shown to improve the capacity of model animals to survive in conditions of oxidative stress. Pharmacological H2S donor (GYY4137) protected wild-type C. elegans against paraquat poisoning [20]. In D. melanogaster the X-ray radiation-induced embryonic mortality were found to be reduced by H2S treatment [21]. Our data demonstrated that overexpression of the CBS and CSE is not sufficient to increase tolerance to oxidative stress in Drosophila.
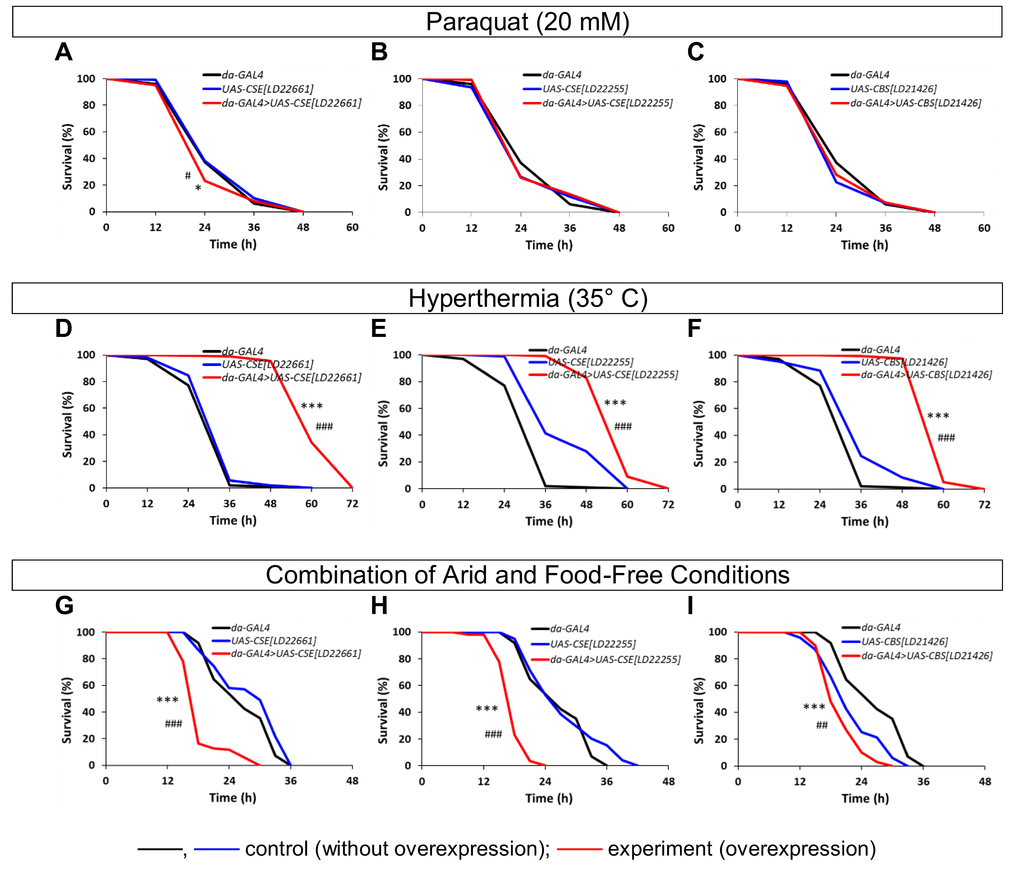
Figure 2. The effects of constitutive ubiquitous overexpression of CSE and CBS genes on resistance to stress factors (paraquat (A-C), hyperthermia (D-F) and combination of arid and food-free conditions (G-I)). *p<0.05, **p<0.01, ***p<0.001, Fisher's exact test (da-GAL4>UAS vs da-GAL4); #p<0.05, ##p<0.01, ###p<0.001, Fisher's exact test (da-GAL4>UAS vs UAS).
We also found that CSE[LD22661], CSE[LD22255] and CBS[LD21426] upregulation provoked significant elevation of resistance to hyperthermia (Figures 2D-F). The obtained results are consistent with the published effects of exogenous H2S. Exposure of C. elegans increased concentrations of H2S has been shown to enhance thermotolerance [15].
The significant decrease of resistance to combination of arid and food-free conditions was observed (p<0.001, Figures 2G-I). As known from the published data exposure to H2S emanated from a K2S donor significantly increased survival of flies under arid and food-free conditions, but not under humid and food-free conditions, demonstrating that H2S plays a role in desiccation tolerance but not in nutritional stress alleviation [22]. On the other hand, overexpression of the CBS, was sufficient to promote fat deposition in flies [4]. It is well known that Drosophila are better able to resist starvation by sequestering more body lipid reserves [23]. Thus the final role of endogenous H2S in the formation of resistance to starvation and desiccation has to be clarified.
Locomotor activity
The influence of CBS and CSE overexpression on locomotor activity was estimated in flies at the age of 5, 15 and 25 days (Figure 3). It was found that UAS-CSE[LD22661] overexpression induced decline of spontaneous locomotor activity in the first part of life and increase of this parameter in the second part of male’s life (Figure 3A). In UAS-CSE[LD22255] overexpressing males locomotor activity is higher than in control group in the first and second third of life but it is lower in the last one. (Figure 3B). The UAS-CBS[LD21426] overexpressing males were characterized by high longlife locomotor activity (Figure 3C).
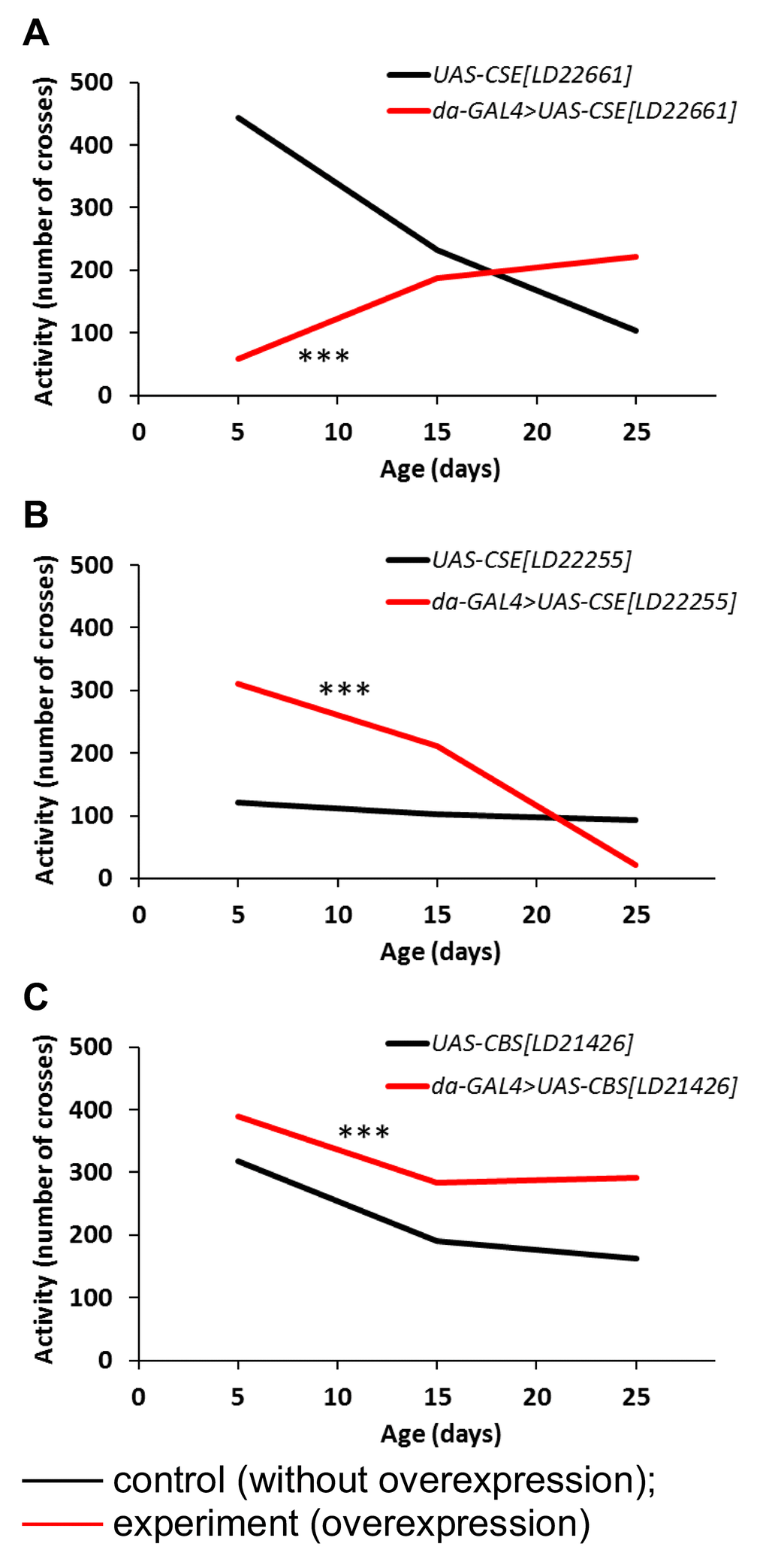
Figure 3. The effects of constitutive ubiquitous overexpression of CSE[LD22661] (A), CSE[LD22255] (B), CBS[LD21426] (C) on age-dependent changes in spontaneous locomotor activity. Locomotor activity was defined as averaged number of sensor crosses during 3 min by 30 flies. *р<0.05, **р<0.01, ***р<0.001, χ2.
Discussion
Therefore, we analyzed the effects of constitutive ubiquitous overexpression of CSE[LD22661], CSE[LD22255] and CBS[LD21426] on the Drosophila melanogaster lifespan, stress resistance (paraquat (oxidative stress), hyperthermia, and combination of arid and food-free conditions) and healthspan (locomotor activity). We showed that the overexpression of cystathionine β-synthase gene (CBS[LD21426]) was characterized by the highest geroprotective efficiency. The CBS[LD21426] upregulation causes increase of median lifespan (by 12.5%) and maximum lifespan (by 6.9%) in males (p<0.05) relative to parental lines. At the same time the overexpression of this gene is resulted in increased thermotolerance (p<0.05) and locomotor activity (p<0.001), but decline resistance to arid and food-free conditions (p<0.001). The overexpression of cystathionine γ-lyase genes (CSE[LD22661] and CSE[LD22255]) did not lead to geroprotective effects and caused decline of resistance to arid and food-free conditions (p<0.001), but simultaneously, the elevation of resistance to hyperthermia was observed (p<0.001). The differences in the effects of CBS and CSE genes overexpression on the lifespan can be partly explained by different roles of the enzymes encoded by these genes in the TSP. While the CBS catalyzes the first and rate-determining step in the TSP, which involves the pyridoxal 5′-phosphate-dependent condensation of serine and homocysteine to form cystathionine and H2S, the CSE uses cystathionine as a substrate for cysteine and H2S production [4]. Thus, CBS can partially limit the effects of CSE overexpression on H2S production and lifespan.
We showed that CBS and CSE overexpression induce elevated resistance to hyperthermia. And in case of CBS overexpression thermotolerance is accompanied by an increase in lifespan. The common role of H2S in thermotolerance and longevity was revealed in publication of Miller and Roth [15]. They showed that elevated resistance to hyperthermia and increased lifespan in C. elegans after exposure to H2S require activity of silent information regulator 2 (SIR2) but are independent of the insulin signaling pathway, mitochondrial dysfunction, and caloric restriction [15]. Our results confirm their assumption that sirtuin-dependent mechanisms by which H2S increases thermotolerance and lifespan in nematodes may be evolutionary conserved. In addition the overexpression of dSir2 extended lifespan in different model organisms including Saccharomyces cerevisiae [24], C. elegans [25], D. melanogaster [26] and Mus musculus males [27]. We also found that overexpression of CBS[LD21426] and CSE[LD22255] is associated with the increase of locomotor activity. This effect can be explained by stimulation of cellular bioenergetics with H2S produced by CSE and CBS enzymes [9].
Taken together, these data demonstrate that of CBS and CSE overexpression implicated in the control of stress resistance and aging process. The transgene overexpression of the CSE and CBS in Drosophila induce similar effects on stress-resistance and locomotor activity, however lifespan extending effect was revealed for CBS overexpression only. The positive effects of CBS and CSE overexpression on lifespan, stress resistance and locomotor activity may be associated with H2S-mediated modulation of autophagy, inflammation, oxidative stress, cell death, cellular metabolism and cellular bioenergetics (Figure 4). It should be taken into account that the effects of H2S are dependent on the experimental system, model organism, concentration, and may vary from beneficial to harmful [6–10].
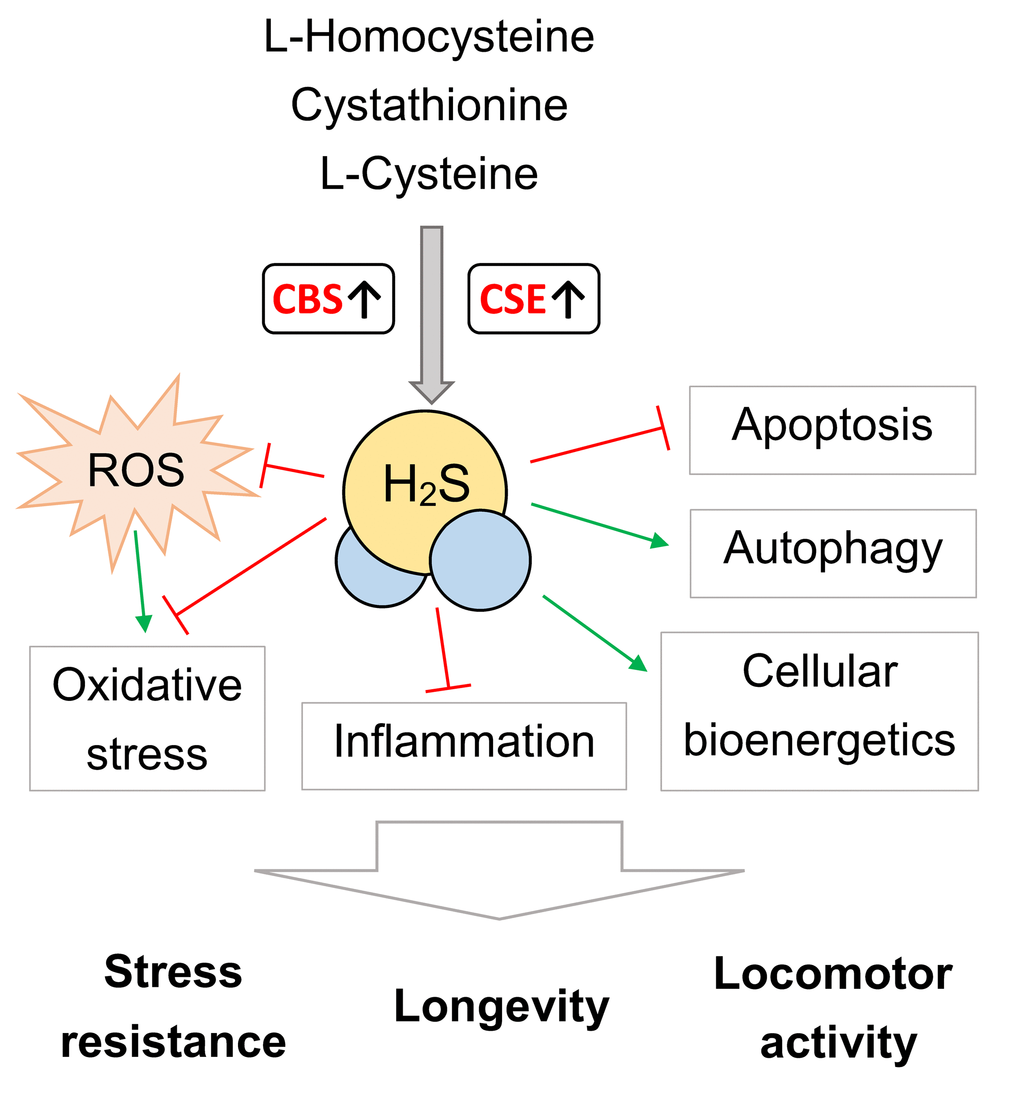
Figure 4. The possible mechanisms of CBS and CSE overexpression on lifespan, stress resistance and locomotor activity. ROS – reactive oxygen species. Please note that this scheme serve as generalized illustration and the mechanisms are dependent on the experimental system and model organism used. For detailed description of these mechanisms see [4–10].
Methods
Drosophila melanogaster lines
Full length sequenced cDNA Clones from Drosophila Genomics Resource Center (DGRC, USA) were used for UAS lines creation. Cloning of cDNAs into pUAST-attB vector, sequence verification of each clone, and φC31 integrase-mediated site-specific germline transformation via embryo injection were ordered in GenetiVision (Houston, Texas, USA) (Table S1). UAS-CSE[LD22661] contains an additional copy of CG12264 gene coding for mitochondria-located enzyme with CSE activity [18]. UAS-CSE[LD22255] carries an additional copy of CG5345 gene that is the Drosophila orthologue of CSE (cytoplasm-located) [11]. UAS-CBS[LD21426] contains an additional copy of CG1753 gene that is the single homolog of CBS in Drosophila [4].
The GAL4 line (GAL4 driver) was used for overexpression of UAS transgenes. da-GAL4 expresses GAL4 ubiquitously and strongly under the control of daughterless [28]. This driver expresses throughout development and in most adult tissues [29]. da-GAL4 line was kindly provided by Dr. Laurent Seroude (Queen’s University, Kingston, Canada).
In order to match the genetic background of UAS and GAL4 lines they were backcrossed into white-eyed standard w1118 (#3605, Bloomington Drosophila Stock Center, USA) for 6-8 times.
Activation of transgenes overexpression
The GAL4-UAS binary transgenic system was used to activate the expression of the studied genes [30]. We used constitutively active (da-GAL4) driver of GAL4 that activates the gene overexpression ubiquitously. The flies with overexpressed transgenes were obtained by mating the females from UAS lines and males from GAL4 driver lines.
Quantitative real time PCR
To confirm overexpression of studied genes in the whole body 10-15 imagoes were used in every variant of the experiment. Gene expression levels were analyzed in flies at the age of 5-6 days after imago hatching. Experiments were performed in 3-4 replicates. Whole flies or heads were homogenized with the Silent Crusher-S homogenizer (Heidolph, Germany) in TRIzol Reagent (Invitrogen, USA). RNA was separated using BCP (Invitrogen, USA), in accordance with the manufacturer’s protocol. To test that RNA samples were DNA-free, control PCR experiments without the reverse transcription step were performed with primers for the β-Tubulin gene. Reverse transcription was performed using an Oligo(dT)20 primer (Invitrogen, USA) and SuperScript III Reverse Transcriptase (Invitrogen, USA), according to manufacturer’s instructions.
Quantitative real-time PCR (qRT-PCR) assays were performed using SYBRGreen PCR Master Mix (Applied Biosystems, USA). The list of primers is presented in Table S2. All reactions were performed using a CFX96 real-time PCR detection system (Bio-Rad Laboratories, USA). The thermal cycle conditions were: initial denaturation step at 95°С for 10 min, followed by 50 cycles of 95°С for 15 s (denaturation), 60°С for 30 s (annealing) and 60°С for 30 s (elongation). Expression levels were normalized against the housekeeping gene β-Tubulin. All target genes and β-Tubulin were amplified in separate PCR tubes. Four measurements were performed for each version of the experiment.
Lifespan analysis
Control and experimental fly males were collected during 24 h after imago hatching and maintained in constant conditions on a sugar-yeast medium at 25°C and 60% humidity in a 12:12 h light-dark cycle. The flies were housed in Drosophila vials at a density of 30 individuals per vial, with 5 vials per experimental variant. Dead flies were recorded daily. Fresh medium vials were provided two times per week. Experiments were performed in 2 replicates. The median lifespan, the age of 90% mortality (maximum lifespan), and the mortality rate doubling time (MRDT) were calculated.
Analysis of locomotor activity
Age-dependent changes in spontaneous locomotor activity were measured using the Drosophila Population Monitor (TriKinetics Inc., USA). Activity was defined as total number of sensor crosses during 3 min by 30 flies. Measurements were carried out every 10 days, starting from the age of 5 days, until 30 flies were alive in control and experimental variants (at the age of 5, 15 and 25 days).
Stress resistance analysis
The resistance to acute stress conditions was evaluated in 5 days old imagoes. The exposure to paraquat (oxidative stress), hyperthermia, and combination of arid and food-free conditions were used. Before as exposed to paraquat flies were deprived of food and water for 3 h and then transferred into vials containing 3-layer filter paper saturated with 300 ml of a 5% sucrose solution with 20 mM paraquat (Sigma-Aldrich, USA). At the variant of hyperthermia, flies were kept in vials on agar-yeast medium at 35 °C. For combination of arid and food-free conditions flies were placed into empty vials. Flies were transferred into new vials every two days, and kept under stressful conditions until the death.
For evaluating the resistances to oxidative stress and heat shock the amount of dead flies were counted two times per day. For evaluating the resistances to combination of arid and food-free conditions dead flies were counted every 3 hours. The survival time for 25%, 50%, 75% and 90% of populations were estimated.
Statistical analysis
Relative levels of expression were calculated using 2-ΔΔСt method [31]. ΔΔCt was calculated according to equation ΔΔCt = ΔCt (experiment) – ΔCt (control), where ΔCt = Ct (target gene) – Ct (β-Tubulin). Statistical significance of expression differences was estimated using Mann-Whitney U-test [32]. To compare the statistical differences in survival functions and median lifespan between control and experimental groups, the modified Kolmogorov-Smirnov and log-rank [33] tests were used, respectively [34,35]. A Wang-Allison test was used to estimate the differences in the age of 90% mortality [36]. The χ2 test was used for locomotor activity and fecundity analysis. To assess the statistical significance of differences in resistance to stress factors, the Fisher's exact test was used [37]. Statistical analyses of the data were performed using R, version 2.15.1 (The R Foundation) and OASIS 2: Online Application for Survival Analysis 2 [37].
Acknowledgements
We are grateful to Dr. Laurent Seroude (Queen’s University, Canada) and the Bloomington stock center (Indiana University, USA) for providing the Drosophila strains, to GenetiVision Corporation (Houston, Texas, USA) for transgenic fly services, to the Drosophila Genomics Resource Center (Indiana University, USA) for cDNA clones. We thank the Institute of Biology of Komi Science Center for assistance in the experiments with Drosophila and Insilico Medicine, Inc. and MIPT for assistance in data analysis.
Conflicts of Interest
Author Alex Zhavoronkov is employed by Insilico Medicine, Inc, a longevity biotechnology and artificial intelligence company. All other authors declare no competing interests.
Funding
This work was supported by the Russian Science Foundation grant N 17-74-30030.
References
- 1. Wang L, Karpac J, Jasper H. Promoting longevity by maintaining metabolic and proliferative homeostasis. J Exp Biol. 2014; 217:109–18. https://doi.org/10.1242/jeb.089920 [PubMed]
- 2. Pan H, Finkel T. Key proteins and pathways that regulate lifespan. J Biol Chem. 2017; 292:6452–60. https://doi.org/10.1074/jbc.R116.771915 [PubMed]
- 3. Moskalev A, Shaposhnikov M, Solovev I. Studying the geroprotective effects of inhibitors suppressing aging-associated signaling cascades in model organisms. Medical news of North Caucasus. 2017; 12: 342-7. .
- 4. Kabil H, Kabil O, Banerjee R, Harshman LG, Pletcher SD. Increased transsulfuration mediates longevity and dietary restriction in Drosophila.. Proc Natl Acad Sci USA. 2011; 108:16831–36. https://doi.org/10.1073/pnas.1102008108 [PubMed]
- 5. Hine C, Harputlugil E, Zhang Y, Ruckenstuhl C, Lee BC, Brace L, Longchamp A, Treviño-Villarreal JH, Mejia P, Ozaki CK, Wang R, Gladyshev VN, Madeo F, et al. Endogenous hydrogen sulfide production is essential for dietary restriction benefits. Cell. 2015; 160:132–44. https://doi.org/10.1016/j.cell.2014.11.048 [PubMed]
- 6. Sen N. Functional and Molecular Insights of Hydrogen Sulfide Signaling and Protein Sulfhydration. J Mol Biol. 2017; 429:543–61. https://doi.org/10.1016/j.jmb.2016.12.015 [PubMed]
- 7. Zhang Y, Tang ZH, Ren Z, Qu SL, Liu MH, Liu LS, Jiang ZS. Hydrogen sulfide, the next potent preventive and therapeutic agent in aging and age-associated diseases. Mol Cell Biol. 2013; 33:1104–13. https://doi.org/10.1128/MCB.01215-12 [PubMed]
- 8. Wu D, Wang H, Teng T, Duan S, Ji A, Li Y. Hydrogen sulfide and autophagy: A double edged sword. Pharmacol Res. 2018; 131:120–27. https://doi.org/10.1016/j.phrs.2018.03.002 [PubMed]
- 9. Szabo C, Ransy C, Módis K, Andriamihaja M, Murghes B, Coletta C, Olah G, Yanagi K, Bouillaud F. Regulation of mitochondrial bioenergetic function by hydrogen sulfide. Part I. Biochemical and physiological mechanisms. Br J Pharmacol. 2014; 171:2099–122. https://doi.org/10.1111/bph.12369 [PubMed]
- 10. Latorre E, Torregrossa R, Wood ME, Whiteman M, Harries LW. Mitochondria-targeted hydrogen sulfide attenuates endothelial senescence by selective induction of splicing factors HNRNPD and SRSF2.. Aging (Albany NY). 2018; 10:1666–81. https://doi.org/10.18632/aging.101500 [PubMed]
- 11. Mathew ND, Schlipalius DI, Ebert PR. Sulfurous gases as biological messengers and toxins: comparative genetics of their metabolism in model organisms. J Toxicol. 2011; 2011:394970. https://doi.org/10.1155/2011/394970 [PubMed]
- 12. Hine C, Mitchell JR. Calorie restriction and methionine restriction in control of endogenous hydrogen sulfide production by the transsulfuration pathway. Exp Gerontol. 2015; 68:26–32. https://doi.org/10.1016/j.exger.2014.12.010 [PubMed]
- 13. Mani S, Yang G, Wang R. A critical life-supporting role for cystathionine γ-lyase in the absence of dietary cysteine supply. Free Radic Biol Med. 2011; 50:1280–87. https://doi.org/10.1016/j.freeradbiomed.2011.01.038 [PubMed]
- 14. Watanabe M, Osada J, Aratani Y, Kluckman K, Reddick R, Malinow MR, Maeda N. Mice deficient in cystathionine beta-synthase: animal models for mild and severe homocyst(e)inemia. Proc Natl Acad Sci USA. 1995; 92:1585–89. https://doi.org/10.1073/pnas.92.5.1585 [PubMed]
- 15. Miller DL, Roth MB. Hydrogen sulfide increases thermotolerance and lifespan in Caenorhabditis elegans.. Proc Natl Acad Sci USA. 2007; 104:20618–22. https://doi.org/10.1073/pnas.0710191104 [PubMed]
- 16. Qabazard B, Ahmed S, Li L, Arlt VM, Moore PK, Stürzenbaum SR. C. elegans aging is modulated by hydrogen sulfide and the sulfhydrylase/cysteine synthase cysl-2.. PLoS One. 2013; 8:e80135. https://doi.org/10.1371/journal.pone.0080135 [PubMed]
- 17. Gramates LS, Marygold SJ, Santos GD, Urbano JM, Antonazzo G, Matthews BB, Rey AJ, Tabone CJ, Crosby MA, Emmert DB, Falls K, Goodman JL, Hu Y, et al, and the FlyBase Consortium. FlyBase at 25: looking to the future. Nucleic Acids Res. 2017; 45:D663–71. https://doi.org/10.1093/nar/gkw1016 [PubMed]
- 18. Lyne R, Smith R, Rutherford K, Wakeling M, Varley A, Guillier F, Janssens H, Ji W, Mclaren P, North P, Rana D, Riley T, Sullivan J, et al. FlyMine: an integrated database for Drosophila and Anopheles genomics. Genome Biol. 2007; 8:R129. https://doi.org/10.1186/gb-2007-8-7-r129 [PubMed]
- 19. Snijder PM, Baratashvili M, Grzeschik NA, Leuvenink HG, Kuijpers L, Huitema S, Schaap O, Giepmans BN, Kuipers J, Miljkovic JL, Mitrovic A, Bos EM, Szabo C, et al. Overexpression of cystathionine gamma-lyase suppresses detrimental effects of spinocerebellar ataxia type 3. Mol Med. 2015; 21:1. https://doi.org/10.2119/molmed.2015.00221 [PubMed]
- 20. Qabazard B, Li L, Gruber J, Peh MT, Ng LF, Kumar SD, Rose P, Tan CH, Dymock BW, Wei F, Swain SC, Halliwell B, Stürzenbaum SR, Moore PK. Hydrogen sulfide is an endogenous regulator of aging in Caenorhabditis elegans.. Antioxid Redox Signal. 2014; 20:2621–30. https://doi.org/10.1089/ars.2013.5448 [PubMed]
- 21. Matter BE, Würgler FE, Ulrich H. On the radioprotective effect of hydrogen sulphide in Drosophila melanogaster.. Int J Radiat Biol Relat Stud Phys Chem Med. 1969; 15:557–62. https://doi.org/10.1080/09553006914550851 [PubMed]
- 22. Zhong JF, Wang SP, Shi XQ, Mu LL, Li GQ. Hydrogen sulfide exposure increases desiccation tolerance in Drosophila melanogaster.. J Insect Physiol. 2010; 56:1777–82. https://doi.org/10.1016/j.jinsphys.2010.07.009 [PubMed]
- 23. Pum LK, Taehwan J. Exploring the nutritional basis of starvation resistance in Drosophila melanogaster.. Funct Ecol. 2014; 28:1144–55. https://doi.org/10.1111/1365-2435.12247
- 24. Kaeberlein M, McVey M, Guarente L. The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes Dev. 1999; 13:2570–80. https://doi.org/10.1101/gad.13.19.2570 [PubMed]
- 25. Tissenbaum HA, Guarente L. Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans.. Nature. 2001; 410:227–30. https://doi.org/10.1038/35065638 [PubMed]
- 26. Whitaker R, Faulkner S, Miyokawa R, Burhenn L, Henriksen M, Wood JG, Helfand SL. Increased expression of Drosophila Sir2 extends life span in a dose-dependent manner. Aging (Albany NY). 2013; 5:682–91. https://doi.org/10.18632/aging.100599 [PubMed]
- 27. Kanfi Y, Naiman S, Amir G, Peshti V, Zinman G, Nahum L, Bar-Joseph Z, Cohen HY. The sirtuin SIRT6 regulates lifespan in male mice. Nature. 2012; 483:218–21. https://doi.org/10.1038/nature10815 [PubMed]
- 28. Wodarz A, Hinz U, Engelbert M, Knust E. Expression of crumbs confers apical character on plasma membrane domains of ectodermal epithelia of Drosophila.. Cell. 1995; 82:67–76. https://doi.org/10.1016/0092-8674(95)90053-5 [PubMed]
- 29. Smith JE
3rd , Cronmiller C. The Drosophila daughterless gene autoregulates and is controlled by both positive and negative cis regulation. Development. 2001; 128:4705–14. [PubMed] - 30. Tower J. Transgenic methods for increasing Drosophila life span. Mech Ageing Dev. 2000; 118:1–14. https://doi.org/10.1016/S0047-6374(00)00152-4 [PubMed]
- 31. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001; 25:402–08. https://doi.org/10.1006/meth.2001.1262 [PubMed]
- 32. Mann HB, Whitney DR. On a Test of Whether one of Two Random Variables is Stochastically Larger than the Other. Ann Math Stat. 1947; 18:50–60. https://doi.org/10.1214/aoms/1177730491
- 33. Mantel N. Evaluation of survival data and two new rank order statistics arising in its consideration. Cancer Chemother Rep. 1966; 50:163–70. [PubMed]
- 34. Breslow N. A generalized Kruskal-Wallis test for comparing K samples subject to unequal patterns of censorship. Biometrika. 1970; 57:579–94. https://doi.org/10.1093/biomet/57.3.579
- 35. Fleming TR, O’Fallon JR, O’Brien PC, Harrington DP. Modified Kolmogorov-Smirnov test procedures with application to arbitrarily right-censored data. Biometrics. 1980; 36:607–25. https://doi.org/10.2307/2556114
- 36. Wang C, Li Q, Redden DT, Weindruch R, Allison DB. Statistical methods for testing effects on “maximum lifespan”. Mech Ageing Dev. 2004; 125:629–32. https://doi.org/10.1016/j.mad.2004.07.003 [PubMed]
- 37. Han SK, Lee D, Lee H, Kim D, Son HG, Yang JS, Lee SV, Kim S. OASIS 2: online application for survival analysis 2 with features for the analysis of maximal lifespan and healthspan in aging research. Oncotarget. 2016; 7:56147–52. https://doi.org/10.18632/oncotarget.11269 [PubMed]