



Introduction
Maintaining a healthy skeletal muscle mass across the lifecourse is essential to preserve locomotory function and whole-body metabolic health [1,2]. Accordingly, progressive muscle wasting such as with ageing (‘sarcopenia’) is associated with increased risk of falls, metabolic diseases and mortality [3,4]. Resistance exercise (RE) training (RET) represents the most effective lifestyle intervention for increasing/maintaining muscle mass and function across the lifespan [5]. One reason it is so important to optimise RET-strategies for maximising muscle mass gains in older age is because older adults exhibit blunted protein synthetic [6,7] and hypertrophic [8] responses to RE/T (commonly termed anabolic resistance). There thus remains a significant need to optimise anabolic responses to RE in older adults and to understand the molecular basis of any adaptive deficits.
Conventional RE involves both shortening (concentric, CON) and lengthening (eccentric, ECC) contractions, which induce distinct architectural adaptations. Indeed, adaptations to CON favour increases in pennation angle while ECC favours increases in fascicle length [9,10]. Chronically, ECC RET may also lead to greater muscle hypertrophy and strength gains versus CON RET [11]. As such, it is being increasingly claimed that ECC-type RET represents an intervention with potential to facilitate muscle maintenance in ageing [12]. Nonetheless, of the limited studies comparing adaptations to conventional RET (i.e. CON+ECC) versus ECC in older adults, conflicting results suggest that isolated ECC either augments [13,14] or confers no additional benefits [15–17] in total muscle mass and/or isometric leg strength of older adults (versus CON or conventional RET).
Transcriptomic responses to acute ECC and CON exercise have also demonstrated specific molecular regulation. For example, in healthy young adults, combined ECC+CON (versus CON alone) uniquely upregulated 28 genes involved in ontologies relating to inflammatory processes, energy metabolism and extracellular matrix function, with no apparent downregulated genes [18]. In a second study, up to 24 h post-exercise, ECC versus CON differentially regulated 51 transcripts involved in processes of protein synthesis, stress responses and sarcolemmal structure [19]. However, just 8 and 11 genes were uniquely regulated by ECC at 3 and 6 h post-exercise, respectively [19]. Transcriptional responses to ECC/CON have not been directly compared in older adults. Nonetheless, 4 h after traditional RE loading (i.e. combined ECC+CON) older versus younger muscle displayed 595 differentially regulated transcripts associated with various signalling processes [20]. Similarly, 318 “age-specific” genes were uniquely expressed 24 h post-RE, enriched for inflammatory, immune system, protein degradation and stress response-related pathways [21]. Despite providing seminal insight, these transcriptomic studies have limitations including i) lack of baseline comparison [18], ii) variable post-exercise time frames (4-8 h [18],), iii) inability to distinguish ECC effects from CON [18], iv) low subject N (n=3 [18,19]), v) inconsistency between ECC- and CON-unique transcriptional profiles precluding alignment of molecular responses and contraction mode. Furthermore, despite a lack of coherence in differentially expressed genes between studies (at least as a factor of age [22]) ageing could be associated with altered transcriptional responses to exercise that could contribute to poorer adaptation of ageing muscle to RET.
Given maladaptation to exercise (RE/RET) in ageing, and the distinct metabolic and molecular responses to contraction mode, we examined the acute transcriptional response to isolated CON and isolated ECC contractions, in young and older adults. We hypothesised that there would be age-related differences in the transcriptional response to exercise that may help explain age-related differences in the adaptive responses to exercise.
Results
Sequence quality and removal of outliers
FastQC was used to judge the quality of the sequence data. As samples contained no over-represented sequences or adapter sequences and the illumina base quality scores were always >30, no filtering or trimming was performed. A multi-dimensional scaling (MDS) plot to characterise the variation between samples identified the presence of two outliers in the samples (Figure S1A). The samples, a young baseline sample and an older 5 h post-ECC exercise sample, were removed from all subsequent analysis. The MDS plot of the remaining data showed a more clustered set of samples with no obvious outliers (Figure S1B).
Age-independent muscle transcriptomic responses to CON and ECC exercise
Comparing baseline to 5 h post-CON and -ECC exercise, the muscle transcriptome displayed a pattern of significant differential gene expression in both age groups (Table 1). We next directly compared these differentially expressed genes between post-CON and post-ECC exercise, within age groups (Figure 1). In young adults the up- and down-regulated transcriptional profiles of CON and ECC contractions are highly similar (209 and 43 commonly up- and down-regulated genes, respectively). However small contraction mode-specific differences exist, most notably with 12 up-regulated genes unique to ECC. Similarly, in older adults there is large overlap between CON and ECC differentially expressed genes (309 and 113 commonly up- and down-regulated genes, respectively), with enrichment in multiple KEGG pathways including the NF-κB, focal adhesion and energy-sensing pathways. Despite the higher number of contraction-unique up-regulated genes in older muscle (Figure 1), these gene sets show no enrichment for gene ontology (GO) terms suggesting that the genes do not coherently represent any specific functions or that these sets comprise poorly annotated genes. However, KEGG analysis showed ECC-specific upregulation of a longevity pathway, and CON-specific upregulation of the AGE-RAGE signalling pathway (Figure 2). Full lists of differentially expressed genes from baseline to post-exercise, and for the overlap between contraction types can be found in Table S1. Complete lists of enriched GO terms and KEGG pathways for all contraction mode-associated differentially expressed genes can be found in Table S2 and Table S3, respectively.
Table 1. Total numbers of significantly differentially expressed genes 5 h after concentric and eccentric exercise, in skeletal muscle of young and older individuals.
| Concentric exercise | Eccentric exercise | ||||
| Up-regulated | Down-regulated | Up-regulated | Down-regulated | ||
| Young | 227 | 63 | 590 | 204 | |
| Older | 665 | 328 | 724 | 198 | |
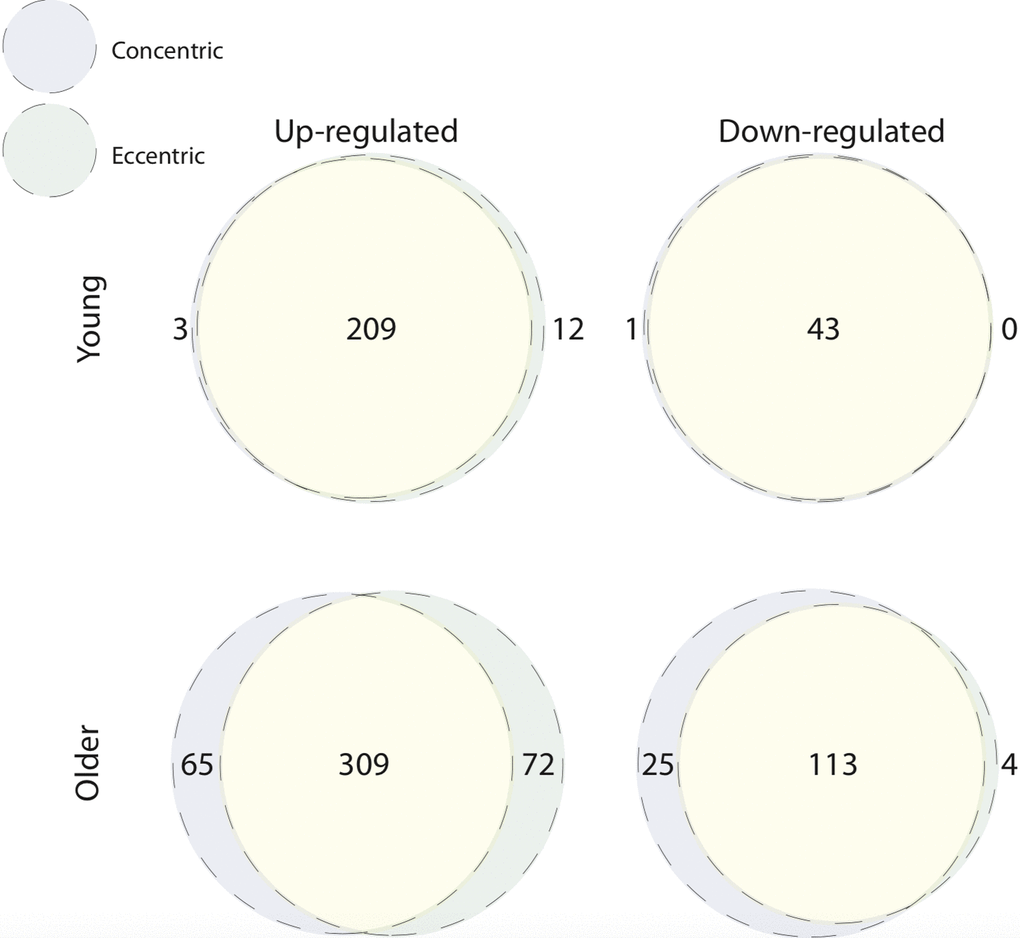
Figure 1. Concentric and eccentric exercise induce a common transcriptomic response in young and older muscle. Overlap of contraction mode-associated differentially expressed genes within age groups. Near complete overlap exists between up-regulated (top left Venn) and down-regulated (top right Venn) genes, 5 h after concentric and eccentric exercise in young muscle. Up-regulated (bottom left Venn) and down-regulated (bottom right Venn) genes in older muscle after concentric and eccentric exercise also display predominant overlap.
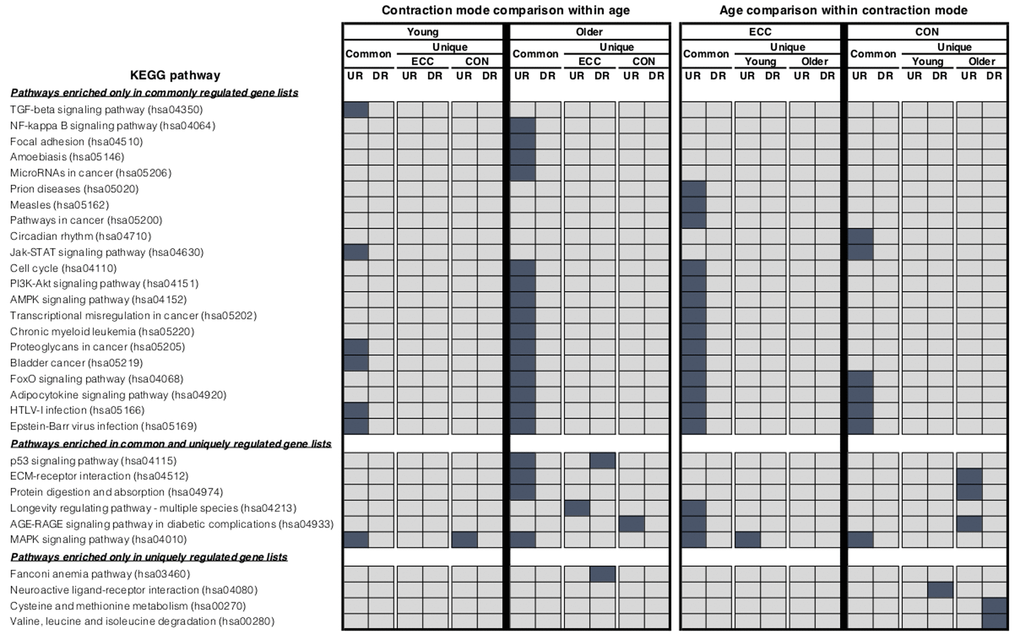
Figure 2. KEGG pathway analysis. Summary of significantly enriched KEGG pathways for the lists of differentially expressed genes that were identified as being either commonly or uniquely regulated when comparing between contraction mode within age group and between age groups within each contraction mode using the rank-rank hypergeometric overlap. Dark blue shading highlights which KEGG terms were found to be significantly enriched (adjusted P-value <0.05) in each condition.
The effect of age on the muscle transcriptomic response to CON and ECC exercise
To determine whether ageing muscle displays a disparate post-exercise transcriptional profile compared to the young, GO term enrichment was first assessed for differentially expressed genes between baseline and post-CON and baseline and post-ECC within age groups. In the young, we identified between 6-115 GO terms in the ‘Biological Processes’ ontology in up- or down-regulated genes after CON and ECC exercise. Whilst more specific terms such as ‘regulation of TOR signalling cascade’ were expectedly identified, more predominant (e.g. common to both CON and ECC responses) were high-level GO terms including ‘signalling,’ ‘multicellular organismal process’ and ‘developmental process’, which are too broad to be informative. Thus, no consistent functional pattern emerges for the post-exercise transcriptional response in young muscle. The older muscle response to ECC similarly identified numerous high-level GO terms enriched in up-regulated (138) and down-regulated (39) genes. Full lists of these GO terms can be viewed in Table S2. However, a clear post-CON specific pattern of GO term enrichment arose in older muscle: terms associated with blood vessel development and cell adhesion were enriched in up-regulated genes, and several terms relating to mitochondrial respiration were enriched in the set of down-regulated genes (Figure 3).
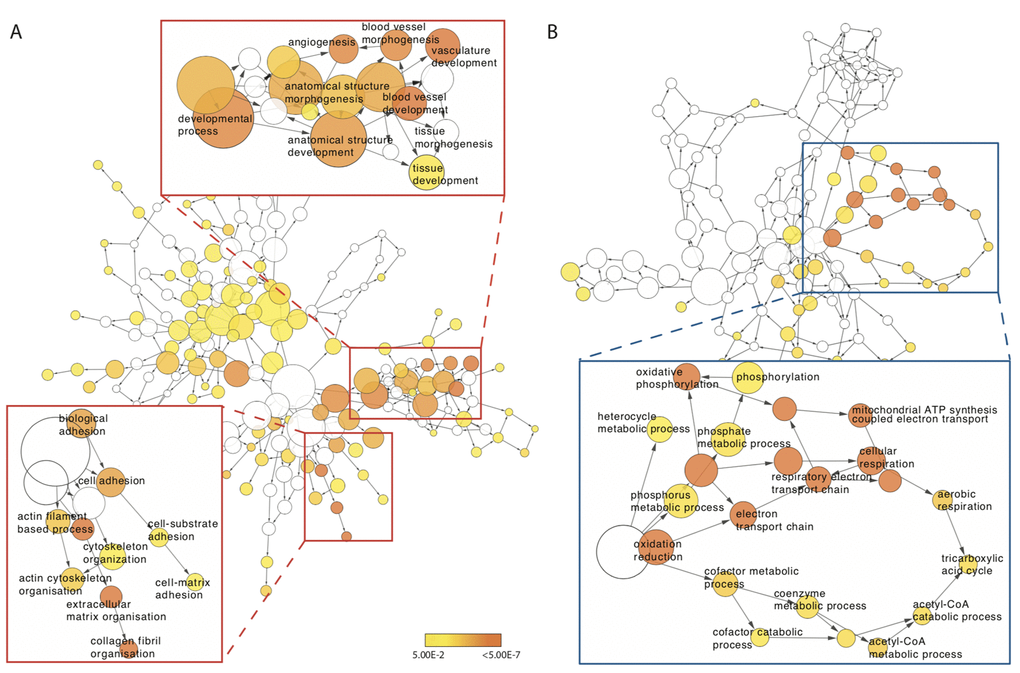
Figure 3. Concentric exercise-mediated transcriptomic profile of older muscle. (A) GO terms enriched for genes in older muscle that display increased expression 5 h post-concentric exercise. Red inserts highlight increased expression of GO terms associated with cytoskeletal, cell adhesion and extra-cellular matrix (bottom right) and blood vessel development (top left). (B) GO terms enriched for genes in older muscle that display reduced expression 5 h post-concentric exercise. Blue insert highlights reduced expression of GO term network associated with mitochondrial metabolism. Each node in the network represents a GO term with the size of each node corresponding to the number of genes associated with that term. Significantly enriched terms are coloured yellow with more significant terms a deeper shade.
We next overlaid post-CON and post–ECC differentially expressed genes between young and older age groups to directly compare age-specific transcriptomic responses (Figure 4). After CON exercise, very few genes were uniquely up-regulated (13 genes) and down-regulated (2 genes) in young muscle, with no clear GO pattern. In contrast (and in line with older baseline to 5 h post-CON expression changes), post-CON older muscle displayed: i) unique up-regulation of 147 genes enriched for terms associated with cell adhesion, extracellular organisation and blood vessel development and, ii) unique down-regulation of 28 genes enriched for terms relating to the metabolism and/ or catabolism of amino acids, lipids, carboxylic acid and DNA (Table 2). KEGG analysis further confirms GO findings that in older muscle, CON exercise specifically down-regulates pathways of amino acid metabolism, and up-regulates extra-cellular matrix-receptor related pathways (Figure 2). Despite comparably high numbers of uniquely up-regulated (151) and down-regulated (19) genes after ECC in the older group, and a differential ECC expression profile in the young (81 up-regulated and 23 down-regulated genes), there was no GO term enrichment for any age-dependent ECC gene sets. Complete lists of enriched GO terms for post-exercise differentially expressed genes between age groups can be viewed in Table S2. Due to the tendency for poor GO term enrichment, we further assessed the top 10 ranked DE genes per condition based on logarithmic fold change in expression to provide a functional overview of the most significantly altered genes (Figure 5), and assessed gene enrichment in KEGG pathways (Figure 2). However, KEGG analysis also failed to find ECC-specific gene enrichment in any biological pathway.
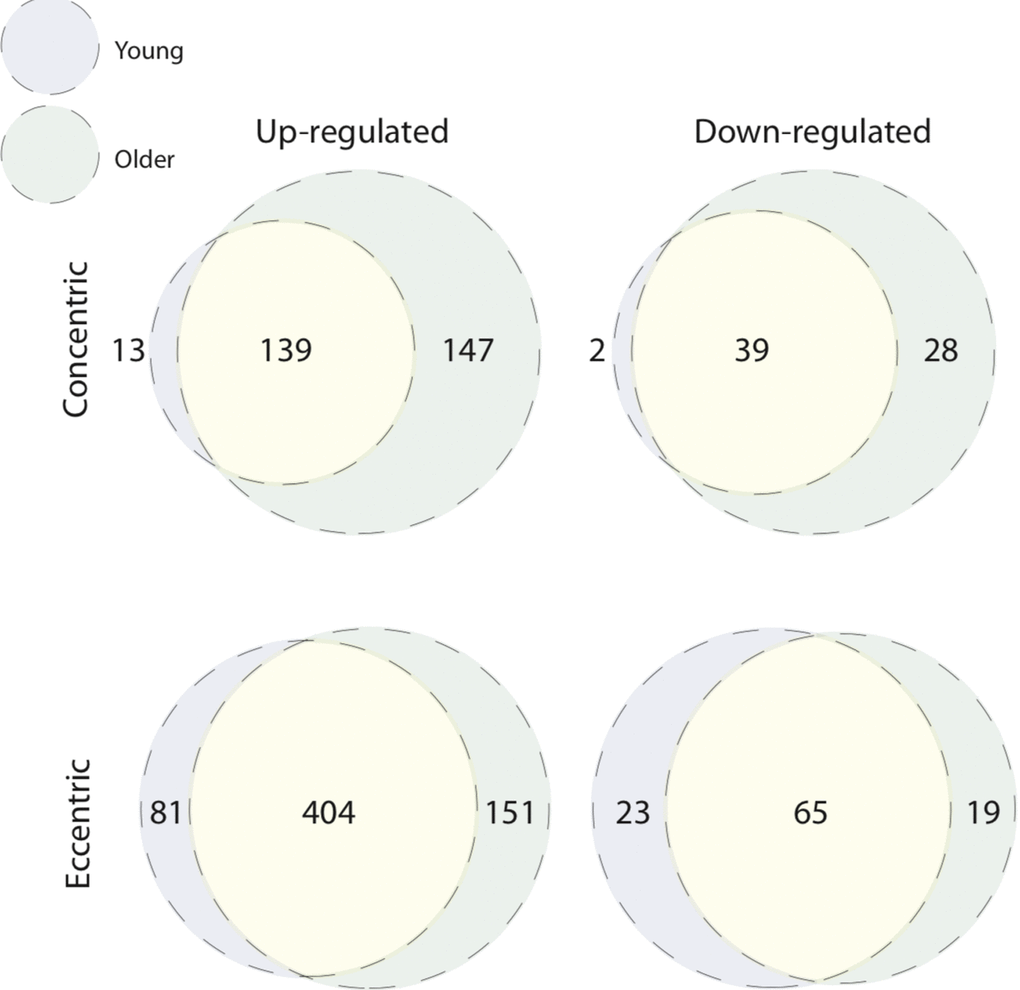
Figure 4. Unique age-related transcriptomic response to concentric and eccentric exercise. Overlap of age-associated differentially expressed genes within contraction modes. Both young and older muscle display unique signatures of differential gene expression 5 h post-concentric (up-regulated, top left Venn; down-regulated, top right Venn) and post-eccentric (up-regulated, bottom left Venn; down-regulated, bottom right Venn) exercise.
Table 2. GO terms enriched for significantly differentially expressed genes in older muscle, 5 h post-concentric exercise. Dashed lines separate broad functional GO term classes.
| GO-ID | GO description | Observed DE genes | Total genes in GO-ID | P-raw | P-corrected |
| Up-regulated post-concentric exercise in older muscle only: | |||||
| 7155 | Cell adhesion | 14 | 355 | 2.12E-04 | 3.24E-02 |
| 22610 | Biological adhesion | 14 | 355 | 2.12E-04 | 3.24E-02 |
| 43062 | Extracellular structure organization | 9 | 96 | 4.18E-06 | 1.71E-03 |
| 30198 | Extracellular matrix organization | 8 | 72 | 4.08E-06 | 1.71E-03 |
| 30199 | Collagen fibril organization | 6 | 22 | 2.81E-07 | 3.44E-04 |
| 1568 | Blood vessel development | 12 | 210 | 1.83E-05 | 4.47E-03 |
| 1944 | Vasculature development | 12 | 215 | 2.31E-05 | 4.71E-03 |
| 43588 | Skin development | 5 | 21 | 6.14E-06 | 1.88E-03 |
| Down-regulated post-concentric exercise in older muscle only: | |||||
| 9081 | Branched chain family amino acid metabolic process | 2 | 16 | 5.24E-04 | 3.10E-02 |
| 9082 | Branched chain family amino acid biosynthetic process | 1 | 2 | 4.33E-03 | 4.60E-02 |
| 9098 | Leucine biosynthetic process | 1 | 2 | 4.33E-03 | 4.60E-02 |
| 19482 | Beta-alanine metabolic process | 1 | 1 | 2.17E-03 | 4.60E-02 |
| 9063 | Cellular amino acid catabolic process | 2 | 42 | 3.64E-03 | 4.60E-02 |
| 9083 | Branched chain family amino acid catabolic process | 2 | 12 | 2.90E-04 | 2.86E-02 |
| 6550 | Isoleucine catabolic process | 1 | 2 | 4.33E-03 | 4.60E-02 |
| 19484 | Beta-alanine catabolic process | 1 | 1 | 2.17E-03 | 4.60E-02 |
| 6574 | Valine catabolic process | 1 | 1 | 2.17E-03 | 4.60E-02 |
| 34440 | Lipid oxidation | 2 | 34 | 2.39E.03 | 4.60E-02 |
| 19395 | Fatty acid oxidation | 2 | 34 | 2.39E-03 | 4.60E-02 |
| 6635 | Fatty acid beta-oxidation | 2 | 29 | 1.74E-03 | 4.60E-02 |
| 9062 | Fatty acid catabolic process | 2 | 37 | 2.83E-03 | 4.60E-02 |
| 46395 | Carboxylic acid catabolic process | 4 | 80 | 2.21E-05 | 3.27E-03 |
| 42732 | D-xylose metabolic process | 1 | 1 | 2.17E-03 | 4.60E-02 |
| 5997 | Xylulose metabolic process | 1 | 1 | 2.17E-03 | 4.60E-02 |
| 45072 | Regulation of interferon-gamma biosynthetic process | 1 | 2 | 4.33E-03 | 4.60E-02 |
| 45078 | Positive regulation of interferon-gamma biosynthetic process | 1 | 2 | 4.33E-03 | 4.60E-02 |
| 46113 | Nucleobase catabolic process | 1 | 2 | 4.33E-03 | 4.60E-02 |
| 6208 | Pyrimidine base catabolic process | 1 | 2 | 4.33E-03 | 4.60E-02 |
| 19859 | Thymine metabolic process | 1 | 2 | 4.33E-03 | 4.60E-02 |
| 6210 | Thymine catabolic process | 1 | 2 | 4.33E-03 | 4.60E-02 |
| 9439 | Cyanate metabolic process | 1 | 2 | 4.33E-03 | 4.60E-02 |
| 9440 | Cyanate catabolic process | 1 | 2 | 4.33E-03 | 4.60E-02 |
| 6082 | Organic acid metabolic process | 5 | 371 | 9.19E-04 | 4.53E-02 |
| 16054 | Organic acid catabolic process | 4 | 80 | 2.21E-05 | 3.27E-03 |
| 44282 | Small molecule catabolic process | 4 | 179 | 5.06E-04 | 3.10E-02 |
| 44270 | Cellular nitrogen compound catabolic process | 2 | 46 | 4.35E-03 | 4.60E-02 |
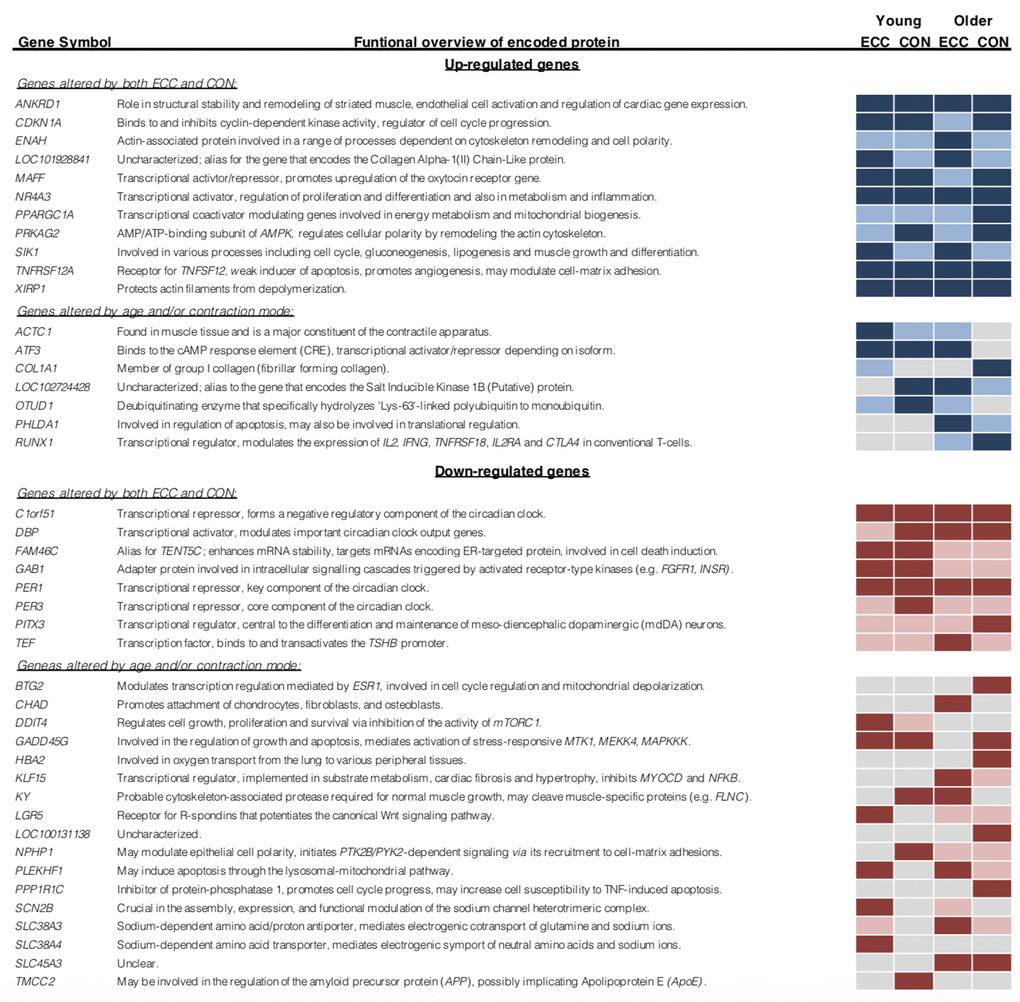
Figure 5. Top 10 ranked differentially expressed genes, classified by contraction mode and age group. Light blue/ red shading denotes gene differential expression for that condition (P<0.05). Dark blue/ red shading indicates gene falls within the top 10 log-fold change ranked differentially expressed genes for that condition.
Discussion
Loading muscle via CON or ECC RET induce divergent architectural and functional adaptations. The role of contraction mode on acute responses of ageing muscle to RE is poorly understood. Here we report that minor transcriptional differences characterise isolated ECC (versus isolated CON) in young muscle and that this pattern persists in older muscle. However, larger differentially expressed gene sets unique to age were observed for both ECC and CON. Whilst ECC-unique genes in older adults failed to cluster into any functional categories, the ageing CON profile was enriched for up-regulated cell adhesion, extracellular organisation and blood vessel development pathways, and down-regulation of genes associated with mitochondrial respiration, amino acid and lipid metabolism, and DNA regulation.
Small differences characterise transcriptional responses to CON versus ECC
In spite of well-established acute and chronic adaptive differences, in young sedentary, healthy muscle we report overlap in the majority of genes acutely responsive to both CON and ECC. The notable exception of 12 up-regulated genes unique to ECC suggests limited contraction mode-specific transcriptional sensitivity, and these failed to cluster to ontology groups. This corroborates previous reports that of 3,300 transcripts examined, 11 genes were uniquely differentially regulated 6 h post-ECC, which did not associate with any functional classifications [19]. Similarly, Chen and colleagues [18] found 28 up-regulated and 0 down-regulated genes (of between 625-1,125 post-exercise differentially expressed genes) 4-8 h after ECC+CON versus CON alone. Thus, irrespective of contraction mode, unaccustomed exercise per se associates with a somewhat “generic” acute global genomic response. It is, however, plausible that transcriptional differences emerge over repeated bouts of RE, as muscle develops phenotypic specificity to ECC and CON. Interestingly, although the pattern of large transcriptional overlap remained, older muscle displayed a higher number of ECC and CON specific differentially expressed genes. Nonetheless, these failed to cluster to any gene GO terms, thus the functional meaning of this slightly more pronounced contraction mode-related difference is unclear. Whilst individual consideration of all CON- and ECC-unique genes is out of scope herein, future detailed analysis of this gene set might provide new mechanistic insight into poor ageing muscle adaptation to exercise.
We further compared our list of contraction mode specific DE genes with those identified in previous microarray studies [18,19,23] and found little overlap (with the exception of USP2 and NR4A1 [23] for young muscle ECC response, and LPP [18] and TM4SF1 [23] for older muscle ECC response). Despite lack of direct alignment in DE genes across human studies in young muscle - in all studies, increased expression of heat shock proteins (HSP) is consistently identified as a post-ECC signature (HSPB1, HSPB8, CRYAB [19], HSPA1B, DNAJB4 [18], DNAJB2 [23] and DNAJB1 presently). Whilst we identified other HSP’s as commonly up-regulated by both ECC and CON (Table S1), ECC appears to associate with an acute HSP response superseding that observed after CON. HSP’s are responsive to environmental stress and serve to stabilise proteins during cellular damage by facilitating protein folding during periods of increased protein synthesis, and saving ‘damaged’ proteins from further unfolding and trafficking to proteolytic machinery for degradation [24]. As such, HSP activation might occur as a post-translational, satellite cell-independent regenerative response to the cellular disruption caused by ECC contractions and the additional (i.e. beyond that required by CON) need for new proteins to replace and repair areas of localised muscle damage.
Age modifies the post-CON and -ECC muscle transcriptome
ECC exercise altered the expression of numerous genes (total 170) uniquely in older muscle versus the young. It is however curious that none of the ECC responsive older muscle-specific genes clustered to any gene ontology terms. For example, no age-specific cell adhesion GO pattern or KEGG pathway enrichment was observed after ECC alone (which, conversely, was found post-CON), where the greater mechanical strain might be anticipated to stimulate a significant mechano-transduction response. This lack of ontological enrichment implies a certain randomness to the ageing muscle transcriptional response to ECC. This might support the ‘disposable soma’ theory of ageing [25] which proposes cellular and molecular ageing as an inherently stochastic process. Such influence of chance events and/ or random responses to external cues could explain the large differences observed in the rate and onset of ageing even among isogenic populations of mice and nematodes [26,27]. Indeed, stochastic ageing might be particularly relevant to muscle, which displays early onset and profound defects across the lifecourse, e.g. versus neuronal decline [26]. Mechanical loads also direct the stochasticity of biochemical reactions [28]. The greater (versus CON) muscular mechanical strain imposed by ECC may thus promote a stochastic transcriptional response, as observed herein. The anticipated net result of such random post-exercise molecular events would be loss of adaptive specificity, such as appropriate sarcomere remodelling and/ or cumulative muscle deterioration. Since these are known sarcopenic phenotypes, stochasticity might underpin at least part of poor muscle ageing.
Virtually all post-CON responsive genes in the young corresponded in older muscle, inferring age per se, does not reduce the transcriptional capacity to appropriately respond to CON. Nonetheless, older muscle displayed additional subsets of up- and down-regulated genes versus the young group. For example, the 147 genes specifically up-regulated in older muscle after CON were enriched for genes associated with blood vessel development and cell-cell adhesion GO terms, and altered cell adhesion pathways was further corroborated by KEGG pathway analysis. Interestingly, this represents an ontological opposite of the baseline ageing expression profile (data not shown) suggestive of a compensatory attempt of older muscle to correct age-related transcriptional abnormalities. Since this presumed corrective mechanism is ineffective at fully mitigating poor ageing muscle adaptation to exercise (i.e. the well-established ‘anabolic blunting’ phenomenon [6–8]), future investigation of the multifaceted role of impaired cell adhesion and associated mechanically-mediated intracellular signalling (e.g. via integrin-adhesome complexes) might provide new insight into the mechanisms of blunted ageing muscle exercise responses. Acute CON also associated with an age-specific down-regulation of certain metabolic pathways. Genes supressed post-CON in the older group clustered to a distinct subset of mitochondrial function, fatty acid and amino acid metabolism ontologies (the latter also confirmed by KEGG analysis). Diminished mitochondrial volume [29] and oxidative capacity [30–32] are frequently reported features of ageing muscle. Nonetheless, such studies are typically poorly controlled for confounders to confer age effects, i.e. physical activity levels [33].
The present study’s recruitment of sedentary young and older adults provides evidence that age per se can influence the muscle mitochondrial response to unaccustomed muscle contraction. Importantly, whilst aerobic and RE-training increase ageing muscle mitochondrial content and function [34], these adaptations remain below that of healthy young counterparts [35,36]. Our data provide preliminary evidence for a transcriptional basis of attenuated adaptation and implicates mitochondrial insensitivity to exercise (with CON at least) in the aetiology of ageing muscle deterioration. Finally, CON also down-regulated several amino acid metabolic and catabolic ontologies/ KEGG pathways, implying a post-CON disturbance in ageing muscles’ ability to process the substrates of muscle protein turnover. It is therefore possible that abnormal amino acid processing contributes to anabolic blunting of ageing muscle during exercise training [6–8].
Methods
Subjects
Eight young (age, 21±1 y; body mass index, 23±2 kg/m2; 80% ECC 1RM, 211±14 kg; 80% CON 1RM, 122±11 kg) and eight older (age, 70±1y; body mass index, 26±1 kg/m2; 80% ECC 1RM, 155±1 kg; 80% CON 1RM, 79±6 kg) healthy male volunteers were recruited to this study. Because muscular adaptation occurs following a single bout of exercise to diminish the post-exercise regenerative response (i.e. the ‘repeated bout effect’ [37]), all recruited volunteers were exercise naïve, defined here as not participating in any form of structured exercise regime, and having no history of exercise training within the previous 12 months. Ethical approval was obtained from the University of Nottingham Medical School Ethics Committee, and all studies carried out in accordance with the Declaration of Helsinki. All volunteers were health-screened prior to participation.
Study design
Visit 1: Volunteers were asked to refrain from exercise for 72 h and before testing performed an overnight fast with water consumed ad libitum. Volunteers arrived at ~9:00 h for a baseline muscle biopsy from the m. vastus lateralis of a randomised leg. Biopsies were taken under sterile conditions using local anaesthetic (1% lidocaine), blotted on gauze to remove excess blood, snap frozen in liquid nitrogen and stored at -80°C. Visit 2: After 96 h having conducted no exercise, volunteers performed an overnight fast and arrived at the laboratory at ~8.30 h after consuming a 250 ml liquid high energy nutritionally complete drink (Fortisip, Nutricia) at 07:00 h. Volunteers remained fasted thereafter until the study was complete at ~17.30 h. The isolated CON and isolated ECC knee extension exercise protocols were then performed; legs were randomised to unilateral CON-only and contralateral ECC-only exercise and the order in which CON and ECC were performed was also randomised. Exercise was performed on a modified leg press designed to permit either CON or ECC contractions in isolation within a single leg [9] with a starting knee angle of 90° for CON contractions and 180° for ECC contractions. For each isolated contraction mode, after an isolated CON/ECC warm-up consisting of 2 sets of 6 repetitions set at 40 kg volunteers, isolated CON/ECC one-repetition maximum (1-RM) was determined at a starting load of 50% estimated 1-RM, with subsequent attempts separated by 3 min and the resistance increased until a controlled contraction (~3 seconds) through the full range of motion could no longer be performed. Immediately after CON/ECC 1-RM had been established, the exercise protocol was performed comprising 7 sets of 10 repetitions (~3 second contractions) at 80% CON-specific or ECC-specific 1-RM, with 2-minute rest between sets. After ~1 h recovery the above exercise protocol was performed for the second contraction mode on the opposing leg. Finally, 5 h following the cessation of each exercise protocol a muscle biopsy was collected from the m. vastus lateralis of each leg.
RNA sequencing
All RNA extraction, library preparations and next generation sequencing was performed by the Beijing Genomics Institute. Total RNA was isolated by using TRIzol reagent. Of the 8 volunteers, RNA integrity was low for 1 young and 1 older volunteer at all time points (i.e. baseline, 5 h post-ECC and 5 h post-CON) and were excluded from further analysis. Samples for 1 older volunteer (5 h post-ECC) and 2 young volunteers (5 h post-ECC and 5 h post-CON) also had low RNA integrity and were also excluded from analysis. Library preparations on the remaining 39 samples were performed using TruSeq RNA library preparation kits. Next generation sequencing was performed using the HiSeq 3000/HiSeq 4000 Illumina sequencing by synthesis chemistry systems, using advanced patterned flow cell technology for maximised level of sample throughput. The data have been deposited with links to BioProject accession number PRJNA509121 in the NCBI BioProject database (https://www.ncbi.nlm.nih.gov/bioproject/).
Alignment of sequencing data
The quality of sequencing reads was evaluated using FastQC (Babraham Bioinformatics). As the quality of reads was judged to be high no additional filtering or trimming was performed. Reads were aligned to the human reference genome (hg38 NCBI - iGenomes) using Bowtie2 [38]. A 'local' alignment was performed on single end reads with the 'very-sensitive' pre-set parameters. Further processing of the alignment files was performed using samtools [39].
Identifying differential expression
Reads mapping to known exons were counted using featureCounts [40] in an unstranded fashion and using the human genome annotation as a reference (hg38 NCBI – iGenomes). To characterise the variation between sequencing samples, edgeR [41] was used to produce a MDS plot. On the basis of the MDS plot two samples (one young baseline and one older ECC) were removed from the analysis. Differential expression was inferred between groups (i.e. young baseline versus young CON) using edgeR. In the edgeR analysis genes were only included in the analysis if the counts per million (CPM) for that gene was greater than 2 in more than 10 samples. Counts were normalised by library size and biological and technical variation (dispersion) was estimated. Significance of differential expression was estimated using the Fisher's exact test and P-values were corrected for a false discovery rate using the method of Benjamini and Hochberg [42] with a significance cut-off of P-corr < 0.05.
Comparing differentially expressed genes across age and exercise modes
To identify overlapping differentially expressed genes we used the rank-rank hypergeometric overlap (RRHO) where genes were ranked on log fold change and P<0.05 defined a significant overlap [43]. The overlap between the exercise modes was identified by comparing each exercise mode against baseline within age groups. The overlap between young and older post-exercise responses was identified by comparing CON or ECC exercise modes across age groups. Sample specific differentially expressed genes were defined as those genes that showed statistically different ranks in RRHO and significant differential expression in that sample. Overlapping differentially expressed genes were defined as those genes that showed no significant difference in rank and were significantly differentially expressed in both samples.
Pathway analysis
Significant differentially expressed genes were tested for functional enrichment using the GO [44] and BiNGO [45]. In this analysis the ‘sample’ was the set of differentially expressed genes and the ‘background’ was all genes that passed the filtering criteria in the edgeR analysis (see above). In particular, we tested for enrichment of GO Biological Process terms using the hypergeometric test and used all annotations for human genes and GO terms. Lists of differentially expressed genes were also subject to pathway enrichment analysis using the Enrichr web server [46,47], with specific focus placed on enrichment for pathway terms contained within the Kyoto Encyclopaedia of Genes and Genomes (KEGG) database [48]. In each case, terms with a Benjamini-Hochberg [42] corrected P < 0.05 were accepted as being enriched. Significantly enriched GO Biological Process terms were subsequently visualised as a network using BiNGO and Cytoscape [49].
Ranking contraction-regulated genes
To rank the top 10 differentially expressed genes for each contraction mode and age group, up-regulated and down-regulated genes for each condition (versus baseline) were ranked separately based on their logarithmic fold-change in expression (log2 fold change [LFC]), and prioritised candidate targets then selected in each instance as the top 10 genes with the greatest LFC. Insight into the functional role(s) of each identified candidate target was subsequently assign using the UniProt database (uniprot.org).
Supplementary Materials
Acknowledgements
The authors would like to acknowledge Margaret Baker and Amanda Gates for their clinical and technical support. The authors would also like to thank all of the volunteers for giving up their time to complete the study.
Conflicts of Interest
The authors declare no conflicts of interest.
Funding
CSD was funded by a doctoral training studentship from Bournemouth University. This work was generously supported by the Wellcome Trust Institutional Strategic Support Award (WT105618MA). RMA is generously supported by the Wellcome Trust Institutional Strategic Support Award (WT105618MA) and an EPSRC/BBSRC Innovation Fellowship (EP/S001352/1). We acknowledge the Medical Research Council [grant number MR/P021220/1] [grant number MR/K00414X/1] and Arthritis Research UK [grant number 19891] as part of the MRC-ARUK Centre for Musculoskeletal Ageing Research awarded to the Universities of Nottingham and Birmingham, and the National Institute for Health Research, Nottingham Biomedical Research Centre. This work was supported by the Biotechnology and Biological Sciences Research Council [grant number BB/N015894/1]. This work was supported by a grant from the Swedish Research Council for Sport Science (dnr 2016/125 and dnr 2017/143). C.R.G.W is supported by the Biotechnology and Biological Sciences Research Council-funded South West Biosciences Doctoral Training Partnership [BB/J014400/1; BB/M009122/1].
References
- 1. Reid KF, Fielding RA. Skeletal muscle power: a critical determinant of physical functioning in older adults. Exerc Sport Sci Rev. 2012; 40:4–12. https://doi.org/10.1097/JES.0b013e31823b5f13 [PubMed]
- 2. Zurlo F, Larson K, Bogardus C, Ravussin E. Skeletal muscle metabolism is a major determinant of resting energy expenditure. J Clin Invest. 1990; 86:1423–27. https://doi.org/10.1172/JCI114857 [PubMed]
- 3. Marquis K, Debigaré R, Lacasse Y, LeBlanc P, Jobin J, Carrier G, Maltais F. Midthigh muscle cross-sectional area is a better predictor of mortality than body mass index in patients with chronic obstructive pulmonary disease. Am J Respir Crit Care Med. 2002; 166:809–13. https://doi.org/10.1164/rccm.2107031 [PubMed]
- 4. von Haehling S, Morley JE, Anker SD. An overview of sarcopenia: facts and numbers on prevalence and clinical impact. J Cachexia Sarcopenia Muscle. 2010; 1:129–33. https://doi.org/10.1007/s13539-010-0014-2 [PubMed]
- 5. Fiatarone MA, O’Neill EF, Ryan ND, Clements KM, Solares GR, Nelson ME, Roberts SB, Kehayias JJ, Lipsitz LA, Evans WJ. Exercise training and nutritional supplementation for physical frailty in very elderly people. N Engl J Med. 1994; 330:1769–75. https://doi.org/10.1056/NEJM199406233302501 [PubMed]
- 6. Fry CS, Drummond MJ, Glynn EL, Dickinson JM, Gundermann DM, Timmerman KL, Walker DK, Dhanani S, Volpi E, Rasmussen BB. Aging impairs contraction-induced human skeletal muscle mTORC1 signaling and protein synthesis. Skelet Muscle. 2011; 1:11. https://doi.org/10.1186/2044-5040-1-11 [PubMed]
- 7. Kumar V, Selby A, Rankin D, Patel R, Atherton P, Hildebrandt W, Williams J, Smith K, Seynnes O, Hiscock N, Rennie MJ. Age-related differences in the dose-response relationship of muscle protein synthesis to resistance exercise in young and old men. J Physiol. 2009; 587:211–17. https://doi.org/10.1113/jphysiol.2008.164483 [PubMed]
- 8. Phillips BE, Williams JP, Greenhaff PL, Smith K, Atherton PJ. Physiological adaptations to resistance exercise as a function of age. JCI Insight. 2017; 2:95581. https://doi.org/10.1172/jci.insight.95581 [PubMed]
- 9. Franchi MV, Atherton PJ, Reeves ND, Flück M, Williams J, Mitchell WK, Selby A, Beltran Valls RM, Narici MV. Architectural, functional and molecular responses to concentric and eccentric loading in human skeletal muscle. Acta Physiol (Oxf). 2014; 210:642–54. https://doi.org/10.1111/apha.12225 [PubMed]
- 10. Reeves ND, Maganaris CN, Longo S, Narici MV. Differential adaptations to eccentric versus conventional resistance training in older humans. Exp Physiol. 2009; 94:825–33. https://doi.org/10.1113/expphysiol.2009.046599 [PubMed]
- 11. Schoenfeld BJ, Ogborn DI, Vigotsky AD, Franchi MV, Krieger JW. Hypertrophic effects of concentric vs. eccentric muscle actions: a systematic review and meta-analysis. J Strength Cond Res. 2017; 31:2599–608. https://doi.org/10.1519/JSC.0000000000001983 [PubMed]
- 12. Hoppeler H. Moderate load eccentric exercise; a distinct novel training modality. Front Physiol. 2016; 7:483. https://doi.org/10.3389/fphys.2016.00483 [PubMed]
- 13. Hortobágyi T, DeVita P. Favorable neuromuscular and cardiovascular responses to 7 days of exercise with an eccentric overload in elderly women. J Gerontol A Biol Sci Med Sci. 2000; 55:B401–10. https://doi.org/10.1093/gerona/55.8.B401 [PubMed]
- 14. Mueller M, Breil FA, Vogt M, Steiner R, Lippuner K, Popp A, Klossner S, Hoppeler H, Däpp C. Different response to eccentric and concentric training in older men and women. Eur J Appl Physiol. 2009; 107:145–53. https://doi.org/10.1007/s00421-009-1108-4 [PubMed]
- 15. Dias CP, Toscan R, de Camargo M, Pereira EP, Griebler N, Baroni BM, Tiggemann CL. Effects of eccentric-focused and conventional resistance training on strength and functional capacity of older adults. Age (Dordr). 2015; 37:99. https://doi.org/10.1007/s11357-015-9838-1 [PubMed]
- 16. LaStayo P, Marcus R, Dibble L, Wong B, Pepper G. Eccentric versus traditional resistance exercise for older adult fallers in the community: a randomized trial within a multi-component fall reduction program. BMC Geriatr. 2017; 17:149. https://doi.org/10.1186/s12877-017-0539-8 [PubMed]
- 17. Symons TB, Vandervoort AA, Rice CL, Overend TJ, Marsh GD. Effects of maximal isometric and isokinetic resistance training on strength and functional mobility in older adults. J Gerontol A Biol Sci Med Sci. 2005; 60:6. [PubMed]
- 18. Chen YW, Hubal MJ, Hoffman EP, Thompson PD, Clarkson PM. Molecular responses of human muscle to eccentric exercise. J Appl Physiol (1985). 2003; 95:2485–94. https://doi.org/10.1152/japplphysiol.01161.2002 [PubMed]
- 19. Kostek MC, Chen YW, Cuthbertson DJ, Shi R, Fedele MJ, Esser KA, Rennie MJ. Gene expression responses over 24 h to lengthening and shortening contractions in human muscle: major changes in CSRP3, MUSTN1, SIX1, and FBXO32. Physiol Genomics. 2007; 31:42–52. https://doi.org/10.1152/physiolgenomics.00151.2006 [PubMed]
- 20. Raue U, Trappe TA, Estrem ST, Qian HR, Helvering LM, Smith RC, Trappe S. Transcriptome signature of resistance exercise adaptations: mixed muscle and fiber type specific profiles in young and old adults. J Appl Physiol (1985). 2012; 112:1625–36. https://doi.org/10.1152/japplphysiol.00435.2011 [PubMed]
- 21. Thalacker-Mercer AE, Dell’Italia LJ, Cui X, Cross JM, Bamman MM. Differential genomic responses in old vs. young humans despite similar levels of modest muscle damage after resistance loading. Physiol Genomics. 2010; 40:141–49. https://doi.org/10.1152/physiolgenomics.00151.2009 [PubMed]
- 22. Phillips BE, Williams JP, Gustafsson T, Bouchard C, Rankinen T, Knudsen S, Smith K, Timmons JA, Atherton PJ. Molecular networks of human muscle adaptation to exercise and age. PLoS Genet. 2013; 9:e1003389. https://doi.org/10.1371/journal.pgen.1003389 [PubMed]
- 23. Mahoney DJ, Safdar A, Parise G, Melov S, Fu M, MacNeil L, Kaczor J, Payne ET, Tarnopolsky MA. Gene expression profiling in human skeletal muscle during recovery from eccentric exercise. Am J Physiol Regul Integr Comp Physiol. 2008; 294:R1901–10. https://doi.org/10.1152/ajpregu.00847.2007 [PubMed]
- 24. Senf SM. Skeletal muscle heat shock protein 70: diverse functions and therapeutic potential for wasting disorders. Front Physiol. 2013; 4:330. https://doi.org/10.3389/fphys.2013.00330 [PubMed]
- 25. Kirkwood TB. Evolution of ageing. Nature. 1977; 270:301–04. https://doi.org/10.1038/270301a0 [PubMed]
- 26. Herndon LA, Schmeissner PJ, Dudaronek JM, Brown PA, Listner KM, Sakano Y, Paupard MC, Hall DH, Driscoll M. Stochastic and genetic factors influence tissue-specific decline in ageing C. elegans. Nature. 2002; 419:808–14. https://doi.org/10.1038/nature01135 [PubMed]
- 27. Tilstra JS, Robinson AR, Wang J, Gregg SQ, Clauson CL, Reay DP, Nasto LA, St Croix CM, Usas A, Vo N, Huard J, Clemens PR, Stolz DB, et al. NF-κB inhibition delays DNA damage-induced senescence and aging in mice. J Clin Invest. 2012; 122:2601–12. https://doi.org/10.1172/JCI45785 [PubMed]
- 28. Xiao B, Cui LQ, Chen TM, Lian B. Stochastic effects in adaptive reconstruction of body damage: implied the creativity of natural selection. J Cell Mol Med. 2015; 19:2521–29. https://doi.org/10.1111/jcmm.12647 [PubMed]
- 29. Conley KE, Jubrias SA, Esselman PC. Oxidative capacity and ageing in human muscle. J Physiol. 2000; 526:203–10. https://doi.org/10.1111/j.1469-7793.2000.t01-1-00203.x [PubMed]
- 30. Tonkonogi M, Fernström M, Walsh B, Ji LL, Rooyackers O, Hammarqvist F, Wernerman J, Sahlin K. Reduced oxidative power but unchanged antioxidative capacity in skeletal muscle from aged humans. Pflugers Arch. 2003; 446:261–69. https://doi.org/10.1007/s00424-003-1044-9 [PubMed]
- 31. Larsen RG, Callahan DM, Foulis SA, Kent-Braun JA. Age-related changes in oxidative capacity differ between locomotory muscles and are associated with physical activity behavior. Appl Physiol Nutr Metab. 2012; 37:88–99. https://doi.org/10.1139/h11-135 [PubMed]
- 32. Short KR, Bigelow ML, Kahl J, Singh R, Coenen-Schimke J, Raghavakaimal S, Nair KS. Decline in skeletal muscle mitochondrial function with aging in humans. Proc Natl Acad Sci USA. 2005; 102:5618–23. https://doi.org/10.1073/pnas.0501559102 [PubMed]
- 33. Payne BA, Chinnery PF. Mitochondrial dysfunction in aging: much progress but many unresolved questions. Biochim Biophys Acta. 2015; 1847:1347–53. https://doi.org/10.1016/j.bbabio.2015.05.022 [PubMed]
- 34. Peterson CM, Johannsen DL, Ravussin E. Skeletal muscle mitochondria and aging: a review. J Aging Res. 2012; 2012:194821. https://doi.org/10.1155/2012/194821 [PubMed]
- 35. Lanza IR, Short DK, Short KR, Raghavakaimal S, Basu R, Joyner MJ, McConnell JP, Nair KS. Endurance exercise as a countermeasure for aging. Diabetes. 2008; 57:2933–42. https://doi.org/10.2337/db08-0349 [PubMed]
- 36. Wang E, Næss MS, Hoff J, Albert TL, Pham Q, Richardson RS, Helgerud J. Exercise-training-induced changes in metabolic capacity with age: the role of central cardiovascular plasticity. Age (Dordr). 2014; 36:665–76. https://doi.org/10.1007/s11357-013-9596-x [PubMed]
- 37. McHugh MP. Recent advances in the understanding of the repeated bout effect: the protective effect against muscle damage from a single bout of eccentric exercise. Scand J Med Sci Sports. 2003; 13:88–97. https://doi.org/10.1034/j.1600-0838.2003.02477.x [PubMed]
- 38. Langmead B, Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nat Methods. 2012; 9:357–59. https://doi.org/10.1038/nmeth.1923 [PubMed]
- 39. Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, Marth G, Abecasis G, Durbin R, and 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics. 2009; 25:2078–79. https://doi.org/10.1093/bioinformatics/btp352 [PubMed]
- 40. Liao Y, Smyth GK, Shi W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics. 2014; 30:923–30. https://doi.org/10.1093/bioinformatics/btt656 [PubMed]
- 41. Robinson MD, McCarthy DJ, Smyth GK. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics. 2010; 26:139–40. https://doi.org/10.1093/bioinformatics/btp616 [PubMed]
- 42. Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc B. 1995; 57:289–300. https://doi.org/10.1111/j.2517-6161.1995.tb02031.x
- 43. Plaisier SB, Taschereau R, Wong JA, Graeber TG. Rank-rank hypergeometric overlap: identification of statistically significant overlap between gene-expression signatures. Nucleic Acids Res. 2010; 38:e169. https://doi.org/10.1093/nar/gkq636 [PubMed]
- 44. Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, Harris MA, Hill DP, Issel-Tarver L, et al, and The Gene Ontology Consortium. Gene ontology: tool for the unification of biology. Nat Genet. 2000; 25:25–29. https://doi.org/10.1038/75556 [PubMed]
- 45. Maere S, Heymans K, Kuiper M. BiNGO: a Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics. 2005; 21:3448–49. https://doi.org/10.1093/bioinformatics/bti551 [PubMed]
- 46. Chen EY, Tan CM, Kou Y, Duan Q, Wang Z, Meirelles GV, Clark NR, Ma’ayan A. Enrichr: interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinformatics. 2013; 14:128. https://doi.org/10.1186/1471-2105-14-128 [PubMed]
- 47. Kuleshov MV, Jones MR, Rouillard AD, Fernandez NF, Duan Q, Wang Z, Koplev S, Jenkins SL, Jagodnik KM, Lachmann A, McDermott MG, Monteiro CD, Gundersen GW, Ma’ayan A. Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016; 44:W90-7. https://doi.org/10.1093/nar/gkw377 [PubMed]
- 48. Kanehisa M, Goto S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Research. 2000; 28:27-30.
- 49. Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003; 13:2498–504. https://doi.org/10.1101/gr.1239303 [PubMed]