



Introduction
Gastric cancer (GC) has been ranked as the second most common cause of cancer-associated mortality in Eastern Asia, particularly in China [1, 2]. Surgical resection is the most effective treatment strategy for patients with early-stage GC [3]. When GC progresses to an advanced stage (unresectable or metastatic), chemotherapy remains the major approach to treatment [4]. As drug resistance commonly occurs, it is urgent to identify novel biomarkers or targets for developing new therapies for GC.
Matrine is one of the bioactive components derived from Sophora flavescens [5]. Traditionally, matrine has been used for the treatment of cancer, arrhythmias, skin diseases, and hepatitis in China [6, 7]. Previous studies have shown that matrine can suppress the proliferation of tumor cells. For example, matrine was shown to inhibit melanoma and cervical cancer tumorigenesis [8, 9]. Notably, matrine was also found to induce GC cell death by regulating various downstream targets [10].
Long non-coding RNAs (lncRNAs) are defined as ~200-nt RNAs without protein translation ability [11, 12]. Previous studies have shown that lncRNAs are implicated in various biological processes, including tumorigenesis and drug resistance [13, 14]. Recently, it has been reported that NUT family member 2A-antisense RNA 1 (NUTM2A-AS1) is highly expressed in non-small cell lung cancer and plays a role in GC [15, 16].
MicroRNAs (miRNAs or miRs) are small non-coding RNAs of ~22 nt in length that bind to the 3′-untranslated region of mRNAs of target genes to regulate their expression [17]. It has been shown that miRNAs are involved in a variety of cancer types, including osteosarcoma and colorectal cancer [18, 19]. LncRNAs act as competitive endogenous RNAs to interact with miRNAs via miRNA recognition elements that sequester the miRNA-RNA-induced silencing complex away from downstream genes [20]. Consequently, lncRNAs might exert their effects via miRNAs during cancer progression. In previous studies, miR-613 was shown to suppress tumor cell proliferation and migration, and it was able to target SRY-box transcription factor 9, 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 2, and cyclin-dependent kinase 9 to inhibit the tumorigenic potential of GC cells [21–23].
Vascular endothelial growth factor A (VEGFA) plays a role in tumor growth and metastasis. In previous studies, VEGFA was shown to be regulated by various miRNAs including miR-150-5p [24], miR-130b [25], and miR-5047 [26]. Thus, it is common to link VEGFA and miRNAs in tumor progression.
The purpose of the present study was to elucidate the molecular mechanism underlying GC progression. We hope that the findings will broaden our current insights into lncRNA-mediated GC progression and facilitate the development of novel therapeutics for GC.
Results
The loss of NUTM2A-AS1 attenuates GC tumorigenesis following matrine treatment
To investigate the role of NUTM2A-AS1 in matrine-regulated GC tumorigenesis, NUTM2A-AS1 knockdown was performed by using two small hairpin (sh)RNAs. The reverse transcription-quantitative polymerase chain reaction (RT-qPCR) results demonstrated that both sh-NUTM2A-AS1-1 and sh-NUTM2A-AS1-2 decreased the expression of NUTM2A-AS1 compared with that caused by sh-negative control (NC) (Figure 1A).
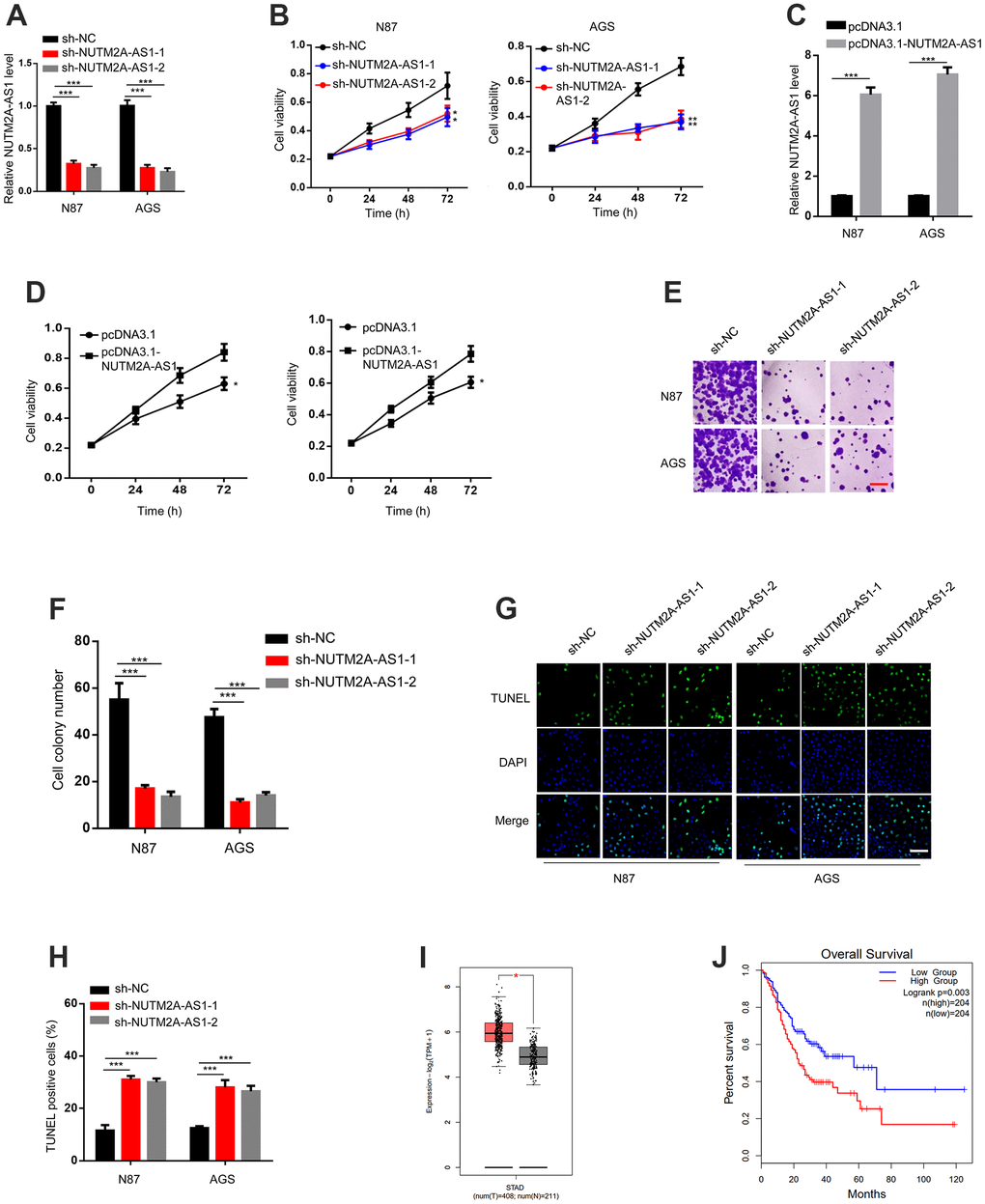
Figure 1. The loss of NUTM2A-AS1 attenuates GC tumorigenesis following matrine treatment. (A) The RT-qPCR was employed to evaluate the levels of NUTM2A-AS1 in N87 and AGS cells transfected with sh-NC, sh-NUTM2A-AS1-1, or sh-NUTM2A-AS1-2. ***P < 0.001. (B) The viability of N87 or AGS cells transfected with sh-NC, sh-NUTM2A-AS1-1, or sh-NUTM2A-AS1-2 following matrine treatment was examined by using the MTT assay. *P < 0.05, **P < 0.01. (C) The RT-qPCR was employed to evaluate the levels of NUTM2A-AS1 in N87 and AGS cells transfected with pcDNA3.1 or pcDNA3.1-NUTM2A-AS1. ***P < 0.001. (D) The viability of N87 or AGS cells transfected with pcDNA3.1 or pcDNA3.1-NUTM2A-AS1 following matrine treatment was examined by using the MTT assay. *P < 0.05. (E, F) Cell colonies of N87 and AGS cells transfected with sh-NC, sh-NUTM2A-AS1-1, or sh-NUTM2A-AS1-2 following matrine treatment. Scale bar, 5 μm. ***P<0.001. (G, H) Apoptosis of N87 and AGS cells transfected with sh-NC, sh-NUTM2A-AS1-1, or sh-NUTM2A-AS1-2 following matrine treatment was examined by using the TUNEL assay. Scale bar, 5 μm. ***P < 0.001. (I) The Cancer Genome Atlas analysis of patients with GC showing the NUTM2A-AS1 levels in normal tissues and tumors. *P < 0.05. (J) Kaplan-Meier plot to evaluate the overall survival of patients with GC and high or low NUTM2A-AS1 levels. P = 0.003, high (n = 204), low (n = 204). RT-qPCR, reverse transcription-quantitative polymerase chain reaction; NUTM2A, NUT family member 2A; AS1, antisense RNA 1; GC, gastric cancer; sh, small hairpin; NC, negative control.
NUTM2A-AS1-knockdown N87 and AGS cells showed reduced viability and cell colony numbers following matrine treatment (Figure 1B, 1E, 1F). In contrast, NUTM2A-AS1-overexpressing N87 and AGS cells showed enhanced viability following matrine treatment (Figure 1C, 1D). NUTM2A-AS1 knockdown led to increased cell apoptosis (Figure 1G, 1H). Taken together, these findings demonstrated that NUTM2A-AS1 contributed to the effect of matrine on GC tumorigenesis.
Next, we determined whether NUTM2A-AS1 was involved in the overall survival of patients with GC. By analyzing The Cancer Genome Atlas (TCGA) datasets, upregulated NUTM2A-AS1 expression was observed in tumor compared with non-tumor tissues (Figure 1I). Importantly, a low NUTM2A-AS1 level was associated with increased overall survival time (Figure 1J).
MiR-613 prevents NUTM2A-AS1-induced resistance of GC cells to matrine
To determine the downstream miRNAs responsible for NUTM2A-AS1-mediated matrine resistance in GC cells, starBase v2.0 was used to identify miR-613 as a potential target of NUTM2A-AS1 (Figure 2A). NUTM2A-AS1 knockdown resulted in increased expression of miR-613 (Figure 2B). 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and cell colony formation assays demonstrated that NUTM2A-AS1 depletion impaired N87 cell viability and proliferation, respectively, following treatment with matrine. The miR-613 inhibitor rescued the impaired cell viability and proliferation (Figure 2C–2E). NUTM2A-AS1 depletion caused enhanced apoptosis of N87 cells under matrine treatment, which was rescued by the miR-613 inhibitor (Figure 2F, 2G). These findings indicate that miR-613 was critical for NUTM2A-AS1-regulated resistance of GC cells to matrine.
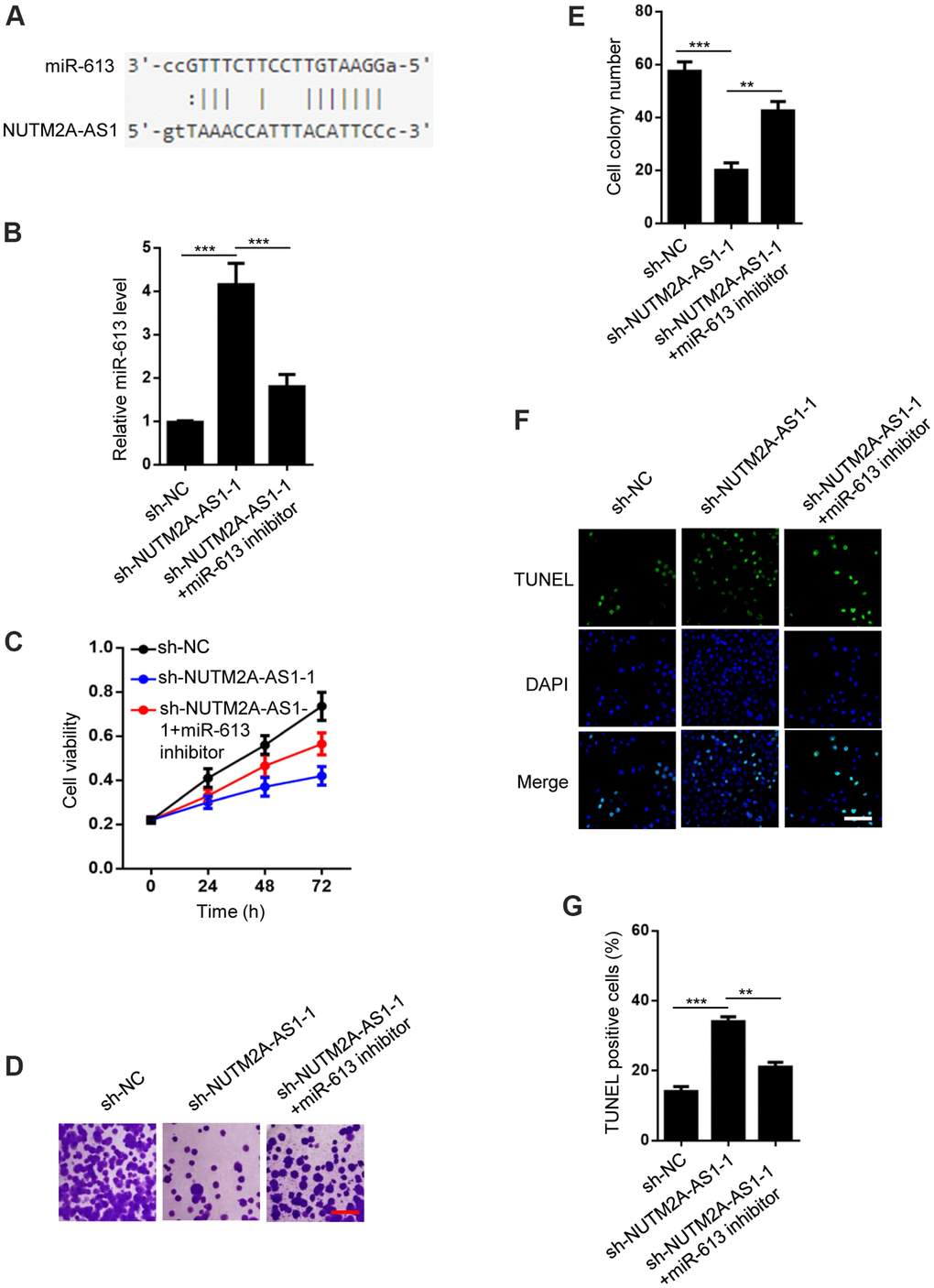
Figure 2. MiR-613 rescues NUTM2A-AS1-regulated matrine resistance in GC cells. (A) StarBase 2.0 shows that miR-613 is a target of NUTM2A-AS1. (B) The reverse transcription-quantitative polymerase chain reaction was employed to evaluate the levels of miR-613 in N87 cells transfected with sh-NC, sh-NUTM2A-AS1-1, or sh-NUTM2A-AS1-1 plus the miR-613 inhibitor. ***P < 0.001. (C) The viability of N87 cells transfected with sh-NC, sh-NUTM2A-AS1-1, or sh-NUTM2A-AS1-1 plus the miR-613 inhibitor under matrine treatment was examined by using the MTT assay. *P < 0.05, **P < 0.01. (D, E) Cell colonies of N87 cells transfected with sh-NC, sh-NUTM2A-AS1-1, or sh-NUTM2A-AS1-1 plus the miR-613 inhibitor under matrine treatment. Scale bar, 5 μm. **P < 0.01, ***P < 0.001. (F, G) Apoptosis of N87 cells transfected with sh-NC, sh-NUTM2A-AS1-1, or sh-NUTM2A-AS1-1 plus the miR-613 inhibitor under matrine treatment was examined by using the TUNEL assay. Scale bar, 5 μm. **P < 0.01, ***P < 0.001. DAPI, 4',6-diamidino-2-phenylindole; NUTM2A, NUT family member 2A; AS1, antisense RNA 1; GC, gastric cancer; miR, microRNA; sh, small hairpin; NC, negative control; TUNEL, terminal deoxynucleotidyl transferase dUTP nick-end labeling.
NUTM2A-AS1 and miR-613 regulate oxidative stress signaling
A previous report showed that miR-613 modulated reactive oxygen species (ROS) in neuronal cells [27]. Based on this observation, we hypothesized that NUTM2A-AS1 and miR-613 may regulate oxidative stress signaling. The miR-613 mimic promoted ROS generation, and increased the glutathione (GSH) level and superoxide dismutase (SOD) activity, while the miR-613 inhibitor decreased ROS generation, the GSH level, and SOD activity in N87 cells treated with matrine (Figure 3A–3C). In addition, NUTM2A-AS1 knockdown increased ROS generation, the GSH level, and SOD activity in N87 cells treated with matrine (Figure 3D–3F). Taken together, these findings demonstrated that NUTM2A-AS1 and miR-613 regulated oxidative stress signaling in GC cells.
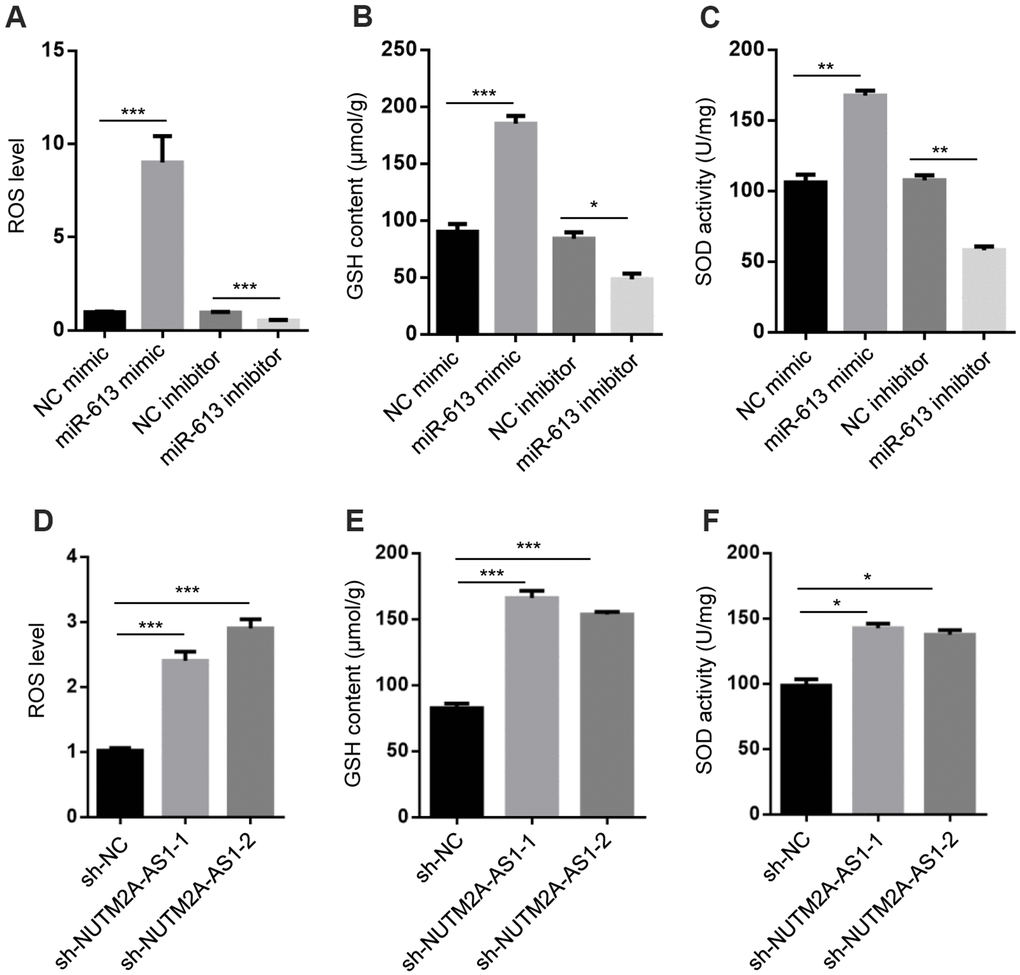
Figure 3. NUTM2A-AS1 and miR-613 regulate oxidative stress signaling. (A) ROS levels were measured in N87 cells transfected with the NC mimic, miR-613 mimic, NC inhibitor, or miR-613 inhibitor. ***P < 0.001. (B) GSH contents were measured in N87 cells transfected with the NC mimic, miR-613 mimic, NC inhibitor, or miR-613 inhibitor. *P < 0.05, ***P < 0.001. (C) SOD activity was measured in N87 cells transfected with the NC mimic, miR-613 mimic, NC inhibitor, or miR-613 inhibitor. **P < 0.01. (D) ROS levels were measured in N87 cells transfected with sh-NC, sh-NUTM2A-AS1-1, or sh-NUTM2A-AS1-2. ***P < 0.001. (E) GSH contents were measured in N87 cells transfected with sh-NC, sh-NUTM2A-AS1-1, or sh-NUTM2A-AS1-2. ***P < 0.001. (F) SOD activity was measured in N87 cells transfected with sh-NC, sh-NUTM2A-AS1-1, or sh-NUTM2A-AS1-2. *P < 0.05. NUTM2A, NUT family member 2A; AS1, antisense RNA 1; miR, microRNA; sh, small hairpin; NC, negative control; ROS, reactive oxygen species; GSH, glutathione; SOD, superoxide dismutase.
VEGFA is critical for miR-613-mediated suppression of GC in response to matrine treatment
TargetScan software was used to predict the potential downstream effector of NUTM2A-AS1/miR-613 in GC (Figure 4A). The RT-qPCR results revealed that the miR-613 mimic suppressed, whereas the miR-613 inhibitor promoted, VEGFA expression (Figure 4B). These data were in agreement with previous findings [19, 28]. In addition, NUTM2A-AS1 knockdown attenuated the VEGFA expression level (Figure 4C).
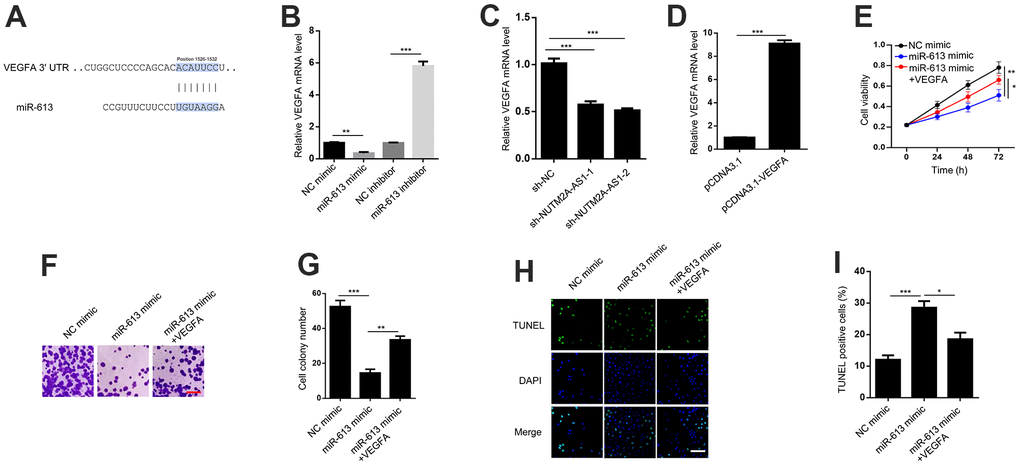
Figure 4. VEGFA is critical for miR-613-mediated suppression of gastric cancer under matrine treatment. (A) TargetScan shows that VEGFA is a potential target of miR-613. (B) VEGFA mRNA levels were detected by the RT-qPCR in N87 cells transfected with the NC mimic, miR-613 mimic, NC inhibitor, or miR-613 inhibitor. **P < 0.01, ***P < 0.001. (C) VEGFA mRNA levels were detected by the RT-qPCR in N87 cells transfected with sh-NC, sh-NUTM2A-AS1-1 or sh-NUTM2A-AS1-2. ***P < 0.001. (D) VEGFA expression levels were determined by the RT-qPCR in N87 cells transfected with pcDNA3.1 or pcDNA3.1-VEGFA. ***P < 0.001. (E) The MTT assay was used to evaluate the viability of N87 cells transfected with the NC mimic, miR-613 mimic, or miR-613 mimic plus VEGFA under matrine treatment. *P < 0.05, **P < 0.01. (F, G) Cell colonies of N87 cells transfected with the NC mimic, miR-613 mimic, or miR-613 mimic plus VEGFA under matrine treatment. Scale bar, 5 μm. **P < 0.01, ***P < 0.001. (H, I) The apoptosis of N87 cells transfected with the NC mimic, miR-613 mimic, or miR-613 mimic plus VEGFA under matrine treatment was examined by the TUNEL assay. Scale bar, 5 μm. *P < 0.05, ***P < 0.001. VEGFA, vascular endothelial growth factor A; miR, microRNA; RT-qPCR, reverse transcription-quantitative polymerase chain reaction; NC, negative control; TUNEL, terminal deoxynucleotidyl transferase dUTP nick-end labeling.
To elucidate whether VEGFA was critical for miR-613-modulated GC tumorigenesis, VEGFA was overexpressed in N87 cells (Figure 4D). The MTT and cell colony formation assays revealed that VEGFA could rescue miR-613-inhibited cell viability and proliferation following matrine treatment (Figure 4E–4G). In addition, VEGFA attenuated miR-613-enhanced cell apoptosis following matrine treatment (Figure 4H, 4I). In summary, VEGFA acted as key effector of miR-613-induced suppression of GC under matrine treatment.
Discussion
During GC treatment worldwide, chemotherapy resistance is a common problem [29]. Thus, novel agents, including natural compounds, must be developed for treating GC. Matrine is a component of traditional Chinese medicine. Previous studies showed that matrine inhibited GC and hepatocellular carcinoma progression by stimulating cell apoptosis [30, 31]. In agreement with published data, the present in vitro results demonstrated that matrine attenuated cell viability and proliferation, and enhanced cell apoptosis.
Non-coding RNAs include lncRNAs, miRNAs, circular RNAs, and small nucleolar RNAs, among others, and have been reported to be involved in diverse biological processes [32]. The present study explored the potential role of lncRNA NUTM2A-AS1 in GC tumorigenesis. NUTM2A-AS1 knockdown markedly reduced GC cell viability and proliferation, and promoted cell apoptosis. In patients with GC, tumors expressed more NUTM2A-AS1 than normal tissue, and a lower level of tumor NUTM2A-AS1 was associated with prolonged survival time.
ROS are generated naturally by cellular metabolism, as well as by some xenobiotics [33]. Intracellular ROS derived from chemotherapeutic agents can kill tumor cells [34]. It has been suggested that ROS could induce GC cell apoptosis and drug sensitivity [35–37]. In the present study, it was observed that NUTM2A-AS1 and miR-613 regulated ROS production in N87 cells. These findings led us to hypothesize that ROS may be a key effector for NUTM2A-AS1/miR-613-mediated matrine-resistant GC.
VEGF has been proposed to serve as a crucial gene for promoting angiogenesis during tumor metastasis [38]. VEGFA overexpression is found in various tumors, including GC [39]. The present study confirmed that VEGFA acted as a downstream effector of miR-613 in N87 cells. The mechanism by which VEGFA influences GC tumorigenesis remains unknown. According to previous research, VEGFA could activate the phosphoinositide 3-kinase/AKT and extracellular signal-regulated kinase 1/2 signaling pathways, which may be potential mechanisms for NUTM2A-AS1/VEGFA-induced GC tumorigenesis [40]. In addition, it is interesting that oxidative stress can regulate VEGFA gene transcription [41]. This observation led us to perform the current study that clarifies the relationship between NUTM2A-AS1, miR-613, ROS, and VEGFA.
Conclusions
The NUTM2A-AS1/miR-613/ROS/VEGFA axis is important for the inhibition of GC progression by matrine. The present findings enrich our understanding of GC treatment with matrine, and may facilitate the development of novel therapeutics for GC by targeting the NUTM2A-AS1/miR-613/ROS/VEGFA axis. However, the role of this axis in GC presented in this study was limited to the cellular and molecular biology levels. Thus, additional efforts are needed to confirm this finding at the animal level and, ultimately, in human clinical trials.
Materials and Methods
Cell culture
Human GC cells (N87 and AGS) and 293T cells were obtained from the The Cell Bank of Type Culture Collection of The Chinese Academy of Sciences (Shanghai, China). Cells were cultured in Dulbecco’s Modified Eagle’s Medium (Gibco, Gaithersburg, MD, USA) supplemented with 10% fetal bovine serum (Gibco) and 1% penicillin/streptomycin at 37° C in the presence of 5% CO2.
Transfection
VEGFA and NUTM2A-AS1 were amplified and cloned into the pcDNA3.1 vector. pcDNA3.1-VEGFA or pcDNA3.1-NUTM2A-AS1 was then transiently transfected into N87 or AGS cells using Lipofectamine® 2000 (Invitrogen). After 48 h of transfection, the cell lysates were collected and used for subsequent experiments.
Stable cell line generation
To generate stable cell lines, ~2 μg of the vectors [1 μg pLKO.1-shRNA, 0.5 μg pVSVG, and 0.5 μg of pPAX2] were transfected into 293T cells by using Lipofectamine® 2000. After 36 h, the lentiviruses were harvested and stored at -80° C. Next, the viruses were used to infect N87 and AGS cells. To obtain resistant cells, ~1 μg/mL puromycin was used. The shRNA sequences were as follows: sh-NC, 5′-AUCGGCAACUAGGCAUCAUCAG-3′; sh-NUTM2A-AS1-1, 5′-GGGACAGUGUAUGCAAGAA-3′; and sh-NUTM2A-AS1-2, 5′-GGACAGUGUAUGCAAGAAU-3′.
MTT assay
N87 and AGS cells were seeded in 96-well plates (~5,000 cells/well). MTT reagent (Sigma-Aldrich, St. Louis, MO, USA) was added and the cells were incubated for 4 h at 37° C in the presence of 5% CO2. At different time points (0, 24, 48, and 72 h), 150 μL dimethyl sulfoxide were added to each well and incubated with the cells for 10 min. Finally, the optical density at 490 nm was examined.
Colony formation assay
N87 and AGS cells (2 × 104 cells) were seeded in 6-well plates and cultured for 2 weeks. The cell colonies were fixed with 4% paraformaldehyde. Crystal violet (0.005%) was used to stain the colonies, and the number was counted under a light microscope.
Terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) assay
Transfected N87 and AGS cells were seeded in each well of a 24-well plate. TUNEL signals were detected according to the manufacturer’s instructions (Beyotime Institute of Biotechnology, Haimen, China). The cell nuclei were stained with 4',6-diamidino-2-phenylindole.
TCGA analysis
TCGA datasets of patients with GC were downloaded and analyzed (408 tumor and 211 normal tissues). The relative expression of NUTM2A-AS1, overall survival, and stage were analyzed. A Kaplan-Meier plot was used to evaluate the overall survival of patients.
ROS determination
GC cells were incubated with 10 μM 2,7-dichlorodi-hydrofluorescin diacetate at 37° C for 30 min to determine ROS generation. The activity of SOD and content of GSH were measured with cell lysates using appropriate kits (Construction Bioengineering Research, Nanjing, China).
RT-qPCR
Total RNA was extracted from GC cells using TRIzol® (Invitrogen) according to the manufacturer’s instructions. The RNA concentration was determined using a NanoDrop instrument (NanoDrop Technologies, Wilmington, DE, USA). For RT-qPCR, a PrimeScript RT Reagent Kit (Takara Bio, Dailan, China) was used to generate cDNA with 1 μg total RNA. RT-qPCR was performed with the SYBR Green I Master Mix Kit. β-Actin or U6 acted as an internal control. Relative gene levels were calculated by using the 2-ΔΔCq method. The primers used were as follows: NUTM2A-AS1: forward, 5′-CTCGACTCAGTCCTCCAGC-3′ and reverse, 5′-GCCTCCTCCTCTTGCTTCAT-3′; miR-613: forward, 5′-GTGAGTGCGTTTCCAAGTGT-3′ and reverse, 5′-GGGTCCCTTCACACTTGGAA-3′; VEGFA: forward, 5′-ATCCAATCGAGACCCTGGTG-3′ and reverse, 5′-ATCTCTCCTATGTGCTGGCC-3′; β-actin: forward, 5′-GGGAAATCGTGCGTGACATT-3′ and reverse, 5′-AGGTAGTTTCGTGGATGCCA-3′; and U6: forward, 5′- CGCTTCGGCAGCACATATACTA-3′ and reverse, 5′- GAATTTGCGTGTCATCCTTGCG-3′.
Statistical analyses
Statistical analyses were performed with Prism 8.0 software (GraphPad Software, La Jolla, CA, USA). Two groups were compared by using the unpaired Student’s t-test, while multiple groups were compared by a one-way analysis of variance followed by Tukey’s post-hoc test. P < 0.05 was considered to indicate a statistically significant difference.
Data availability statement
The data supporting this research are available from the corresponding author upon reasonable request.
Author Contributions
LZ and QL designed the study; HY and YJ performed the experiments, analyzed the data and wrote the manuscript; YG, QL, MR, WZ and CY provided essential assistant through this work; and all authors reviewed the final manuscript.
Acknowledgments
We would like to thank EdiTar Bio-Tech Ltd. (Nanking, China) for English language editing.
Conflicts of Interest
We declare there are no conflicts of interest.
Funding
This work was supported by the Youth Fund Project of NSFC (No. 81403242) and Scientific Research Project of Traditional Chinese Medicine of Shanghai Health Commission (No. 2020LZ007).
References
- 1. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015; 65:87–108. https://doi.org/10.3322/caac.21262 [PubMed]
- 2. Chen W, Zheng R, Baade PD, Zhang S, Zeng H, Bray F, Jemal A, Yu XQ, He J. Cancer statistics in China, 2015. CA Cancer J Clin. 2016; 66:115–32. https://doi.org/10.3322/caac.21338 [PubMed]
- 3. Zhang J, Li Y, Chen X, Liu T, Chen Y, He W, Zhang Q, Liu S. Autophagy is involved in anticancer effects of matrine on SGC-7901 human gastric cancer cells. Oncol Rep. 2011; 26:115–24. https://doi.org/10.3892/or.2011.1277 [PubMed]
- 4. Li Y, Zhang J, Ma H, Chen X, Liu T, Jiao Z, He W, Wang F, Liu X, Zeng X. Protective role of autophagy in matrine-induced gastric cancer cell death. Int J Oncol. 2013; 42:1417–26. https://doi.org/10.3892/ijo.2013.1817 [PubMed]
- 5. Zhang JW, Su K, Shi WT, Wang Y, Hu PC, Wang Y, Wei L, Xiang J, Yang F. Matrine inhibits the adhesion and migration of BCG823 gastric cancer cells by affecting the structure and function of the vasodilator-stimulated phosphoprotein (VASP). Acta Pharmacol Sin. 2013; 34:1084–92. https://doi.org/10.1038/aps.2013.15 [PubMed]
- 6. Zhang MJ, Huang J. [Recent research progress of anti-tumor mechnism matrine]. Zhongguo Zhong Yao Za Zhi. 2004; 29:115–8. [PubMed]
- 7. Liu Y, Xu Y, Ji W, Li X, Sun B, Gao Q, Su C. Anti-tumor activities of matrine and oxymatrine: literature review. Tumour Biol. 2014; 35:5111–9. https://doi.org/10.1007/s13277-014-1680-z [PubMed]
- 8. Liu XY, Fang H, Yang ZG, Wang XY, Ruan LM, Fang DR, Ding YG, Wang YN, Zhang Y, Jiang XL, Chen HC. Matrine inhibits invasiveness and metastasis of human malignant melanoma cell line A375 in vitro. Int J Dermatol. 2008; 47:448–56. https://doi.org/10.1111/j.1365-4632.2008.03627.x [PubMed]
- 9. Zhang L, Wang T, Wen X, Wei Y, Peng X, Li H, Wei L. Effect of matrine on HeLa cell adhesion and migration. Eur J Pharmacol. 2007; 563:69–76. https://doi.org/10.1016/j.ejphar.2007.01.073 [PubMed]
- 10. Liu ZM, Yang XL, Jiang F, Pan YC, Zhang L. Matrine involves in the progression of gastric cancer through inhibiting miR-93-5p and upregulating the expression of target gene AHNAK. J Cell Biochem. 2020; 121:2467–77. https://doi.org/10.1002/jcb.29469 [PubMed]
- 11. Derrien T, Johnson R, Bussotti G, Tanzer A, Djebali S, Tilgner H, Guernec G, Martin D, Merkel A, Knowles DG, Lagarde J, Veeravalli L, Ruan X, et al. The GENCODE v7 catalog of human long noncoding RNAs: analysis of their gene structure, evolution, and expression. Genome Res. 2012; 22:1775–89. https://doi.org/10.1101/gr.132159.111 [PubMed]
- 12. Guttman M, Amit I, Garber M, French C, Lin MF, Feldser D, Huarte M, Zuk O, Carey BW, Cassady JP, Cabili MN, Jaenisch R, Mikkelsen TS, et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature. 2009; 458:223–7. https://doi.org/10.1038/nature07672 [PubMed]
- 13. Ulitsky I, Bartel DP. lincRNAs: genomics, evolution, and mechanisms. Cell. 2013; 154:26–46. https://doi.org/10.1016/j.cell.2013.06.020 [PubMed]
- 14. Yang G, Lu X, Yuan L. LncRNA: a link between RNA and cancer. Biochim Biophys Acta. 2014; 1839:1097–109. https://doi.org/10.1016/j.bbagrm.2014.08.012 [PubMed]
- 15. Acha-Sagredo A, Uko B, Pantazi P, Bediaga NG, Moschandrea C, Rainbow L, Marcus MW, Davies MP, Field JK, Liloglou T. Long non-coding RNA dysregulation is a frequent event in non-small cell lung carcinoma pathogenesis. Br J Cancer. 2020; 122:1050–8. https://doi.org/10.1038/s41416-020-0742-9 [PubMed]
- 16. Wang J, Yu Z, Wang J, Shen Y, Qiu J, Zhuang Z. LncRNA NUTM2A-AS1 positively modulates TET1 and HIF-1A to enhance gastric cancer tumorigenesis and drug resistance by sponging miR-376a. Cancer Med. 2020; 9:9499–510. https://doi.org/10.1002/cam4.3544 [PubMed]
- 17. Sun K, Lai EC. Adult-specific functions of animal microRNAs. Nat Rev Genet. 2013; 14:535–48. https://doi.org/10.1038/nrg3471 [PubMed]
- 18. Zhu Y, Tang L, Zhao S, Sun B, Cheng L, Tang Y, Luo Z, Lin Z, Zhu J, Zhu W, Zhao R, Lu B, Long H. CXCR4-mediated osteosarcoma growth and pulmonary metastasis is suppressed by MicroRNA-613. Cancer Sci. 2018; 109:2412–22. https://doi.org/10.1111/cas.13653 [PubMed]
- 19. Yang X, Zhang L, Song X, He W, Zhang D, Lu Q, Wu J, Wu C, Jiang J. MicroRNA-613 promotes colon cancer cell proliferation, invasion and migration by targeting ATOH1. Biochem Biophys Res Commun. 2018; 504:827–33. https://doi.org/10.1016/j.bbrc.2018.09.054 [PubMed]
- 20. Thomson DW, Dinger ME. Endogenous microRNA sponges: evidence and controversy. Nat Rev Genet. 2016; 17:272–83. https://doi.org/10.1038/nrg.2016.20 [PubMed]
- 21. Liu H, Chen K, Wang L, Zeng X, Huang Z, Li M, Dong P, Chen X. miR-613 inhibits Warburg effect in gastric cancer by targeting PFKFB2. Biochem Biophys Res Commun. 2019; 515:37–43. https://doi.org/10.1016/j.bbrc.2019.05.001 [PubMed]
- 22. Lu Y, Tang L, Zhang Q, Zhang Z, Wei W. MicroRNA-613 inhibits the progression of gastric cancer by targeting CDK9. Artif Cells Nanomed Biotechnol. 2018; 46:980–4. https://doi.org/10.1080/21691401.2017.1351983 [PubMed]
- 23. Xue M, Li G, Sun P, Zhang D, Fang X, Li W. MicroRNA-613 induces the sensitivity of gastric cancer cells to cisplatin through targeting SOX9 expression. Am J Transl Res. 2019; 11:885–94. [PubMed]
- 24. Chen X, Xu X, Pan B, Zeng K, Xu M, Liu X, He B, Pan Y, Sun H, Wang S. miR-150-5p suppresses tumor progression by targeting VEGFA in colorectal cancer. Aging (Albany NY). 2018; 10:3421–37. https://doi.org/10.18632/aging.101656 [PubMed]
- 25. Mu HQ, He YH, Wang SB, Yang S, Wang YJ, Nan CJ, Bao YF, Xie QP, Chen YH. MiR-130b/TNF-α/NF-κB/VEGFA loop inhibits prostate cancer angiogenesis. Clin Transl Oncol. 2020; 22:111–21. https://doi.org/10.1007/s12094-019-02217-5 [PubMed]
- 26. Guo J, Chen M, Ai G, Mao W, Li H, Zhou J. Hsa_circ_0023404 enhances cervical cancer metastasis and chemoresistance through VEGFA and autophagy signaling by sponging miR-5047. Biomed Pharmacother. 2019; 115:108957. https://doi.org/10.1016/j.biopha.2019.108957 [PubMed]
- 27. Di G, Wang Z, Wang W, Cheng F, Liu H. AntagomiR-613 protects neuronal cells from oxygen glucose deprivation/re-oxygenation via increasing SphK2 expression. Biochem Biophys Res Commun. 2017; 493:188–94. https://doi.org/10.1016/j.bbrc.2017.09.049 [PubMed]
- 28. Yu X, Wang W. Tumor suppressor microRNA-613 inhibits glioma cell proliferation, invasion and angiogenesis by targeting vascular endothelial growth factor A. Mol Med Rep. 2017; 16:6729–35. https://doi.org/10.3892/mmr.2017.7422 [PubMed]
- 29. Gravalos C, Jimeno A. HER2 in gastric cancer: a new prognostic factor and a novel therapeutic target. Ann Oncol. 2008; 19:1523–9. https://doi.org/10.1093/annonc/mdn169 [PubMed]
- 30. Luo C, Zhu Y, Jiang T, Lu X, Zhang W, Jing Q, Li J, Pang L, Chen K, Qiu F, Yu X, Yang J, Huang J. Matrine induced gastric cancer MKN45 cells apoptosis via increasing pro-apoptotic molecules of Bcl-2 family. Toxicology. 2007; 229:245–52. https://doi.org/10.1016/j.tox.2006.10.020 [PubMed]
- 31. Zhang JQ, Li YM, Liu T, He WT, Chen YT, Chen XH, Li X, Zhou WC, Yi JF, Ren ZJ. Antitumor effect of matrine in human hepatoma G2 cells by inducing apoptosis and autophagy. World J Gastroenterol. 2010; 16:4281–90. https://doi.org/10.3748/wjg.v16.i34.4281 [PubMed]
- 32. Matsui M, Corey DR. Non-coding RNAs as drug targets. Nat Rev Drug Discov. 2017; 16:167–79. https://doi.org/10.1038/nrd.2016.117 [PubMed]
- 33. Devasagayam TP, Tilak JC, Boloor KK, Sane KS, Ghaskadbi SS, Lele RD. Free radicals and antioxidants in human health: current status and future prospects. J Assoc Physicians India. 2004; 52:794–804. [PubMed]
- 34. Renschler MF. The emerging role of reactive oxygen species in cancer therapy. Eur J Cancer. 2004; 40:1934–40. https://doi.org/10.1016/j.ejca.2004.02.031 [PubMed]
- 35. Ba MC, Long H, Cui SZ, Gong YF, Yan ZF, Wang S, Wu YB. Mild hyperthermia enhances sensitivity of gastric cancer cells to chemotherapy through reactive oxygen species-induced autophagic death. Tumour Biol. 2017; 39:1010428317711952. https://doi.org/10.1177/1010428317711952 [PubMed]
- 36. Liang W, Cai A, Chen G, Xi H, Wu X, Cui J, Zhang K, Zhao X, Yu J, Wei B, Chen L. Shikonin induces mitochondria-mediated apoptosis and enhances chemotherapeutic sensitivity of gastric cancer through reactive oxygen species. Sci Rep. 2016; 6:38267. https://doi.org/10.1038/srep38267 [PubMed]
- 37. Wang S, Chen Z, Zhu S, Lu H, Peng D, Soutto M, Naz H, Peek R
Jr , Xu H, Zaika A, Xu Z, El-Rifai W. PRDX2 protects against oxidative stress induced by H. pylori and promotes resistance to cisplatin in gastric cancer. Redox Biol. 2020; 28:101319. https://doi.org/10.1016/j.redox.2019.101319 [PubMed] - 38. Fu D, Li P, Cheng W, Tian F, Xu X, Yi X, Tang C, Wang Y, Hu Q, Zhang Z. Impact of vascular endothelial growth factor gene-gene and gene-smoking interaction and haplotype combination on bladder cancer risk in Chinese population. Oncotarget. 2017; 8:22927–35. https://doi.org/10.18632/oncotarget.15287 [PubMed]
- 39. Ping SY, Shen KH, Yu DS. Epigenetic regulation of vascular endothelial growth factor a dynamic expression in transitional cell carcinoma. Mol Carcinog. 2013; 52:568–79. https://doi.org/10.1002/mc.21892 [PubMed]
- 40. An G, Liang S, Sheng C, Liu Y, Yao W. Upregulation of microRNA-205 suppresses vascular endothelial growth factor expression-mediated PI3K/Akt signaling transduction in human keloid fibroblasts. Exp Biol Med (Maywood). 2017; 242:275–85. https://doi.org/10.1177/1535370216669839 [PubMed]
- 41. Schäfer G, Cramer T, Suske G, Kemmner W, Wiedenmann B, Höcker M. Oxidative stress regulates vascular endothelial growth factor-A gene transcription through Sp1- and Sp3-dependent activation of two proximal GC-rich promoter elements. J Biol Chem. 2003; 278:8190–8. https://doi.org/10.1074/jbc.M211999200 [PubMed]