



Introduction
Clinically, many diseases not only involve a single organ or system, but also have obvious overlap in their pathophysiological mechanisms, so it is necessary to comprehensively consider the relationship between various organs or systems to take positive and effective prevention and treatment measures [1, 2]. Among them, the relationship between chronic kidney disease and cardiovascular disease is a famous model [3, 4]. It has been found that the incidence of cardiovascular disease is significantly increased in patients with chronic kidney disease, and some renal transplant patients have coronary artery disease, cardiac structural abnormalities, and heart failure [5]. Cardiovascular death accounts for more than 50% of patients with chronic kidney disease. Even if the renal function decreases slightly, it also has an adverse effect on the clinical prognosis of many patients with heart disease [6, 7]. For this reason, it is necessary to actively explore new potentially mechanism for cardiac disease with chronic kidney disease.
Cardiac fibrosis is caused by cardiomyocyte injury caused by various causes. Damaged cardiomyocytes can release exosomes, and the components of exosomes released by damaged cardiomyocytes are quite different from those released by normal cardiomyocytes, which can trigger the repair process and eventually lead to myocardial fibrosis [8–10]. For example, patients with diabetic cardiomyopathy are usually accompanied by cardiac fibrosis. [11] The content of heat shock protein 20 (HSP20) in the exosomes released by cardiomyocytes in patients with diabetic cardiomyopathy was significantly lower than that in the control group, indicating that the decreased expression of HSP20 may be one of the causes of myocardial fibrosis in patients with diabetic cardiomyopathy [12]. Some interventions can promote the release of exosome from normal cardiomyocytes, which has the function of reducing cardiac fibrosis, such as exercise can effectively prevent and treat diabetic cardiomyopathy. Sporting can reduce myocardial fibrosis in patients with diabetic cardiomyopathy by promoting the release of exosomes from cardiomyocytes, which can increase the levels of series of miRNA (miR-19a [13] miR-100-5p [14], miR-21 [15] and miR-155-5p [16]) carried by these exosomes, and then down-regulate the expression of MMP-9 and block the remodeling of downstream extracellular matrix [17].
At present, the studies on the contents of exosomes are mostly focused on macromolecules such as protein, mRNA and miRNA. In contrast, the studies on exosome-related lncRNA are rare, and most studies focus on tumors, but there are few reports on circulatory, respiratory and digestive system diseases [18]. It was reported that lncRNA ZFAS1 as a serca2a inhibitor to cause intracellular ca2+ overload and contractile dysfunction in myocardial infarction mice [19]. Down-regulated of lncRNA ZFAS1 could protect cardiomyocytes against acute myocardial infarction through anti-apoptosis by regulating miR-150/CRP [20]. However, the pathway of lncRNA ZFAS1 has not been deeply studied.
Wnt/β-catenin signaling pathway is widely involved in the development of embryos and organs and controls self-balance and self-renewal in most mature tissues. Wnt signal pathways, namely classical wnt signal pathway and non-classical Wnt signal pathway, are strictly related to cell development and play an essential role in the event of tissue and organ fibrosis. Wnt signaling pathway may play different roles in the occurrence, development, and inhibition of fibrosis in the lung, heart, kidney, liver, and other organs. Here, we would explore the effect of Wnt/β-catenin signaling pathway in cardiac fibrosis of CKD mice.
In our research, we explored the function of lncRNA ZFAS1 transferred by exosomes regulated WNT4/β-catenin signal pathways in cardiac fibrosis of CKD mice.
Results
LncRNA ZFAS1 was increased in the heart tissue of CKD mice
In previous reports, lncRNA ZFAS1 involved in CVD; however, the research on heart disease caused by CKD is not detailed. To explore the role of lncRNA ZFAS1, we constructed the CKD model in C57BL/6 mice, the sham group was indicated as the control group. In exploring new therapeutic targets for treating cardiomyopathy associated with CKD, we performed lncRNA deep sequence analysis from heart samples of CKD mice. The data revealed 196 lncRNAs that were altered in CKD mice compared with sham mice. 119 lncRNAs were up-regulated, and 77 lncRNAs were down-regulation. Among 119 up-regulated lncRNA, lncRNA ZFAS1 was a significant increase in heart tissues of CKD mice (Figure 1A). Volcano map also showed that the expression of lncRNA ZFAS1 in CKD heart tissue was higher than that in the sham group (Figure 1B). Then we isolated the total RNA from heart tissues for assessing the expression of lncRNA ZFAS1.RT-PCR result performed that lncRNA ZFAS1 was abnormally increased in heart tissues of CKD mice (Figure 1C). Based on the above findings, we speculate that lncRNA ZFAS1 involved in the heart injury caused by CKD.
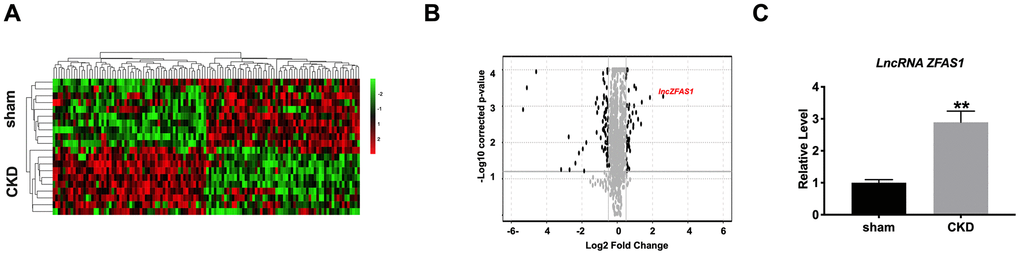
Figure 1. Increased level of lncRNA ZFAS1 in the heart tissue of CKD mice. (A) The volcano plot of lncRNAs. (B) Heat map of microarray data was used to show the significant expression of lncRNAs in heart tissues of sham and CKD mice. (C) The expression of lncRNA ZFAS1 was detected in the heart tissues of sham and CKD mice. *P<0.05.
Isolated and confirmed of exosome
Exosomes can be derived from many cell types; the further study was performed to assess whether lncRNA ZFAS1 regulated heart injury via the exosome pathway. Then we isolated exosome from the supernatant of HCM, and confirmed the characteristics of the exosome. Then we detected the exosome specific expressed protein (CD81, CD63) to confirm exosome (Figure 2A). In contrast, we discovered the particle size of the extract to establish the characteristics of exosome (Figure 2B). Then we transfected lncRNA ZFAS1/NC into HCM cells. RT-PCR was performed to detect the expression level of lncRNA ZFAS1 (Figure 2C). Next, we isolated exosome and observed the morphology of exosome by transmission electron microscopy (TEM, Figure 2D, 2E). Further, exosome derived from lncRNA ZFAS1 transfection HCM performed up-regulated of lncRNA ZFAS1.
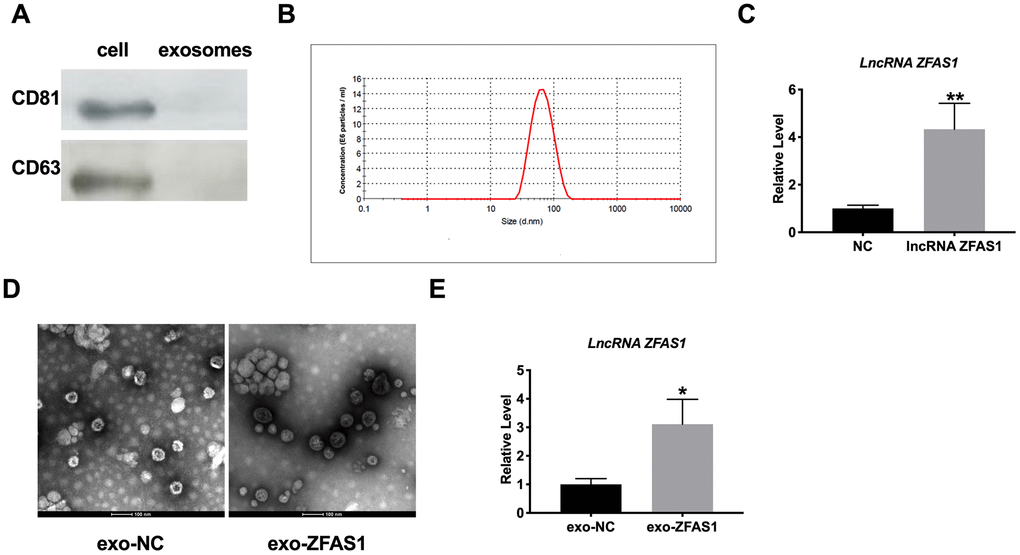
Figure 2. Isolation and characterization of exosomes derived from HCMs. (A) CD81 and CD63 were detected by western blot. (B) Exosome particle size detection. (C) The expression level of lncRNA ZFAS1 was detected in HCMs. (D) TEM was used to determine the existence and morphology of exosomes. (E) The expression level of lncRNA ZFAS1 was detected in exosome. *P<0.05, **P<0.01.
Exosome-lncRNA ZFAS1 regulated fibrosis in human cardiac fibroblast
We add exosome derived from lncRNA ZFAS1/NC (exo-ZFAS1/exo-NC) transfection cell into HCF. Compared with TGF-β1 treatment, we also observed that exo-ZFAS1 induced the morphology of cells changed from fibroblasts to myofibroblasts (Figure 3A). Then we detected the expression level of Col1a1, Col3a1, MMP2, MMP9, α-SMA, and CTGF by performing RT-PCR assay. The results showed that exo-ZFAS1 administrated also induced the increase in levels of Col1a1, Col3a1, MMP2, MMP9, α-SMA, CTGF, which were similar to TGF-β1 (Figure 3B). Further, we assessed the protein level of fibrosis-related markers, Col1a1, TGF-β1, Fibronectin, α-SMA. While exo-ZFAS1 performed a similar function with TGF-β1 in HCF (Figure 3C).
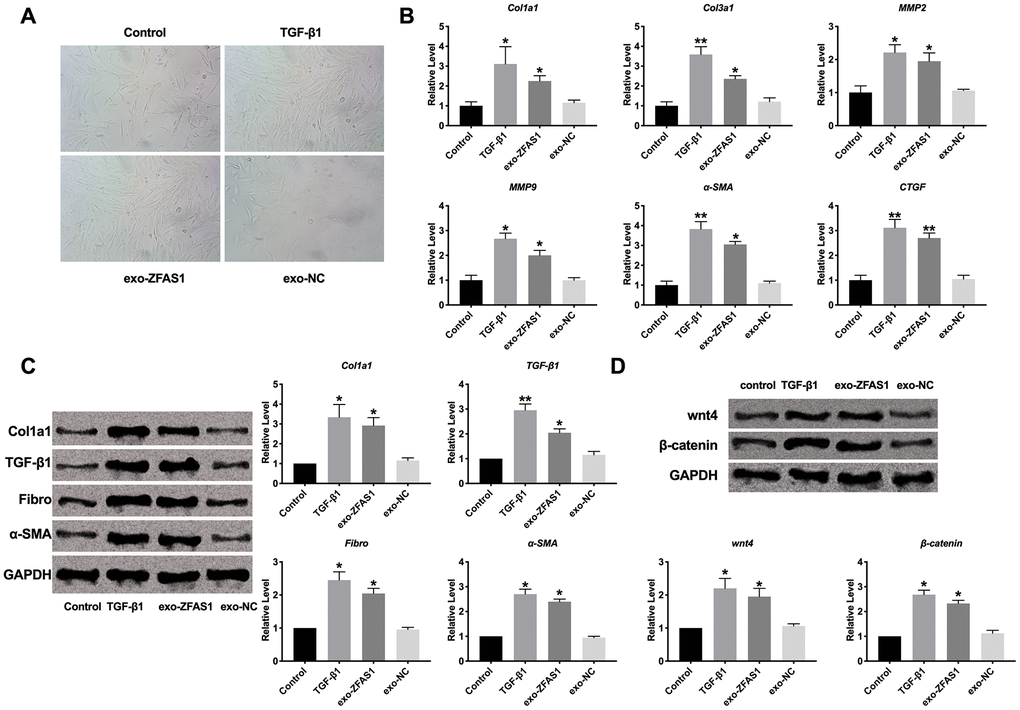
Figure 3. Exosome-lncRNA ZFAS1 regulated fibrosis in human cardiac fibroblasts. (A) Cell morphology detection. (B) RT-PCR assay was performed to measure the expression of Col1a1, Col3a1, MMP2, MMP9, α-SMA, and CTGF. (C) The expression level of Col1a1, TGF-β1, Fibronectin, α-SMA were assessed in HCFs. (D) The effect of exosome-lncRNA ZFAS1 on the wnt4/β-catenin signal pathway. *P<0.05.
After TGF-β1 treatment, the expression of wnt4 and β-catenin were up-regulated, the similar results were also obtained after exo-ZFAS1 administrated (Figure 3D). In summary, exo-ZFAS1 may regulate fibrosis in HCF by effecting wnt4/β-catenin.
Exosome-lncRNA ZFAS1 aggravated cardiac fibrosis in CKD mice
Next, we treated exosomes into mice via tail vein injection. The cardiac function in mice was detected by echocardiography. Exo-ZFAS1 significantly aggravated the decrease of cardiac function in CKD mice by comparing the changes of Ejection fraction (EF) and fraction shortening (FS) (Figure 4A). RT-PCR was employed to determine the expression of Col1a1, Col3a1, MMP2, MMP9, α-SMA, and CTGF in heart tissue. We found that exo-ZFAS1 significantly aggravated the level of fibrosis markers (Figure 4B). Masson experimental showed that exo-ZFAS1 accentuated fibrosis in CKD mice (Figure 4C). The level of Fibronectin was detected by immunohistochemical experiment. Exo-ZFAS1 induced expression of Fibronectin in CKD mice (Figure 4D). Further, we assessed the protein level of fibrosis-related markers, Col1a1, TGF-β1, Fibronectin, α-SMA. Meanwhile, exo-ZFAS1 induced expression of fibrosis-related markers in CKD mice (Figure 4E). The expression of wnt4 and β-catenin were induced by exo-ZFAS1 administrated (Figure 4F). Taken together, exo-ZFAS1 could promote cardiac fibrosis in CKD mice.
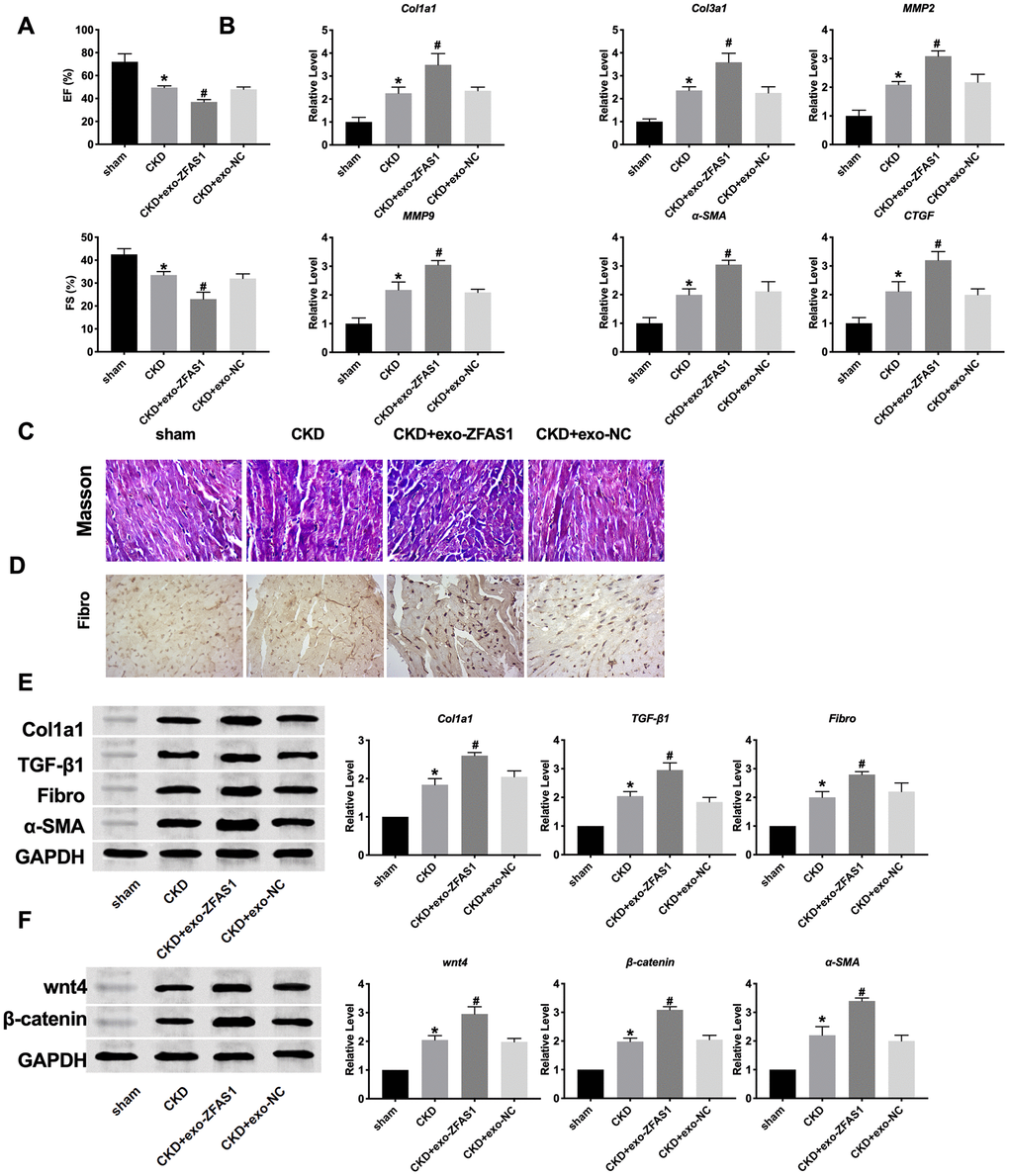
Figure 4. Exosome-lncRNA ZFAS1 aggravated cardiac fibrosis in CKD mice. (A) The cardiac function in mice was detected by echocardiography. (B) RT-PCR assay was performed to measure the expression of Col1a1, Col3a1, MMP2, MMP9, α-SMA, and CTGF. (C) The represented image of Masson. (D) The represented image of Immunohistochemistry for staining Fibronectin. (E) The expression level of Col1a1, TGF-β1, Fibronectin, α-SMA were assessed in heart tissues. (F) The effect of exosome-lncRNA ZFAS1 on the wnt4/β-catenin signal pathway. *P <0.05 vs. sham, #P <0.05 vs. CKD.
Exosome-lncRNA ZFAS1 promoted fibrosis in ex-vivo
For further study, we isolated CFs from the heart of sham and CKD mice. Then the CFs were cultured and treated with exosomes. The mRNA of Col1a1, Col3a1, MMP2, MMP9, α-SMA, and CTGF was induced by exo-ZFAS1 in CFs (Figure 5A). Then we observed that vimentin and α-SMA were up-regulated in exo-ZFAS1 by immunofluorescence (Figure 5B). Further, we assessed the protein level of Col1a1, TGF-β1, Fibronectin, α-SMA. Meanwhile, exo-ZFAS1 induced expression of fibrosis-related markers in CFs (Figure 5C, 5D). Consistent with the results in vivo, exo-ZFAS1 could also activate the wnt4/β-catenin signal pathway.
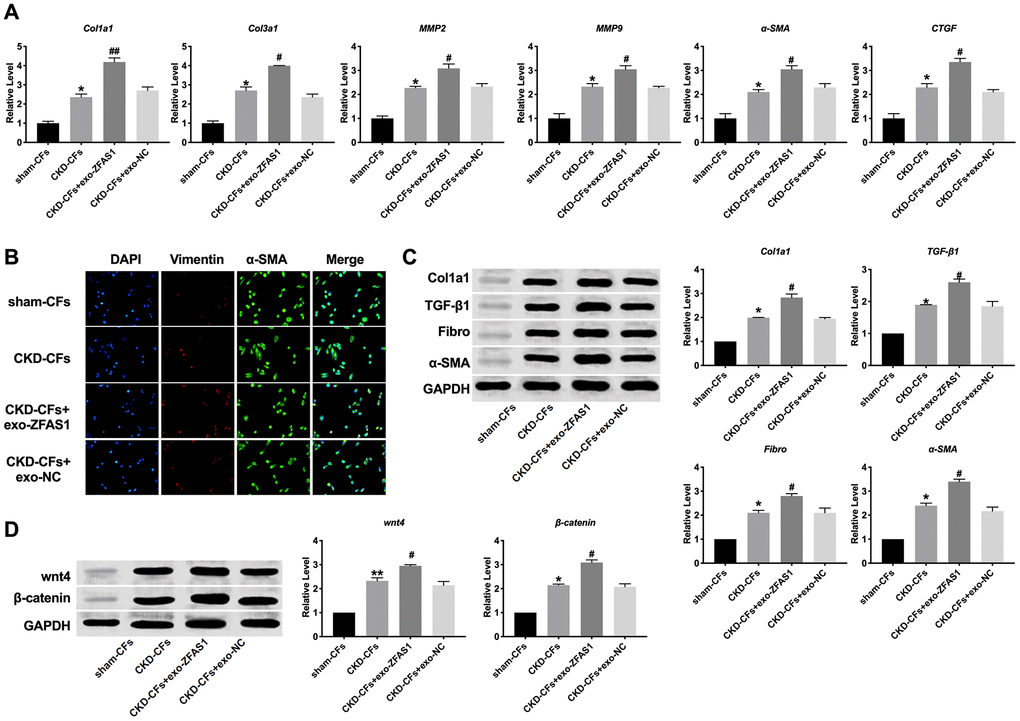
Figure 5. Exosome-lncRNA ZFAS1 promoted fibrosis in ex-vivo. (A) RT-PCR assay was performed to measure the expression of Col1a1, Col3a1, MMP2, MMP9, α-SMA, and CTGF. (B) The represented image of immunofluorescence for staining vimentin and α-SMA. (C) The expression level of Col1a1, TGF-β1, Fibronectin, α-SMA were assessed in CFs. (D) The effect of exosome-lncRNA ZFAS1 on the wnt4/β-catenin signal pathway. *P<0.05.,**P <0.01 vs. sham-CFs, #P <0.05, ##P <0.01 vs. CKD-CFs.
Silencing of lncRNA ZFAS1 prevents fibrosis in HCFs
Next, we constructed shRNA for interference with the expression of lncRNA ZFAS1. Then HCFs were transfection with sh-ZFAS1/sh-NC after TGF-β1 treatment. Then we explored the expression of Col1a1, Col3a1, MMP2, MMP9, α-SMA, and CTGF. Silencing of ZFAS1 inhibited the expression of Col1a1, Col3a1, MMP2, MMP9, α-SMA, and CTGF (Figure 6A). Then we observed that sh-ZFAS1 prevented the expression level of Col1a1, TGF-β1, Fibronectin, α-SMA (Figure 6B). Meanwhile, we found that sh-ZFAS1 inhibited the expression of wnt4 and β-catenin (Figure 6C). In summary, the silencing of ZFAS1 would prevent fibrosis in HCFs.
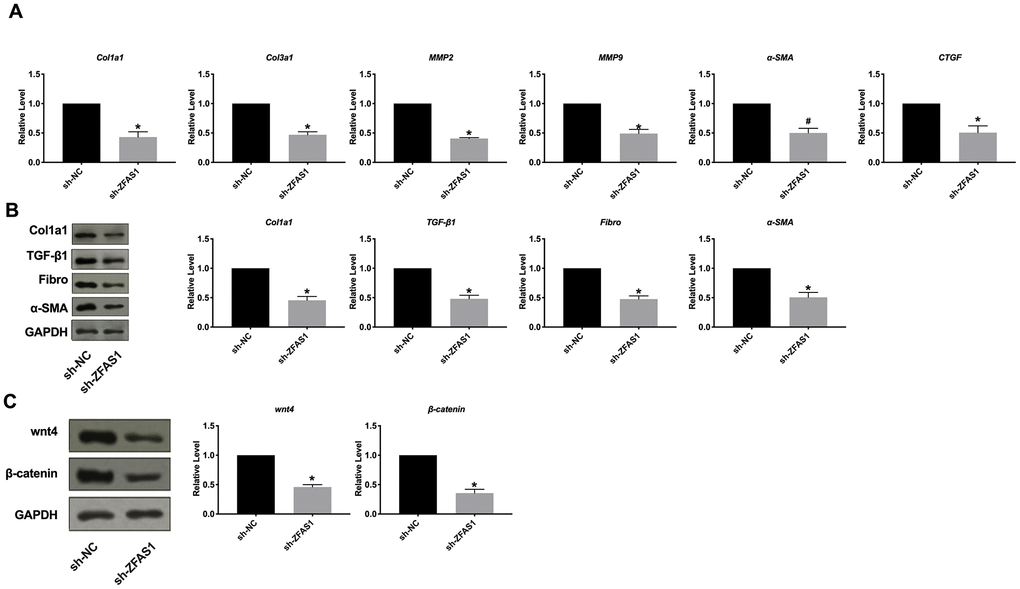
Figure 6. Silencing of lncRNA ZFAS1 blocked cardiac fibrosis in HCFs. (A) RT-PCR assay was performed to measure the expression of Col1a1, Col3a1, MMP2, MMP9, α-SMA, and CTGF. (B) The expression level of Col1a1, TGF-β1, Fibronectin, α-SMA were assessed in HCFs after sh-ZFAS1 transfection. (C) The effect of lncRNA ZFAS1 on the wnt4/β-catenin signal pathway. *P <0.05 vs. sh-NC.
LncRNA ZFAS1 regulated fibrosis via interacting with miR-4711-5p/wnt4
Previous studies have found that lncRNA regulates miRNA, miRNA mainly by acting as a precursor of miRNA, competitively combining mRNA with miRNA and "sponge effect". It can form a complex regulatory network by directly combining lncRNA or indirectly regulating lncRNA and miRNA through intermediate factors. The biological website predicted the existence of binding sites between lncRNA ZFAS1 and miR-4711-5p. Luciferase assay reported that lncRNA ZFAS could bind with miR-4711-5p (Figure 7A). HCFs were transfected with miR-4711-5p mimic after exosome and TGF-β1 treatment. Overexpression of miR-4711-5p could block fibroblasts transforming into myofibroblasts (Figure 7B). The increased mRNA level of Col1a1, Col3a1, MMP2, MMP9, α-SMA, and CTGF was prevented by a miR-4711-5p mimic in HCFs (Figure 7C). Similarly, Overexpression of miR-4711-5p inhibited the level of vimentin and α-SMA (Figure 7D). Meanwhile, wnt4 was an underlying target of miR-4711-5p (Figure 7E). The protein level of wnt4 and β-catenin were prevented by force of miR-4711-5p (Figure 7F). Taken together, lncRNA ZFAS1 could regulate the wnt4/ β-catenin signal pathway by interacting with miR-4711-5p.
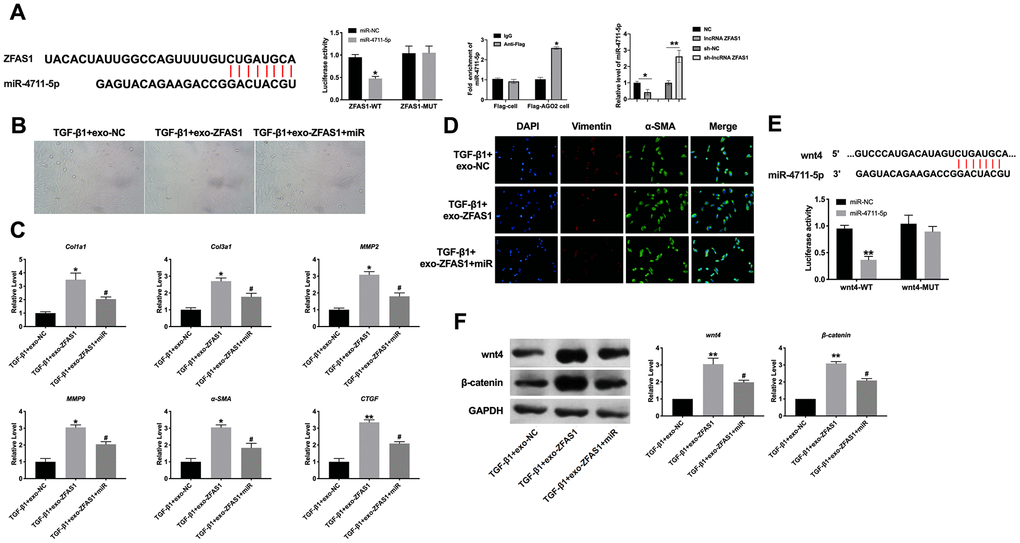
Figure 7. LncRNA ZFAS1 regulated fibrosis via interacting with miR-4711-5p/wnt4. (A) The biological website predicted the existence of binding sites between lncRNA ZFAS1 and miR-4711-5p. Luciferase assay confirmed the relationship between lncRNA ZFAS1 with miR-4711-5p. (B) Cell morphology detection. (C) RT-PCR assay was performed to measure the expression of Col1a1, Col3a1, MMP2, MMP9, α-SMA, and CTGF. (D) The represented image of immunofluorescence for staining vimentin and α-SMA. (E) The biological website predicted the existence of binding sites between wnt4 and miR-4711-5p. (F) The effect of lncRNA ZFAS1 and miR-4711-5p on the wnt4/β-catenin signal pathway. *P<0.05, **P<0.01 vs. TGF-β1+exo-NC group. #P <0.05 vs. TGF-β1+exo-ZFAS1.
Discussion
Chronic kidney disease is an independent risk factor for cardiovascular disease (CVD). Mild renal dysfunction can lead to an increase in the incidence of CVD. CVD in patients with CKD includes coronary artery disease, cardiomyopathy, cardiac valvular disease, cerebrovascular disease, and peripheral vascular disease [21]. Early evaluation and timely intervention of the risk factors of CVD are of considerable significance to reduce complications, improve the quality of life, and improve the prognosis of patients with CKD [22].
Exosomes are a kind of nano-vesicles released by cells, which can be released by a variety of cells, including cardiomyocytes, hepatocytes, and a variety of stem cells. Exosomes act as communication between cells. They carry messenger RNA, non-coding RNA, and proteins involved in almost all pathophysiological processes. As a communication medium between cells, exosomes participate in the pathophysiological process of tissue fibrosis. It has been found that exosomes derived from mesenchymal stem cells can improve the cardiac microenvironment after myocardial injury, promote angiogenesis, reduce inflammation, and inhibit ventricular remodeling. Exosomes contain miR-221 and miR-22 can also inhibit p53-induced cardiomyocyte apoptosis and reduce cardiac fibrosis [23]. This series of protective effects are closely related to the activation of the IGF-1/PI3K/Akt signal pathway. When acute myocardial infarction occurs, the secretion of cardiomyocyte exosomes increases. The secretion of exosomes around infarction is different from that of healthy myocardium. The released exosomes contain heart-specific miRNA-133, which has anti-apoptosis and anti-fibrosis effects [24].
At present, little is known about the mechanism of the formation and release of exosomes, which is a required field to be studied in the future. The functional molecules carried by exosomes are affected by the type of mother cells and the environment of mother cells, and the functions of different target cells are also different [25, 26]. The exosomes released after tissue cell injury can trigger inflammation, cause repair and regeneration response, and lead to tissue fibrosis [27, 28]. Therefore, exosomes and their contents can be used as new markers for the diagnosis of tissue fibrosis to avoid tissue and organ damage caused by biopsy, but the specific exosomes of tissue fibrosis need to be further studied.
Wnt/β-catenin signal pathway is one of the classic signal pathways to regulate cell proliferation, differentiation, and apoptosis, and the activation of the wnt/β-catenin signal pathway has been proved to affect the formation of cardiac fibroblasts and induce cardiomyocyte hypertrophy and fibrosis [29]. Wnt/β-catenin signaling pathway can regulate the progression of myocardial fibrosis by promoting the activation of fibroblasts, secretion of cytokines, and production of collagen [30]. It was reported that the wnt4/β-catenin signal pathway involved in renal fibrosis progression. However, there was no research on wnt4 in cardiac fibrosis [31]. Here, we found that wnt4 would be downstream of lncRNA ZFAS1 in regulating cardiac fibrosis. In future research, we would explore the underlying mechanism of wnt4 in cardiac diseases with CKD.
Cardiovascular disease is the leading cause of death in patients with chronic kidney disease, and chronic kidney disease also has a negative effect on the prognosis of patients with cardiovascular disease. To this end, based on the experiences and lessons learned from the study of nephropathy and cardiovascular disease, strengthening the relationship between cardiology and nephrology and establishing integrated therapy will provide the best diagnosis and treatment for these patients and significantly improve their clinical prognosis.
Conclusions
In our results performed an underlying mechanism on lncRNA ZFAS1 in cardiac fibrosis of CKD. We indicated that lncRNA ZFAS1 could transfer via exosome from cardiomyocytes to cardiac fibroblasts. We also showed lncRNA ZFAS1 regulated the cardiac fibrosis by WNT4/β-catenin signaling pathway, and miR-4711-5p involved in this pathway.
Materials and Methods
Mice model of chronic kidney disease
C57BL/6 mice were purchased from the Beijing Charles river and fed in the experimental Animal Center of Cangzhou Central Hospital of Hebei Province with the standard condition. For creating the CKD model, mice were fed with 0.2% adenine (# D191101) for 6 weeks. A week before the establishment of the CKD model, mice were injected with exosome via the tail vein. Six weeks after the establishment of the model, mice were intraperitoneally injected with 3% pentobarbital sodium and were euthanatized by excessive anesthesia with a dose of 90 mL/kg, and the organs and tissues were removed for follow-up study The research protocol of this study was approved by the Animal Care and Use Committee of Cangzhou Central Hospital of Hebei Province
Cell culture
Human cardiomyocytes and human cardiac fibroblasts were cultured in Dulbecco’s modified Eagle’s medium (Waltham, USA) supplemented with 10 % FBS (Cromwell, USA) and 100 μL/mL penicillin and streptomycin (Sigma-Aldrich, USA) and placed at 37° C with 5% CO2.
For TGFβ1 treatment, TGFβ1 (R&D Systerm, USA) were firstly dissolved in DMSO (Sigma, USA) and further diluted to 2 ng/ml.
Isolation and culture of adult mouse cardiac fibroblasts
The mice heart was removed and put into precooled PBS, the atrium and excess connective tissue were cut off, the cardiac cavity was cut open to remove blood clots, and the heart was rinsed repeatedly with precooled PBS for 3 or 4 times. The ventricle was cut into tissue blocks of about 1 mm × 1 mm × 1 mm and inhaled into the sterilized test tube. The tissue blocks were rewashed with precooled PBS until the liquid was clear and the supernatant was absorbed. Digestive juices (1 g Ca2+ L trypsin and 0.8 g / L type IV collagenase) were prepared with PBS without GMP and filtered to remove bacteria. Add 6 mL digestive juice to the tissue block, shake and digest 6 min; in a water bath at 37° C, gently blow away the sticky tissue block with an elbow straw, put it in a water bath shaker for 6 min, then blow repeatedly; after standing for a while, move most of the supernatant into a 50 mL centrifuge tube, add the same volume of DMEM containing 100 mL/L FBS to terminate digestion, and place on ice. 6 mL digestive juice was added to the remaining tissue mass, and the above digestion steps were repeated until the tissue mass was digested, leaving only a white transparent collagen fiber network (about 2 to 3 times). All the cell suspensions were filtered with a 200-mesh filter, then centrifuged for 10 min at 4° C, 250° C and 300 g, and precipitated with 30 mL DMEM resuspension cells containing 100 mL FBS, which were evenly divided into 3 petri dishes with a diameter of 10 cm. After 90 min, differential adhesion in absolute static culture at 37° C incubator, the unattached cells were washed and discarded by cross shaking petri dish (the unattached cells contained cardiomyocytes, and the supernatant could be transferred into a new petri dish to continue culture and purification if necessary). After repeated washing of PBS at 37° C, 10 mL of DMEM, containing 100 mL of FBS was put into the incubator for static culture, and the liquid was changed every 3 days.
Isolation and identification of exosome
When the cell fusion reached 70%, it was washed three times with 0.01mol/L phosphate buffer (PBS) and incubated with serum-free medium or exosome-free FBS medium for 48 hours. the supernatant was collected, centrifuged, 300xg centrifuged 10min at 4° C to remove cells, 2000xg centrifugation 20min to remove cell fragments, and exudates were stored at-80° C. Exosome morphology was observed by transmission electron microscope (TEM). The exosome particle size was detected by PSS Nicomp 380Z3000 analyzer (USA).
Quantitative real-time PCR
RNA isolation, reverse transcription and quantitative expression were carried according to manufacturer’s instructions. All the kits were purchased from Vazyme, and gene expression was calculated using 2-ΔΔCt method. The primer sequences (5’-3’) were shown as follow.
ZFAS1-F: CTATTGTCCTGCCCGTTAGAGp; ZFAS1-R: GTCAGGAGATCGAAGGTTGTAG; GAPDH-F: GATTCCACCCATGGCAAATTC; GAPDH-R: CTGGAAGATGGTGATGGGATT; Col1a1-F: CAAGACGAAGACATCCCACCAATC; Col1a1-R: ACAGATCACGTCATCGCACAACA; Col3a1-F: TCGCTCTGCTTCATCCCACTAT; Col3a1-R: CTTCCAGACATCTCTATCCGCAT; MMP2-F: AGTTTCCATTCCGCTTCCAG; MMP2-R: CGGTCGTAGTCCTCAGTGGT; MMP9-F: GTCCACCCTTGTGCTCTTCC; MMP9-R: GACTCTCCACGCATCTCTG; CTGF-F: GAAAGACAGGTACTAGCTGA; CTGF-R: CGTACCATATGTTCTGACAG; miR-4711-5P-F: UGCAUCAGGCCAGAAGACAUGAG; miR-4711-5P-R: CUCAUGUCUUCUGGCCUGAUGCA.
Dual luciferase reporter assay
20 mmol/L miR-4711-5p or negative control (NC) and lncRNA ZFAS1/wnt4 were co-transfected into HEK293T cells. Luciferase activity was detected with Luciferase Reporter Assay Kit (Biovision, China) on luminometer (Berthold, Germany) 48 hr after the transfection.
Western blot analysis
Total protein was collected from heart tissues or cardiomyocytes with RIPA lysis Mix. The western blotting assay was performed as previously described. Briefly, 50-80 μg protein extraction was loaded via SDS-PAGE and transferred onto nitrocellulose membranes (absin, China), then incubated with primary antibodies for 2 hrs at temperature, then plated at 4 °C overnight, the membranes were incubated in 5% non-fat milk blocking buffer for horizontal mode 3 h. After incubation with secondary antibodies, the membranes were scanned using an Odyssey, and data were analyzed with Odyssey software (LI-COR, USA).
Histological analysis
The heart tissues were gathered and fixed in 4% paraformaldehyde for 24 hrs. Then the fixed tissues were embedded in paraffin. Next, Paraffin slicer machine was used to cut slices (5-mm cross-sectional). H&E staining was used to evaluate cardiac morphology.
Immunohistochemical staining
Paraffin sections of lung tissue were dewaxing to water in xylene and descending series of ethanol. We penetrated parts using 0.5% Triton X-100. After 3 times wash, we blocked sections with 50% goat serum. Then, sections were incubated with the claudin-5 antibody overnight. We incubated the sections using a secondary antibody followed by DAPI staining. The sections were photographed by light scope under an IX73 fluorescence microscope (Olympus, Valley, PA) and analyzed by Image J software.
Immunofluorescence
In the culture plate, the slides that had already climbed the cells were washed with PBS for 3 times, fixed with 4% paraformaldehyde for15 min and washed 3 times with PBS. 0.5% Triton X Mel 100 (prepared by PBS) permeable at room temperature for 20 min. PBS slides were washed for 3 times, each time 3 min, the absorbent paper was used to dry PBS, normal goat serum was dripped on the slides, 30 min; the absorbent paper was sealed at room temperature to absorb the sealing solution, and no washing was done. Each slide was dripped with enough diluted first antibody and put into a wet box, incubated overnight at 4° C. Add fluorescent second antibody: PBST soak the climbing tablets for 3 times, each time 3 min, absorbent paper absorbs the excess liquid on the climbing tablets, then add diluted fluorescent secondary antibodies, incubate PBST sections 3 times at 37° C for 1 hour in a wet box, 3 min; re-staining nuclei each time: add DAPI to avoid light and incubate for 5 min, and PBST 5 min × 4 times to wash off the excess DAPI; Dry the liquid on the climbing sheet with absorbent paper, seal the film with the sealing solution containing anti-fluorescence quenching agent, and then observe and collect the image under the fluorescence microscope.
Statistical analysis
All data is statistical as a mean ± S.E.M. We performed Student's t-test or a one-way ANOVA for statistical analysis. P < 0.05 was described as statistically significant.
Author Contributions
Yunfei Wang, Xufen Cao, and Liqiu Yan made substantial contributions to the design of the study; Ye Zheng, Jing Yu, and Fuyun Sun drafted the work or revised it; Zheng Lian and Lina Sun approved the version to be published.
Conflicts of Interest
The authors declare that there are no conflicts of interest.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
References
- 1. Damman K, Testani JM. The kidney in heart failure: an update. Eur Heart J. 2015; 36:1437–44. https://doi.org/10.1093/eurheartj/ehv010 [PubMed]
- 2. Nessim SJ, Bargman JM. The peritoneal-renal syndrome. Nat Rev Nephrol. 2013; 9:302–06. https://doi.org/10.1038/nrneph.2013.43 [PubMed]
- 3. Rafiq K, Noma T, Fujisawa Y, Ishihara Y, Arai Y, Nabi AH, Suzuki F, Nagai Y, Nakano D, Hitomi H, Kitada K, Urushihara M, Kobori H, et al. Renal sympathetic denervation suppresses de novo podocyte injury and albuminuria in rats with aortic regurgitation. Circulation. 2012; 125:1402–13. https://doi.org/10.1161/CIRCULATIONAHA.111.064097 [PubMed]
- 4. van Kimmenade RR, Januzzi JL
Jr , Baggish AL, Lainchbury JG, Bayes-Genis A, Richards AM, Pinto YM. Amino-terminal pro-brain natriuretic Peptide, renal function, and outcomes in acute heart failure: redefining the cardiorenal interaction? J Am Coll Cardiol. 2006; 48:1621–7. https://doi.org/10.1016/j.jacc.2006.06.056 [PubMed] - 5. Ruiz-Hurtado G, Sarafidis P, Fernández-Alfonso MS, Waeber B, Ruilope LM. Global cardiovascular protection in chronic kidney disease. Nat Rev Cardiol. 2016; 13:603–08. https://doi.org/10.1038/nrcardio.2016.48 [PubMed]
- 6. Global Burden of Metabolic Risk Factors for Chronic Diseases Collaboration. Cardiovascular disease, chronic kidney disease, and diabetes mortality burden of cardiometabolic risk factors from 1980 to 2010: a comparative risk assessment. Lancet Diabetes Endocrinol. 2014; 2:634–47. https://doi.org/10.1016/S2213-8587(14)70102-0 [PubMed]
- 7. Sarnak MJ, Amann K, Bangalore S, Cavalcante JL, Charytan DM, Craig JC, Gill JS, Hlatky MA, Jardine AG, Landmesser U, Newby LK, Herzog CA, Cheung M, et al, and Conference Participants. Chronic kidney disease and coronary artery disease: JACC state-of-the-art review. J Am Coll Cardiol. 2019; 74:1823–38. https://doi.org/10.1016/j.jacc.2019.08.1017 [PubMed]
- 8. Ranjan P, Kumari R, Verma SK. Cardiac fibroblasts and cardiac fibrosis: precise role of exosomes. Front Cell Dev Biol. 2019; 7:318. https://doi.org/10.3389/fcell.2019.00318 [PubMed]
- 9. Chen GH, Xu J, Yang YJ. Exosomes: promising sacks for treating ischemic heart disease? Am J Physiol Heart Circ Physiol. 2017; 313:H508–23. https://doi.org/10.1152/ajpheart.00213.2017 [PubMed]
- 10. Davidson SM, Yellon DM. Exosomes and cardioprotection - a critical analysis. Mol Aspects Med. 2018; 60:104–14. https://doi.org/10.1016/j.mam.2017.11.004 [PubMed]
- 11. Salem ES, Fan GC. Pathological effects of exosomes in mediating diabetic cardiomyopathy. Adv Exp Med Biol. 2017; 998:113–38. https://doi.org/10.1007/978-981-10-4397-0_8 [PubMed]
- 12. Wang X, Gu H, Huang W, Peng J, Li Y, Yang L, Qin D, Essandoh K, Wang Y, Peng T, Fan GC. Hsp20-mediated activation of exosome biogenesis in cardiomyocytes improves cardiac function and angiogenesis in diabetic mice. Diabetes. 2016; 65:3111–28. https://doi.org/10.2337/db15-1563 [PubMed]
- 13. Shen T, Zheng Q, Luo H, Li X, Chen Z, Song Z, Zhou G, Hong C. Exosomal miR-19a from adipose-derived stem cells suppresses differentiation of corneal keratocytes into myofibroblasts. Aging (Albany NY). 2020; 12:4093–110. https://doi.org/10.18632/aging.102802 [PubMed]
- 14. Luo P, Jiang C, Ji P, Wang M, Xu J. Exosomes of stem cells from human exfoliated deciduous teeth as an anti-inflammatory agent in temporomandibular joint chondrocytes via miR-100-5p/mTOR. Stem Cell Res Ther. 2019; 10:216. https://doi.org/10.1186/s13287-019-1341-7 [PubMed]
- 15. Zhou Y, Ren H, Dai B, Li J, Shang L, Huang J, Shi X. Hepatocellular carcinoma-derived exosomal miRNA-21 contributes to tumor progression by converting hepatocyte stellate cells to cancer-associated fibroblasts. J Exp Clin Cancer Res. 2018; 37:324. https://doi.org/10.1186/s13046-018-0965-2 [PubMed]
- 16. Zhou X, Yan T, Huang C, Xu Z, Wang L, Jiang E, Wang H, Chen Y, Liu K, Shao Z, Shang Z. Melanoma cell-secreted exosomal miR-155-5p induce proangiogenic switch of cancer-associated fibroblasts via SOCS1/JAK2/STAT3 signaling pathway. J Exp Clin Cancer Res. 2018; 37:242. https://doi.org/10.1186/s13046-018-0911-3 [PubMed]
- 17. Weiskirchen R, Weiskirchen S, Tacke F. Organ and tissue fibrosis: Molecular signals, cellular mechanisms and translational implications. Mol Aspects Med. 2019; 65:2–15. https://doi.org/10.1016/j.mam.2018.06.003 [PubMed]
- 18. Andersson R, Refsing Andersen P, Valen E, Core LJ, Bornholdt J, Boyd M, Heick Jensen T, Sandelin A. Nuclear stability and transcriptional directionality separate functionally distinct RNA species. Nat Commun. 2014; 5:5336. https://doi.org/10.1038/ncomms6336 [PubMed]
- 19. Zhang Y, Jiao L, Sun L, Li Y, Gao Y, Xu C, Shao Y, Li M, Li C, Lu Y, Pan Z, Xuan L, Zhang Y, et al. LncRNA ZFAS1 as a SERCA2a inhibitor to cause intracellular Ca2+ overload and contractile dysfunction in a mouse model of myocardial infarction. Circ Res. 2018; 122:1354–68. https://doi.org/10.1161/CIRCRESAHA.117.312117 [PubMed]
- 20. Wu T, Wu D, Wu Q, Zou B, Huang X, Cheng X, Wu Y, Hong K, Li P, Yang R, Li Y, Cheng Y. Knockdown of Long Non-Coding RNA-ZFAS1 Protects Cardiomyocytes Against Acute Myocardial Infarction Via Anti-Apoptosis by Regulating miR-150/CRP. J Cell Biochem. 2017; 118:3281–289. https://doi.org/10.1002/jcb.25979 [PubMed]
- 21. Rossignol P, Agarwal R, Canaud B, Charney A, Chatellier G, Craig JC, Cushman WC, Gansevoort RT, Fellström B, Garza D, Guzman N, Holtkamp FA, London GM, et al. Cardiovascular outcome trials in patients with chronic kidney disease: challenges associated with selection of patients and endpoints. Eur Heart J. 2019; 40:880–86. https://doi.org/10.1093/eurheartj/ehx209 [PubMed]
- 22. Lees JS, Welsh CE, Celis-Morales CA, Mackay D, Lewsey J, Gray SR, Lyall DM, Cleland JG, Gill JMR, Jhund PS, Pell J, Sattar N, Welsh P, Mark PB. Glomerular filtration rate by differing measures, albuminuria and prediction of cardiovascular disease, mortality and end-stage kidney disease. Nat Med. 2019; 25:1753–1760. https://doi.org/10.1038/s41591-019-0627-8 [PubMed]
- 23. Powers JC, Sabri A, Al-Bataineh D, Chotalia D, Guo X, Tsipenyuk F, Berretta R, Kavitha P, Gopi H, Houser SR, Khan M, Tsai EJ, Recchia FA. Differential microRNA-21 and microRNA-221 upregulation in the biventricular failing heart reveals distinct stress responses of right versus left ventricular fibroblasts. Circ Heart Fail. 2020; 13:e006426. https://doi.org/10.1161/CIRCHEARTFAILURE.119.006426 [PubMed]
- 24. Lin F, Zeng Z, Song Y, Li L, Wu Z, Zhang X, Li Z, Ke X, Hu X. YBX-1 mediated sorting of miR-133 into hypoxia/reoxygenation-induced EPC-derived exosomes to increase fibroblast angiogenesis and MEndoT. Stem Cell Res Ther. 2019; 10:263. https://doi.org/10.1186/s13287-019-1377-8 [PubMed]
- 25. Shi Y, Wang Y, Li Q, Liu K, Hou J, Shao C, Wang Y. Immunoregulatory mechanisms of mesenchymal stem and stromal cells in inflammatory diseases. Nat Rev Nephrol. 2018; 14:493–507. https://doi.org/10.1038/s41581-018-0023-5 [PubMed]
- 26. Dougherty JA, Patel N, Kumar N, Rao SG, Angelos MG, Singh H, Cai C, Khan M. Human cardiac progenitor cells enhance exosome release and promote angiogenesis under physoxia. Front Cell Dev Biol. 2020; 8:130. https://doi.org/10.3389/fcell.2020.00130 [PubMed]
- 27. Wermuth PJ, Piera-Velazquez S, Rosenbloom J, Jimenez SA. Existing and novel biomarkers for precision medicine in systemic sclerosis. Nat Rev Rheumatol. 2018; 14:421–32. https://doi.org/10.1038/s41584-018-0021-9 [PubMed]
- 28. Jung JH, Fu X, Yang PC. Exosomes generated from iPSC-derivatives: new direction for stem cell therapy in human heart diseases. Circ Res. 2017; 120:407–17. https://doi.org/10.1161/CIRCRESAHA.116.309307 [PubMed]
- 29. Tao H, Yang JJ, Shi KH, Li J. Wnt signaling pathway in cardiac fibrosis: new insights and directions. Metabolism. 2016; 65:30–40. https://doi.org/10.1016/j.metabol.2015.10.013 [PubMed]
- 30. Yang D, Fu W, Li L, Xia X, Liao Q, Yue R, Chen H, Chen X, An S, Zeng C, Wang WE. Therapeutic effect of a novel Wnt pathway inhibitor on cardiac regeneration after myocardial infarction. Clin Sci (Lond). 2017; 131:2919–2932. https://doi.org/10.1042/CS20171256 [PubMed]
- 31. DiRocco DP, Kobayashi A, Taketo MM, McMahon AP, Humphreys BD. Wnt4/β-catenin signaling in medullary kidney myofibroblasts. J Am Soc Nephrol. 2013; 24:1399–412. https://doi.org/10.1681/ASN.2012050512 [PubMed]