




Introduction
Alcohol is widely consumed in the United States, with 86% and 27% of individuals 18 years of age or older having ever consumed alcohol and having binged alcohol in the past month, respectively [1]. The effects of alcohol on health are complex, with moderate consumption associated with lower risks of diabetes and ischemic heart disease, although the risk of these diseases increase with greater consumption [2]. In comparison, the risk of cancer, including oral, liver, female breast, and colorectal, monotonically increases with greater alcohol consumption [2, 3]. In addition to the amount of alcohol consumed, the type of alcohol has demonstrated different effects on health outcomes. For example, a meta-analysis observed increased risk of lung cancer with greater consumption of beer and liquor, whereas modest consumption of wine was associated with lower risk [4]. Additionally, alcohol consumption has been shown to differ by demographic characteristics, with declining consumption with increasing age and higher consumption among males and self-reported White individuals [5, 6]. While previous studies identified associations between alcohol and age-related health outcomes, studies investigating the effect, type, and pattern of alcohol consumption, in conjunction with demographic characteristics, on the aging process at a molecular level remain limited. Furthermore, a better understanding of the molecular and biological processes associated with alcohol consumption, both chronically and acutely, may improve our understanding of its complex relationship with health and potentially provide new modalities to screen for and prevent age- and alcohol-related chronic diseases.
Molecular markers of biological aging have provided insights into the aging process and age-related conditions. As an indicator of biological age estimated from DNA methylation levels, epigenetic age captures age-related changes to the epigenome and is highly correlated with chronological age [7]. Furthermore, the divergence between epigenetic age and chronological age is used to estimate epigenetic age acceleration (EAA), with positive values indicating an older epigenetic age relative to chronological age and vice versa. Numerous epigenetic metrics of biological age have been developed, including those by Horvath (intrinsic epigenetic age), Hannum (extrinsic epigenetic age), Levine (PhenoAge), and Lu (GrimAge), and have been associated with lifestyle factors, age-related diseases, physical functioning, and lifespan [7–10].
Studies have previously identified associations between alcohol consumption and several EAA metrics [11–16]. In addition, alcohol use disorders have been associated with epigenetic aging in blood and liver tissues, suggesting potential alcohol induced tissue-specific age related changes [17, 18]. Despite these findings, few studies have examined both cumulative and binged alcohol consumption, and the type of alcohol consumed, on epigenetic age-related changes. Therefore, we investigated the associations between cumulative and binged alcohol consumption, as well as alcohol type, on four measures of EAA in the Coronary Artery Risk Development in Young Adults (CARDIA) Study.
Results
Sample characteristics
Characteristics of study participants who underwent DNA methylation profiling at examination year (Y) 15 and Y20 were previously found to be largely similar to those participants who did not undergo DNA methylation profiling [19]. Table 1 presents descriptive characteristics for the 1,030 and 945 participants who underwent DNA methylation profiling by binge drinking status at Y15 and Y20, respectively. Among participants who drink, those who recently binge drank had an average of 24.6 ± 22.6 and 31.3 ± 27.5 total alcohol years at Y15 and Y20, compared to an average of 7.1 ± 8.4 and 10.8 ± 17.0 total alcohol years among those who did not recently binge drink, respectively. At both examination years, participants who recently binge drank exhibited a higher GAA compared to non-drinkers and participants who did not recently binge drink (P < 0.001), while the other EAA metrics did not show a similar trend.
Table 1. Descriptive statistics of study participants at examination years 15 and 20.
| N | Year 15 | Year 20 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Non-drinkers | No recent binge | Recent binge | P | Non-drinkers | No recent binge | Recent binge | P | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 178 | 592 | 260 | 155 | 566 | 224 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Female, n | 112 (62.9%) | 336 (56.8%) | 78 (30.0%) | <0.001 | 95 (61.3%) | 308 (45.6%) | 80 (35.7%) | <0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Race, n | 0.003 | 0.006 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Black | 91 (51.1%) | 235 (39.7%) | 91 (35.0%) | 81 (52.3%) | 219 (38.7%) | 85 (38.0%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| White | 87 (48.9%) | 357 60.3%) | 169 (65.0%) | 74 (47.7%) | 347 (61.3%) | 139 (62.0%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Chronological Age, years | 40.1 (3.8) | 40.7 (3.5) | 40.0 (3.3) | 0.019 | 45.2 (3.6) | 45.6 (3.6) | 45.0 (3.3) | 0.067 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IEAA, years | 0.1 (4.2) | 0.1 (4.2) | –0.2 (4.6) | 0.656 | 0.6 (4.5) | –0.1 (4.6) | –0.2 (3.9) | 0.220 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EEAA, years | –0.1 (5.2) | –0.1 (5.2) | 0.1 (5.2) | 0.893 | 0.7 (5.2) | –0.1 (5.1) | –0.1 (4.8) | 0.160 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PAA, years | –0.5 (5.8) | 0.0 (6.0) | 0.3 (6.3) | 0.437 | 0.4 (6.0) | –0.2 (6.0) | 0.3 (6.2) | 0.406 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GAA, years | –1.3 (3.8) | –0.3 (4.4) | 1.4 (5.0) | <0.001 | –1.4 (4.0) | –0.3 (4.4) | 1.6 (4.8) | <0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Education, years | 15.0 (2.3) | 15.2 (2.6) | 14.9 (2.6) | 0.304 | 14.8 (2.4) | 15.2 (2.5) | 14.8 (2.5) | 0.091 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Center, n | <0.001 | 0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Birmingham, AL | 68 (38.2%) | 134 (22.6%) | 50 (19.2%) | 56 (36.1%) | 118 (20.8%) | 44 (19.6%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Chicago, IL | 30 (16.9%) | 127 (21.5%) | 66 (25.4%) | 28 (18.1%) | 118 (20.8%) | 60 (26.8%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Minneapolis, MN | 38 (21.3%) | 157 (26.5%) | 81 (31.2%) | 37 (23.9%) | 163 (28.9%) | 56 (25.0%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Oakland, CA | 42 (23.6%) | 174 (29.4%) | 63 (24.2%) | 34 (21.9%) | 167 (29.5%) | 64 (28.6%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Pack years of smoking, year | 1.5 (4.7) | 4.0 (7.7) | 6.6 (9.9) | <0.001 | 1.7 (5.0) | 4.8 (9.1) | 6.6 (9.3) | <0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Physical activity, intensity score | 278.1 (246.9) | 328.3 (253.3) | 449.0 (311.1) | <0.001 | 288.7 (264.6) | 337.4 (267.6) | 423.8 (289.1) | <0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BMI, kg/m2 | 29.6 (6.8) | 28.4 (6.6) | 28.1 (4.9) | 0.031 | 30.7 (7.1) | 29.2 (6.7) | 28.6 (5.2) | 0.008 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Alcohol years, years | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Beer | 0 (0) | 3.6 (6.1) | 15.5 (17.4) | <0.001 | 0 (0) | 5.6 (13.2) | 17.6 (18.9) | <0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Liquor | 0 (0) | 1.4 (2.9) | 5.2 (8.9) | <0.001 | 0 (0) | 2.2 (5.5) | 6.9 (11.0) | <0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Wine | 0 (0) | 2.2 (3.8) | 3.9 (6.6) | <0.001 | 0 (0) | 3.0 (5.0) | 6.8 (11.3) | <0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Total Alcohol | 0 (0) | 7.1 (8.4) | 24.6 (22.6) | <0.001 | 0 (0) | 10.8 (17.0) | 31.3 (27.5) | <0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Binge in last 30 days, days | 0 (0) | 0 (0) | 4.1 (5.5) | <0.001 | 0 (0) | 0 (0) | 3.7 (4.8) | <0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| All statistics shown are mean and standard deviation, except for sex, race, and center, which are shown as number of participants and percentages. Abbreviations: IEAA: intrinsic epigenetic age acceleration; EEAA: extrinsic epigenetic age acceleration; PAA: PhenoAge acceleration; GAA: GrimAge acceleration; BMI: body mass index. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Cumulative alcohol consumption on epigenetic age acceleration
Table 2 presents results for the associations between the cumulative alcohol consumption variables and EAA. After adjusting for covariates, beer years (P = 0.035 and P = 0.001) and liquor years (P = 0.003 and P < 0.001) were positively associated with GAA at Y15 and Y20, respectively. Each additional five beer years was associated with a 0.12-year [95% CI: 0.01, 0.22] and a 0.15-year [95% CI: 0.06, 0.24] higher GAA at Y15 and Y20, respectively. Each additional five liquor years was associated with a 0.32-year [95% CI: 0.11, 0.54] and a 0.33-year [95% CI: 0.16, 0.50] higher GAA at Y15 and Y20, respectively. Total alcohol years were positively associated with GAA (P = 0.005 and P < 0.001), with a 0.11-year [95% CI: 0.03, 0.19] and 0.13-year [95% CI: 0.07, 0.19] higher GAA per five alcohol years at Y15 and Y20, respectively. Wine years were not associated with GAA at either Y15 or Y20. Findings from GEE analyses yielded similar results as Y15 and Y20. IEAA (intrinsic epigenetic age acceleration), EEAA (extrinsic epigenetic age acceleration), and PAA (PhenoAge acceleration) were not associated with the cumulative alcohol consumption variables. Restricting to study participants with complete follow up yielded consistent conclusions (Supplementary Table 1). We observed similar findings when alcohol consumption was categorized into non-drinkers, low, intermediate, and high alcohol consumption (Supplementary Tables 2–4). Similar results as total alcohol years were observed using the cumulative amount of absolute alcohol consumed (Supplementary Table 5). When analyzing weekly alcohol consumption, only liquor consumption was positively associated with GAA (Supplementary Table 6). We observed correlations between beer, liquor, and total alcohol years and several GrimAge surrogate biomarkers of blood plasma proteins at Y15 and Y20, including adrenomedullin, leptin, and plasminogen activation inhibitor 1 (PAI-1) (Supplementary Figures 1, 2). Telomere length estimates were derived from DNA methylation and no cumulative alcohol variable was associated with telomere length (Supplementary Table 7).
Table 2. Analysis results for the association between cumulative alcohol consumption and EAA at examination years 15 and 20.
| Year 15 | Year 20 | GEE | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| β [95% CI] | P | β [95% CI] | P | β [95% CI] | P | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Beer Years | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IEAA | –0.04 [–0.17, 0.09] | 0.526 | –0.08 [–0.19, 0.02] | 0.129 | –0.07 [–0.17, 0.04] | 0.216 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EEAA | 0.01 [–0.15, 0.16] | 0.909 | –0.03 [–0.15, 0.09] | 0.600 | –0.02 [–0.13, 0.09] | 0.758 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PAA | 0.11 [–0.07, 0.29] | 0.241 | 0.05 [–0.10, 0.19] | 0.516 | 0.07 [–0.05, 0.19] | 0.275 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GAA | 0.12 [0.01, 0.22] | 0.035 | 0.15 [0.06, 0.24] | 0.001 | 0.13 [–0.01, 0.27] | 0.075 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Liquor Years | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IEAA | 0.12 [–0.13, 0.38] | 0.344 | –0.11 [–0.32, 0.10] | 0.306 | –0.02 [–0.22, 0.18] | 0.832 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EEAA | –0.13 [–0.44, 0.17] | 0.396 | –0.08 [–0.30, 0.15] | 0.512 | –0.11 [–0.29, 0.07] | 0.247 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PAA | 0.19 [–0.16, 0.55] | 0.291 | 0.13 [–0.15, 0.41] | 0.372 | 0.14 [–0.13, 0.41] | 0.313 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GAA | 0.32 [0.11, 0.54] | 0.003 | 0.33 [0.16, 0.50] | <0.001 | 0.31 [0.11, 0.51] | 0.002 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Wine Years | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IEAA | 0.17 [–0.13, 0.48] | 0.265 | 0.03 [–0.19, 0.24] | 0.813 | 0.08 [–0.08, 0.23] | 0.328 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EEAA | –0.01 [–0.38, 0.35] | 0.952 | –0.06 [–0.29, 0.17] | 0.610 | –0.05 [–0.26, 0.16] | 0.645 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PAA | 0.18 [–0.24, 0.61] | 0.405 | 0.02 [–0.27, 0.31] | 0.892 | 0.07 [–0.22, 0.35] | 0.649 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GAA | 0.08 [–0.18, 0.33] | 0.546 | 0.11 [–0.07, 0.28] | 0.228 | 0.09 [–0.11, 0.30] | 0.359 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Total Alcohol Years | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IEAA | 0.01 [–0.08, 0.10] | 0.820 | –0.05 [–0.12, 0.02] | 0.182 | –0.03 [–0.10, 0.04] | 0.436 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EEAA | –0.01 [–0.13, 0.10] | 0.806 | –0.03 [–0.11, 0.05] | 0.438 | –0.03 [–0.10, 0.05] | 0.455 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PAA | 0.10 [–0.03, 0.23] | 0.138 | 0.04 [–0.06, 0.14] | 0.414 | 0.06 [–0.03, 0.15] | 0.202 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GAA | 0.11 [0.03, 0.19] | 0.005 | 0.13 [0.07, 0.19] | <0.001 | 0.12 [0.02, 0.21] | 0.013 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Results are adjusted for chronological age, sex, race, center, education, pack years of smoking, BMI, and physical activity. Beta coefficients represent the gain in EAA for each additional 5 alcohol years. Abbreviations: IEAA: intrinsic epigenetic age acceleration; EEAA: extrinsic epigenetic age acceleration; PAA: PhenoAge acceleration; GAA: GrimAge acceleration. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Figure 1 displays quantile regression plots for the cumulative alcohol consumption variables at Y15 and Y20 on GAA. We plotted regression estimates for 19 quantiles ranging from 0.05 to 0.95. As displayed in the plots, beer years displayed an increasing effect on GAA across the distribution at Y15 with almost a three and a half times greater effect in the upper conditional quartiles compared to the lower conditional quartiles (0.31-year vs. 0.09-year, respectively; Figure 1A), with similar effects at Y20 (0.34-year vs. 0.10-year, respectively; Figure 1B). Liquor years demonstrated a thirty-four times greater effect in the upper conditional quartiles compared to the lower conditional quartiles at Y15 (0.68-year vs. 0.02-year, respectively; Figure 1C) and five times greater effect in the upper conditional quartiles compared to the lower conditional quartiles at Y20 (0.54-year vs. 0.11-year, respectively; Figure 1D). Wine years displayed a relatively negative effect on GAA in the lower conditional quartiles with a positive effect in the upper conditional quartiles at both Y15 and Y20. Notably, the effect estimate of wine years can be six and a half times greater in the upper conditional quartiles compared to the lower conditional quartiles of the distribution at Y15 (0.20-year gain vs. 0.03-year loss, respectively; Figure 1E) and nine times greater effect in the upper conditional quartiles compared to the lower conditional quartiles at Y20 (0.36-year gain vs. 0.04-year loss, respectively; Figure 1F). Total alcohol years demonstrated a four and a half times greater effect in the upper conditional quartiles of the distribution compared to the lower conditional quartiles (0.27-year vs. 0.06-year, respectively; Figure 1G) at Y15 and an approximately six times greater effect in the upper conditional quartiles compared to the lower conditional quartiles (0.25-year vs. 0.04-year, respectively; Figure 1H) at Y20.
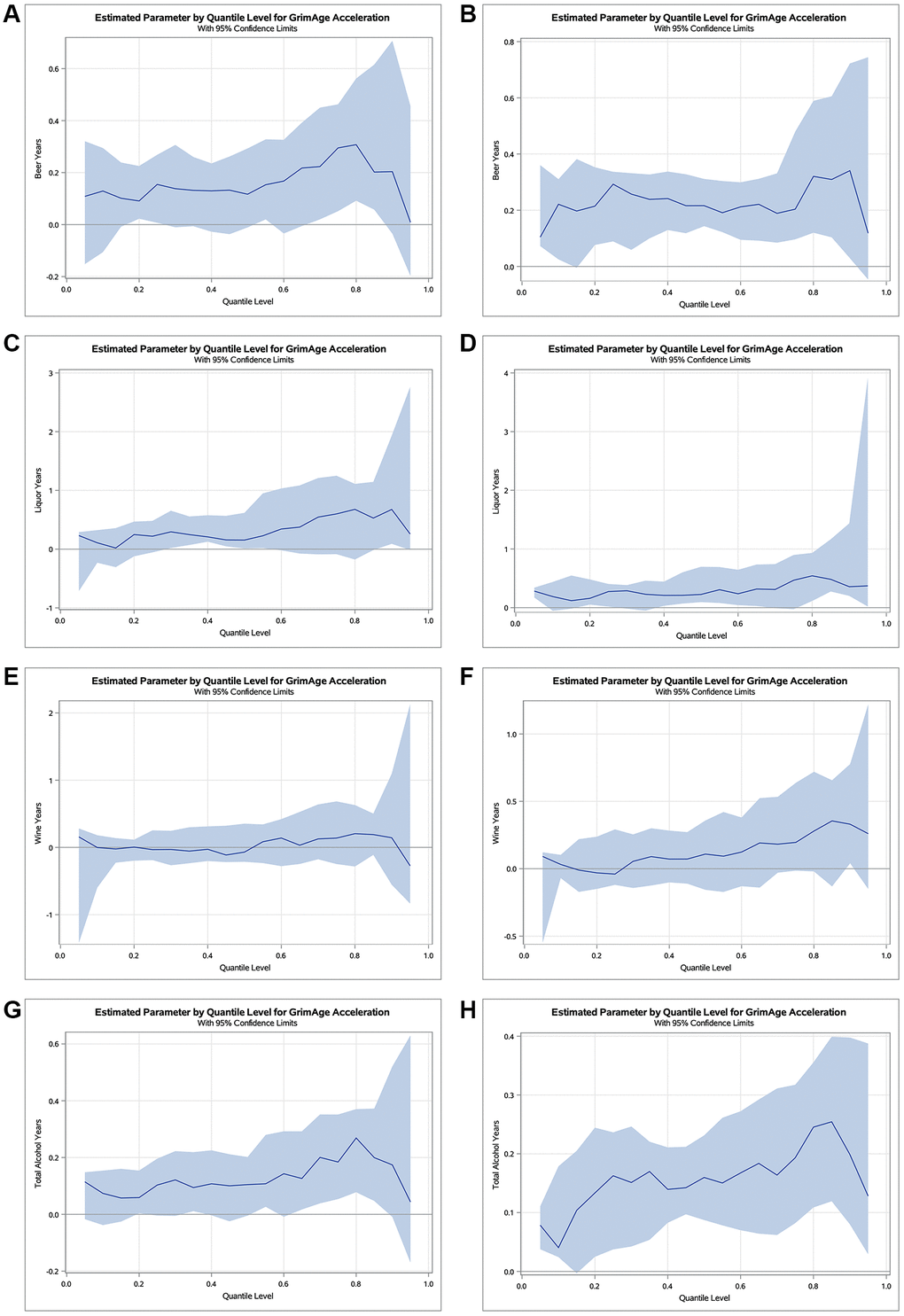
Figure 1. Estimated parameters by quantile with 95% confidence limits for the effect of cumulative alcohol consumption on GAA at examination years 15 and 20. Quantile regression plots for beer (A, B), liquor (C, D), wine (E, F), and total alcohol (G, H) years at Y15 and Y20, respectively. The x-axis represents the quantile scale, and the y-axis represents the effect of alcohol on GAA for a given quantile. Results are adjusted for chronological age, sex, race, center, education, pack years of smoking, BMI, and physical activity.
Recent binge drinking on epigenetic age acceleration
Table 3 presents results for the associations between binge drinking and EAA. Recent binge drinking was positively associated with GAA at Y15 (P < 0.001) and Y20 (P < 0.001). Compared to participants who did not binge drink in the past 30 days, those who did exhibited a 1.22-year [95% CI: 0.69, 1.76] and a 1.55-year [95% CI: 0.99, 2.11] higher GAA at Y15 and Y20, respectively. Additionally, the number of days of binge drinking was associated with GAA at Y15 (P = 0.002) and Y20 (P < 0.001). Each additional day of binge drinking was associated with a 0.11-year [95% CI: 0.04, 0.18] and a 0.21-year [95% CI: 0.12, 0.29] higher GAA at Y15 and Y20, respectively. GEE yielded comparable associations as Y15 and Y20. IEAA, EEAA, and PAA were not associated with either binge drinking variable. Restricting to study participants with complete follow up yielded consistent conclusions (Supplementary Table 8). We observed similar correlations between days of recent binge drinking and several GrimAge surrogate biomarkers of blood plasma proteins at Y15 and Y20 (Supplementary Figures 1, 2). When modeling both cumulative alcohol consumption and recent binge drinking during GAA analyses, we observed recent binge drinking remained associated with GAA, while the cumulative alcohol consumption variables became less significant, although liquor years remained associated with GAA (Supplementary Table 9). Recent binge drinking was associated with telomere length derived from DNA methylation (Supplementary Table 7).
Table 3. Analysis results for the association between recent binge drinking and EAA at examination years 15 and 20.
| Year 15 | Year 20 | GEE | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| β [95% CI] | P | β [95% CI] | P | β [95% CI] | P | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Recent Binge | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IEAA | –0.33 [–0.97, 0.32] | 0.320 | –0.30 [–0.99, 0.38] | 0.386 | –0.32 [–0.80, 0.16] | 0.195 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EEAA | –0.31 [–1.08, 0.46] | 0.429 | –0.60 [–1.36, 0.15] | 0.115 | –0.45 [–1.02, 0.13] | 0.126 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PAA | 0.22 [–0.67, 1.12] | 0.624 | 0.27 [–0.65, 1.20] | 0.563 | 0.24 [–0.44, 0.93] | 0.486 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GAA | 1.22 [0.69, 1.76] | <0.001 | 1.55 [0.99, 2.11] | <0.001 | 1.38 [0.92, 1.83] | <0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Recent Binge Quantity | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IEAA | –0.04 [–0.12, 0.05] | 0.389 | 0.00 [–0.10, 0.10] | 0.983 | –0.02 [–0.12, 0.08] | 0.654 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EEAA | –0.06 [–0.16, 0.04] | 0.244 | 0.03 [–0.08, 0.15] | 0.575 | –0.02 [–0.10, 0.06] | 0.635 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PAA | –0.04 [–0.16, 0.07] | 0.475 | 0.14 [0.00, 0.28] | 0.055 | 0.03 [–0.07, 0.13] | 0.579 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GAA | 0.11 [0.04, 0.18] | 0.002 | 0.21 [0.12, 0.29] | <0.001 | 0.15 [0.07, 0.23] | <0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Results are adjusted for chronological age, sex, race, center, education, pack years of smoking, BMI, and physical activity. Beta coefficients for recent binge represents the gain in EAA for participants who binge drank in the past 30 days and the beta coefficients for recent binge quantity represents the gain in EAA for each additional day of binge drinking in the past 30 days. Abbreviations: IEAA: intrinsic epigenetic age acceleration; EEAA: extrinsic epigenetic age acceleration; PAA: PhenoAge acceleration; GAA: GrimAge acceleration. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Cumulative alcohol consumption and chronological age interaction on GrimAge acceleration
Table 4 presents the interaction and stratified results for the joint associations of cumulative alcohol consumption and chronological age on GAA. Summary statistics by age quartiles are presented in Supplementary Table 10. At Y15, we observed a 0.35-year [95% CI: 0.14, 0.57] higher GAA per 5 beer years among participants in quartile 1 compared to a 0.05-year [95% CI: –0.30, 0.19] loss in GAA among those in quartile 4 (Pinteraction < 0.001). Additionally, participants in quartile 1 displayed a 0.26-year [95% CI: 0.11, 0.41] higher GAA per 5 total alcohol years compared to participants in quartile 4 who experienced a 0.06-year [95% CI: –0.11, 0.22] higher GAA (Pinteraction = 0.002). No statistical interactions were observed between liquor and wine years with chronological age.
Table 4. Interaction and stratified analysis results for the association between cumulative alcohol consumption and GAA at examination years 15 and 20 by strata of chronological age quartiles.
| Year 15 | Year 20 | GEE | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Βalcohol [95% CI] | P | Βalcohol [95% CI] | P | Βalcohol [95% CI] | P | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Beer Years | –0.05 [–0.07, –0.02] | <0.001* | –0.04 [–0.06, –0.03] | <0.001* | –0.03 [–0.05, –0.02] | <0.001* | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 1 | 0.35 [0.14, 0.57] | 0.002 | 0.29 [0.12, 0.46] | <0.001 | 0.31 [0.16, 0.46] | <0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 2 | 0.30 [0.10, 0.50] | 0.004 | 0.36 [0.15, 0.57] | <0.001 | 0.32 [0.15, 0.49] | <0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 3 | –0.07 [–0.28, 0.13] | 0.488 | 0.29 [0.13, 0.45] | <0.001 | 0.13 [–0.03, 0.28] | 0.119 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 4 | –0.05 [–0.30, 0.19] | 0.660 | –0.17 [–0.34, –0.01] | 0.043 | –0.16 [–0.34, 0.03] | 0.094 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Liquor Years | –0.03 [–0.08, 0.02] | 0.297 | –0.01 [–0.05, 0.04] | 0.766 | –0.02 [–0.06, 0.03] | 0.500 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 1 | 0.50 [0.14, 0.87] | 0.007 | 0.22 [–0.05, 0.50] | 0.115 | 0.32 [–0.01, 0.65] | 0.060 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 2 | 0.18 [–0.29, 0.66] | 0.440 | 0.21 [–0.25, 0.67] | 0.363 | 0.18 [–0.25, 0.60] | 0.416 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 3 | 0.00 [–0.51, 0.51] | 0.999 | 0.43 [0.08, 0.78] | 0.017 | 0.23 [–0.19, 0.65] | 0.278 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 4 | 0.39 [–0.01, 0.79] | 0.055 | 0.26 [–0.11, 0.63] | 0.164 | 0.31 [–0.07, 0.70] | 0.114 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Wine Years | –0.04 [–0.12, 0.04] | 0.312 | –0.07 [–0.13, –0.01] | 0.015* | –0.04 [–0.09, 0.01] | 0.154 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 1 | 0.46 [–0.44, 1.37] | 0.317 | 0.64 [–0.07, 1.34] | 0.077 | 0.54 [–0.08, 1.16] | 0.090 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 2 | –0.07 [–0.57, 0.43] | 0.777 | 0.05 [–0.29, 0.39] | 0.784 | 0.00 [–0.27, 0.26] | 0.991 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 3 | –0.03 [–0.52, 0.46] | 0.908 | 0.23 [–0.07, 0.53] | 0.129 | 0.12 [–0.10, 0.34] | 0.290 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 4 | 0.12 [–0.31, 0.56] | 0.579 | –0.17 [–0.48, 0.15] | 0.297 | –0.08 [–0.45, 0.30] | 0.690 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Total Alcohol Years | –0.03 [–0.05, –0.01] | 0.002* | –0.03 [–0.04, –0.02] | <0.001* | –0.02 [–0.04, –0.01] | 0.004* | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 1 | 0.26 [0.11, 0.41] | <0.001 | 0.20 [0.08, 0.31] | 0.001 | 0.22 [0.10, 0.33] | <0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 2 | 0.18 [0.03, 0.34] | 0.019 | 0.23 [0.08, 0.38] | 0.004 | 0.19 [0.06, 0.33] | 0.006 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 3 | –0.05 [–0.21, 0.12] | 0.558 | 0.23 [0.12, 0.35] | <0.001 | 0.11 [0.00, 0.22] | 0.045 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quartile 4 | 0.06 [–0.11, 0.22] | 0.497 | –0.07 [–0.19, 0.04] | 0.192 | –0.05 [–0.20, 0.11] | 0.550 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| *Interaction terms with P ≤ 0.05. Bolded values represent the beta coefficient [95% CI] and P for the joint association between alcohol years and chronological age. Results are adjusted for sex, race, center, education, pack years of smoking, BMI, and physical activity. Beta coefficients represent the gain in EAA for each additional 5 alcohol years. Quartile 1: Y15[32≤age≤37], Y20[37≤age≤42]; Quartile 2: Y15[38≤age≤40], Y20[43≤age≤45]; Quartile 3: Y15[41≤age≤43], Y20[46≤age≤48]; and Quartile 4: Y15[44≤age≤49], Y20[49≤age≤54]. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
At Y20, we observed a 0.29-year [95% CI: 0.12, 0.46] higher GAA in quartile 1 per 5 beer years compared to a 0.17-year [95% CI: –0.34, –0.01] loss in GAA in quartile 4 (Pinteraction < 0.001). Moreover, quartile 1 experienced a 0.20-year [95% CI: 0.08, 0.31] higher GAA compared to a 0.07-year [95% CI: –0.19, 0.04] loss in GAA in quartile 4 per 5 total alcohol years (Pinteraction < 0.001). Interaction and stratified results from GEE provided similar findings. Restricting to study participants with complete follow up yielded consistent conclusions (Supplementary Table 11). We observed similar results when adjusting for both cumulative alcohol consumption and recent binge drinking (Supplementary Table 12). While interactions of cumulative alcohol consumption with sex yielded primarily non-significant associations, female participants displayed higher GAA with greater consumption of beer, wine, and total alcohol compared to male participants (Supplementary Table 13). Additionally, Black participants experienced a higher GAA with greater consumption of liquor and total alcohol compared to White participants, although these interactions were non-significant (Supplementary Table 14).
Discussion
We observed positive associations between cumulative alcohol consumption and binge drinking with GAA in young adults. Specifically, we observed positive associations between liquor and total alcohol years with GAA. Beer and wine years were marginally and not associated with GAA, respectively. We also observed positive associations between recent binge drinking and each additional day of recent binge drinking with GAA, respectively. Additionally, we identified statistical interactions between beer and total alcohol years with chronological age, with younger participants exhibiting a higher average in GAA compared to older participants. Our results provide novel insight into the association of cumulative and recent alcohol consumption, as well as the type of alcohol, on age-related epigenetic changes.
Estimated biological age via DNA methylation has provided unique molecular insights into the aging process and age-related conditions. Additionally, these metrics have furthered our understanding of the impact comorbidities, behaviors, and lifestyle factors, including alcohol consumption, have on epigenetic age-related changes. We observed cumulative alcohol consumption was positively associated with GAA across time, suggesting long-term alcohol consumption may accelerate epigenetic aging. Alcohol has previously been associated with several of the GrimAge surrogate biomarkers of blood plasma proteins, including PAI-1 [20] and leptin [21]. We observed similar correlations between these surrogate biomarkers of blood plasma proteins and several alcohol variables at Y15 and Y20, suggesting the observed associations between alcohol consumption and epigenetic aging as estimated by GAA may be partially explained by the correlations between alcohol and the surrogate biomarkers of blood plasma proteins of GrimAge. Our findings, i.e., positive association between total alcohol consumption and epigenetic aging, are consistent with previously observed associations [11–16]. Findings from our study add to the current literature by identifying associations between cumulative, lifetime alcohol consumption and epigenetic age acceleration in a diverse study population, as well as showing increasing effects on epigenetic aging with higher alcohol consumption, as demonstrated during the quantile regression analyses. Our findings, together with previous studies, illustrate the GrimAge surrogate biomarkers of blood plasma proteins may be modulated by alcohol consumption, with potential subsequent negative impacts on the aging process. Behavioral modifications to limit alcohol consumption may reduce alcohol induced epigenetic age-related changes and potentially, increase lifespan.
The type of alcohol consumed has been shown to have varying associations with numerous age-related diseases [4, 22, 23]. In our study, we observed positive associations between beer, liquor, and total alcohol years with GAA and notably, null associations with wine years. Prior studies have investigated the molecular and cellular mechanisms underlying the varying effects of alcohol consumption on the aging process. Polyphenols, a group of naturally occurring anti-oxidants, have been suggested to modulate adaptive immune responses and influence anti-aging mechanisms, including preventing cellular senescence, and are present in alcohol by various quantities [24]. The hops and malt in beer contain phenolic compounds that exhibit anti-carcinogenic and anti-inflammatory properties, such as inhibiting inducible nitric oxide (NO) synthase and cyclooxygenase 2 (COX2) [25]. Production of NO and COX2 have been associated with cellular aging and may modulate epigenetic aging through similar mechanisms [26, 27]. Additionally, moderate beer consumption has been associated with lower risk of cardiovascular disease, potentially in part by the cardioprotective effects of polyphenols [28, 29]. Contrary to these findings, our results showed accelerated epigenetic aging with beer consumption, demonstrating potential independent associations between age-related epigenetic changes and disease. Compared to beer and wine, liquor has the highest alcohol content but the lowest polyphenolic concentration, which may impact the beneficial health gains as seen with wine and beer [25]. Liquor has been associated with greater risk of lung [4], and upper digestive tract cancers [30] and overall mortality [31], suggesting potentially negative impacts of liquor on the aging process and age related diseases. Wine contains resveratrol, a polyphenol that exhibits anti-aging properties by acting as a potent SIRT1 activator to regulate longevity and has shown to improve health and longevity in mice models [32]. As a part of the Mediterranean diet, moderate wine consumption has been shown to protect against age-related diseases including cancer [4, 33] and dementia [34] and increase life expectancy [35]. While we observed associations with alcohol consumption and GAA, the null findings with the other EAA metrics may be partially explained by different sets of CpGs and covariates used in the development of the epigenetic age measures, which captures different biological processes of aging. While previous studies examined total alcohol consumption and EAA [11–16], we identified associations with cumulative alcohol specific consumption and epigenetic aging, providing novel insight into alcohol specific effects on the epigenome. In sum, these findings suggest the type of alcohol may influence epigenetic age-related changes differently and as such, altering beverage preferences may modulate the aging process.
Alcohol’s association on age-related epigenetic changes appear to differ by the pattern and severity of consumption. We observed large effect estimates for recent binge drinking on GAA, although the effect estimates for the number of days of binge drinking were similar to those observed for cumulative alcohol consumption. The large, although transient, effect of binge drinking on GAA may be due to pharmacokinetics, where the rate of absorption is greater than the rate of elimination during binging episodes, resulting in high blood alcohol concentrations. Compared to non-binge drinking consumption, binge drinking leads to acutely elevated levels of alcohol, which can negatively impact the body through organ injury, local and systemic inflammation, and endotoxemia, leading to potential multisystemic pathophysiological consequences [36]. Furthermore, the generation of reactive oxygen species and alcohol metabolites and disruption of anti-oxidant mechanisms, leads to cellular dysfunction and immune system dysfunction and exhaustion [36]. Binge drinking has also been associated with PAI-1 [37], suggesting the observed association between binge drinking and GAA may be partially explained by the correlations between binge drinking and the surrogate biomarkers of blood plasma proteins of GrimAge. Our results demonstrate differences in the effect of cumulative and binge alcohol consumption patterns on epigenetic aging, and suggests lifestyle changes, such as limiting binge drinking, may aid in slowing biological aging.
Consumption of alcohol varies with chronological age, with the prevalence and quantity inversely associated with age [5, 38]. While the mechanisms for the observed statistical interactions by chronological age remains unclear, one possible explanation involves age-related changes in alcohol preference. Findings from five national alcohol surveys showed beer and liquor consumption decline with increasing age, while wine consumption exhibited a more complex pattern [39]. In the current study, older participants exhibited greater wine years and lower beer and liquor years, suggesting that changes in alcohol consumption patterns may modulate age-related epigenetic changes. An additional explanation maybe due to alterations in the pattern of alcohol consumption. For example, binge drinking has previously been inversely associated with chronological age [40]. We observed participants in the highest age quartile reported the lowest binging events compared to other quartiles, consistent with declining excessive drinking with increasing age. Our findings suggest age-related changes to alcohol consumption may influence epigenetic aging and may serve to slow biological aging later in life.
As a longitudinal study with a large sample size, we were able to obtain multiple alcohol and DNA methylation measurements. Additionally, the detailed collection of type-specific alcohol consumption data allowed us to elucidate specific effects on EAA. This study, however, is not without limitations. Alcohol consumption was self-reported, which may be subject to social desirability bias in which participants report levels of consumption that are socially acceptable. However, consistent findings were observed with multiple time points and all responses at each examination were confidential. Despite being a diverse study population, CARDIA enrolled Black and White participants at four locations across the United States and as such, additional studies with more diverse populations are needed to better generalize these findings. Additionally, null associations between wine and the EAA metrics may be related to the sample size of this study and thus, replication of the cumulative alcohol specific associations in larger studies are needed to validate the findings presented here. Furthermore, race is a social construct and studies investigating genetic ancestry, as estimated by single nucleotide polymorphisms, may provide additional insights into the association between alcohol consumption and biological aging. Finally, due to the age of the study population, we were unable to link associations to any clinically significant, alcohol-related outcomes. Future research in older populations, and as CARDIA ages, will help elucidate the complete relationship between alcohol, epigenetics, and health.
In conclusion, we observed significant associations between cumulative and recent alcohol consumption, as well as the type of alcohol, with GAA. Moreover, we observed statistical interactions between cumulative alcohol consumption and chronological age on GAA. The findings presented here add to the existing literature on alcohol and the aging process and provide novel insights into the heterogeneous effects of the type of alcohol on epigenetic aging. Replication of our findings is needed as well as further exploration of the causal mechanisms of alcohol on epigenetic age-related changes. With the high prevalence of alcohol consumption and the growing aging population in the United States, elucidating changes to the epigenome due to alcohol consumption may yield novel insights into the aging process and potentially, longevity facilitating lifestyle modifications.
Methods
Study sample
Details of the original CARDIA study design and examinations have previously been described [41]. Briefly, CARDIA is a population-based, cohort study designed to investigate the determinants of subclinical and clinical cardiovascular disease. Four centers across the United States recruited 5,115 Black and White participants ages 18 to 30 years between 1985 to 1986 and received in person examinations at 1985-1986 (baseline visit; year 0 [Y0]), 1987-88 (Y2), 1990-91 (Y5), 1992-93 (Y7), 1995-96 (Y10), 2000-01 (Y15), 2005-06 (Y20), 2010-11 (Y25), 2015-16 (Y30), and currently participating in 2020-21 (Y35).
Alcohol consumption measurements
Alcohol consumption was assessed at Y0 and at each follow up examination. Participants were asked “Did you drink any alcoholic beverages in the past year?” and if answered yes, participants were asked “How many drinks of [alcohol type] do you usually have per week?” for beer (a 12-ounce glass, can, or bottle), liquor (a shot or 1.5-ounces), and wine (a 5-ounce glass). Binge drinking was also obtained by asking participants “During the past 30 days, on how many days did you have five or more drinks on the same occasion?” We considered six alcohol variables at Y15 and Y20: four continuous alcohol variables capturing cumulative consumption from Y0 to Y15 and from Y0 to Y20, i.e., beer, liquor, wine, and total alcohol (the sum of the individual alcohol types); one binary variable indicating whether a participant binge drank in the past 30 days at Y15 and Y20; and one continuous variable indicating the number of days a participant binge drank during those 30 days at Y15 and Y20. In order to calculate cumulative alcohol consumption from all available CARDIA data from Y0 to Y15 and Y20 separately, we assumed weekly consumption of alcohol measured at each CARDIA exam was consistent in-between study examinations, i.e., weekly alcohol consumption represented consumption throughout the year and between examinations. For each weekly alcohol variable, we estimated ‘alcohol years’ by multiplying each alcohol variable by 52 weeks and the number of years between visits and dividing by 365 days, where an alcohol year is equivalent to 365 days of alcohol consumption. We then estimated alcohol years for each alcohol type separately and total alcohol by summing across examinations from Y0 to Y15 and Y20.
DNA methylation profiling
Details of methylation profiling and quality control have previously been described [19, 42–44]. Briefly, a randomly selected subset of 1,200 study participants with available whole blood repeatedly collected at both Y15 and Y20 (a total of 2,400 samples) underwent DNA methylation profiling using the Illumina MethylationEPIC Beadchip. The R package ENmix was used to perform data preprocessing and quality control using default parameter settings [45]. Low quality methylation measurements were defined as having less than 3 beads or a detection P < 1 × 10−6. A total of 87 samples with low quality methylation measurements >5% or bisulfite conversion process of extremely low intensity (defined as less than 3 × standard deviation of the intensity across samples below the mean intensity) and 6,209 CpG sites with a detection rate <95% were excluded from further analysis. Furthermore, 95 samples were identified as extreme outliers via the average total intensity value [intensity of the unmethylated signal (U) + intensity of the methylated signal (M)] or β value [M/(U + M + 100)] across all CpG sites and Tukey’s method [46]. Application of a model-based background correction was conducted using ENmix and dye bias correction was performed using RELIC [47]. M and U intensities for Infinium I or II probes were quantile-normalized separately, respectively. Low quality methylation measurements and β value outliers were set to missing. After applying these quality control measures, the final methylation dataset for epigenetic age calculation included 1,042 and 957 samples at Y15 and Y20, respectively. Supplementary Figure 3 presents a graphical timeline of the examinations with phenotype and blood draw collection.
Epigenetic age calculation
Four epigenetic ages were calculated at Y15 and Y20. Horvath’s age, intrinsic epigenetic age acceleration (IEAA), was calculated with 353 CpGs and is associated with cell-intrinsic aging [7]. Hannum’s age, extrinsic epigenetic age acceleration (EEAA), was estimated from 71 CpGs and is associated with immunological aging [8]. Levine’s age, PhenoAge acceleration (PAA), was calculated with 513 CpGs and is associated with comorbidities and physical functionality [9]. Lu’s age, GrimAge acceleration (GAA), was estimated from 1,030 CpGs and is associated with lifespan [10]. A publicly available online calculator was used to calculate the DNA-methylation epigenetic age estimates (https://dnamage.genetics.ucla.edu/new). EAA for each metric was estimated as the residuals from a linear model of chronological age and the top eight technical principal components, capturing technical biases and batch effect, on epigenetic age as previously described [42].
Statistical analysis
Multiple linear regression and generalized estimating equations (GEE) were used to examine the associations between the cumulative and binge drinking alcohol variables (independent variables) and each EAA (outcome variables) at Y15 and Y20. Each cumulative alcohol variable was rescaled to units of five drink years (i.e., divided by 5) to aid in the interpretation of beta coefficients. Drinkers were further categorized into low, intermediate, or high consumption via tertiles and compared to non-drinkers for each cumulative alcohol variable. Quantile regression was additionally performed to evaluate the conditional quantiles of each cumulative alcohol variable on EAA. This approach provides a more comprehensive view of the covariate effects on variables under study [48]. Due to previously observed differences in alcohol consumption by demographic characteristics [5, 6], interaction and stratified analyses were conducted by chronological age, sex, and race with the cumulative alcohol variables. Age was categorized into quartiles during stratified analyses: quartile 1 (nY15[32≤age≤37] = 242, nY20[37≤age≤42] = 223), quartile 2 (nY15[38≤age≤40] = 246, nY20[43≤age≤45] = 224), quartile 3 (nY15[41≤age≤43] = 294, nY20[46≤age≤48] = 271), and quartile 4 (nY15[44≤age≤49] = 248, nY20[49≤age≤54] = 227). Sensitivity analyses using study participants with complete follow up were performed. All models were adjusted for chronological age, sex, race, center, education, pack years of smoking, body mass index, and physical activity. Pack years of smoking was estimated by converting the number of daily smoked cigarettes measured at each examination to packs of cigarettes, multiplying by the number of years between examinations, and summing the products from Y0 to Y15 and Y20, separately. Associations were declared significant if P ≤ 0.05. All statistical analyses were performed using SAS 9.4.
Abbreviations
BMI: body mass index; CARDIA: Coronary Artery Risk Development in Young Adults; EAA: epigenetic age acceleration; EEAA: extrinsic epigenetic age acceleration; GAA: GrimAge acceleration; IEAA: intrinsic epigenetic age acceleration; PAA: PhenoAge acceleration.
Author Contributions
Study conception and design by LF and BJ. Data processing and quality control by YZ, TG, and JW. Statistical analyses and manuscript drafting by DN. Manuscript writing and revisions by LF, BJ, and YZ. All authors reviewed and provided comments to the final manuscript (LL, DJ, PS, CL, QD, SH, AL, KY, PG, DJ). All authors read and approved the final manuscript.
Acknowledgments
We would like to thank the CARDIA study participants, as well as the staff that aided in data collection and processing.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Ethical Statement and Consent
The Institutional Review Boards at all participating institutions approved this study and all study participants provided written consent.
Funding
The Coronary Artery Risk Development in Young Adults Study (CARDIA) is conducted and supported by the National Heart, Lung, and Blood Institute (NHLBI) in collaboration with the University of Alabama at Birmingham (HHSN268201800005I and HHSN268201800007I), Northwestern University (HHSN268201800003I), University of Minnesota (HHSN268201800006I), and Kaiser Foundation Research Institute (HHSN268201800004I). The DNA methylation laboratory work and analytical component were funded by American Heart Association (17SFRN33700278 and 14SFRN20790000, Northwestern University) and National Institute on Aging (R21AG063370 and R21AG068955). The Longer Life Foundation (2019-008) provided additional support.
References
- 1. Substance Abuse and Mental Health Services Administration (SAMHSA). 2018 National Survey on Drug Use and Health (NSDUH): Table 2.1B - Tobacco Product and Alcohol Use in Lifetime, Past Year, and Past Month among Persons Aged 12 or Older, by Age Group: Percentages, 2017 and 2018. 2018.
- 2. GBD 2016 Alcohol Collaborators. Alcohol use and burden for 195 countries and territories, 1990-2016: a systematic analysis for the Global Burden of Disease Study 2016. Lancet. 2018; 392:1015–35. https://doi.org/10.1016/S0140-6736(18)31310-2 [PubMed]
- 3. Bagnardi V, Rota M, Botteri E, Tramacere I, Islami F, Fedirko V, Scotti L, Jenab M, Turati F, Pasquali E, Pelucchi C, Galeone C, Bellocco R, et al. Alcohol consumption and site-specific cancer risk: a comprehensive dose-response meta-analysis. Br J Cancer. 2015; 112:580–93. https://doi.org/10.1038/bjc.2014.579 [PubMed]
- 4. Chao C. Associations between beer, wine, and liquor consumption and lung cancer risk: a meta-analysis. Cancer Epidemiol Biomarkers Prev. 2007; 16:2436–47. https://doi.org/10.1158/1055-9965.EPI-07-0386 [PubMed]
- 5. Moore AA, Gould R, Reuben DB, Greendale GA, Carter MK, Zhou K, Karlamangla A. Longitudinal patterns and predictors of alcohol consumption in the United States. Am J Public Health. 2005; 95:458–65. https://doi.org/10.2105/AJPH.2003.019471 [PubMed]
- 6. Platt A, Sloan FA, Costanzo P. Alcohol-consumption trajectories and associated characteristics among adults older than age 50. J Stud Alcohol Drugs. 2010; 71:169–79. https://doi.org/10.15288/jsad.2010.71.169 [PubMed]
- 7. Horvath S. DNA methylation age of human tissues and cell types. Genome Biol. 2013; 14:R115. https://doi.org/10.1186/gb-2013-14-10-r115 [PubMed]
- 8. Hannum G, Guinney J, Zhao L, Zhang L, Hughes G, Sadda S, Klotzle B, Bibikova M, Fan JB, Gao Y, Deconde R, Chen M, Rajapakse I, et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol Cell. 2013; 49:359–67. https://doi.org/10.1016/j.molcel.2012.10.016 [PubMed]
- 9. Levine ME, Lu AT, Quach A, Chen BH, Assimes TL, Bandinelli S, Hou L, Baccarelli AA, Stewart JD, Li Y, Whitsel EA, Wilson JG, Reiner AP, et al. An epigenetic biomarker of aging for lifespan and healthspan. Aging (Albany NY). 2018; 10:573–91. https://doi.org/10.18632/aging.101414 [PubMed]
- 10. Lu AT, Quach A, Wilson JG, Reiner AP, Aviv A, Raj K, Hou L, Baccarelli AA, Li Y, Stewart JD, Whitsel EA, Assimes TL, Ferrucci L, Horvath S. DNA methylation GrimAge strongly predicts lifespan and healthspan. Aging (Albany NY). 2019; 11:303–27. https://doi.org/10.18632/aging.101684 [PubMed]
- 11. Fiorito G, McCrory C, Robinson O, Carmeli C, Ochoa-Rosales C, Zhang Y, Colicino E, Dugué PA, Artaud F, McKay GJ, Jeong A, Mishra PP, Nøst TH, et al, and BIOS Consortium, and Lifepath consortium. Socioeconomic position, lifestyle habits and biomarkers of epigenetic aging: a multi-cohort analysis. Aging (Albany NY). 2019; 11:2045–70. https://doi.org/10.18632/aging.101900 [PubMed]
- 12. Quach A, Levine ME, Tanaka T, Lu AT, Chen BH, Ferrucci L, Ritz B, Bandinelli S, Neuhouser ML, Beasley JM, Snetselaar L, Wallace RB, Tsao PS, et al. Epigenetic clock analysis of diet, exercise, education, and lifestyle factors. Aging (Albany NY). 2017; 9:419–46. https://doi.org/10.18632/aging.101168 [PubMed]
- 13. Kresovich JK, Martinez Lopez AM, Garval EL, Xu Z, White AJ, Sandler DP, Taylor JA. Alcohol Consumption and Methylation-Based Measures of Biological Age. J Gerontol A Biol Sci Med Sci. 2021; 76:2107–11. https://doi.org/10.1093/gerona/glab149 [PubMed]
- 14. Bøstrand SMK, Vaher K, de Nooij L, Harris MA, Cole JH, Cox SR, Marioni RE, McCartney DL, Walker RM, McIntosh AM, Evans KL, Whalley HC, Wootton RE, Clarke TK. Associations between alcohol use and accelerated biological ageing. Addict Biol. 2022; 27:e13100. https://doi.org/10.1111/adb.13100 [PubMed]
- 15. Zhao W, Ammous F, Ratliff S, Liu J, Yu M, Mosley TH, Kardia SLR, Smith JA. Education and Lifestyle Factors Are Associated with DNA Methylation Clocks in Older African Americans. Int J Environ Res Public Health. 2019; 16:3141. https://doi.org/10.3390/ijerph16173141 [PubMed]
- 16. Schmitz LL, Zhao W, Ratliff SM, Goodwin J, Miao J, Lu Q, Guo X, Taylor KD, Ding J, Liu Y, Levine M, Smith JA. The Socioeconomic Gradient in Epigenetic Ageing Clocks: Evidence from the Multi-Ethnic Study of Atherosclerosis and the Health and Retirement Study. Epigenetics. 2022; 17:589–611. https://doi.org/10.1080/15592294.2021.1939479 [PubMed]
- 17. Rosen AD, Robertson KD, Hlady RA, Muench C, Lee J, Philibert R, Horvath S, Kaminsky ZA, Lohoff FW. DNA methylation age is accelerated in alcohol dependence. Transl Psychiatry. 2018; 8:182. https://doi.org/10.1038/s41398-018-0233-4 [PubMed]
- 18. Luo A, Jung J, Longley M, Rosoff DB, Charlet K, Muench C, Lee J, Hodgkinson CA, Goldman D, Horvath S, Kaminsky ZA, Lohoff FW. Epigenetic aging is accelerated in alcohol use disorder and regulated by genetic variation in APOL2. Neuropsychopharmacology. 2020; 45:327–36. https://doi.org/10.1038/s41386-019-0500-y [PubMed]
- 19. Nannini DR, Joyce BT, Zheng Y, Gao T, Liu L, Yoon G, Huan T, Ma J, Jacobs DR
Jr , Wilkins JT, Ren J, Zhang K, Khan SS, et al. Epigenetic age acceleration and metabolic syndrome in the coronary artery risk development in young adults study. Clin Epigenetics. 2019; 11:160. https://doi.org/10.1186/s13148-019-0767-1 [PubMed] - 20. Djoussé L, Pankow JS, Arnett DK, Zhang Y, Hong Y, Province MA, Ellison RC. Alcohol consumption and plasminogen activator inhibitor type 1: the National Heart, Lung, and Blood Institute Family Heart Study. Am Heart J. 2000; 139:704–9. https://doi.org/10.1016/s0002-8703(00)90052-8 [PubMed]
- 21. Röjdmark S, Calissendorff J, Brismar K. Alcohol ingestion decreases both diurnal and nocturnal secretion of leptin in healthy individuals. Clin Endocrinol (Oxf). 2001; 55:639–47. https://doi.org/10.1046/j.1365-2265.2001.01401.x [PubMed]
- 22. Huang J, Wang X, Zhang Y. Specific types of alcoholic beverage consumption and risk of type 2 diabetes: A systematic review and meta-analysis. J Diabetes Investig. 2017; 8:56–68. https://doi.org/10.1111/jdi.12537 [PubMed]
- 23. Costanzo S, Di Castelnuovo A, Donati MB, Iacoviello L, de Gaetano G. Wine, beer or spirit drinking in relation to fatal and non-fatal cardiovascularevents: a meta-analysis. Eur J Epidemiol. 2011; 26:833–50. https://doi.org/10.1007/s10654-011-9631-0 [PubMed]
- 24. Luo J, Si H, Jia Z, Liu D. Dietary Anti-Aging Polyphenols and Potential Mechanisms. Antioxidants (Basel). 2021; 10:283. https://doi.org/10.3390/antiox10020283 [PubMed]
- 25. Arranz S, Chiva-Blanch G, Valderas-Martínez P, Medina-Remón A, Lamuela-Raventós RM, Estruch R. Wine, beer, alcohol and polyphenols on cardiovascular disease and cancer. Nutrients. 2012; 4:759–81. https://doi.org/10.3390/nu4070759 [PubMed]
- 26. Pourbagher-Shahri AM, Farkhondeh T, Talebi M, Kopustinskiene DM, Samarghandian S, Bernatoniene J. An Overview of NO Signaling Pathways in Aging. Molecules. 2021; 26:4533. https://doi.org/10.3390/molecules26154533 [PubMed]
- 27. Kim J, Vaish V, Feng M, Field K, Chatzistamou I, Shim M. Transgenic expression of cyclooxygenase-2 (COX2) causes premature aging phenotypes in mice. Aging (Albany NY). 2016; 8:2392–406. https://doi.org/10.18632/aging.101060 [PubMed]
- 28. Bobak M, Skodova Z, Marmot M. Effect of beer drinking on risk of myocardial infarction: population based case-control study. BMJ. 2000; 320:1378–9. https://doi.org/10.1136/bmj.320.7246.1378 [PubMed]
- 29. Marcos A, Serra-Majem L, Pérez-Jiménez F, Pascual V, Tinahones FJ, Estruch R. Moderate Consumption of Beer and Its Effects on Cardiovascular and Metabolic Health: An Updated Review of Recent Scientific Evidence. Nutrients. 2021; 13:879. https://doi.org/10.3390/nu13030879 [PubMed]
- 30. Grønbaek M, Becker U, Johansen D, Tønnesen H, Jensen G, Sørensen TI. Population based cohort study of the association between alcohol intake and cancer of the upper digestive tract. BMJ. 1998; 317:844–7. https://doi.org/10.1136/bmj.317.7162.844 [PubMed]
- 31. Grønbaek M, Deis A, Sørensen TI, Becker U, Schnohr P, Jensen G. Mortality associated with moderate intakes of wine, beer, or spirits. BMJ. 1995; 310:1165–9. https://doi.org/10.1136/bmj.310.6988.1165 [PubMed]
- 32. Baur JA, Pearson KJ, Price NL, Jamieson HA, Lerin C, Kalra A, Prabhu VV, Allard JS, Lopez-Lluch G, Lewis K, Pistell PJ, Poosala S, Becker KG, et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature. 2006; 444:337–42. https://doi.org/10.1038/nature05354 [PubMed]
- 33. Barstad B, Sørensen TI, Tjønneland A, Johansen D, Becker U, Andersen IB, Grønbaek M. Intake of wine, beer and spirits and risk of gastric cancer. Eur J Cancer Prev. 2005; 14:239–43. https://doi.org/10.1097/00008469-200506000-00007 [PubMed]
- 34. Truelsen T, Thudium D, Grønbaek M, and Copenhagen City Heart Study. Amount and type of alcohol and risk of dementia: the Copenhagen City Heart Study. Neurology. 2002; 59:1313–9. https://doi.org/10.1212/01.wnl.0000031421.50369.e7 [PubMed]
- 35. Streppel MT, Ocké MC, Boshuizen HC, Kok FJ, Kromhout D. Long-term wine consumption is related to cardiovascular mortality and life expectancy independently of moderate alcohol intake: the Zutphen Study. J Epidemiol Community Health. 2009; 63:534–40. https://doi.org/10.1136/jech.2008.082198 [PubMed]
- 36. Molina PE, Nelson S. Binge Drinking's Effects on the Body. Alcohol Res. 2018; 39:99–109. [PubMed]
- 37. Numminen H, Syrjälä M, Benthin G, Kaste M, Hillbom M. The effect of acute ingestion of a large dose of alcohol on the hemostatic system and its circadian variation. Stroke. 2000; 31:1269–73. https://doi.org/10.1161/01.str.31.6.1269 [PubMed]
- 38. Eigenbrodt ML, Mosley TH
Jr , Hutchinson RG, Watson RL, Chambless LE, Szklo M. Alcohol consumption with age: a cross-sectional and longitudinal study of the Atherosclerosis Risk in Communities (ARIC) study, 1987-1995. Am J Epidemiol. 2001; 153:1102–11. https://doi.org/10.1093/aje/153.11.1102 [PubMed] - 39. Kerr WC, Greenfield TK, Bond J, Ye Y, Rehm J. Age, period and cohort influences on beer, wine and spirits consumption trends in the US National Alcohol Surveys. Addiction. 2004; 99:1111–20. https://doi.org/10.1111/j.1360-0443.2004.00820.x [PubMed]
- 40. Naimi TS, Brewer RD, Mokdad A, Denny C, Serdula MK, Marks JS. Binge drinking among US adults. JAMA. 2003; 289:70–5. https://doi.org/10.1001/jama.289.1.70 [PubMed]
- 41. Friedman GD, Cutter GR, Donahue RP, Hughes GH, Hulley SB, Jacobs DR
Jr , Liu K, Savage PJ. CARDIA: study design, recruitment, and some characteristics of the examined subjects. J Clin Epidemiol. 1988; 41:1105–16. https://doi.org/10.1016/0895-4356(88)90080-7 [PubMed] - 42. Joyce BT, Gao T, Zheng Y, Ma J, Hwang SJ, Liu L, Nannini D, Horvath S, Lu AT, Bai Allen N, Jacobs DR
Jr , Gross M, Krefman A, et al. Epigenetic Age Acceleration Reflects Long-Term Cardiovascular Health. Circ Res. 2021; 129:770–81. https://doi.org/10.1161/CIRCRESAHA.121.318965 [PubMed] - 43. Zheng Y, Joyce BT, Hwang SJ, Ma J, Liu L, Allen NB, Krefman AE, Wang J, Gao T, Nannini DR, Zhang H, Jacobs DR
Jr , Gross MD, et al. Association of Cardiovascular Health Through Young Adulthood With Genome-Wide DNA Methylation Patterns in Midlife: The CARDIA Study. Circulation. 2022; 146:94–109. https://doi.org/10.1161/CIRCULATIONAHA.121.055484 [PubMed] - 44. Nannini DR, Zheng Y, Joyce BT, Gao T, Liu L, Jacobs DR
Jr , Schreiner P, Liu C, Horvath S, Lu AT, Yaffe K, Sidney S, Greenland P, et al. Marijuana use and DNA methylation-based biological age in young adults. Clin Epigenetics. 2022; 14:134. https://doi.org/10.1186/s13148-022-01359-8 [PubMed] - 45. Xu Z, Niu L, Li L, Taylor JA. ENmix: a novel background correction method for Illumina HumanMethylation450 BeadChip. Nucleic Acids Res. 2016; 44:e20. https://doi.org/10.1093/nar/gkv907 [PubMed]
- 46. Tukey JW. Exploratory Data Analysis: Pearson. 1977. http://theta.edu.pl/wp-content/uploads/2012/10/exploratorydataanalysis_tukey.pdf.
- 47. Xu Z, Langie SA, De Boever P, Taylor JA, Niu L. RELIC: a novel dye-bias correction method for Illumina Methylation BeadChip. BMC Genomics. 2017; 18:4. https://doi.org/10.1186/s12864-016-3426-3 [PubMed]
- 48. Koenker R, Hallock KF. Quantile Regression. J Econ Perspect. 2001; 15:143–56. https://doi.org/10.1257/jep.15.4.143