



Introduction
Lung cancer is a frequently diagnosed cancer worldwide, and the leading cause of cancer-related death [1]. Lung adenocarcinoma (LUAD), which comprises 40% of all lung cancers cases, is responsible for over 500,000 deaths per year worldwide [2]. Considerable efforts have been made to develop novel anticancer drugs for LUAD patients over the last decade, but their 5-year overall survival (OS) rates are still poor, probably due to the high heterogeneity of the disease and lack of effective diagnostic and prognostic biomarkers [3–6]. The advancement of immunotherapy can serve as a promising rescue option for LUAD patients [3]. Several studies have demonstrated that immune checkpoint inhibitors could improve OS, progression-free survival (PFS), and objective response rates in patients with non-small cell lung cancer when used in the first- or subsequent-line settings [7–9]. However, a substantial portion of patients failed to respond to this therapy [5, 7–10]. Therefore, it is of utmost urgency to develop novel therapeutic targets and reliable biomarkers for LUAD treatment.
Kinesin family member 14 (KIF14), is an emerging molecular motor that involved in cytokinesis completion, midbody formation, chromosome segregation, mitotic spindle formation and cargo-containing vesicle transport [11–14]. KIF14 is potentially oncogenic and can act as a chromokinesin via binding to microtubules and chromatin during the bipolar spindle formation [11, 14, 15]. High expression of KIF14 is demonstrated to be a significant prognostic biomarker in various cancers, including pancreatic adenocarcinoma (PAAD), hepatocellular carcinoma, cervical cancer and ovarian cancer [16–19]. Knocking-down of KIF14 could result in the failure of cytokinesis and inhibition of cancer cell growth, while overexpression of KIF14 induced cancer cell proliferation [15, 20]. As an ATPase, it also has potential as a therapeutic target [21]. However, the expression, prognosis, mechanisms and tumor immune regulation of KIF14 in patients with LUAD remain obscure.
In the present study, both expression and survival analyses were conducted on KIF14 in pan-cancer with an emphasis on LUAD. Moreover, the association between KIF14 and long non-coding RNAs (lncRNA) or microRNAs (miRNAs) in LUAD were evaluated. In addition, we explored the relationships of KIF14 with immune checkpoints-related genes, immune cell biomarkers and immune cell infiltration (ICI) in LUAD. The results indicated that lncRNAs- and miRNAs-regulated overexpression of KIF14 was associated with tumor immune infiltration and unfavorable prognosis in patients with LUAD.
Results
The expression of KIF14 in pan-cancer
Expression analysis of KIF14 through the Oncomine database (http://www.oncomine.org) revealed that KIF14 was overexpressed in numerous cancers, including LUAD, compared with their respective adjacent tissues (Figure 1A). The results of difference expression analyses through The Cancer Genome Atlas (TCGA) database (http://portal.gdc.cancer.gov) confirmed that KIF14 was upregulated not only in LUAD, but also in uterine corpus endometrial carcinoma (UCEC), thyroid carcinoma (THCA), stomach adenocarcinoma (STAD), sarcoma (SARC), rectum adenoma (READ), prostate adenocarcinoma (PRAD), PAAD, lung squamous cell carcinoma (LUSC), liver hepatocellular carcinoma (LIHC), kidney renal papillary cell carcinoma (KIRP), kidney renal clear cell carcinoma (KIRC), head and neck squamous cell carcinoma (HNSC), glioblastoma multiforme (GBM), esophageal carcinoma (ESCA), colon adenocarcinoma (COAD), cholangiocarcinoma (CHOL), endocervical adenocarcinoma (CESC), breast invasive carcinoma (BRCA), cervical squamous cell carcinoma and bladder urothelial carcinoma (BLCA) (Figure 1B).
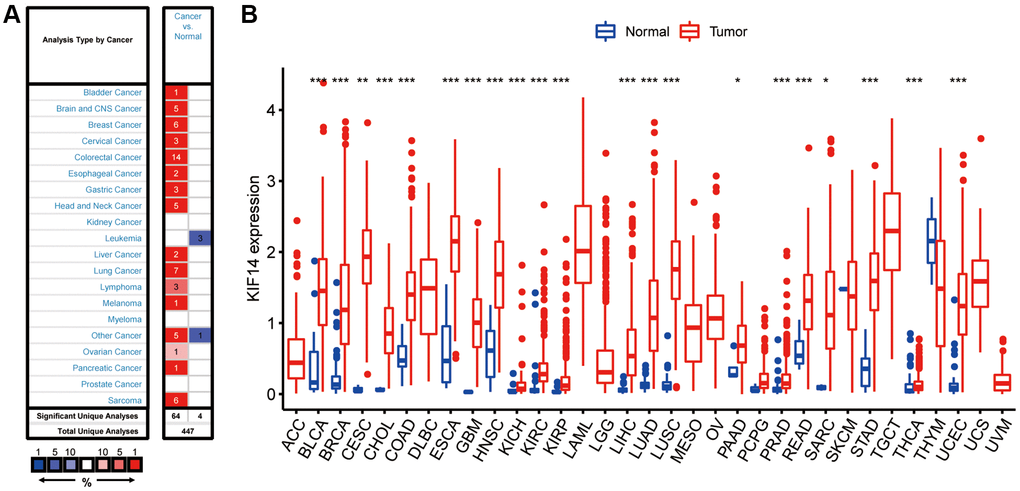
Figure 1. Expression of KIF14 in pan-cancers. (A) Upregulated or downregulated levels of KIF14 in different tumors collected from the Oncomine database. (B). Expression levels of KIF14 in various tumors from The Cancer Genome Atlas (TCGA) database analyzed by R package. *P-value < 0.05, **P-value < 0.01, ***P-value < 0.001.
High-expression of KIF14 predicts unfavorable prognosis
The Kaplan-Meier (K-M) survival analysis was conducted to assess the correlation between KIF14 expression and survival outcomes (PFS and OS) in pan-cancer according to the TCGA database. Both OS and PFS results demonstrated that high KIF14 expression could predict unfavorable prognosis in LUAD, PAAD, mesothelioma (MESO), LIHC, KIRP, KIRC, and adrenocortical carcinoma (ACC) (Figure 2A and 2G, Supplementary Figures 1 and 2). Next, we verified the results of LUAD via the PrognoScan database (http://dna00.bio.kyutech.ac.jp/PrognoScan/index.html) [22] (Table 1). Notably, high KIF14 expression was significantly correlated with shorter OS (Figure 2B–2F) and relapse-free survival (RFS) (Figure 2H and 2I) in seven out of ten independent cohorts. In the remaining three cohorts [23] (jacob-00182-CANDF, jacob-00182-UM, and jacob-00182-HLM cohorts, Figure 2J–2L), the OS of KIF14 upregulation group was still shorter than that of KIF14 downregulation group, but no significant difference was observed. For TCGA-LUAD cohort, both univariate and multivariate Cox regression analysis revealed that KIF14 was an independent prognostic factor for OS (Figure 2M and 2N), with Akaike information criterion (AIC) = 1094.23, p-value = 1e-04. Altogether, these results demonstrate that KIF14 can serve as a prognostic biomarker for patients with LUAD.
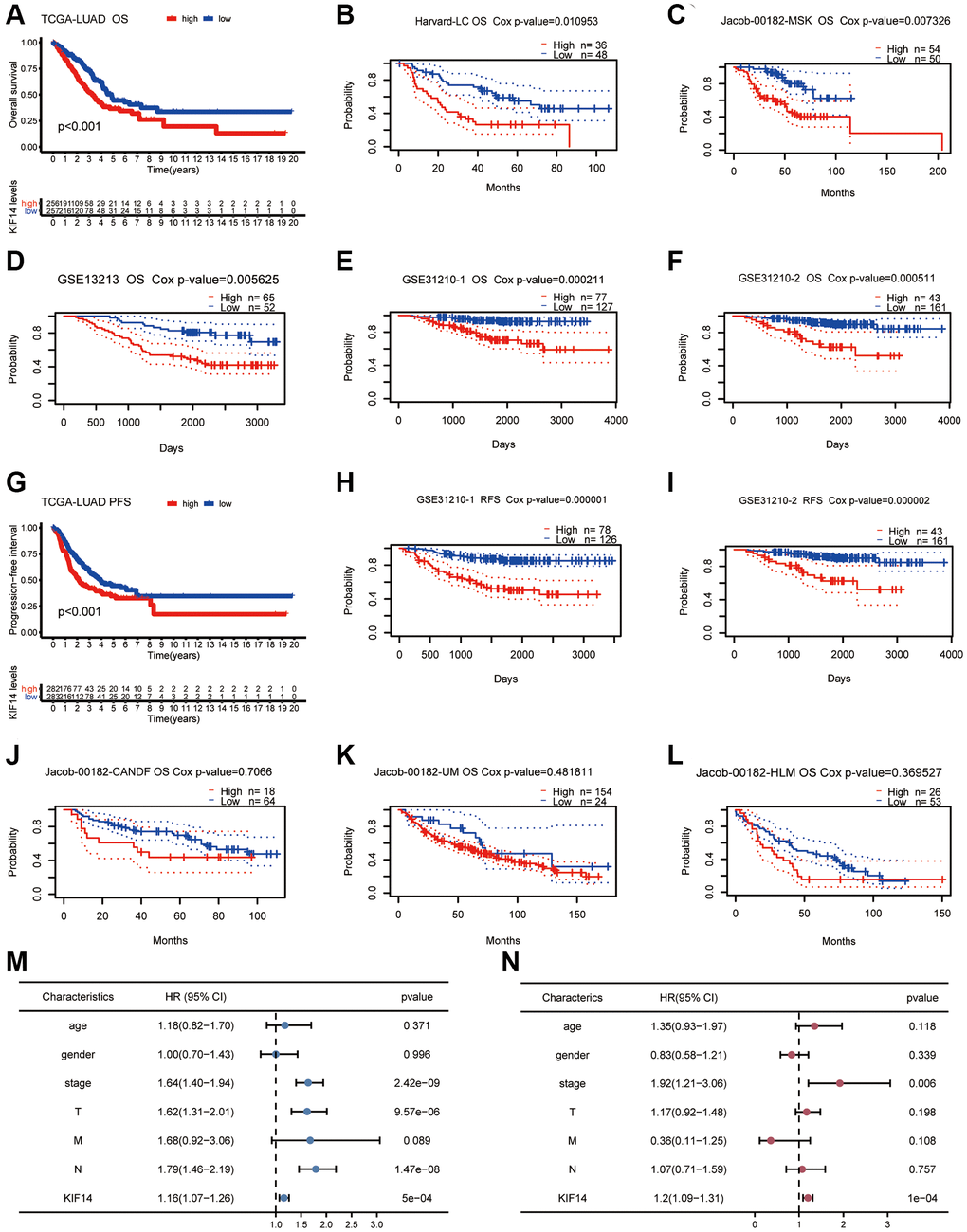
Figure 2. Survival analyses of KIF14 in LUAD. (A–F) K-M plots for OS in different LUAD cohorts (TCGA-LUAD cohort, Harvard -LC cohort, jacob-00182-MSK cohort, GSE13212 cohort, GSE31210-1 cohort, and GSE31210-2 cohort). (G–I) Survival curves of PFS, RFS in LUAD cohorts (TCGA-LUAD cohort, GSE31210-1 cohort, and GSE31210-2 cohort). (J–L) OS survival analyses of KIF14 in LUAD cohorts with negative results (jacob-00182-CANDF cohort, jacob-00182-UM cohort and jacob-00182-HLM cohort). (M, N) Univariate (M) and multivariate (N) Cox regression analyses to evaluate the independent factors of KIF14 in TCGA-LUAD cohort.
Table 1. Expression of KIF14 in lung adenocarcinoma in PrognoScan database.
| Cancer | Dataset | Endpoint | Probe ID | N | Cox P-value | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LUAD | jacob-00182-CANDF | OS | 206364_at | 82 | 0.7066 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LUAD | HARVARD-LC | OS | 34563_at | 84 | 0.003813 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LUAD | jacob-00182-HLM | OS | 206364_at | 79 | 0.369527 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LUAD | jacob-00182-MSK | OS | 206364_at | 104 | 0.001179 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LUAD | GSE13213 | OS | A_23_P149668 | 117 | 0.001809 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LUAD | GSE31210 | OS | 236641_at | 204 | 0.000691 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LUAD | GSE31210 | OS | 206364_at | 204 | 0.001082 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LUAD | GSE31210 | RFS | 206364_at | 204 | 0.000028 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LUAD | GSE31210 | RFS | 236641_at | 204 | 0.000005 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LUAD | jacob-00182-UM | OS | 206364_at | 178 | 0.481811 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Abbreviations: LUAD: Lung adenocarcinoma; OS: Overall survival; RFS: Relapse free survival. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Overexpression of KIF14 correlated with adverse clinical characteristics in LUAD
The correlation between KIF14 expression and clinical characteristics in LUAD were analyzed. The results showed that KIF14 was positively related to advanced tumor stage (Figure 3A, p = 0.021) and lymph node metastasis (Figure 3B, p = 0.014). Moreover, high expression of KIF14 were more common in patients with uncontrolled disease (Figure 3C, p < 0.001), smoker (Figure 3D, p = 0.001), younger patients (Figure 3E, p = 0.001), and male (Figure 3F, p = 0.001). The associations between the expression of KIF14 and T stage and M stage were not statistically significant (Figure 3G and 3H, p > 0.05). Hence, all the above findings suggest that KIF14 might be involved in the tumorigenesis of LUAD.
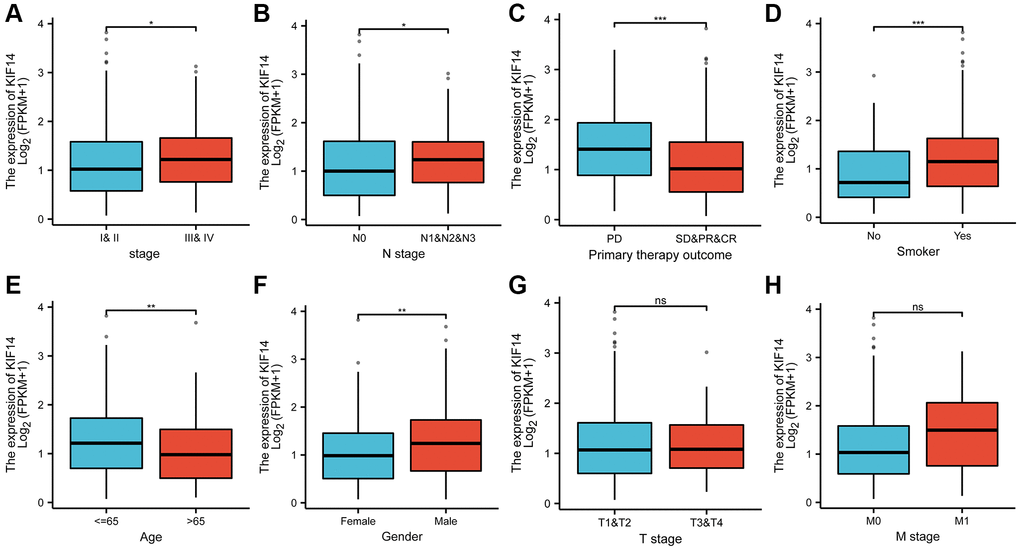
Figure 3. The correlations between KIF14 expression and pathological characteristics in LUAD. Upregulation of KIF14 is positively related to higher pathological stage (A), N stage (B), uncontrolled disease (C), smoking (D), younger patients(E), and male (F) in TCGA-LUAD cohort. The associations between the expression of KIF14 and T stage (G) and M stage (H) were not statistically significant. *P-value < 0.05, **P-value < 0.01, ***P-value < 0.001. Abbreviation: ns: non-statical significant.
Prediction and construction of the ceRNA network of KIF14 in LUAD
Identification of upstream miRNAs of KIF14
To explore the upstream miRNAs regulating KIF14 expression, the binding of miRNAs to KIF14 was assessed using the starBase database (http://starbase.sysu.edu.cn) [24], and then visualized in a miRNAs-KIF14 regulatory network constructed by Cytoscape tool [25]. In view of the action mechanisms by which miRNAs negatively regulate KIF14 expression, the miRNAs were then filtered according to the following criteria: (i) downregulation patterns in LUAD, (ii) positive correlation with LUAD patients’ prognosis, and (iii) negative correlation with KIF14 expression. Finally, a total of 7 miRNAs were included (Table 2). Among them, hsa-miR-101-3p was further selected for subsequent analysis, which exhibited the highest correlation with KIF14 expression in LUAD patients (Table 2, Figure 4A and 4D–4F). The binding target sequence information can be found in Supplementary Figure 3.
Table 2. Potential upstream miRNAs of KIF14.
| Gene | miRNA | R-value | P-value |
| KIF14 | hsa-miR-101-3p | −0.404705546 | 0 |
| KIF14 | hsa-let-7c-5p | −0.362901213 | 3.12E-16 |
| KIF14 | hsa-miR-133a-3p | −0.352791372 | 2.28E-15 |
| KIF14 | hsa-miR-195-5p | −0.35086189 | 4.22E-15 |
| KIF14 | hsa-miR-218-5p | −0.262940449 | 6.79E-09 |
| KIF14 | hsa-miR-34c-5p | −0.243877863 | 8.08E-08 |
| KIF14 | hsa-miR-133b | −0.223939 | 8.20E-07 |
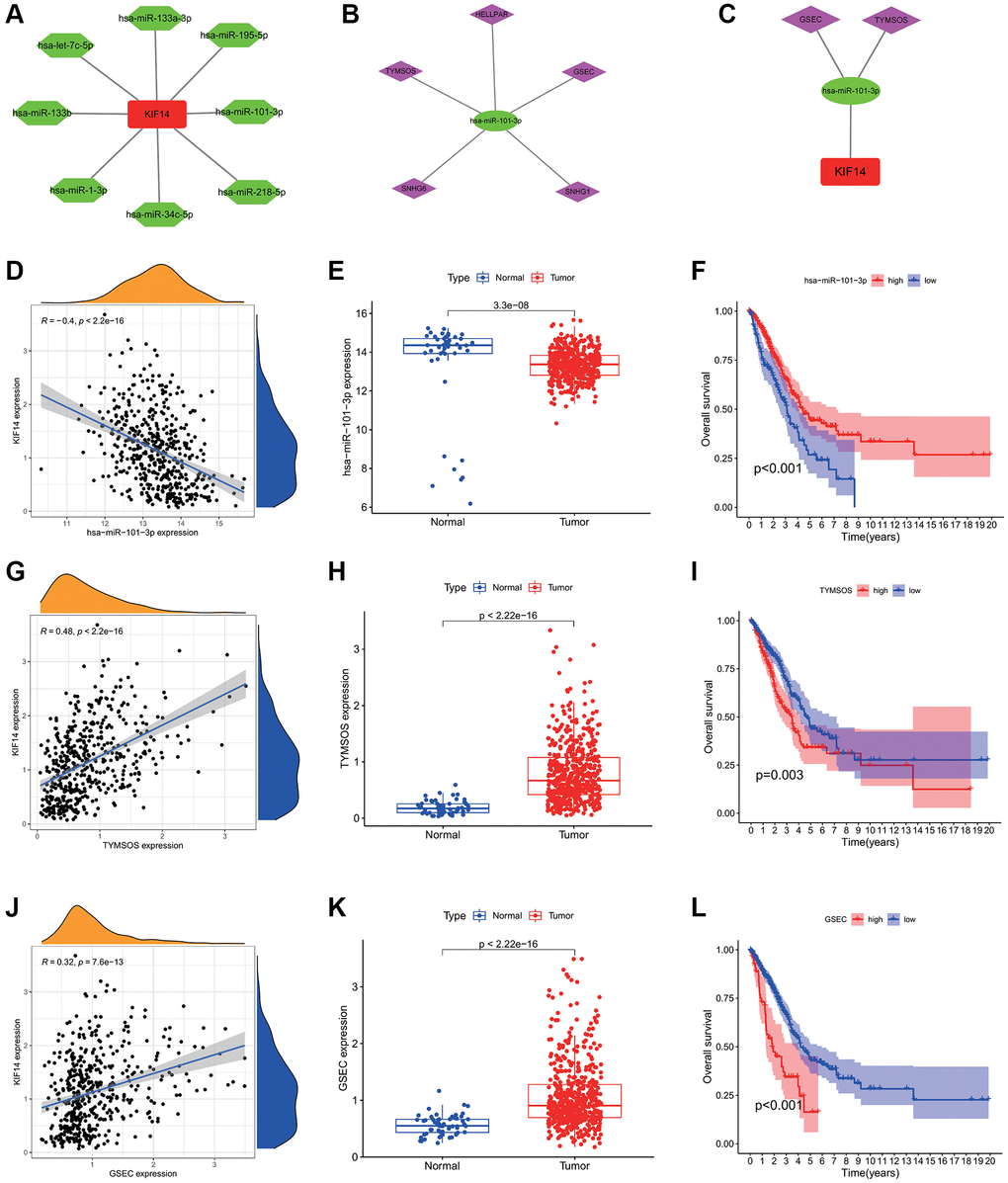
Figure 4. Construction of the ceRNA network of KIF14 in LUAD. (A) The potential miRNAs-KIF14 regulatory network constructed via Cytoscape tool. (B) The potential lncRNAs-hsa-miR-101-3p regulatory network constructed via Cytoscape tool. (C) The ceRNA network of GSEC/TYMSOS-hsa-miR-101-3p-KIF14 in LUAD. (D) The correlation between hsa-miR-101-3p and KIF14 expression in LUAD. (E) Expression levels of hsa-miR-101-3p in LUAD and control tissues in TCGA-LUAD cohort. (F) Survival analysis of hsa-miR-101-3p in TCGA-LUAD cohort. (G) The correlation between TYMSOS and KIF14 expression in LUAD. (H) Expression levels of TYMSOS in LUAD and control tissues in TCGA-LUAD cohort. (I) Survival analysis of TYMSOS in TCGA-LUAD cohort. (J) The correlation analysis of GSEC and KIF14 expressions in LUAD. (K) The differential expression of GSEC in LUAD and normal tissues in TCGA-LUAD cohort. (L) Survival analysis of GSEC in TCGA-LUAD cohort.
Prediction and construction of the ceRNA network of KIF14
Through the starBase database, the potential upstream lncRNAs of hsa-miR-101-3p were predicted (Figure 4B). According to the ceRNA hypothesis, these lncRNAs were also filtered by the following criteria [26, 27]: (i) upregulation patterns in LUAD, (ii) positive correlation with KIF14 expression, (iii) negative correlation with hsa-miR-101-3p expression, (iv) negative correlation with LUAD patients’ prognosis, and (v) significant association with unfavorable prognosis in LUAD patients. According to the results listed in Tables 3–5, GSEC and TYMSOS (Figure 4B, 4C and 4G–4L), which were negatively correlated LUAD patients’ OS, were considered as the two most potential upstream lncRNAs of hsa-miR-101-3p/KIF14 axis in LUAD patients (Figure 5), while SNHG6, HELLPAR, and SNHG1 were excluded (Supplementary Figure 4).
Table 3. Association analysis between lncRNA and hsa-miR-101-3p or lncRNA and KIF14 in LUAD determined via starBase database.
| lncRNA | miRNA | R-value | P-value |
| GSEC | hsa-miR-101-3p | −0.224264048 | 8.45E-07 |
| HELLPAR | hsa-miR-101-3p | −0.243521462 | 8.45E-08 |
| SNHG1 | hsa-miR-101-3p | −0.222764208 | 1.00E-06 |
| SNHG6 | hsa-miR-101-3p | −0.216861364 | 1.95E-06 |
| TYMSOS | hsa-miR-101-3p | −0.287171466 | 2.19E-10 |
Table 4. Association analysis between lncRNA and KIF14 in LUAD determined via starBase database.
| lncRNA | Gene | R-value | P-value |
| SNHG6 | KIF14 | 0.166466613 | 0.000273472 |
| TYMSOS | KIF14 | 0.484935757 | 0 |
| HELLPAR | KIF14 | 0.51181722 | 0 |
| GSEC | KIF14 | 0.32269557 | 7.57E-13 |
| SNHG1 | KIF14 | 0.368245931 | 7.11E-17 |
Table 5. Correlation analysis between lncRNA and hsa-miR101-3p in LUAD.
| lncRNA | miRNA | R-value | P-value |
| SNHG6 | hsa-miR-101-3p | −0.216861364 | 1.95E-06 |
| TYMSOS | hsa-miR-101-3p | −0.287171466 | 2.19E-10 |
| HELLPAR | hsa-miR-101-3p | −0.243521462 | 8.45E-08 |
| GSEC | hsa-miR-101-3p | −0.224264048 | 8.45E-07 |
| SNHG1 | hsa-miR-101-3p | −0.222764208 | 1.00E-06 |
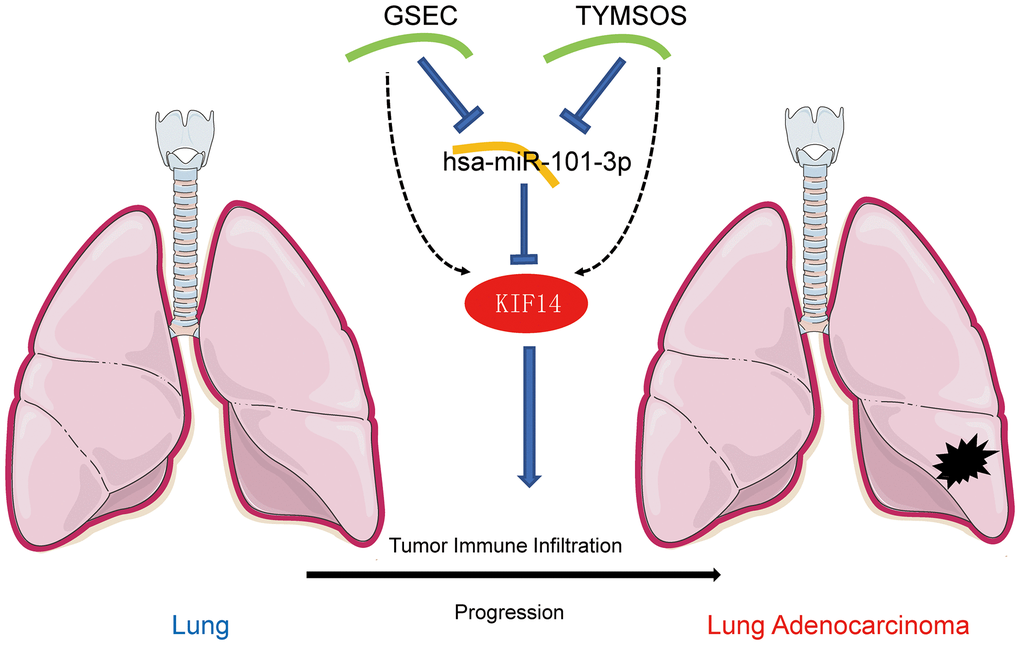
Figure 5. The model of GSEC/TYMSOS-hsa-miR-101-3p axis in tumorigenesis of LUAD.
Association of KIF14 expression with tumor ICI in LUAD
Association of KIF14 expression with TMB and MSI
TMB is defined as the distribution density of nonsynonymous mutations in the protein- coding sequences, and is closely association with tumor prognosis. MSI refers to the changes in the length of microsatellite sequences due to deletion or insertion mutations during the process of DNA replication, which is mostly caused by functional defects such as the mismatch repair system (MMRs), and is involved in tumor development. Both TMB and MSI have been found to be closely associated with immunotherapeutic response. The expression of KIF14 was remarkably correlated with TMB in LUAD, UCEC, THYM, STAD, skin cutaneous melanoma (SKCM), SARC, READ, PRAD, PAAD, MESO, LUSC, KIRC, KICH, HNSC, COAD, BRCA, brain lower grade glioma (BLGG), BLCA, acute myeloid leukemia (AML) and ACC (Figure 6A). Meanwhile, the expression of KIF14 was markedly associated with MSI in LUAD, UCEC, testicular germ cell tumors (TGCT), STAD, SKCM, SARC, READ, LUSC, GBM, diffuse large B-cell lymphoma (DLBC) in lymphoid neoplasm, COAD and ACC (Figure 6B). Interestingly, the expression of KIF14 was closely related to both TMB and MSI in LUAD, indicating that KIF14 may be used as a biomarker for immunotherapy in LUAD patients.
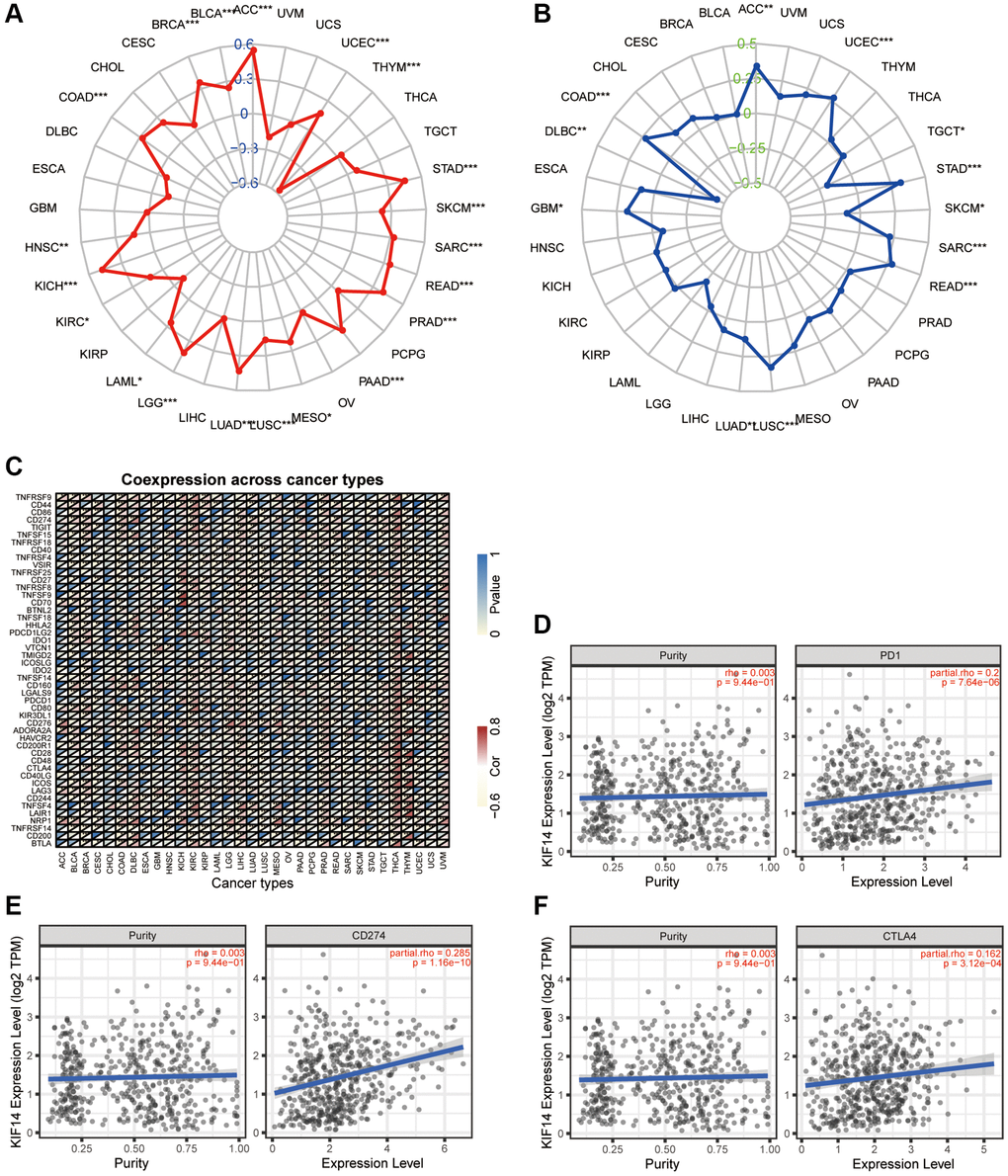
Figure 6. Association between KIF14 expression and immune infiltration in LUAD. (A–C) The association of KIF14 expression with TMB (A), MSI (B) and immune checkpoint-related genes (C) in pan-cancer. (D–F) The associations of KIF14 expression with PD-1 (D), CD274 (E) and CTLA4 (F) expression in LUAD, which were determined using the TIMER database. *P-value < 0.05, **P-value < 0.01, ***P-value < 0.001.
Discussion
Increasing evidence have demonstrated KIF14 plays vital roles in regulating tumorigeneses and progression of various cancer [16, 21, 32, 33]. Abnormal expression of KIF14 contributed to unfavorable prognosis in patients with cancer, and was associated with cancer cell proliferation and progression [16–20, 34]. However, the role of KIF14 in mediating LUAD carcinogenesis is still unclarified, and awaits further research.
In the present study, pan-cancer analyses of KIF14 expression were performed via Oncomine and TCGA databases. Survival analyses indicated that high expression of KIF14 was correlated with worse prognosis in various cancer types, including LUAD.
High expression of KIF14 was positively related to advanced tumor stage, N stage, uncontrolled disease, smoking history and male. All the above findings suggest that KIF14 might be involved in the tumorigenesis of LUAD. Moreover, both univariate and multivariate Cox regression analyses demonstrated that KIF14 was an independent prognostic factor for LUAD. Furthermore, survival analyses through the PrognoScan database confirmed that KIF14 contributed to unfavorable prognosis in patients with LUAD. Altogether, these results reveal the oncogenic role of KIF14 in LUAD.
It is well-known that miRNAs participate in tumorigenic process by primarily regulating the abnormal expression of target genes [35–38].To investigate the upstream regulatory mRNAs of KIF14, several prediction programs, such as TargetScan, PicTar, miRanda, microT, miRmap, RNA22 and PITA, were used to assess the binding of miRNAs with KIF14. The identified miRNAs could act as tumor-suppressive molecules in cancer. For instance, hsa-miR-101-3p is involved in LINC00943-mediated gastric cancer cell proliferation and chemosensitivity [39]. Knocking down of hsa-miR-181c-5p depletes pediatric cancer stem-like cells and impairs tumor sphere formation [40]. Taking differential expression, correlation and survival analyses into consideration, hsa-miR-101-3p was identified as the most promising upstream binding tumor suppressive miRNA of KIF14.
Next, based on the ceRNA hypothesis [26], the upstream lncRNAs of hsa-miR-101-3p/KIF14 axis were predicted through the starBase database. Again, taking differential expression, correlation and survival analyses into consideration, GSEC and TYMSOS were identified as the most promising upstream upregulated lncRNAs. Previous studies have shown that GSEC and TYMSOS contribute the proliferation and migration of cancer cells. Matsumura and colleagues reported that GSEC could inhibit the function of DHX36 to promote colon cancer cell migration [41]. Liu and colleagues found that GSEC accelerated the proliferation and invasion of osteosarcoma via regulating miR-588/EIF5A2 axis [42]. TYMSOS was upregulated in gastric cancer cells and functioned as a ceRNA to post-transcriptionally regulate gastric cancer progression [43]. Taken together, GSEC and TYMSOS/hsa-miR-101-3p/KIF14 axis were identified as potential regulatory pathways in LUAD.
Immunotherapy is an emerging novel therapy for various cancers, including LUAD. TMB, MSI, and PD-1/PD-L1 have been proposed as potential biomarkers of immunotherapy in LUAD patients, but the results are still not satisfactory [5]. The discovery of accurate biomarker for immunotherapy remains a great challenge. Our results demonstrated that KIF14 was significantly correlated with TMB, MSI and immune checkpoint-related genes in various cancers. Impressively, KIF14 expression was positively associated with both TMB and MSI as well as CTLA4, PD-L1 and PD-1 in LUAD, which was further verified through the TIMER database. It has been reported that tumor ICI can influence the prognosis of patients with cancer. In this study, both TIMER database and biomarker analyses with R showed that KIF14 expression was positively related to CD8+ T cell and negatively associated with dendritic and B cell. These findings indicate that KIF14 expression may influence the tumor ICI of LUAD, thus highlighting its potential as an immunotherapy target.
Although our results improve our understanding of the correlation between KIF14 and LUAD, several limitations should be noted. Firstly, this study was primarily based on the molecular data downloaded from various public databases. Thus, further experimental studies are needed to verify our findings. Secondly, the function and mechanism of how KIF14 promotes the progression of LUAD should also be experimentally investigated.
Conclusions
In conclusion, KIF14 was upregulated in various cancers including LUAD, and contributed to unfavorable prognosis in LUAD patients. GSEC and TYMSOS/hsa-miR-101-3p/KIF14 ceRNA network was identified as a novel upstream regulatory mechanism of KIF14 in LUAD. Moreover, KIF14 exerted an oncogenic role via influencing tumor ICI and immune checkpoint-related gene expression. Nevertheless, further experimental studies are warranted to verify our findings in the near future.
Materials and Methods
Pan-cancer evaluation of KIF14 expression
The transcriptome data, miRNA data of pan-cancer combined with corresponding patients’ records were download from the TCGA database, and then analyzed with “limma” package in R version:3.6.3. P-values < 0.05 were deemed as statistically significant. Then, the mRNA levels of KIF14 in pan-cancers were analyzed using the Oncomine database (http://www.oncomine.org), with thresholds of p-value = 0.001 and fold change = 1.5.
Survival analysis of KIF14 in LUAD
The Kaplan-Meier (K-M) survival analysis was used to assess the differences between the high-expression group and low-expression groups based on median expression of KIF14 via the “survminer” and “survival” packages in R. Next, univariate and multivariate Cox regression analyses were conducted to explore the relationships among KIF14 expression, clinical prognostic indicators and survival time in LUAD patients, which was generated via the “survival” package in R with p-values < 0.05 regarded as statistical significance. The correlations between KIF14 and survival endpoints in LUAD was then confirmed using the PrognoScan database, with a fixed threshold of Cox p-value < 0.05.
Evaluation of the correlation between KIF14 expression and clinicopathological features
To evaluate the correlation between KIF14 expression level and clinicopathological features (e.g., TNM stage and age in diagnosis), Chi-squared tests were performed via using the “beeswarm” package in R.
Candidate miRNA prediction
Upstream binding miRNAs of KIF14 in LUAD were assessed using the starBase database (http://starbase.sysu.edu.cn) that involves TargetScan, PicTar, miRanda, microT, miRmap, RNA22 and PITA programs. Only the miRNAs identified from at least 2 of the above-mentioned programs were subjected to further analysis. The correlation analysis of miRNAs with KIF14 was performed according to their negative regulation of a target mRNA and those differentially expressed in tumor tissue than in normal tissues, which was carried out via the “limma”, “reshape2”, “ggpubr”, “ggExtra” packages in R based on TCGA-LUAD dataset. The survival analyses of miRNAs in LUAD were performed using the “survminer” and “survival” packages in R, with statistical significance set as log rank p-value < 0.05. Hsa-miR-101-3p showed highest correlation with KIF14, lowest expression in LUAD tissues and most favorable prognosis, which was selected as the candidate miRNA for KIF14.
Candidate lncRNA prediction and ceRNA network construction
StarBase database was also utilized to identify candidate lncRNAs bound to hsa-miR-101-3p, which included the correlation analysis of lncRNA-KIF14 with lncRNA-hsa-miR-101-3p in LUAD. Then, the correlation, expression and survival analyses of candidate lncRNAs were performed based on TCGA-LUAD dataset. Finally, the competing endogenous RNA (ceRNA) network of KIF14 was established via Cytoscape tool.
Evaluation of tumor immune infiltration
Tumor mutation burden (TMB) is defined as the total number of somatic mutations per megabase of a cancer genome [44]. Microsatellite instability (MSI) refers to the insertion or deletion of one or more repeat units in tumor cells [45]. Both TMB and MSI have been regarded as potential biomarkers for predicting responses to immunotherapy. The TMB and MSI values of each tumor in pan-caner were calculated, and their association with KIF14 expression were analyzed using the Spearman rank correlation coefficient. The radar charts were generated with “fmsb” package in R.
TIMER (http://timer.cistrome.org/), a website for the comprehensive analysis of ICI, was employed to determine the association between KIF14 expression and immune checkpoint-related gene expression or tumor ICI in LUAD. Level of statistical significance was set as p-value < 0.05.
Statistical analysis
The difference between normal and pan-cancer were calculated with “limma” package in R. For survival analyses, log rank test was employed for comparison. Univariate and multivariate analyses were carried out via Cox logistic regression model to identify independent clinicopathologic characteristics. The Chi-squared test was applied to evaluate the significant association between KIF14 expression and clinicopathological characteristics. The correlation of KIF14 expression with immune-related genes or tumor ICI was determined via the Spearman’s correlation analysis. A p-value < 0.05 was regarded as statistical significance.
Supplementary Materials
Author Contributions
(i) Conception and design: Yuejun Li; (ii) Administrative support: Yuejun Li; (iii) Provision of study materials or patients: Hong Wang, Fangting Tang, Ping Tang, Liang Zhang; (iv) Collection and assembly of data: Hong Wang, Fangting Tang, Ping Tang, Liang Zhang, Qixin Gan; (v) Data analysis and interpretation: All authors; (vi) Manuscript writing: All authors; (vii) Final approval of manuscript: All authors.
Acknowledgments
The authors would like to express their gratitude to EditSprings (https://www.editsprings.com/) for the expert linguistic services provided.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as potential conflicts of interest.
Ethical Statement
All of our data were directly acquired from open-access public databases, and this study was carried out strictly in accordance with the publishing guidelines provided by TCGA.
Funding
This study was supported by the National Natural Science Foundation of China [No. 81703916], Natural Science Foundation of Hunan Province [No. 2018JJ6042].
References
- 1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2020. CA Cancer J Clin. 2020; 70:7–30. https://doi.org/10.3322/caac.21590 [PubMed]
- 2. Imielinski M, Berger AH, Hammerman PS, Hernandez B, Pugh TJ, Hodis E, Cho J, Suh J, Capelletti M, Sivachenko A, Sougnez C, Auclair D, Lawrence MS, et al. Mapping the hallmarks of lung adenocarcinoma with massively parallel sequencing. Cell. 2012; 150:1107–20. https://doi.org/10.1016/j.cell.2012.08.029 [PubMed]
- 3. Thai AA, Solomon BJ, Sequist LV, Gainor JF, Heist RS. Lung cancer. Lancet. 2021; 398:535–54. https://doi.org/10.1016/S0140-6736(21)00312-3 [PubMed]
- 4. Yang CY, Yang JC, Yang PC. Precision Management of Advanced Non-Small Cell Lung Cancer. Annu Rev Med. 2020; 71:117–36. https://doi.org/10.1146/annurev-med-051718-013524 [PubMed]
- 5. Memmott RM, Wolfe AR, Carbone DP, Williams TM. Predictors of Response, Progression-Free Survival, and Overall Survival in Patients With Lung Cancer Treated With Immune Checkpoint Inhibitors. J Thorac Oncol. 2021; 16:1086–98. https://doi.org/10.1016/j.jtho.2021.03.017 [PubMed]
- 6. Xu Q, Chen Y. An Aging-Related Gene Signature-Based Model for Risk Stratification and Prognosis Prediction in Lung Adenocarcinoma. Front Cell Dev Biol. 2021; 9:685379. https://doi.org/10.3389/fcell.2021.685379 [PubMed]
- 7. Herbst RS, Giaccone G, de Marinis F, Reinmuth N, Vergnenegre A, Barrios CH, Morise M, Felip E, Andric Z, Geater S, Özgüroğlu M, Zou W, Sandler A, et al. Atezolizumab for First-Line Treatment of PD-L1-Selected Patients with NSCLC. N Engl J Med. 2020; 383:1328–39. https://doi.org/10.1056/NEJMoa1917346 [PubMed]
- 8. Powles T, van der Heijden MS, Castellano D, Galsky MD, Loriot Y, Petrylak DP, Ogawa O, Park SH, Lee JL, De Giorgi U, Bögemann M, Bamias A, Eigl BJ, et al, and DANUBE study investigators. Durvalumab alone and durvalumab plus tremelimumab versus chemotherapy in previously untreated patients with unresectable, locally advanced or metastatic urothelial carcinoma (DANUBE): a randomised, open-label, multicentre, phase 3 trial. Lancet Oncol. 2020; 21:1574–88. https://doi.org/10.1016/S1470-2045(20)30541-6 [PubMed]
- 9. Sezer A, Kilickap S, Gümüş M, Bondarenko I, Özgüroğlu M, Gogishvili M, Turk HM, Cicin I, Bentsion D, Gladkov O, Clingan P, Sriuranpong V, Rizvi N, et al. Cemiplimab monotherapy for first-line treatment of advanced non-small-cell lung cancer with PD-L1 of at least 50%: a multicentre, open-label, global, phase 3, randomised, controlled trial. Lancet. 2021; 397:592–604. https://doi.org/10.1016/S0140-6736(21)00228-2 [PubMed]
- 10. Grant MJ, Herbst RS, Goldberg SB. Selecting the optimal immunotherapy regimen in driver-negative metastatic NSCLC. Nat Rev Clin Oncol. 2021; 18:625–44. https://doi.org/10.1038/s41571-021-00520-1 [PubMed]
- 11. Benoit MPMH, Asenjo AB, Paydar M, Dhakal S, Kwok BH, Sosa H. Structural basis of mechano-chemical coupling by the mitotic kinesin KIF14. Nat Commun. 2021; 12:3637. https://doi.org/10.1038/s41467-021-23581-3 [PubMed]
- 12. Chew YM, Cross RA. Molecular Motors: Kif14's Disordered Dongle. Curr Biol. 2020; 30:R988–91. https://doi.org/10.1016/j.cub.2020.06.095 [PubMed]
- 13. Pejskova P, Reilly ML, Bino L, Bernatik O, Dolanska L, Ganji RS, Zdrahal Z, Benmerah A, Cajanek L. KIF14 controls ciliogenesis via regulation of Aurora A and is important for Hedgehog signaling. J Cell Biol. 2020; 219:e201904107. https://doi.org/10.1083/jcb.201904107 [PubMed]
- 14. Zhernov I, Diez S, Braun M, Lansky Z. Intrinsically Disordered Domain of Kinesin-3 Kif14 Enables Unique Functional Diversity. Curr Biol. 2020; 30:3342–51.e5. https://doi.org/10.1016/j.cub.2020.06.039 [PubMed]
- 15. Xu H, Choe C, Shin SH, Park SW, Kim HS, Jung SH, Yim SH, Kim TM, Chung YJ. Silencing of KIF14 interferes with cell cycle progression and cytokinesis by blocking the p27(Kip1) ubiquitination pathway in hepatocellular carcinoma. Exp Mol Med. 2014; 46:e97. https://doi.org/10.1038/emm.2014.23 [PubMed]
- 16. Klimaszewska-Wiśniewska A, Neska-Długosz I, Buchholz K, Durślewicz J, Grzanka D, Kasperska A, Antosik P, Zabrzyński J, Grzanka A, Gagat M. Prognostic Significance of KIF11 and KIF14 Expression in Pancreatic Adenocarcinoma. Cancers (Basel). 2021; 13:3017. https://doi.org/10.3390/cancers13123017 [PubMed]
- 17. Cheng C, Wu X, Shen Y, Li Q. KIF14 and KIF23 Promote Cell Proliferation and Chemoresistance in HCC Cells, and Predict Worse Prognosis of Patients with HCC. Cancer Manag Res. 2020; 12:13241–57. https://doi.org/10.2147/CMAR.S285367 [PubMed]
- 18. Liu J, Li D, Zhang X, Li Y, Ou J. Histone Demethylase KDM3A Promotes Cervical Cancer Malignancy Through the ETS1/KIF14/Hedgehog Axis. Onco Targets Ther. 2020; 13:11957–73. https://doi.org/10.2147/OTT.S276559 [PubMed]
- 19. Thériault BL, Pajovic S, Bernardini MQ, Shaw PA, Gallie BL. Kinesin family member 14: an independent prognostic marker and potential therapeutic target for ovarian cancer. Int J Cancer. 2012; 130:1844–54. https://doi.org/10.1002/ijc.26189 [PubMed]
- 20. Ahmed SM, Thériault BL, Uppalapati M, Chiu CW, Gallie BL, Sidhu SS, Angers S. KIF14 negatively regulates Rap1a-Radil signaling during breast cancer progression. J Cell Biol. 2012; 199:951–67. https://doi.org/10.1083/jcb.201206051 [PubMed]
- 21. Basavarajappa HD, Corson TW. KIF14 as an oncogene in retinoblastoma: a target for novel therapeutics? Future Med Chem. 2012; 4:2149–52. https://doi.org/10.4155/fmc.12.158 [PubMed]
- 22. Mizuno H, Kitada K, Nakai K, Sarai A. PrognoScan: a new database for meta-analysis of the prognostic value of genes. BMC Med Genomics. 2009; 2:18. https://doi.org/10.1186/1755-8794-2-18 [PubMed]
- 23. Madhavan S, Zenklusen JC, Kotliarov Y, Sahni H, Fine HA, Buetow K. Rembrandt: helping personalized medicine become a reality through integrative translational research. Mol Cancer Res. 2009; 7:157–67. https://doi.org/10.1158/1541-7786.MCR-08-0435 [PubMed]
- 24. Li JH, Liu S, Zhou H, Qu LH, Yang JH. starBase v2.0: decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014; 42:D92–7. https://doi.org/10.1093/nar/gkt1248 [PubMed]
- 25. Otasek D, Morris JH, Bouças J, Pico AR, Demchak B. Cytoscape Automation: empowering workflow-based network analysis. Genome Biol. 2019; 20:185. https://doi.org/10.1186/s13059-019-1758-4 [PubMed]
- 26. Salmena L, Poliseno L, Tay Y, Kats L, Pandolfi PP. A ceRNA hypothesis: the Rosetta Stone of a hidden RNA language? Cell. 2011; 146:353–8. https://doi.org/10.1016/j.cell.2011.07.014 [PubMed]
- 27. Liu M, Li J, Huang Z, Li Y. Gastric cancer risk-scoring system based on analysis of a competing endogenous RNA network. Transl Cancer Res. 2020; 9:3889–902. https://doi.org/10.21037/tcr-19-2977 [PubMed]
- 28. Kornepati AVR, Vadlamudi RK, Curiel TJ. Programmed death ligand 1 signals in cancer cells. Nat Rev Cancer. 2022; 22:174–89. https://doi.org/10.1038/s41568-021-00431-4 [PubMed]
- 29. Xia X, Tang P, Liu H, Li Y. Identification and Validation of an Immune-related Prognostic Signature for Hepatocellular Carcinoma. J Clin Transl Hepatol. 2021; 9:798–808. https://doi.org/10.14218/JCTH.2021.00017 [PubMed]
- 30. Yi M, Zheng X, Niu M, Zhu S, Ge H, Wu K. Combination strategies with PD-1/PD-L1 blockade: current advances and future directions. Mol Cancer. 2022; 21:28. https://doi.org/10.1186/s12943-021-01489-2 [PubMed]
- 31. Li T, Fan J, Wang B, Traugh N, Chen Q, Liu JS, Li B, Liu XS. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017; 77:e108–10. https://doi.org/10.1158/0008-5472.CAN-17-0307 [PubMed]
- 32. Corson TW, Gallie BL. KIF14 mRNA expression is a predictor of grade and outcome in breast cancer. Int J Cancer. 2006; 119:1088–94. https://doi.org/10.1002/ijc.21954 [PubMed]
- 33. Corson TW, Zhu CQ, Lau SK, Shepherd FA, Tsao MS, Gallie BL. KIF14 messenger RNA expression is independently prognostic for outcome in lung cancer. Clin Cancer Res. 2007; 13:3229–34. https://doi.org/10.1158/1078-0432.CCR-07-0393 [PubMed]
- 34. Wang Q, Wang L, Li D, Deng J, Zhao Z, He S, Zhang Y, Tu Y. Kinesin family member 14 is a candidate prognostic marker for outcome of glioma patients. Cancer Epidemiol. 2013; 37:79–84. https://doi.org/10.1016/j.canep.2012.08.011 [PubMed]
- 35. Goodall GJ, Wickramasinghe VO. RNA in cancer. Nat Rev Cancer. 2021; 21:22–36. https://doi.org/10.1038/s41568-020-00306-0 [PubMed]
- 36. Kalogirou C, Linxweiler J, Schmucker P, Snaebjornsson MT, Schmitz W, Wach S, Krebs M, Hartmann E, Puhr M, Müller A, Spahn M, Seitz AK, Frank T, et al. MiR-205-driven downregulation of cholesterol biosynthesis through SQLE-inhibition identifies therapeutic vulnerability in aggressive prostate cancer. Nat Commun. 2021; 12:5066. https://doi.org/10.1038/s41467-021-25325-9 [PubMed]
- 37. Lou W, Wang W, Chen J, Wang S, Huang Y. ncRNAs-mediated high expression of SEMA3F correlates with poor prognosis and tumor immune infiltration of hepatocellular carcinoma. Mol Ther Nucleic Acids. 2021; 24:845–55. https://doi.org/10.1016/j.omtn.2021.03.014 [PubMed]
- 38. Wu Q, Luo X, Terp MG, Li Q, Li Y, Shen L, Chen Y, Jacobsen K, Bivona TG, Chen H, Zeng R, Ditzel HJ. DDX56 modulates post-transcriptional Wnt signaling through miRNAs and is associated with early recurrence in squamous cell lung carcinoma. Mol Cancer. 2021; 20:108. https://doi.org/10.1186/s12943-021-01403-w [PubMed]
- 39. Xu Y, Ji T, An N, Wang X, Zhang H, Xu F. LINC00943 is correlated with gastric cancer and regulates cancer cell proliferation and chemosensitivity via hsa-miR-101-3p. Int J Clin Oncol. 2021; 26:1650–60. https://doi.org/10.1007/s10147-021-01945-5 [PubMed]
- 40. Sanchez-Diaz PC, Hsiao TH, Chang JC, Yue D, Tan MC, Chen HI, Tomlinson GE, Huang Y, Chen Y, Hung JY. De-regulated microRNAs in pediatric cancer stem cells target pathways involved in cell proliferation, cell cycle and development. PLoS One. 2013; 8:e61622. https://doi.org/10.1371/journal.pone.0061622 [PubMed]
- 41. Matsumura K, Kawasaki Y, Miyamoto M, Kamoshida Y, Nakamura J, Negishi L, Suda S, Akiyama T. The novel G-quadruplex-containing long non-coding RNA GSEC antagonizes DHX36 and modulates colon cancer cell migration. Oncogene. 2017; 36:1191–9. https://doi.org/10.1038/onc.2016.282 [PubMed]
- 42. Liu R, Ju C, Zhang F, Tang X, Yan J, Sun J, Lv B, Guo Y, Liang Y, Lv XB, Zhang Z. LncRNA GSEC promotes the proliferation, migration and invasion by sponging miR-588/ EIF5A2 axis in osteosarcoma. Biochem Biophys Res Commun. 2020; 532:300–7. https://doi.org/10.1016/j.bbrc.2020.08.056 [PubMed]
- 43. Gu Y, Wan C, Zhou G, Zhu J, Shi Z, Zhuang Z. TYMSOS drives the proliferation, migration, and invasion of gastric cancer cells by regulating ZNF703 via sponging miR-4739. Cell Biol Int. 2021; 45:1710–9. https://doi.org/10.1002/cbin.11610 [PubMed]
- 44. Campbell BB, Light N, Fabrizio D, Zatzman M, Fuligni F, de Borja R, Davidson S, Edwards M, Elvin JA, Hodel KP, Zahurancik WJ, Suo Z, Lipman T, et al. Comprehensive Analysis of Hypermutation in Human Cancer. Cell. 2017; 171:1042–56.e10. https://doi.org/10.1016/j.cell.2017.09.048 [PubMed]
- 45. Cortes-Ciriano I, Lee S, Park WY, Kim TM, Park PJ. A molecular portrait of microsatellite instability across multiple cancers. Nat Commun. 2017; 8:15180. https://doi.org/10.1038/ncomms15180 [PubMed]