



Introduction
Primary angle-closure glaucoma (PACG) is the leading cause of irreversible blindness, which affected the worldwide population estimated to exceed 20 million in 2020 and over 30 million by 2040, with Asia accounting for 76% of PACG cases [1–2]. The prevalence of glaucoma varies significantly between different geographical regions and racial groups [3]. It is more common (60%) among women [4]. Based on recent reports, the prevalence of PACG in mainland China is estimated to be 1.4% (95% credible interval 1.0% to 1.7%) [5]. However, the prevalence of PACG among European derived populations aged 40 and over is only 0.4% (95% credible interval 0.3% to 0.5%) [6]. The pathogenesis of PACG is caused by direct contact between the iris and smooth muscle-like trabecular meshwork, which obstructs the aqueous humor pathway [7]. This obstruction prevents the exit of aqueous humor, thereby increasing the intraocular pressure, which is thought to degenerate the trabecular meshwork and damage the optic nerve [8]. The underlying causes of PACG remain unclear so far, but the genetic factors are well recognized in contributing to the pathogenesis of PACG [9].
During the past few years, genome-wide association studies have identified several PACG loci and genes, including GLC2A on chromosome 10q, PLEKHA7 on chromosome 11p, COL11A1 on chromosome 1p, MFRP on chromosome 11q [10]. SNPs in other candidate genes, including EPDR1, CHAT, GLIS3, FERMT2, DPM2-FAM102, also have a significant association with PACG [11]. Although numerous publications have pointed out these susceptibility genes that appear to contribute to developing PACG, strictly heritable disease-causing mutation has not been identified until Waseem et al. reported the first disease-causing mutation in SPATA13 gene [12]. The discovery of strictly heritable disease-causing mutation can shed light on the molecular mechanisms of PACG.
In the present study, we recruited a five-generation Chinese PACG family. The strictly heritable disease-causing mutation of this family was identified by whole-genome sequencing with bioinformatics analysis. Additionally, this mutation was confirmed as a loss-of-function mutation by various experiments.
Results
Clinical findings
This Chinese PACG family (Figure 1) comprises 18 members (10 males and 8 females). The proband III:2 at the age of 49 (black arrow) was enquired about a family history of glaucoma. The accuracy of family history information was confirmed by the corroboration of relatives attending the hospital. I:2 had been treated with conservative drug therapy for ocular hypertension for 35 years from age 60. On examination, all affected members were found to have the same phenotype as the proband III:2. The thickness of the optic nerve fiber layer in both eyes was significantly thinner. Direct contact of the iris root with the trabecular meshwork led to a closure of the anterior chamber angle (Figure 2). The proband’s mother II:3 and her uncle II:5 underwent surgery for glaucoma. This family has a strong history of glaucoma.
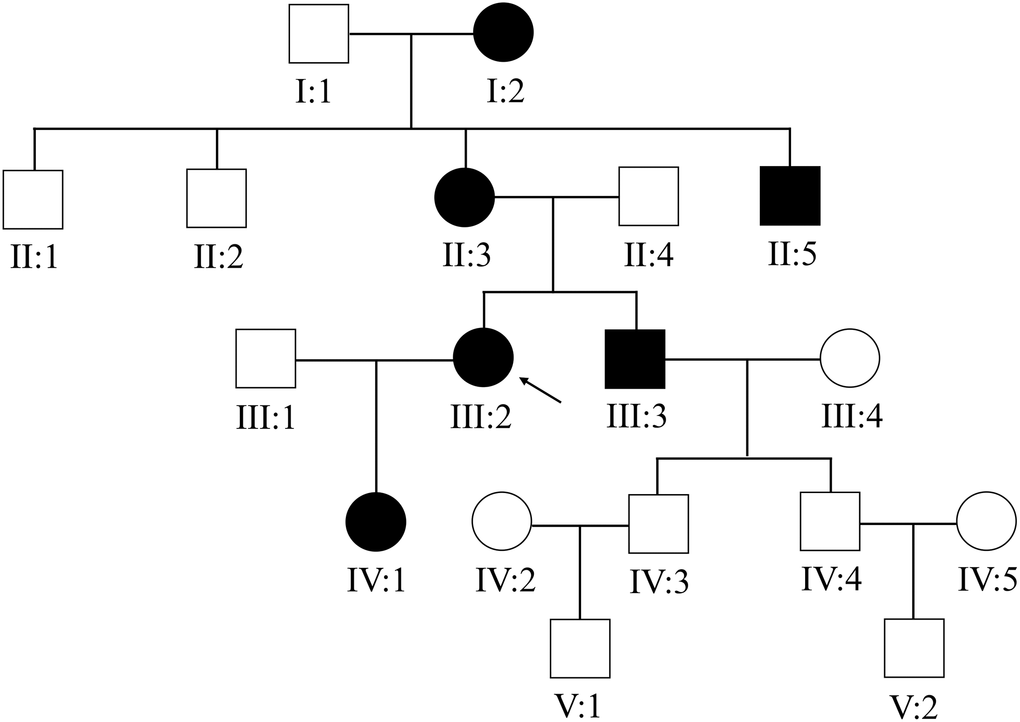
Figure 1. The Chinese family with primary angle-closure glaucoma. The pedigree of the five-generation Chinese PACG family is shown. Roman numerals indicate generations, and individuals within a generation are numbered from left to right. The proband (III:2) is denoted with an arrow.
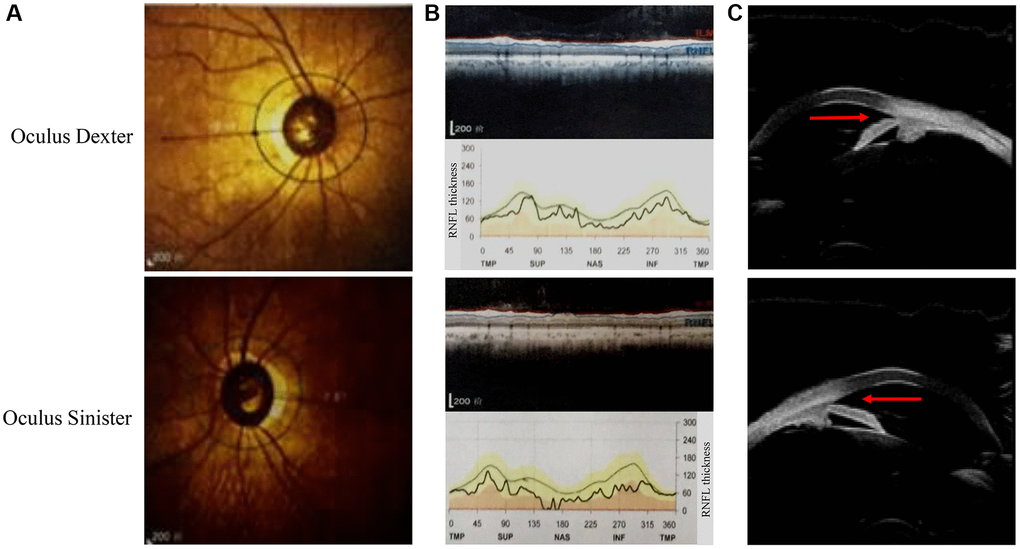
Figure 2. Ophthalmic examination of the proband in this pedigree. (A) The fundus angiography exam of the proband (III:2). (B) Retinal nerve fiber layer (RNFL) thickness is assessed by optical coherence tomography. (C) The results of ultrasonic biomicroscopy reveal relative pupillary block and a narrow-angle inlet.
Whole-genome sequencing and sanger sequencing
The whole-genome sequencing was performed to identify the disease-causing mutation. By using ClinPred, an efficient tool for identifying disease-relevant nonsynonymous variants, 18 variants (the scores >15) were identified as possible disease-causing mutations (Table 1). Sanger sequencing was applied to screen these potential pathogenic variants. A cytosine (C) to thymine (T) heterozygous missense mutation c.977C>T in phosphoenolpyruvate carboxykinase 2 (PCK2) gene was detected in all five affected family members (Figure 3), but not in any of the unaffected members and 86 unrelated individuals. The C to T nucleotide change at this locus is predicted to result in proline (P) to leucine (L) substitution p.Pro326Leu.
Table 1. The disease-causing mutation in candidate genes.
| Chr | Start | End | Ref | Alt | Gene | code | pro | ClinPred | CADD_Pred | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr10 | 112541205 | 112541205 | A | G | RBM20 | c.A838G | p.K280E | 0.610286035 | 16.53 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr10 | 32306144 | 32306144 | C | A | KIF5B | c.G2688T | p.Q896H | 0.991513669 | 21.8 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr12 | 58220831 | 58220831 | C | G | CTDSP2 | c.G302C | p.R101T | 0.965011537 | 22.7 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr12 | 76453592 | 76453592 | C | T | NAP1L1 | c.G415A | p.E139K | 0.888694346 | 19.66 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr13 | 32823672 | 32823672 | A | C | FRY | c.A7018C | p.I2340L | 0.893504195 | 33 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr14 | 24572375 | 24572375 | C | T | PCK2 | c.C977T | p.P326L | 0.986453235 | 31 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr14 | 65259743 | 65259743 | G | C | SPTB | c.C2638G | p.L880V | 0.957589209 | 17.56 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr15 | 54838931 | 54838931 | C | T | UNC13C | c.C5708T | p.S1903L | 0.666001618 | 16.71 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr16 | 61761045 | 61761045 | C | T | CDH8 | c.G1489A | p.A497T | 0.894139826 | 35 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr18 | 28914109 | 28914109 | T | C | DSG1 | c.T949C | p.W317R | 0.998477757 | 18.39 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr1 | 227069657 | 227069657 | C | T | PSEN2 | c.C49T | p.R17W | 0.887850463 | 20.4 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr1 | 227288725 | 227288725 | T | G | CDC42BPA | c.A1974C | p.L658F | 0.955309629 | 18.56 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr3 | 172737337 | 172737337 | G | A | SPATA16 | c.C787T | p.R263W | 0.998399913 | 24.7 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr5 | 162880987 | 162880987 | T | C | NUDCD2 | c.A460G | p.N154D | 0.776593506 | 16.95 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr5 | 96117517 | 96117517 | G | A | ERAP1 | c.C2327T | p.A776V | 0.823308527 | 19.39 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr6 | 34008409 | 34008409 | G | A | GRM4 | c.C937T | p.P313S | 0.982662201 | 21.9 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr7 | 75615259 | 75615259 | C | T | POR | c.C1688T | p.T563M | 0.837663054 | 19.31 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| chr8 | 144812605 | 144812605 | C | T | FAM83H | c.G148A | p.E50K | 0.966016054 | 23 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Abbreviations: Chr: Chromone; Start: variant of start position; End: variant of end position; Ref: reference base; Alt: mutant base; pro: protein; ClinPred: score >0.5; CADD_pred: score>15. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
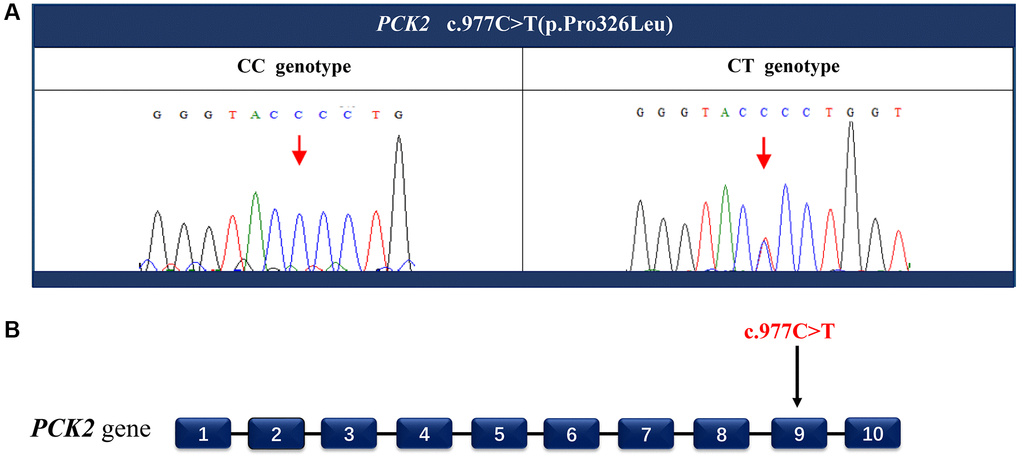
Figure 3. Identify the disease-causing mutation in the PACG family. A heterozygous missense mutation c.977C>T (A) located in exon 9 (B) of the PCK2 gene was detected by Sanger sequencing. This nucleotide substitution results in a proline to leucine substitution p.Pro326Leu.
Influence of mutations on cell cycle and apoptosis
To further investigate the function of PCK2 and determine the activity of the mutation on endogenous target genes, RGC-5 cells were stably transfected with constructs expressing PCK2 with c.977C>T mutation. The mutation induces apoptosis of RGC-5, which was demonstrated by Hoechst 33342/PI double-fluorescent chromatin staining (Figure 4A) and flow cytometry (Figure 4B). In order to explore the mechanism of c.977C>T mutation-induced apoptosis, the levels of proteins associated with cell apoptosis were determined by Western blotting and quantitative analysis. A decreased BcL-XL was detected in RGC-5 cell lines transferred with c.977C>T mutation (Figure 5). To further investigate the effect of c.977C>T mutation on RGC-5 cells, the expression of P53, P21, P27, AKT, and GSK3 were measured. The elevated P53, P21, and P27 were observed in RGC-5 cell lines transferred with c.977C>T mutation (Figure 6). In addition, we found that the expression of AKT and phosphorylation of GSK3α were increased (Figure 7).
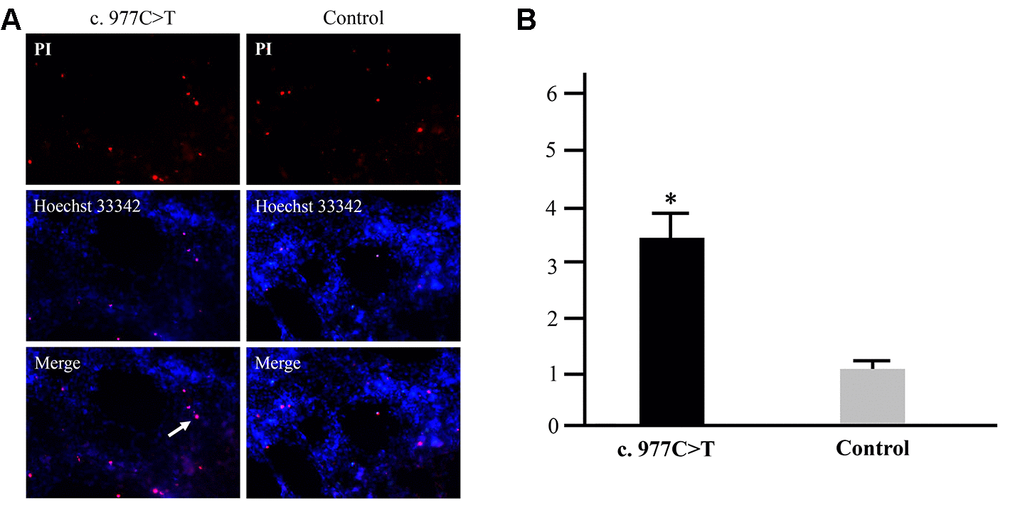
Figure 4. Apoptosis analysis of the mutation p.Pro326Leu. RGC-5 cells were transfected with wild-type PCK2 or p.Pro326Leu mutation, respectively. Multiple apoptotic bodies (a, pink dots) were observed in the cells transferred with p.Pro326Leu mutation (A), and the apoptosis was further proved by flow cytometry (B). *P < 0.05.
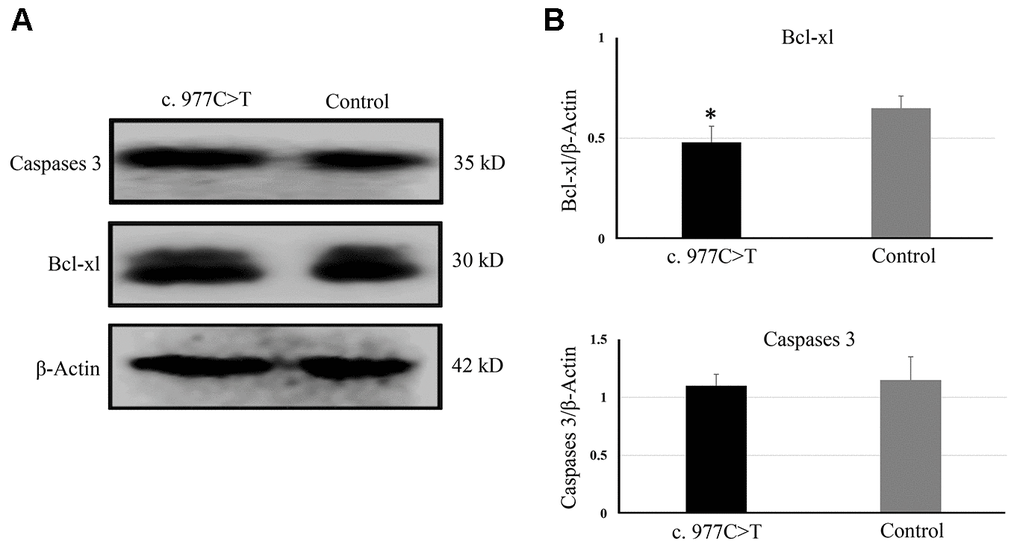
Figure 5. The expressions of genes related to apoptosis were determined by Western blotting (A) and quantitative analysis (B). *P < 0.05.
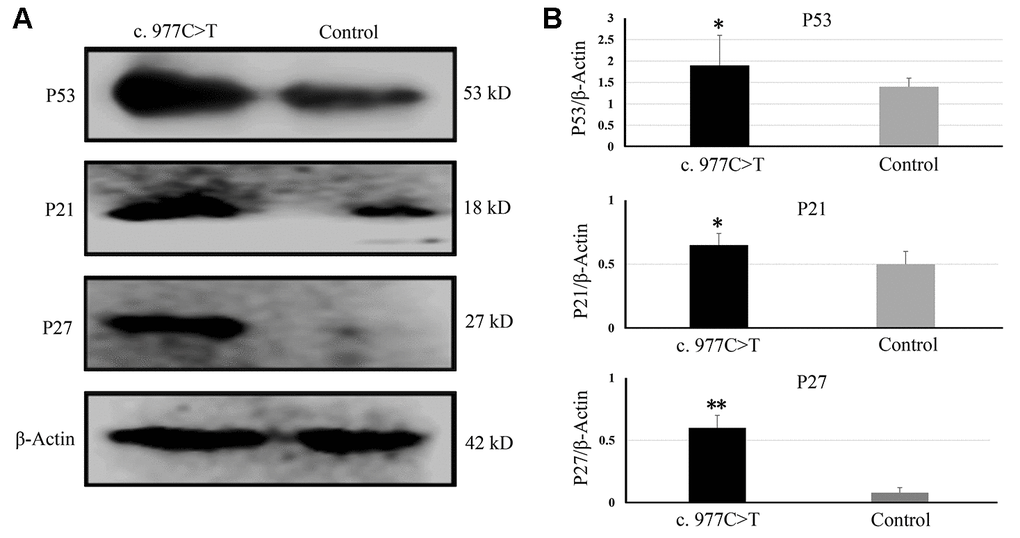
Figure 6. The expressions of genes related to the cell cycle and growth were determined by Western blotting (A) and quantitative analysis (B). *P < 0.05; **P < 0.01.
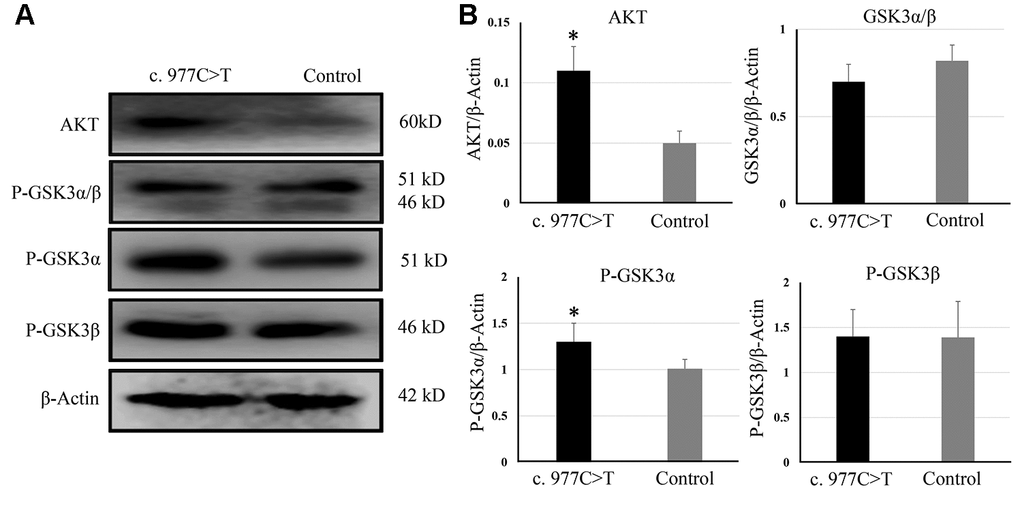
Figure 7. The expression of AKT, GSK3, P-GSK3α, and P-GSK3β were analyzed by Western blotting (A) and quantitative analysis (B). *P < 0.05.
Discussion
The pathogenesis of PACG is complex. Many different factors affect the progression of the anterior chamber angle from narrow to angle-closure [13]. Among them, heritable factors play an important role in the etiology of PACG [14]. Therefore, singling out heritable factors contributing to the disease is crucial for unraveling the pathway of PACG development. In 2020, Waseem et al. identified the first disease-causing mutation c.1432_1440del in the SPATA13 gene by analyzing a seven-generation British PACG family [12]. In the present study, we identified a novel disease-causing mutation c.977C>T in PCK2 in a five-generation Chinese PACG family. PCK2 is a mitochondrial enzyme that catalyzes the conversion of oxaloacetate to phosphoenolpyruvate [15]. A previous study reported that the expression PCK2 significantly was down-regulated in PACG [16], but the role of PCK2 in the pathogenies and development of PACG remains largely unclear. The previous study demonstrated that the function of PCK2 was to promote the growth of smooth muscle [17]. Hence, PCK2 might involve the pathogenies of PACG by regulating the growth of smooth muscle-like trabecular meshwork, which plays a central role in the development of PACG. PCK2 also is associated with lung cancer by promoting tumorigenesis through its gluconeogenic function [18]. Due to its gluconeogenic function, PCK2 deficiency is expected to disrupt glucose homeostasis and result in hypoglycemia related-disorder [19]. The non-synonymous somatic variant in PCK2 was identified in hepatocellular carcinoma tissue, suggesting that PCK2 was associated with hepatocellular carcinoma [20].
To further explore the effect of mutation c.977C>T, the mutation was constructed into lentiviral vector pCD513B and transfected into RGC-5 cells. The c.977C>T mutation induces apoptosis of RGC-5, suggesting that c.977C>T mutation is a function-loss mutation. A decreased Bcl-xl indicates that c.977C>T mutation induces apoptosis by regulating the Bcl-xl induced-apoptosis pathways. Bcl-xL is a transmembrane molecule in the mitochondria, which acts as an anti-apoptotic protein by preventing mitochondrial release, leading to caspase activation and programmed cell apoptosis [21–22]. In addition, elevated P53, P21, and P27 were observed in RGC-5 cells transfected with c.977C>T mutation. P53 and its primary target P21 is linked with DNA damage to cell cycle arrest [23–24]. P27 is a negative regulator of cell proliferation and arrests the cell cycle by inhibiting cyclin-dependent kinases [25]. All of these proteins have been widely confirmed to be involved in cell cycle arrest to cell apoptosis.
In the present study, we also detected an increase of AKT and phosphorylation of GSK3α in RGC-5 cells containing c.977C>T mutation. Impairment of AKT/GSK3 signaling pathway in some eye disorders such as glaucoma, diabetic retinopathy, and retinitis pigmentosa have been reported during the past few years [26–27]. The function of AKT is to phosphorylate GSK3, which is a crucial regulator of differentiation, metabolism, and apoptosis [28]. The mechanism of the AKT/GSK3 signaling pathway involved in eye disorder is that increased GSK3 promotes inflammation and therefore may play a role in glaucoma and age-related macular degeneration activity under disease conditions [29–30]. Interestingly, we found that c.977C>T mutation only induced elevated phosphorylation of GSK3α, suggesting that GSK3α was the target of PCK2 in the development of PACG. GSK3α and GSK3β are the two functional isoforms of GSK3, but whose functions remain ill-defined because of a lack of inhibitors that can distinguish between the two highly homologous isozymes [31–32]. The role of AKT and GSK3α in the pathogenesis of PACG needs to be further investigated.
In conclusion, we reported a novel heterozygous missense mutation in the PCK2 gene causing PACG by analyzing a five-generation Chinese PACG family. Moreover, this mutation-induced RGC-5 cells apoptosis and elevated P53, P21, P27, AKT, and GSK3α, suggesting that this mutation c.977C>T within PCK2 gene cause PACG through impairment of AKT/GSK3α signaling pathway. Identification of strictly heritable disease-causing mutation of PACG helps us to understand the glaucoma etiology.
Methods
Ethical compliance
Written consent was obtained from all participants. The study was approved by the Hospital Research Ethics Committee (2017-002-SX), following the principles of the Declaration of Helsinki of 1975, as revised in 2000.
Subjects and ophthalmic examination
All family members aged 18 years and older were invited to attend comprehensive ophthalmic examination, including fundus angiography, dark room gonioscopy, optical coherence tomography, ultrasonic biomicroscopy, visual field testing. The primary defining feature of cases was contact between the iris and trabecular meshwork identified on darkroom gonioscopy. PACG was diagnosed when there was evidence of iridotrabecular contact and glaucomatous damage to the optic nerve.
Whole-genome sequencing and bioinformatics analysis
DNA was extracted from peripheral blood using Gentra PureGene kit (Qiagen, China). 86 random human DNAs were set as healthy control. Exome capture was performed with an Illumina HiSeq 2500 (Illumina Inc., CA, USA), and the raw sequencing reads were aligned by the BIOTECAN (Shanghai, China) using the Burrows-Wheeler Aligner. After removing duplicates from the sorted alignment using Picard, variants were called using the Genome Analysis Toolkit (GATK v3.70) pipeline. ANNOVAR was used to annotate SNPs and insertions/deletions. Filtrations of all identified variations were performed with data from public databases, including the database in human reference genome hg19 from UCSC, dbSNP, 1000 genomes project, and AVSIFT. After excluding common variants, retained variants were considered to be ‘novel’. Pathogenicity of the variants was evaluated ClinPred Prediction Tool. The criteria to filter candidate mutations were: 1. Located in exonic and splitting regions; 2. The variants frequency <0.05 in 1000g2012apr_all, esp6500si_all, and ExAC_ALL database; 3. The variants with CADD_Pred score >15 suggests potentially pathogenic variants. Direct Sanger sequencing was applied to confirm these potential pathogenic variants with the ABI3500 sequencer (Applied Biosystems, CA, USA). The primers for PCK2 gene is 5′-CAGAGTTCTCCCCTGGTGAAT-3′ and 5′-AAAAGGAAGGTCTGGCTCAG-3′.
Site-directed mutagenesis
Total mRNA was isolated from cultured cells using the Oligotex mRNA kit (QIAGEN, China) and transcribed into cDNA using the Transcriptor Reverse Transcriptase System (Roche, China). The coding regions of PCK2 were amplified using high-fidelity DNA polymerase (New England Biolabs, China) and ligated into a lentiviral vector pCD513B. For site-directed mutagenesis, phosphorylated primers harboring the desired mutations were used to amplify the PCK2 constructs with DNA polymerase and ligated with Taq DNA ligase. Sanger sequencing was used to verify all cloned inserts.
Cell culture, virus packaging, and transfections
RGC-5 cell line and HEK293T were purchased from iCell Bioscience Inc, Shanghai, China. The cells were maintained in DMEM/F12 supplemented with 10% FBS and 1g/L glucose. HEK-293T cells were cultured in DMEM containing 10% FBS and 1% penicillin-streptomycin. All cells were grown in a 6-well plate at 1.2 × 106 cells per well at 37°C in an atmosphere of 5% CO2. The packaging plasmids (pGag/Pol, pRev, and pVSV-G), pCD513B-PCK2 or pCD513B-PCK2-977C>T mutation were co-transfected into HEK 293T cells. Lentivirus-containing supernatants were collected and stored at −80°C.
Flow cytometry
The effects of c.977C>T mutation on cell apoptosis were quantitatively analyzed by flow cytometry. RGC-5 cells in the logarithmic growth phase were inoculated into 6 cm dish and cultured for 48 hrs until the cells adhered to the wall. Following incubation, the cell culture medium was collected in a 10-mL centrifuge tube, and the cells were washed with PBS. Subsequently, the cells were centrifuged to remove the supernatant, re-mixed with binding buffer, and mixed with Annexin V-FITC and propidium iodide (PI) staining solution. After thorough mixing, the cells were incubated at room temperature in the dark for 15 min. PBS was added to the flow tube cell suspension, and a flow cytometry assay (Beckman Coulter) was performed.
SDS electrophoresis and western blotting
Proteins were separated by SDS-polyacrylamide gel electrophoresis and transferred to a polyvinylidene difluoride membrane. Immunoreactive bands were visualized using the ECL Plus detection kit (HaiGene Biotechnology, Harbin, China). Anti-P53 and anti-P21 were purchased from Proteintech, Wuhan, China. Other antibodies (anti-P27, anti-P21, anti-Caspase 3, anti-Bcl-xl, anti-AKT, anti- P-GSK3α, anti- P-GSK3β, and anti-β-actin) were purchased from Sangon Biotech, Shanghai, China.
Statistical analysis
Data are reported as the mean ± SD. Differences were considered statistically significant at P < 0.05. All statistical analyses were performed by using SPSS20.0 (SPSS Inc., Chicago, IL, USA).
Author Contributions
Z.L. and T.L. designed and conceived the study. M.X., J.Y., J.S., and X.X. performed the analysis of mutations in all the families. M.X., J.Y., and J.S. performed the functional experiments and analysis of data. J.Y., J.S., and T.L. evaluated and diagnosed the patients. M.X., J.Y., J.S., Z.L. and T.L. wrote the manuscript. Z.L. and T.L. critically revised the manuscript. All authors participated in discussion of the final manuscript and interpretation of results.
Acknowledgments
The authors want to thank the patients for their participation in the study.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Funding
This study was funded by Key Research and Development Program of Shaanxi Province (2017ZDXM-SF-067), Guangxi Science and Technology Base and Special Fund for Talents (2018AD19267/AD19110161), and Natural Science Foundation of Guangxi (2018JJA140045/2018GXNSFAA281048).
References
- 1. Kondkar AA. Updates on Genes and Genetic Mechanisms Implicated in Primary Angle-Closure Glaucoma. Appl Clin Genet. 2021; 14:89–112. https://doi.org/10.2147/TACG.S274884 [PubMed]
- 2. Liu C, Nongpiur ME, Cheng CY, Khor CC, Yu M, Husain R, Ho CL, Wong TT, Boey PY, Perera S, Wong TY, Vithana EN, Aung T. Evaluation of Primary Angle-Closure Glaucoma Susceptibility Loci for Estimating Angle Closure Disease Severity. Ophthalmology. 2021; 128:403–09. https://doi.org/10.1016/j.ophtha.2020.07.027 [PubMed]
- 3. Tham YC, Li X, Wong TY, Quigley HA, Aung T, Cheng CY. Global prevalence of glaucoma and projections of glaucoma burden through 2040: a systematic review and meta-analysis. Ophthalmology. 2014; 121:2081–90. https://doi.org/10.1016/j.ophtha.2014.05.013 [PubMed]
- 4. Zhang N, Wang J, Chen B, Li Y, Jiang B. Prevalence of Primary Angle Closure Glaucoma in the Last 20 Years: A Meta-Analysis and Systematic Review. Front Med (Lausanne). 2021; 7:624179. https://doi.org/10.3389/fmed.2020.624179 [PubMed]
- 5. Cheng JW, Cheng SW, Ma XY, Cai JP, Li Y, Wei RL. The prevalence of primary glaucoma in mainland China: a systematic review and meta-analysis. J Glaucoma. 2013; 22:301–06. https://doi.org/10.1097/IJG.0b013e31824083ca [PubMed]
- 6. Day AC, Baio G, Gazzard G, Bunce C, Azuara-Blanco A, Munoz B, Friedman DS, Foster PJ. The prevalence of primary angle closure glaucoma in European derived populations: a systematic review. Br J Ophthalmol. 2012; 96:1162–67. https://doi.org/10.1136/bjophthalmol-2011-301189 [PubMed]
- 7. Jonas JB, Aung T, Bourne RR, Bron AM, Ritch R, Panda-Jonas S. Glaucoma. Lancet. 2017; 390:2183–93. https://doi.org/10.1016/S0140-6736(17)31469-1 [PubMed]
- 8. Soh ZD, Thakur S, Majithia S, Nongpiur ME, Cheng CY. Iris and its relevance to angle closure disease: a review. Br J Ophthalmol. 2021; 105:3–8. https://doi.org/10.1136/bjophthalmol-2020-316075 [PubMed]
- 9. Wang J, Yusufu M, Khor CC, Aung T, Wang N. The genetics of angle closure glaucoma. Exp Eye Res. 2019; 189:107835. https://doi.org/10.1016/j.exer.2019.107835 [PubMed]
- 10. Wiggs JL, Pasquale LR. Genetics of glaucoma. Hum Mol Genet. 2017; 26:R21–27. https://doi.org/10.1093/hmg/ddx184 [PubMed]
- 11. Ahram DF, Alward WL, Kuehn MH. The genetic mechanisms of primary angle closure glaucoma. Eye (Lond). 2015; 29:1251–59. https://doi.org/10.1038/eye.2015.124 [PubMed]
- 12. Waseem NH, Low S, Shah AZ, Avisetti D, Ostergaard P, Simpson M, Niemiec KA, Martin-Martin B, Aldehlawi H, Usman S, Lee PS, Khawaja AP, Ruddle JB, et al. Mutations in SPATA13/ASEF2 cause primary angle closure glaucoma. PLoS Genet. 2020; 16:e1008721. https://doi.org/10.1371/journal.pgen.1008721 [PubMed]
- 13. Zhang X, Liu Y, Wang W, Chen S, Li F, Huang W, Aung T, Wang N. Why does acute primary angle closure happen? Potential risk factors for acute primary angle closure. Surv Ophthalmol. 2017; 62:635–47. https://doi.org/10.1016/j.survophthal.2017.04.002 [PubMed]
- 14. Sun X, Dai Y, Chen Y, Yu DY, Cringle SJ, Chen J, Kong X, Wang X, Jiang C. Primary angle closure glaucoma: What we know and what we don't know. Prog Retin Eye Res. 2017; 57:26–45. https://doi.org/10.1016/j.preteyeres.2016.12.003 [PubMed]
- 15. Wu Y, Xia R, Dai C, Yan S, Xie T, Liu B, Gan L, Zhuang Z, Huang Q. Dexamethasone inhibits the proliferation of tumor cells. Cancer Manag Res. 2019; 11:1141–54. https://doi.org/10.2147/CMAR.S187659 [PubMed]
- 16. Ahram D, Grozdanic SD, Kecova H, Kuehn MH. Investigation of Gene-Expression Patterns in Familial Angle-Closure Glaucoma in the Basset Hound. Invest Ophthalmol Vis Sci. 2012; 53:4509. https://iovs.arvojournals.org/article.aspx?articleid=2357484.
- 17. Yu S, Meng S, Xiang M, Ma H. Phosphoenolpyruvate carboxykinase in cell metabolism: Roles and mechanisms beyond gluconeogenesis. Mol Metab. 2021; 53:101257. https://doi.org/10.1016/j.molmet.2021.101257 [PubMed]
- 18. Vincent EE, Sergushichev A, Griss T, Gingras MC, Samborska B, Ntimbane T, Coelho PP, Blagih J, Raissi TC, Choinière L, Bridon G, Loginicheva E, Flynn BR, et al. Mitochondrial Phosphoenolpyruvate Carboxykinase Regulates Metabolic Adaptation and Enables Glucose-Independent Tumor Growth. Mol Cell. 2015; 60:195–207. https://doi.org/10.1016/j.molcel.2015.08.013 [PubMed]
- 19. Nessler J, Hug P, Mandigers PJJ, Leegwater PAJ, Jagannathan V, Das AM, Rosati M, Matiasek K, Sewell AC, Kornberg M, Hoffmann M, Wolf P, Fischer A, et al. Mitochondrial PCK2 Missense Variant in Shetland Sheepdogs with Paroxysmal Exercise-Induced Dyskinesia (PED). Genes (Basel). 2020; 11:774. https://doi.org/10.3390/genes11070774 [PubMed]
- 20. Liu MX, Jin L, Sun SJ, Liu P, Feng X, Cheng ZL, Liu WR, Guan KL, Shi YH, Yuan HX, Xiong Y. Metabolic reprogramming by PCK1 promotes TCA cataplerosis, oxidative stress and apoptosis in liver cancer cells and suppresses hepatocellular carcinoma. Oncogene. 2018; 37:1637–53. https://doi.org/10.1038/s41388-017-0070-6 [PubMed]
- 21. Mejías-Peña Y, Estébanez B, Rodriguez-Miguelez P, Fernandez-Gonzalo R, Almar M, de Paz JA, González-Gallego J, Cuevas MJ. Impact of resistance training on the autophagy-inflammation-apoptosis crosstalk in elderly subjects. Aging (Albany NY). 2017; 9:408–18. https://doi.org/10.18632/aging.101167 [PubMed]
- 22. Lee EF, Fairlie WD. The Structural Biology of Bcl-xL. Int J Mol Sci. 2019; 20:2234. https://doi.org/10.3390/ijms20092234 [PubMed]
- 23. Venkatesh D, Stockwell BR, Prives C. p21 can be a barrier to ferroptosis independent of p53. Aging (Albany NY). 2020; 12:17800–14. https://doi.org/10.18632/aging.103961 [PubMed]
- 24. Lee YY, Choi YS, Kim DW, Cheong JY, Song KY, Ryu MS, Lim IK. Mitochondrial nucleoid remodeling and biogenesis are regulated by the p53-p21WAF1-PKCζ pathway in p16INK4a-silenced cells. Aging (Albany NY). 2020; 12:6700–32. https://doi.org/10.18632/aging.103029 [PubMed]
- 25. Razavipour SF, Harikumar KB, Slingerland JM. p27 as a Transcriptional Regulator: New Roles in Development and Cancer. Cancer Res. 2020; 80:3451–58. https://doi.org/10.1158/0008-5472.CAN-19-3663 [PubMed]
- 26. Liu X, Yao Z. Insensitivity of PI3K/Akt/GSK3 signaling in peripheral blood mononuclear cells of age-related macular degeneration patients. J Biomed Res. 2017; 31:248–55. https://doi.org/10.7555/JBR.31.20160096 [PubMed]
- 27. Sánchez-Cruz A, Martínez A, de la Rosa EJ, Hernández-Sánchez C. GSK-3 Inhibitors: From the Brain to the Retina and Back Again. Adv Exp Med Biol. 2019; 1185:437–41. https://doi.org/10.1007/978-3-030-27378-1_72 [PubMed]
- 28. Beurel E, Grieco SF, Jope RS. Glycogen synthase kinase-3 (GSK3): regulation, actions, and diseases. Pharmacol Ther. 2015; 148:114–31. https://doi.org/10.1016/j.pharmthera.2014.11.016 [PubMed]
- 29. Sánchez-Cruz A, Villarejo-Zori B, Marchena M, Zaldivar-Díez J, Palomo V, Gil C, Lizasoain I, de la Villa P, Martínez A, de la Rosa EJ, Hernández-Sánchez C. Modulation of GSK-3 provides cellular and functional neuroprotection in the rd10 mouse model of retinitis pigmentosa. Mol Neurodegener. 2018; 13:19. https://doi.org/10.1186/s13024-018-0251-y [PubMed]
- 30. Gobrecht P, Leibinger M, Andreadaki A, Fischer D. Sustained GSK3 activity markedly facilitates nerve regeneration. Nat Commun. 2014; 5:4561. https://doi.org/10.1038/ncomms5561 [PubMed]
- 31. Sampedro J, Bogdanov P, Ramos H, Solà-Adell C, Turch M, Valeri M, Simó-Servat O, Lagunas C, Simó R, Hernández C. New Insights into the Mechanisms of Action of Topical Administration of GLP-1 in an Experimental Model of Diabetic Retinopathy. J Clin Med. 2019; 8:339. https://doi.org/10.3390/jcm8030339 [PubMed]
- 32. Wei G, Xu X, Tong H, Wang X, Chen Y, Ding Y, Zhang S, Ju W, Fu C, Li Z, Zeng L, Xu K, Qiao J. Salidroside inhibits platelet function and thrombus formation through AKT/GSK3β signaling pathway. Aging (Albany NY). 2020; 12:8151–66. https://doi.org/10.18632/aging.103131 [PubMed]