



Introduction
ZnF (zinc finger) proteins are the most abundant proteins in eukaryotic genomes [1], which are categorized into several types, namely C2H2, C2HC, C2HC5, C2C2, CCCH, C3HC4, C4, C4HC3, C6 and C8 based on the position as well as the number of histidine and cysteine residues [2, 3]. The functions of ZnF proteins are extraordinarily diverse, which have roles in transcriptional activation, protein degradation, DNA repair, cell migration and cell apoptosis [4, 5]. Most ZnF proteins bind to cognate DNA in the nuclei to regulate the expression of target genes [6]. In addition to DNA, many ZnF proteins also bind to proteins or RNA, which fold into a mini domain around a central zinc ion and ‘grab’ the DNA/RNA through the surrounding ZnFs [7].
Zbtb34 (zinc finger and BTB domain containing 34) is a novel zinc finger protein that was first reported by Qi et al. in 2006 [8], who found that Zbtb34 was expressed ubiquitously in most adult human tissues and was mainly localized to the nuclei. Over the next ten years, there had been few studies on Zbtb34. It was found in a previous study that reactive oxygen species-induced anticancer agents could kill cancer cells by downregulating the miR-27a or miR-17/miR-20a [9], which could regulate the expression of Zbtb34, resulting in the suppression of the expression of specificity protein (Sp) transcription factors [10]. This result indicates that Zbtb34 is a transcriptional repressor. It was found in a recent study that Zbtb34 and lnc-CPLC strongly correlated in colorectal cancer progression, indicating that lnc-CPLC/miR-4319/Zbtb34 signal axis played a role in regulating the development of colorectal cancer [11].
Mouse embryonic stem cells (mESCs) are considered to be a unique tool for many researchers since they have the ability to self-renew and differentiate into all cell types [12]. mESCs derived from knock-in/knockout blastocysts are very useful for studying mutant mice, especially when the phenotype is embryonic-lethal at early stages of development [13]. In the present study, we found that Zbtb34 promoted the proliferation of embryonic stem cells and elongated telomere length by competing with Pot1 (protection of telomere 1) b for the telomere binding sites. Since the deletion of both Pot1a and Pot1b resulted in early embryonic lethality [14], the biological functions of Zbtb34 are investigated through various experiments on mESCs.
Results
Zbtb34 induces the proliferation of mESCs
MTT assay demonstrated that the upregulation of Zbtb34 induced the proliferation of mESCs (Figure 1). However, the rate of apoptotic cells was not significantly different between the upregulated- and downregulated-Zbtb34 mESCs through a flow cytometry analysis.
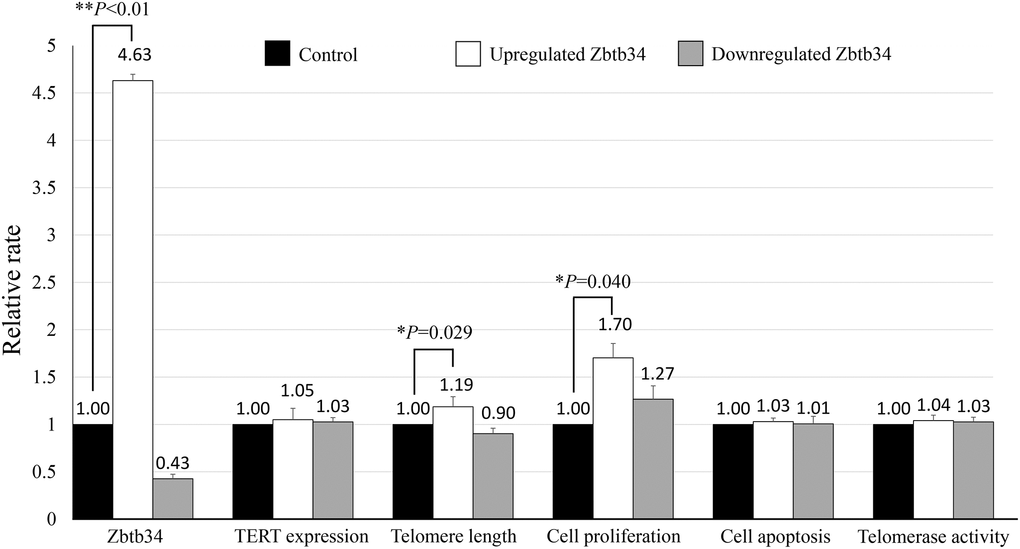
Figure 1. Zbtb34 elongates telomere length and increases cell proliferation. The mESCs were transfected with a Zbtb34-EGFP vector or Zbtb34-siRNA. The values of Zbtb34, TERT expression, telomere length, cell proliferation, cell apoptosis and telomerase activity are expressed as fold changes compared with control values normalized to 1. *P < 0.05, **P < 0.01. Abbreviations: mESCs: mouse embryonic stem cells; TERT: telomerase reverse transcriptase.
Zbtb34 elongates telomere length
The telomere length was detected in mESC lines transfected with Zbtb34-EGFP or siZbtb34. The result revealed that the upregulation of Zbtb34 significantly elongated the telomere length (Figure 1), which, interestingly, did not affect the expression of TERT (telomerase reverse transcriptase), indicating that Zbtb34 did not regulate the TERT expression. Moreover, there was no difference in telomerase activity between upregulated- and downregulated-Zbtb34 mESCs (Figure 1).
Zbtb34 has 3 zinc finger motifs
Mouse Zbtb34 was predicted to have 3 C2H2 ZnFs (Figure 2A). Human Zbtb34 also contains 3 ZnFs, but its location is different from that of the mouse (Figure 2B). Since our research group is working on mESCs, in this study, we aim to clarify the functions of mouse Zbtb34. The alignment of the amino acid sequences of Mus musculus, Homo sapiens, Rattus norvegicus, Macaca mulatta, Felis catus, Panthera tigris, Vicugna pacos and Castor canadensis shows that the amino acids within ZnFs are highly conserved among different species (Figure 2C).
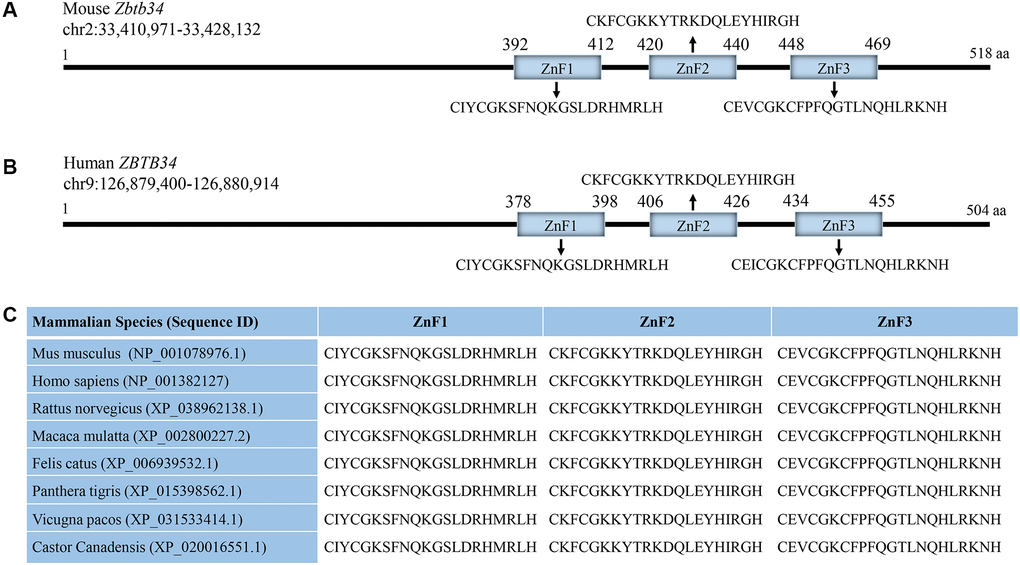
Figure 2. Zbtb34 contains 3 highly-conserved ZnFs. (A) Mouse Zbtb34 is located on Chromosome 2. (B) Human Zbtb34 is located on Chromosome 9. (C) The alignment of the amino acid sequences within ZnFs is highly conserved among different mammalian species. The number of parentheses is the accession number. Abbreviation: ZnF: zinc finger.
The ZnF3 is necessary for Zbtb34 binding to telomeres
C57BL/6 mESCs cultured in OriCell C57BL/6 × 129 medium (Figure 3A) were positive for the pluripotency markers NANOG (red), OCT4 (green), and SSEA1 (red) (Figure 3B). Using the telomere ChIP, we found that Zbtb34 could bind to the telomere DNA sequences (repeat TTAGGG) (Figure 4A). The co-location of Zbtb34 and telomeres was confirmed through the FISH (Fluorescence in situ hybridization) experiment (Figure 4B). We further investigated the functions of each zinc finger motif. A series of alanine scan mutants were generated to substitute the amino acids within each ZnF. The absence of ZnF2/ZnF3 and ZnF1/ZnF3 abolishes the Zbtb34 binding to telomeres (Figure 4A). However, the absence of ZnF1/ZnF2 (Figure 4A) did not affect the binding between Zbtb34 and telomeres, which suggests that ZnF3 at the C-terminus of amino acid protein 448 to 469 is necessary for Zbtb34 binding to the telomeres.
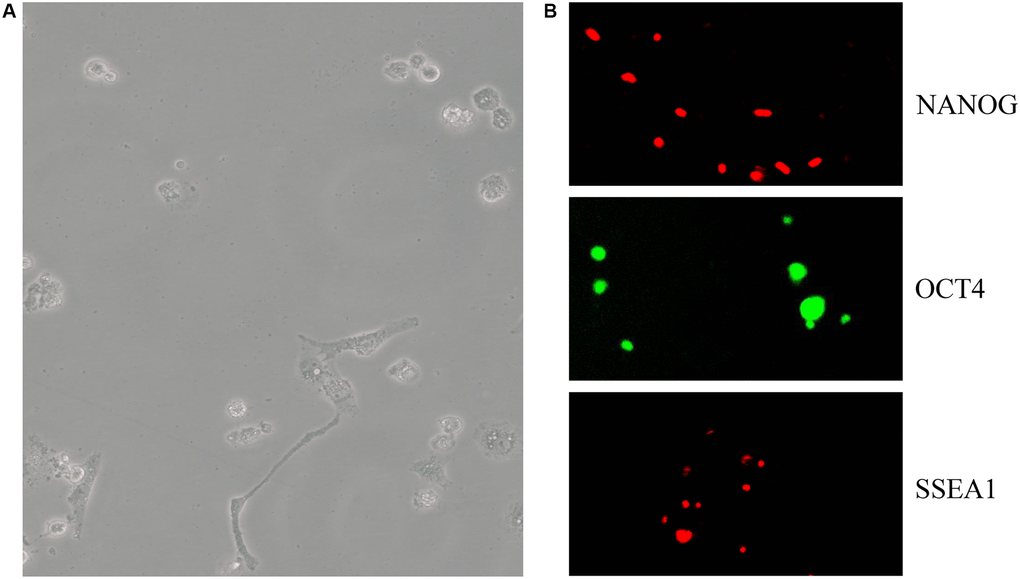
Figure 3. mESCs express pluripotency markers. (A) C57BL/6 mESC culture in OriCell C57BL/6 × 129 medium. (B) The pluripotency marker NANOG (red), OCT4 (green), SSEA1 (red) and staining was visualized through fluorescent microscopy. Abbreviation: mESCs: mouse embryonic stem cells.
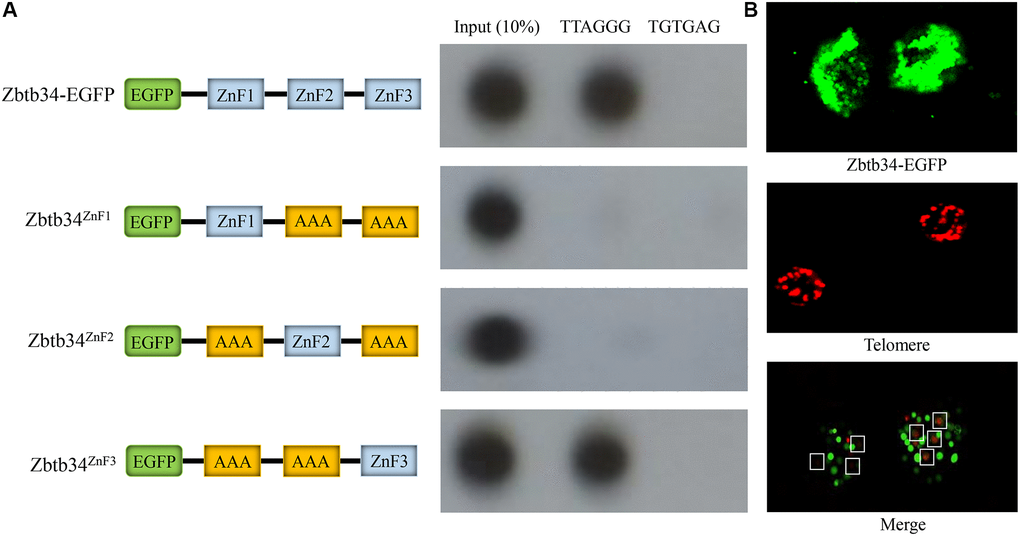
Figure 4. Zbtb34 binds to the telomere DNA. (A) The ZnFs within Zbtb34 of mESCs are analyzed through telomere co-immunoprecipitation. Schemata represents the construct of the Zbtb34-EGFP vector and the alanine substitution of ZnF1, ZnF2 or ZnF3. The results show that Zbtb34 is co-immunoprecipitated with the sequence (TTAGGG) of the telomeres. The association of ZnF1, ZnF2 or ZnF3 with telomere DNA is further analyzed. The results reveal that ZnF3 is co-immunoprecipitated with the telomere DNA. The random sequence (TGTGAG) is the control. (B) Fluorescence in situ hybridization shows the localization of Zbtb34 (green) and telomeres (red). The dots in the white boxes indicate the merger of green and red dots. Abbreviations: ZnF: zinc finger; mESCs: mouse embryonic stem cells.
Zbtb34 competes for the binding sites with Pot1b
Since Pot1 was the only protein binding to the single-stranded DNA TTAGGG at the end of the telomeres, we investigated the relationship between Zbtb34 and Pot1a or Pot1b, two highly homologous isoforms in mice. The results revealed that the upregulation of Zbtb34 decreased the binding of Pot1b to telomeres, which also reduced the binding of Zbtb34 to the telomere DNA (Figure 5). This result indicates that Zbtb34 and Pot1b have the same binding site at the end of the telomeres and a competitive relationship in telomere binding. Interestingly, the upregulation of Zbtb34 does not affect the binding between Pot1a and telomeres.
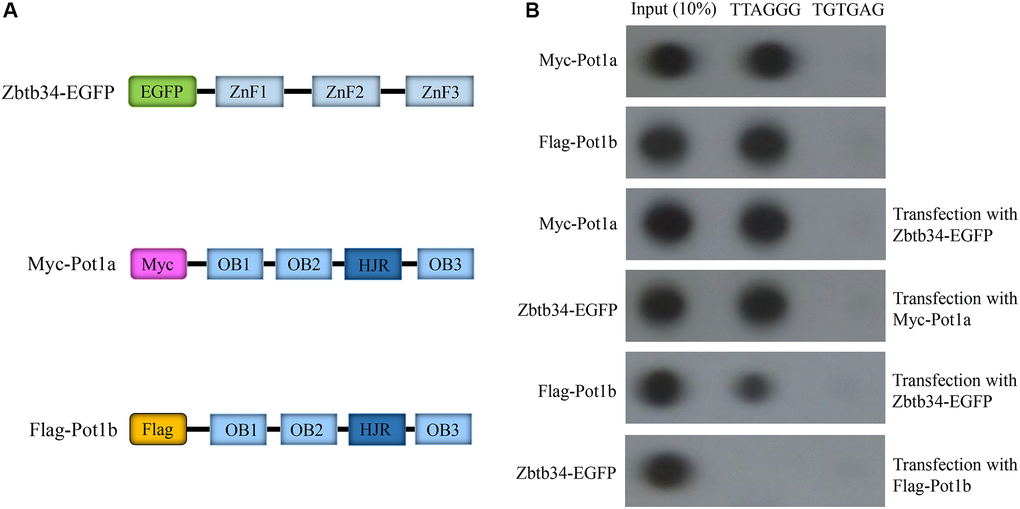
Figure 5. Zbtb34 competes for the binding sites on telomeres with Pot1b. (A) Schemata represents the Myc-Pot1a and Flag-Pot1b constructs. (B) Telomere co-immunoprecipitation shows the association of Zbtb34 and Pot1a or Pot1b with telomere DNA in mESCs. The random sequence (TGTGAG) is the control. Abbreviations: ZnF: zinc finger; OB: oligonucleotide binding; HJR: holiday junction resolvase; mESCs: mouse embryonic stem cells.
Discussion
To date, the relationship between ZnF proteins and telomeres remains largely unknown [15]. Our study demonstrated that Zbtb34 could bind to telomere DNA directly. Since the alanine substitution of ZnF3 inhibited the binding, it suggested that the presence of ZnF3 was necessary for Zbtb34 to bind to telomere DNA. Based on its localization at the telomeres, we reasoned that Zbtb34 might play a role in telomere length regulation. The result confirmed our hypothesis that the upregulation of Zbtb34 promoted telomere elongation. In recent years, the association between ZnF proteins and telomeres has been pointed toward in more and more studies. The most prominent example is the ZBTB48, which can bind to long telomeres through its ZnF11, competing with the telomeric-repeat binding factors TRF1 and TRF2, setting an upper limit for telomere length through triggering telomere trimming [16]. ZBTB10 is another reported protein that can bind to telomeres, which binds to the telomeres with its two C2H2 zinc fingers and interacts with the N-terminal domain of TRF2 [17]. However, ZBTB10 is not involved in telomere homeostasis, indicating that the functions of ZBTB10 binding to telomeres need to be further studied. An interesting finding in this study is that only the third ZnF of Zbtb34 binds to the telomeres, while the functions of the other two ZnFs are unclear. It was found in previous studies that ZBTB48 and ZBTB10 were involved in the progression of different cancers [18, 19]. Zbtb34 has also been shown to play a role in tumor growth as a transcriptional repressor [20]. Therefore, we believe that the other two ZnFs in Zbtb34 are involved in transcriptional repression of tumor growth.
Telomeres consist of DNA repeating sequences (TTAGGG) and shelterin complex at the end of chromosomes, protecting the chromosomes from fusion or degradation [21, 22]. This shelterin complex consists of six telomere-specific proteins, namely TRF1, TRF2, TIN2, POT1, TPP1 and RAP1 (Figure 6) [23]. TIN2 plays a pivotal role in suppressing ATM and ATR signaling pathways through interacting with TPP1/POT1 heterodimer as well as in stabilizing TRF1 and TRF2 at the telomeric DNA [24]. TRF2/RAP1 heterodimer prevent telomere fragility [25]. Among these shelterin proteins, only Pot1 directly recognizes single TTAGGG repeats at the 3′ telomere terminus [26]. Mammalian shelterin proteins Pot1 and TPP1 form a stable heterodimer regulating telomerase-mediated telomere extension [27]. Pot1 plays a negative role in preventing telomerase recruitment to telomeres, resulting in telomere shortening [28]. In mice, Pot1 has two highly homologous isoforms, namely Pot1a and Pot1b. However, the functions of these two isoforms are ill-defined because of the lack of inhibitors that can distinguish them [29]. Since Pot1 can bind to single TTAGGG repeats, we speculated that Pot1 and Zbtb34 had the same binding site on the telomeres. To test this hypothesis, we generated constructs Myc-Pot1a and Flag-Pot1b. The upregulation of Zbtb34 in mESCs caused a drastic reduction in the localization of Pot1b at the telomeres. Similarly, the upregulation of Pot1b also affected the localization of Zbtb34 at telomeres, suggesting that Zbtb34 had a competitive relationship with Pot1b in telomere binding. It can be concluded from our experimental results that Zbtb34, ZBTB48 and ZBTB10, which are 3 ZnF proteins known to bind to telomeres, have different binding activity.
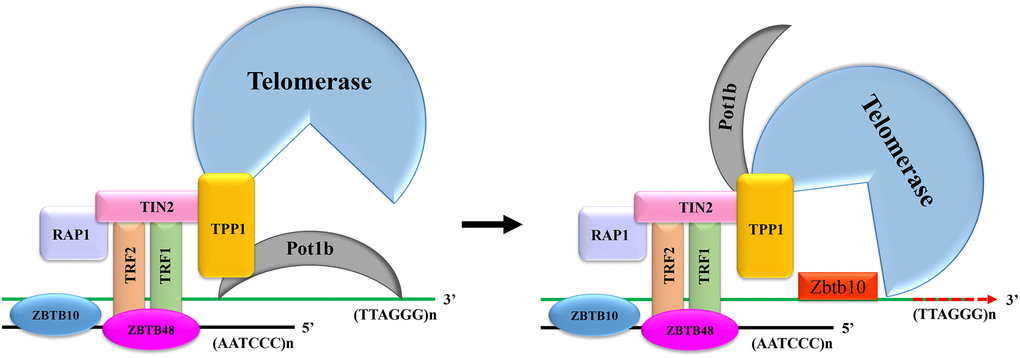
Figure 6. The mechanism for Zbtb34 to regulate telomere length. When the expression of Zbtb34 is more than that of Pot1b, Zbtb34 prevents Pot1b from binding to telomere DNA, which increases the opportunity for telomerase to access telomere DNA. The telomerase adds DNA repeats (TTAGGG)n to the 3′-ends of telomeres. When the expression of Pot1b is more than that of Zbtb34, Pot1b inhibits telomerase recruitment to telomeres, resulting in telomere shortening with cell division.
The depletion of Pot1 has been demonstrated to expose the 3′ telomere terminus and significantly promote telomere elongation [30, 31], which regulates telomere length by leaving telomeres occasionally, allowing telomerase to bind to telomeres, thus resulting in telomerase-mediated telomere lengthening [32]. Therefore, we speculated that the mechanism for Zbtb34 to facilitate telomere elongation was that Zbtb34 physically occupied the position of Pot1b at the telomeres and then helped telomerase interact with 3′ telomere terminus, resulting in telomere elongation (Figure 6). This result is consistent with the study of Gu et al., who recently found that Pot1 negatively regulated telomere length by inhibiting telomerase recruitment to telomeres [33]. Pot1b has been found to have important protective functions for telomeres. Pot1b knockout cells and mice resulted in telomere degradation caused by initiating a DNA damage response [34]. Based on the above experimental results, we speculated that Zbtb34 occupied the telomere binding sites, preventing the activation of DNA damage response. It also provided space for telomerase to bind to telomere DNA, resulting in telomere elongation and mESC proliferation (Figure 6). Telomere length maintenance is important to facilitating cell division [35], which is also critical for the unlimited self-renewal, pluripotency and chromosomal stability of mESCs [36]. The risk of diseases associated with a reduced cell proliferation, including aging or aging-associated diseases, is correlated with an increase in the shortening of telomeres [37]. An interesting finding of this study was that Zbtb34 did not affect the localization of Pot1a at telomeres, indicating that Pot1a and Pot1b played different roles in maintaining telomere functions in mESCs. The biological functions of Pot1a and Pot1b in telomere homeostasis need to be further investigated.
Conclusions
In conclusion, our results demonstrated that Zbtb34 could bind to telomeres, and for which ZnF3 was responsible. Furthermore, the upregulation of Zbtb34 reduced the Pot1b binding to telomere DNA sequences, indicating that the main function of Zbtb34 was to compete for the binding sites on telomeres through Pot1b, resulting in the telomere elongation and cell proliferation. The limitation of this study is that the functions of the other two ZnFs are still unclear. Therefore, ZnF1 and ZnF2 in Zbtb34 need to be further investigated.
Materials and Methods
Constructs
Zbtb34 cDNA was amplified by PCR using Pfu Turbo DNA polymerase (Agilent Corporation, USA) and appropriate primers (Table 1). After digesting the PCR products with Xho I and BamH I, the cDNA fragments were inserted into the Xho I/BamH I site of pEGFP-N1 to generate Zbtb34-EGFP fusion protein. Mouse Pot1 has two highly homologous isoforms, namely Pot1a and Pot1b, whose cDNA was amplified by PCR and then inserted into the Xho I/EcoR I sites of eukaryotic-expression vector pcDNA3.1 to generate pcDNA3.1-Myc-Pot1a as well as pcDNA3.1-Flag-Pot1b. The primers used in the study are listed in Table 1.
Table 1. The primers and siRNA used in this study.
| Primers | Sequences (5′→3′) | Target |
| Zbtb34 cDNA | Forward: CTCGAG ATGGAAAGCACCCTGG | Zbtb34 |
| Reverse: GAGCTCTTAGTCAGGCGCATC | ||
| Pot1a cDNA | Forward: CTCGAGATGTCTTTGGTTTCAAC | Pot1a |
| Reverse: GAATTCCTAGACAACATTTTCTG | ||
| Pot1b cDNA | Forward: CTCGAGATGTCTTCGGCCCCAG | Pot1b |
| Reverse: GAATTCCTAGATGTGTCTTC | ||
| ZnF1 | Forward: GTTGATCGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTATGGG | ZnF1 |
| Reverse: AGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCGATCAACCTCTCG | ZnF1 | |
| ZnF2 | Forward: TTTGTGGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTACTGATG | ZnF2 |
| Reverse: AGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCCACAAAGGGGGTGATC | ZnF2 | |
| ZnF3 | Forward: ATTCCGAGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTGCTCCGGGGGTCAC | ZnF3 |
| Reverse: AGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCAGCTCGGAATGGTTTGTC | ZnF3 | |
| siRNA 1 | Guide: UUCACUAAGCUAAAUUCAGCA | Zbtb34 |
| Passenger: CUGAAUUUAGCUUAGUGAAGG | Zbtb34 | |
| siRNA 2 | Guide: AACAGAAAAUAACCAUUAGAA | Zbtb34 |
| Passenger: CUAAUGGUUAUUUUCUGUUGA | Zbtb34 |
Bioinformatics analysis
Using the batch PWM predictor software (http://zf.princeton.edu/index.php), the positions of ZnFs within Zbtb34 were predicted. The Zbtb34-siRNA was designed through siDirect version 2.0 (http://sidirect2.rnai.jp/).
Alanine scanning mutagenesis
The expression vectors Zbtb34ZnF1 (alanine substitution of ZnF2 and ZnF3), Zbtb34ZnF2 (alanine substitution of ZnF1 and ZnF3) and Zbtb34ZnF3 (alanine substitution of ZnF1 and ZnF2) were constructed using site-directed mutagenesis kits (Sangon Biotech, Shanghai, China). Details of primers used in the study are listed in Table 1. The sequence and the mutants were confirmed through DNA sequencing.
Cell culture and transfection
C57BL/6 mESCs generated from the inner cell mass of a 3.5-day mouse blastocyst and OriCell C57BL/6129 culture media comprising fetal bovine serum, glutamine as well as 2-mercaptoethanol were purchased from Cyagen Bioscience Inc, Suzhou, China. The cells were transfected with constructs (4.0 μg) using Liperfactamine 3000 (Invitrogen, USA) and stained with Hoechst 33342 (10 mg/ml). The fluorescence was visualized through an Olympus BX51 fluorescence microscope.
Immunocytochemistry
Immunocytochemistry assay was performed as previously described [38]. The cells were fixed in 4% paraformaldehyde solution at room temperature for 30 min, and then washed and permeabilized in 0.5 ml of PBST solution for 30 min. Cells were then treated with primary antibodies Rabbit monoclonal to OCT4 (Abcam, ab200834), Rabbit monoclonal to CD15/SSEA1, (Abcam, ab135377), Rabbit monoclonal to NANONG (Abcam, ab203919), overnight at 4°C followed by fluorochrome labelled secondary antibodies Goat anti-Rabbit IgG Alexa Fluor 488 (Abcam, ab150077) and Goat Anti-Rabbit IgG Alexa Fluor 647 (Abcam, ab150083) at room temperature for 1 h. The images were captured with an Olympus BX51 microscope.
Fluorescence in situ hybridization (FISH)
mESCs transferred through vectors grown on coverslips were fixed for 5 min in 1% paraformaldehyde. Cells were then washed in PBS for 3 times. A Cy5-[CCCTAA]3-labelled probe (Sangon Biotech, Shanghai, China) was added to the coverslips in a buffer containing 70% formamide, 1 mg/ml blocking solution (1 mg/ml BSA, 3% goat serum, 0.1% Triton X-100, 1 mM EDTA in PBS) and 10 mM Tris-HCl pH 7.2, denatured at 80°C for 5 min, and hybridized for 1 h at room temperature in the dark. The coverslips were washed twice with 70% formamide and 10 mM Tris-HCl with a pH of 7.2 for 15 min each, which were then mounted for image analysis. This experiment was repeated for 3 times independently.
Telomere chromatin immunoprecipitation (ChIP)
The method refers to the study of Liu [39]. mESCs were cross-linked in 1% formaldehyde for 60 min and then added to 1 ml of SDS lysis buffer (1% SDS, 10 mM EDTA, 50 mM tris- HCl and 1 mM PMSF). Ultrasonically-induced cell lysis was used to obtain chromatin fragments <1 kB. The supernatant was added to 1 ml of IP buffer (0.01% SDS, 1.1% Triton X-100, 1.2 mM EDTA, 16.7 mM Tris, 167 mM NaCl and 1 mM PMSF), and then incubated with antibodies at 4°C overnight. Eluting buffer (10 mM Tris, 5 mM EDTA, 1% Triton X-100 and 0.1% SDS) was added at 65°C for 30 min and then de-crosslinked at 65°C. The protease K with a final concentration of 100 μg/ml was added and incubated at 50°C for 2 h. The immunoprecipitated DNA was purified through QIAEX II System. DNA was denatured and hybridized to a GeneScreen plus hybrid membrane, which was hybridized with a 32P-labelled-telomere repeats (4× TTAGGG) probe overnight at 42°C. The exposure and scanning were performed with a typhoon 9500 imager (GE Healthcare Life Sciences, USA). The anti-GFP (No. D191040), anti-Myc (No. D110006) and anti-Flag (No. D110005) antibodies were purchased from Sangon Biotech, Shanghai, China. This experiment was repeated for 3 times independently.
Real-time PCR
The quantitative real-time PCR was performed to determine the mRNA abundance of telomerase reverse transcriptase (TERT) and Zbtb34. mESCs were transfected through a Zbtb34-EGFP vector or Zbtb34-siRNA for 48 h. Total RNA was isolated from the cells using RNAzol (Sigma-Aldrich, China). mRNA was reversely-transcribed to cDNA using RevertAid first-strand cDNA synthesis kits (No. K1621, Sangon Biotech, Shanghai, China). Real-time PCR was run in a Roche 480 real-time PCR system. Master mix in each reaction tube includes cDNA, ddH2O, SYBR Green, forward and reverse primer of genes (Table 1). This experiment was repeated for at least 3 times independently.
Telomere length measurement
Real-time PCR was employed to measure telomere length using a relative mouse telomere length quantification qPCR assay kit (No.M8908, Shanghai ZhongQiaoXin Zhou Biotechnology, China) according to the manufacturer’s protocol. 200 μL of ddH2O was added to the telomeres and the SCR primer set. The 20 μL of qPCR in reaction included 1 ng of genomic DNA template, 2 μL of telomere primers and SCR primers, 10 μL of 2× qPCR master mix and ddH2O. The qPCR process lasted for 10 min at 95°C, followed by 32 cycles (denaturation at 95°C for 20 s; annealing at 52°C for 20 s and extension at 72°C for 45 s). This experiment was repeated for at least 3 times independently with similar results.
Telomerase activity measurement
Telomerase activity was analyzed using a telomerase enzyme-linked immunosorbent assay (ELISA) kit (No. ml037815, Mlbio, China). Briefly, 1 × 106 mESCs were lysed with 200 μL of RIPA lysis buffer (Beyotime, China). Subsequently, the protein supernatant and 50 μL of biotin-labeled antibodies were mixed at 37°C for 1 h. After washing with PBS to remove the unbound antibodies, 80 μL of horseradish peroxidase was added and incubated for 30 min at 37°C. 50 μL of substrates A and B were then added to each well after the washing was completed. Finally, after reading the absorbance values at 450 nm, telomerase activity (IU/L) of cells was calculated based on the standard curve created according to the standards of the kit. This experiment was repeated for 3 times independently with similar results.
MTT assay
A total of 1 × 104 cells per well were seeded in 96-well plates and grew to 80% confluence. The next day, the culture medium was completely removed, and 90 μL of PBS was added to each well. Then, the plates were incubated for 1 h at room temperature. 50 μL of MTT solution (5 mg/ml) was added, and cells were incubated in a CO2 incubator in the dark for 2 h. The medium was removed, and formazan crystals formed by the cells were dissolved in 500 μl of DMSO, which were then transferred to 96-well plates. The absorbance was read at a wavelength of 490 nm through a PT-3502C plate reader (Potenov, Beijing, China). This experiment was repeated for 3 times independently with similar results.
Flow cytometry
The cells were inoculated into a 6-cm dish and cultured for 48 h until they were adhered to the wall. The cell culture medium was collected in a 10 mL centrifuge tube and washed with PBS after incubation. Subsequently, the cells were centrifuged to remove the supernatant, remixed with binding buffer and mixed with Annexin V-FITC as well as propidium iodide (PI) staining solution. After a thorough mixing, the cells were incubated at room temperature in the dark for 15 min. PBS was added to the cell suspension in the flow tube, and a flow cytometry assay (Beckman Coulter, USA) was performed. This experiment was repeated for 3 times independently with similar results.
Statistical analysis
The data was expressed by mean ± SD. Differences were considered statistically significant at P < 0.05. All statistical analyses were performed using SPSS 20 (SPSS Inc., USA). All experiments were repeated for 3 independent times.
Author Contributions
Z.L. and T.L. designed the experiments and supervised the work. Z. L., X.W., Y. G., X. G., X.L., Y. Z., and X.W. performed the cell culture, construct, telomere ChIP, and qPCR. C. L., T.S., J. L. conduced bioinformatics analysis, MTT, and prepared the figures. Z. L. and T. L. analyzed the data and wrote the manuscript. All authors participated in discussion of the final manuscript and interpretation of results.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Funding
This research was supported by Guangxi Science and Technology Base and Talent Project under Grant No. AD19110161 (Application No: 2018AD19267), Guangxi Natural Science Foundation under Grant No. 2018GXNSFAA281048 (Application No: 2018JJA140045), Innovation Project of Guangxi Graduate Education (No. YCSW2021256), and Innovation and Entrepreneurship Training Program for College Students (No.202010601139).
References
- 1. Cassandri M, Smirnov A, Novelli F, Pitolli C, Agostini M, Malewicz M, Melino G, Raschellà G. Zinc-finger proteins in health and disease. Cell Death Discov. 2017; 3:17071. https://doi.org/10.1038/cddiscovery.2017.71 [PubMed]
- 2. Ding W, Wang Y, Fang W, Gao S, Li X, Xiao K. TaZAT8, a C2H2-ZFP type transcription factor gene in wheat, plays critical roles in mediating tolerance to Pi deprivation through regulating P acquisition, ROS homeostasis and root system establishment. Physiol Plant. 2016; 158:297–311. https://doi.org/10.1111/ppl.12467 [PubMed]
- 3. Jamil W, Wu W, Gong H, Huang JW, Ahmad M, Zhu QG, Jin R, Liu XF, Yin XR. C2H2-Type Zinc Finger Proteins (DkZF1/2) Synergistically Control Persimmon Fruit Deastringency. Int J Mol Sci. 2019; 20:5611. https://doi.org/10.3390/ijms20225611 [PubMed]
- 4. He K, Zhang J, Liu J, Cui Y, Liu LG, Ye S, Ban Q, Pan R, Liu D. Functional genomics study of protein inhibitor of activated STAT1 in mouse hippocampal neuronal cells revealed by RNA sequencing. Aging (Albany NY). 2021; 13:9011–27. https://doi.org/10.18632/aging.202749 [PubMed]
- 5. Singh JK, van Attikum H. DNA double-strand break repair: Putting zinc fingers on the sore spot. Semin Cell Dev Biol. 2021; 113:65–74. https://doi.org/10.1016/j.semcdb.2020.09.003 [PubMed]
- 6. Hudson NO, Buck-Koehntop BA. Zinc Finger Readers of Methylated DNA. Molecules. 2018; 23:2555. https://doi.org/10.3390/molecules23102555 [PubMed]
- 7. Kim DN, Thiel BC, Mrozowich T, Hennelly SP, Hofacker IL, Patel TR, Sanbonmatsu KY. Zinc-finger protein CNBP alters the 3-D structure of lncRNA Braveheart in solution. Nat Commun. 2020; 11:148. https://doi.org/10.1038/s41467-019-13942-4 [PubMed]
- 8. Qi J, Zhang X, Zhang HK, Yang HM, Zhou YB, Han ZG. ZBTB34, a novel human BTB/POZ zinc finger protein, is a potential transcriptional repressor. Mol Cell Biochem. 2006; 290:159–67. https://doi.org/10.1007/s11010-006-9183-x [PubMed]
- 9. Kasiappan R, Jutooru I, Mohankumar K, Karki K, Lacey A, Safe S. Reactive Oxygen Species (ROS)-Inducing Triterpenoid Inhibits Rhabdomyosarcoma Cell and Tumor Growth through Targeting Sp Transcription Factors. Mol Cancer Res. 2019; 17:794–805. https://doi.org/10.1158/1541-7786.MCR-18-1071 [PubMed]
- 10. Jutooru I, Guthrie AS, Chadalapaka G, Pathi S, Kim K, Burghardt R, Jin UH, Safe S. Mechanism of action of phenethylisothiocyanate and other reactive oxygen species-inducing anticancer agents. Mol Cell Biol. 2014; 34:2382–95. https://doi.org/10.1128/MCB.01602-13 [PubMed]
- 11. Liao J, Liu J, Wang J, Wang M. Lnc-CPLC promotes the progression of colorectal cancer via regulating ZBTB34 by competitively binding miR-4319. J Cell Physiol. 2022; 237:1573–85. https://doi.org/10.1002/jcp.30628 [PubMed]
- 12. D'Angelo W, Acharya D, Wang R, Wang J, Gurung C, Chen B, Bai F, Guo YL. Development of Antiviral Innate Immunity During In Vitro Differentiation of Mouse Embryonic Stem Cells. Stem Cells Dev. 2016; 25:648–59. https://doi.org/10.1089/scd.2015.0377 [PubMed]
- 13. Evans M. Discovering pluripotency: 30 years of mouse embryonic stem cells. Nat Rev Mol Cell Biol. 2011; 12:680–6. https://doi.org/10.1038/nrm3190 [PubMed]
- 14. Martínez P, Blasco MA. Role of shelterin in cancer and aging. Aging Cell. 2010; 9:653–66. https://doi.org/10.1111/j.1474-9726.2010.00596.x [PubMed]
- 15. Vilas CK, Emery LE, Denchi EL, Miller KM. Caught with One's Zinc Fingers in the Genome Integrity Cookie Jar. Trends Genet. 2018; 34:313–25. https://doi.org/10.1016/j.tig.2017.12.011 [PubMed]
- 16. Li JS, Miralles Fusté J, Simavorian T, Bartocci C, Tsai J, Karlseder J, Lazzerini Denchi E. TZAP: A telomere-associated protein involved in telomere length control. Science. 2017; 355:638–41. https://doi.org/10.1126/science.aah6752 [PubMed]
- 17. Bluhm A, Viceconte N, Li F, Rane G, Ritz S, Wang S, Levin M, Shi Y, Kappei D, Butter F. ZBTB10 binds the telomeric variant repeat TTGGGG and interacts with TRF2. Nucleic Acids Res. 2019; 47:1896–907. https://doi.org/10.1093/nar/gky1289 [PubMed]
- 18. Jung SJ, Seo YR, Park WJ, Heo YR, Lee YH, Kim S, Lee JH. Clinicopathological Characteristics of TZAP Expression in Colorectal Cancers. Onco Targets Ther. 2020; 13:12933–42. https://doi.org/10.2147/OTT.S274394 [PubMed]
- 19. Hedrick E, Li X, Safe S. Penfluridol Represses Integrin Expression in Breast Cancer through Induction of Reactive Oxygen Species and Downregulation of Sp Transcription Factors. Mol Cancer Ther. 2017; 16:205–16. https://doi.org/10.1158/1535-7163.MCT-16-0451 [PubMed]
- 20. Hedrick E, Crose L, Linardic CM, Safe S. Histone Deacetylase Inhibitors Inhibit Rhabdomyosarcoma by Reactive Oxygen Species-Dependent Targeting of Specificity Protein Transcription Factors. Mol Cancer Ther. 2015; 14:2143–53. https://doi.org/10.1158/1535-7163.MCT-15-0148 [PubMed]
- 21. Semeraro MD, Smith C, Kaiser M, Levinger I, Duque G, Gruber HJ, Herrmann M. Physical activity, a modulator of aging through effects on telomere biology. Aging (Albany NY). 2020; 12:13803–23. https://doi.org/10.18632/aging.103504 [PubMed]
- 22. Turner KJ, Vasu V, Griffin DK. Telomere Biology and Human Phenotype. Cells. 2019; 8:73. https://doi.org/10.3390/cells8010073 [PubMed]
- 23. Penev A, Markiewicz-Potoczny M, Sfeir A, Lazzerini Denchi E. Stem cells at odds with telomere maintenance and protection. Trends Cell Biol. 2022; 32:527–36. https://doi.org/10.1016/j.tcb.2021.12.007 [PubMed]
- 24. Frescas D, de Lange T. TRF2-tethered TIN2 can mediate telomere protection by TPP1/POT1. Mol Cell Biol. 2014; 34:1349–62. https://doi.org/10.1128/MCB.01052-13 [PubMed]
- 25. Cicconi A, Chang S. Shelterin and the replisome: at the intersection of telomere repair and replication. Curr Opin Genet Dev. 2020; 60:77–84. https://doi.org/10.1016/j.gde.2020.02.016 [PubMed]
- 26. Ghilain C, Gilson E, Giraud-Panis MJ. Multifunctionality of the Telomere-Capping Shelterin Complex Explained by Variations in Its Protein Composition. Cells. 2021; 10:1753. https://doi.org/10.3390/cells10071753 [PubMed]
- 27. Xin H, Liu D, Wan M, Safari A, Kim H, Sun W, O'Connor MS, Songyang Z. TPP1 is a homologue of ciliate TEBP-beta and interacts with POT1 to recruit telomerase. Nature. 2007; 445:559–62. https://doi.org/10.1038/nature05469 [PubMed]
- 28. Kelleher C, Kurth I, Lingner J. Human protection of telomeres 1 (POT1) is a negative regulator of telomerase activity in vitro. Mol Cell Biol. 2005; 25:808–18. https://doi.org/10.1128/MCB.25.2.808-818.2005 [PubMed]
- 29. Doksani Y, Wu JY, de Lange T, Zhuang X. Super-resolution fluorescence imaging of telomeres reveals TRF2-dependent T-loop formation. Cell. 2013; 155:345–56. https://doi.org/10.1016/j.cell.2013.09.048 [PubMed]
- 30. Yu Y, Tan R, Ren Q, Gao B, Sheng Z, Zhang J, Zheng X, Jiang Y, Lan L, Mao Z. POT1 inhibits the efficiency but promotes the fidelity of nonhomologous end joining at non-telomeric DNA regions. Aging (Albany NY). 2017; 9:2529–43. https://doi.org/10.18632/aging.101339 [PubMed]
- 31. Aramburu T, Plucinsky S, Skordalakes E. POT1-TPP1 telomere length regulation and disease. Comput Struct Biotechnol J. 2020; 18:1939–46. https://doi.org/10.1016/j.csbj.2020.06.040 [PubMed]
- 32. Smith EM, Pendlebury DF, Nandakumar J. Structural biology of telomeres and telomerase. Cell Mol Life Sci. 2020; 77:61–79. https://doi.org/10.1007/s00018-019-03369-x [PubMed]
- 33. Gu P, Jia S, Takasugi T, Tesmer VM, Nandakumar J, Chen Y, Chang S. Distinct functions of POT1 proteins contribute to the regulation of telomerase recruitment to telomeres. Nat Commun. 2021; 12:5514. https://doi.org/10.1038/s41467-021-25799-7 [PubMed]
- 34. Lazzerini-Denchi E, Sfeir A. Stop pulling my strings - what telomeres taught us about the DNA damage response. Nat Rev Mol Cell Biol. 2016; 17:364–78. https://doi.org/10.1038/nrm.2016.43 [PubMed]
- 35. Guerrero EN, Vega S, Fu C, De León R, Beltran D, Solis MA. Increased proliferation and differentiation capacity of placenta-derived mesenchymal stem cells from women of median maternal age correlates with telomere shortening. Aging (Albany NY). 2021; 13:24542–59. https://doi.org/10.18632/aging.203724 [PubMed]
- 36. Xu C, Xie N, Su Y, Sun Z, Liang Y, Zhang N, Liu D, Jia S, Xing X, Han L, Li G, Tong T, Chen J. HnRNP F/H associate with hTERC and telomerase holoenzyme to modulate telomerase function and promote cell proliferation. Cell Death Differ. 2020; 27:1998–2013. https://doi.org/10.1038/s41418-019-0483-6 [PubMed]
- 37. Celtikci B, Erkmen GK, Dikmen ZG. Regulation and Effect of Telomerase and Telomeric Length in Stem Cells. Curr Stem Cell Res Ther. 2021; 16:809–23. https://doi.org/10.2174/1574888X15666200422104423 [PubMed]
- 38. Long P, Liu Z, Wu B, Chen J, Sun C, Wang F, Huang Y, Chen H, Li Q, Ma Y. Generation of an induced pluripotent stem cell line from chorionic villi of a Patau syndrome spontaneous abortion. Stem Cell Res. 2020; 45:101789. https://doi.org/10.1016/j.scr.2020.101789 [PubMed]
- 39. Liu F, Feng X, Ma W. Analysis of Telomere Proteins by Chromatin Immunoprecipitation (ChIP). Methods Mol Biol. 2017; 1587:205–14. https://doi.org/10.1007/978-1-4939-6892-3_19 [PubMed]