



Introduction
Osteosarcoma (OS) is the most common bone malignancy in children and adolescents [1, 2] and it accounts for 0.2% of all malignancy cases [3]. The current standard of care for OS includes neoadjuvant chemotherapy, radical resection and adjuvant chemotherapy [4]. Because of the use of these comprehensive treatments, the 5-year overall survival rate of primary patients OS has reached approximately 60–70% [4]. However, the 5-year overall survival rate decreases to lower than 15% after the occurrence of distant metastasis [4, 5]. Although new drugs for OS treatment, such as immune checkpoint inhibitors and targeted drugs [6, 7], are constantly being explored, the overall survival rate of OS has still not significantly improved in the past 30 years [8]. There is still an urgent need to discover new treatment options.
Angelicin is one of the main active ingredients of Angelica and Angelica dahurica [9]. Existing evidence has proven that angelicin exerts inhibitory effects on tumor cells by inhibiting proliferation, apoptosis, migration, and invasion [9–12]. However, research related to the role of Angelicin inhibiting OS and its underlying mechanism is insufficient.
Recently, network pharmacology has offered a new strategy for exploring the relationships between drugs and diseases by integrating systems biology, multidirectional pharmaceutical biology, bioinformatics, and computer science [13, 14]. Currently, there are no published studies on the effect of angelicin on the biological behavior of OS cells. In this study, we used network pharmacology and molecular docking techniques and in vitro experiments to explore whether angelicin affects the biological behavior of OS and its specific mechanism.
Materials and Methods
The flow chart of the study design is shown in Figure 1.
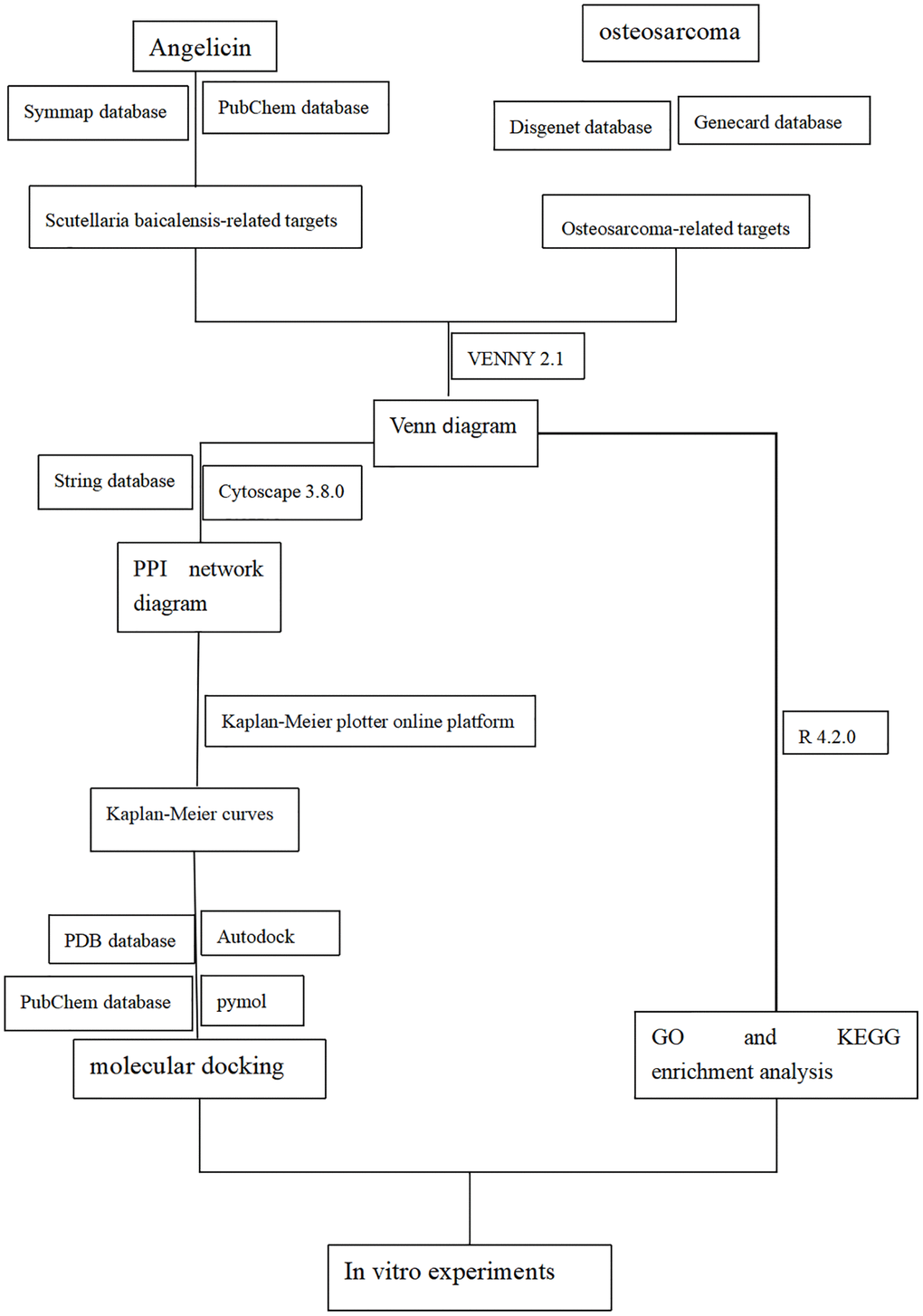
Figure 1. Flow diagram of the experimental design for studying the function of angelicin in the treatment of osteosarcoma.
Identification of angelicin targets
All the active ingredients of Angelicin were obtained from the Pubchem (https://pubchem.ncbi.nlm.nih.gov/) and SymMap (http://www.symmap.org) databases with the keyword “Angelicin”. PubChem: that includes a substantial amount of data related to small organic molecule bioactivity and is funded by the National Institutes of Health (NIH). The SymMap database is a traditional Chinese medicine (TCM) evidence association database that includes an immeasurable amount of information about herbal medicines, their ingredients, and their drug targets; the TCM symptoms, Western medicine symptoms, and diseases associated with these symptoms that these medicines are used to treat; and correlations among these data. In this way, the SymMap database correlates TCM and Western medicine at levels ranging from the phenotypic level to the molecular level.
Identification of OS targets
A search was performed in the DisGeNET database (https://www.disgenet.org/) and the GeneCards database (https://www.genecards.org/) using the keyword “Osteosarcoma” and using “Homo sapiens” as the filtering criterion. In addition, the targets obtained from the DisGeNET database with a score >0.8 were screened. The GeneCards database is a search platform that retrieves genes that are associated with human diseases from 150+ web sources. Genes were searched on the two platforms using the keyword “Osteosarcoma”. The retrieved data include information about Osteosarcoma, such as gene names and gene IDs. The targets that were obtained from these two databases were then integrated, and duplicates were removed, ultimately yielding genes related to “Homo sapiens OS”.
Construction of a PPI network diagram
The identified angelicin-related targets and Homo sapiens OS-related targets were imported into an online Venn diagram mapping platform (https://bioinfogp.cnb.csic.es/tools/venny/). The region where the two circles of the Venn diagram intersect indicates the potential targets of angelicin in the treatment of OS. The targets that were related to the mechanism by which angelicin treats OS were imported into the STRING database. A PPI network diagram was generated with a confidence level of 0.7, and it was imported into the STRING app in Cytoscape software for further processing and analysis.
Hub gene analysis
To further analyze the PPI network and identify hub targets, we imported the PPI network into Cytoscape 3.8.0 software and analyzed the network graph with Network Analyzer. Information on the degree, betweenness centrality (BC), and closeness centrality (CC) of the corresponding targets was obtained. We used degree≥degree median, CC≤CC median, and BC≥BC median as the screening criteria to identify the hub targets.
Gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses
To further analyze the potential mechanism underlying the function of angelicin in OS treatment, we enriched the targets mentioned above with the Bioconductor Cluster Profiler package in R software. According to the GO enrichment analysis, the potential therapeutic targets of angelicin in the treatment of OS are described in terms of biological processes, cellular components, and molecular functions. Additionally, the potential signaling pathways by which angelicin functions in the treatment of OS were explored by KEGG enrichment analysis.
Molecular docking
To investigate the potential interaction between angelicin and its potential targets for the treatment of OS, we performed molecular docking between angelicin and the hub genes based on the principle of semiflexible molecular docking. The leading software for molecular docking is AutoDock, which includes AutoDock tools and AutoDock Vina. Angelicin and hub targets were processed with AutoDock tools, and the docking center of the hub targets was determined. The 3D structures of the target proteins were obtained from the PDB online platform, and the 3D structures of angelicin were obtained from the PubChem online platform.
Cell culture
The human OS cell line MG63 (Procell CL-0157) was kindly provided by Procell Life Science and Technology Co., Ltd. MG63 Cells were grown in 90% modified Eagle’s medium (MEM) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μmol/L streptomycin, and incubated at 37°C in a humidified atmosphere of 5% CO2.
Cell counting Kit-8 (CCK8) assay
MG63 cells were treated with trypsin, grown in 96-well plates (3000 cells/well), and cultured with media supplemented with various concentrations of angelicin. The cells were incubated in a 37°C cell culture incubator for 24 h, 48 h or 72 h. At each time point, the plates were removed, the medium in the corresponding wells was gently aspirated, and the cells were gently washed with sterile PBS. CCK8 and serum-free medium were mixed at a ratio of 1:9, and 100 μl of the mixture was slowly added to each well. The cells were incubated in a cell culture chamber at 37°C for 1 h. The OD value of each well was measured at 450 nm by a multifunctional enzyme marker. Cell activity (%) = (OD value of treatment group-OD value of blank control group)/(OD value of experimental control group-OD value of blank control group) × 100%.
Flow cytometry
According to the instructions, we analyzed the effect of angelicin on the apoptosis of MG63 cells by flow cytometry. MG63 cells were seeded in 6-well plates at a density of 5 × 105 cells per well; when the cells had completely adhered to the well, the original medium was discarded, and the cells were washed 2 times with PBS. Angelicin was added at different concentrations and incubated for 24 h. The cells were stained with propidium iodide (PI) and annexin V-fluorescein isothiocyanate (FITC). ACEA NovoCyteTM (Biosciences, San Diego, CA, USA) equipment was used to perform the flow cytometry analysis. FlowJo (Version 10.0, Stanford, CA, USA) software was used to analyze the results.
Wound healing experiment
MG63 OS cells in the logarithmic phase of growth were seeded in 6-well plates at a density of 3 × 105 cells per well. Then, the cell culture plates were gently shaken to uniformly distribute the cells to form a monolayer, and the cells were prepared for scoring when they reached 95% confluence. A 200 μL pipette tip was used to form a vertical scratch along a sterile straightedge. Then, the treated wells were gently washed 3 times with PBS to remove the nonadherent cells, and the plates were imaged and photographed (0 h). Media supplemented with different concentrations of angelicin (50 and 200 μmol/L) were added, and the control group was incubated in the same volume of MEM supplemented with DMSO; the cells were cultured in an incubator at a constant temperature of 37°C with 5% CO2 for 24 h or 48 h, and then, the cells were photographed. The scratch area S was obtained using the scratch area analysis module of ImageJ software, and the scratch healing rate was calculated according to the following formula: wound healing rate = (1-(S24h/S0h)) × 100%.
Reverse transcription (RT) PCR technology
Total RNA was extracted from cells using an RNA extraction kit, first-strand complementary DNA (cDNA) was synthesized from the total RNA using an RNA reverse transcriptase kit, and quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR) was performed using SYBR Green reagent (TaKaRa, Japan) according to the instructions. The primer sequences are shown in Table 1.
Table 1. The primer sequences of Bax, Bcl-2, CASP9, BIRC2.
| Primer name | Primer sequences (5′ to 3′) |
| Bcl2 Forward | CAGGATAACGGAGGCTGGGATG |
| Bcl2 Reverse | AGAAATCAAACAGAGGCCGCA |
| Bax Forward | CCCGAGAGGTCTTTTTCCGAG |
| Bax Reverse | CCAGCCCATGATGGTTCTGAT |
| CASP9 Forward | CGCTAATGCTGTTTCGGT |
| CASP9 Reverse | AAGATAAGGCAGGGTGAGG |
| BIRC2 Forward | AGTGGTTTCCAAGGTGTGAGT |
| BIRC2 Reverse | AGCCCATTTCCAAGGCAGAT |
Statistical analysis
Statistical analysis of the bioinformatics data and mapping was performed by R ×64 4.0.3, Cytoscape 3.8.0, and PyMOL software. Statistical analysis of the in vitro experimental data was conducted using GraphPad Prism 9. The difference between two groups was assessed using one-way ANOVA. The statistical significance of the CCK8 results was assessed using two-way ANOVA. The data are presented as the means ± standard errors of the means (SEM), and a p value < 0.05 indicates statistical significance.
Data availability statement
The original contributions presented in the study are included in the article Material, further inquiries can be directed to the corresponding authors.
Results
Targets of angelicin
Using “angelicin” as the keyword, we screened its targets in the PubChem and SymMap databases. We identified 36 targets of angelicin by integrating and deweighting the screened targets, as shown in Table 2.
Table 2. Information about 36 angelicin targets.
| Gene symbol | Gene name | Gene symbol | Gene name |
| BCL2 | BCL2, Apoptosis Regulator | ABCB11 | ATP-binding cassette subfamily B member 11 |
| CASP3 | Caspase 3 | ABCB5 | ATP-binding cassette sub-family B member 5 |
| CYP1A1 | Cytochrome P450 family 1 subfamily A member 1 | ABCC1 | Multidrug resistance-associated protein 1 |
| ICAM1 | Intercellular adhesion molecule 1 | ABCC2 | Multidrug resistance-associated protein 2 |
| BAX | Apoptosis regulator BAX | ABCC3 | Multidrug resistance-associated protein 3 |
| BIRC2 | Baculoviral IAP Repeat Containing 2 | ABCC4 | Multidrug resistance-associated protein 4 |
| CASP9 | Caspase 9 | ACHE | Acetylcholinesterase |
| FAM102A | Protein FAM102A | AHR | Aryl hydrocarbon receptor |
| CYP1A1 | Cytochrome P450 1A1 | NR1H4 | Bile acid receptor |
| CYP1A2 | Cytochrome P450 1A2 | SLC22A1 | Solute carrier family 22 member 1 |
| CYP2E1 | Cytochrome P450 2E1 | SLC22A2 | Solute carrier family 22 member 2 |
| CYP3A11 | Cytochrome P450 3A11 | SLC22A4 | Solute carrier family 22 member 4 |
| CYP7A1 | Cytochrome P450 7A1 | SLC22A5 | Solute carrier family 22 member 5 |
| FABP2 | Fatty acid-binding protein 2 | SLC22A6 | Solute carrier family 22 member 6 |
| GPT | Alanine aminotransferase 1 | SLC22A8 | Solute carrier family 22 member 8 |
| SOD1 | Superoxide dismutase (Cu-Zn) | SLC2A9 | Solute carrier family 2, facilitated glucose transporter member 9 |
| SULT2A1 | Sulfotransferase 2A1 |
Targets of OS
The keyword “osteosarcoma” was used to screen the DisGeNET and GeneCards databases. A total of 4389 relevant targets were identified after integration and removal of duplicates.
Constructing and analyzing the PPI network
Angelicin targets and OS therapeutic targets are shown in a Venn diagram (Figure 2). In total, 19 potential targets of angelicin in the treatment of OS were identified (Supplementary Table 1). The potential therapeutic targets were imported into the STRING online platform to generate PPI network diagrams, which were imported into Cytoscape software for processing and analysis (Figure 3). We determined the correlation coefficients of the PPI network, including degree, closeness centrality, and betweenness centrality (Supplementary Table 2). We used the screening criteria of degree≥degree median, CC≤CC median, and BC≥BC median to identify hub targets and showed median degree = 2, median CC = 0.714286, and median BC = 0. The hub targets, including the apoptosis regulator BAX (BAX), b-cell lymphoma-2 (Bcl-2), baculoviral IAP repeat-containing protein 2 (BIRC 2), and caspase 9 (Casp9), were identified based on degree≥2, CC≤0.714286, and BC≥0.
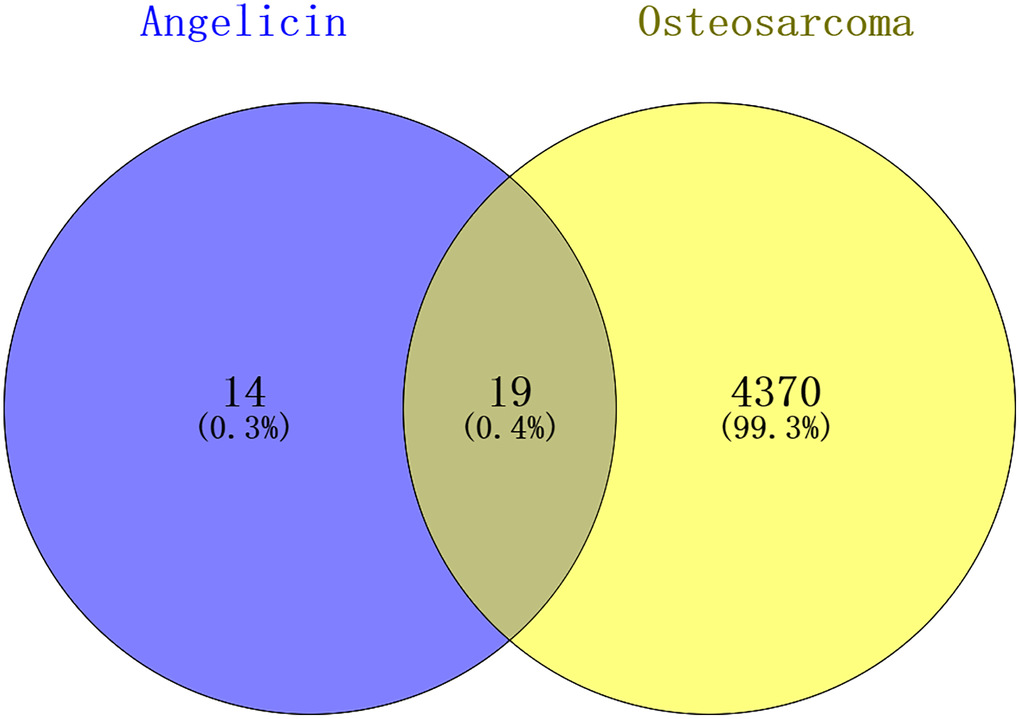
Figure 2. Venn diagram showing the intersection of angelicin-related genes and osteosarcoma-related genes. The angelicin-related targets are shown in the blue circle, and the osteosarcoma-related targets are shown in the yellow circle. The intersection of the two circles indicates potential targets of angelicin in osteosarcoma treatment.
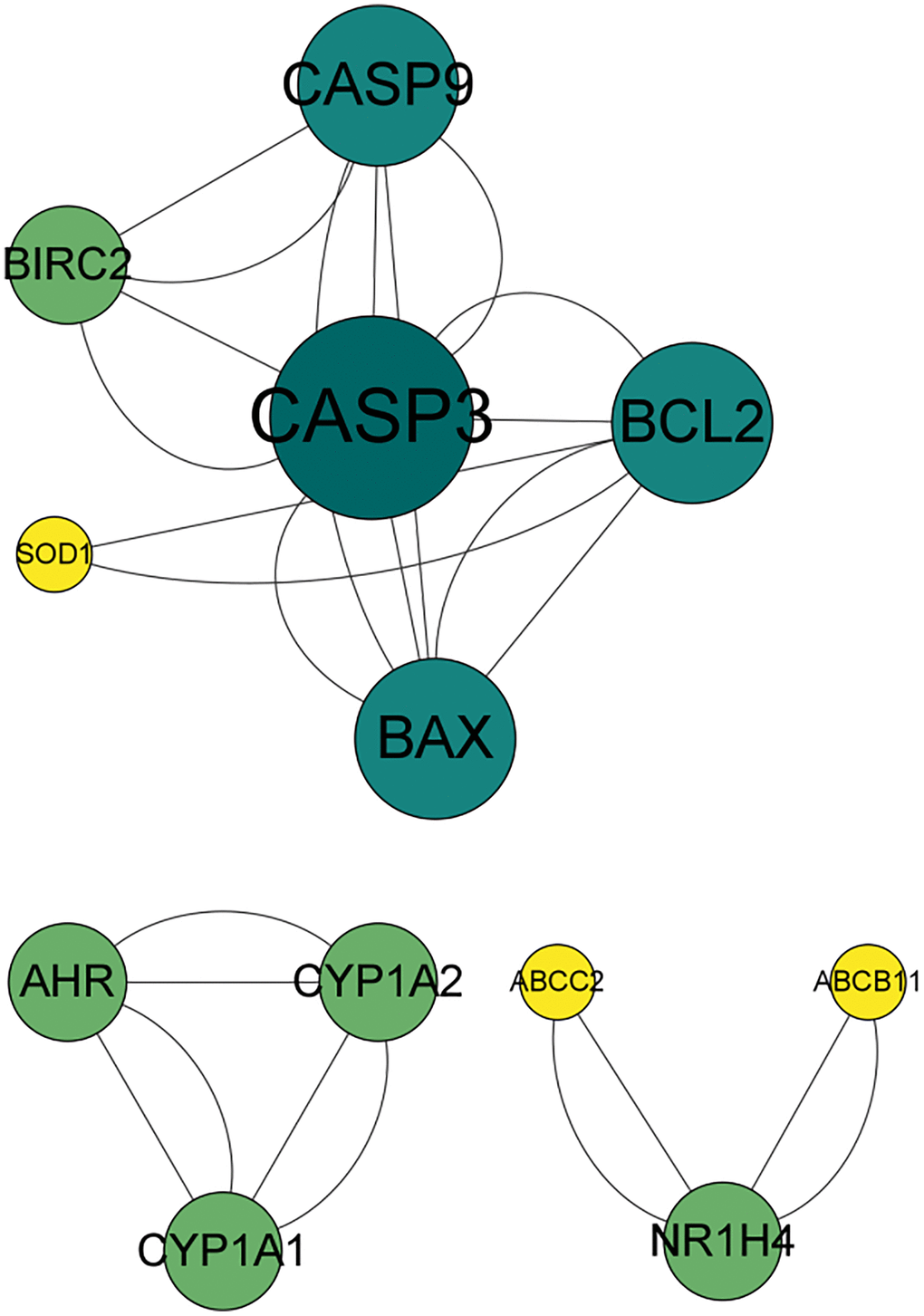
Figure 3. PPI network of the effects of angelicin in the treatment of osteosarcoma. The nodes represent potential therapeutic targets of angelicin in the treatment of osteosarcoma. The larger the node, the darker the color, the higher the target degree, and the higher the number of connections with other nodes.
GO and KEGG analysis
The obtained potential targets of angelicin in the treatment of OS were subjected to GO and KEGG enrichment analyses. The results of biological processes, cellular components, and molecular functions are shown in Supplementary Table 3. The top 10 biological processes and molecular functions and all the cellular components are shown in Figures 4–6; the relationships of the top 10 biological processes and cellular components and all the molecular functions with the potential targets are shown in Figures 4–6, respectively. The potential targets of angelicin in the treatment of OS are shown in Supplementary Table 4, and they were analyzed by KEGG enrichment analysis; the results showed that angelicin impacts osteosarcoma through multiple signaling pathways, such as apoptosis, the p53-related signaling pathway, the NF-kappa B signaling pathway, and the TNF signaling pathway. The top 30 signaling pathways are shown in Figure 7.
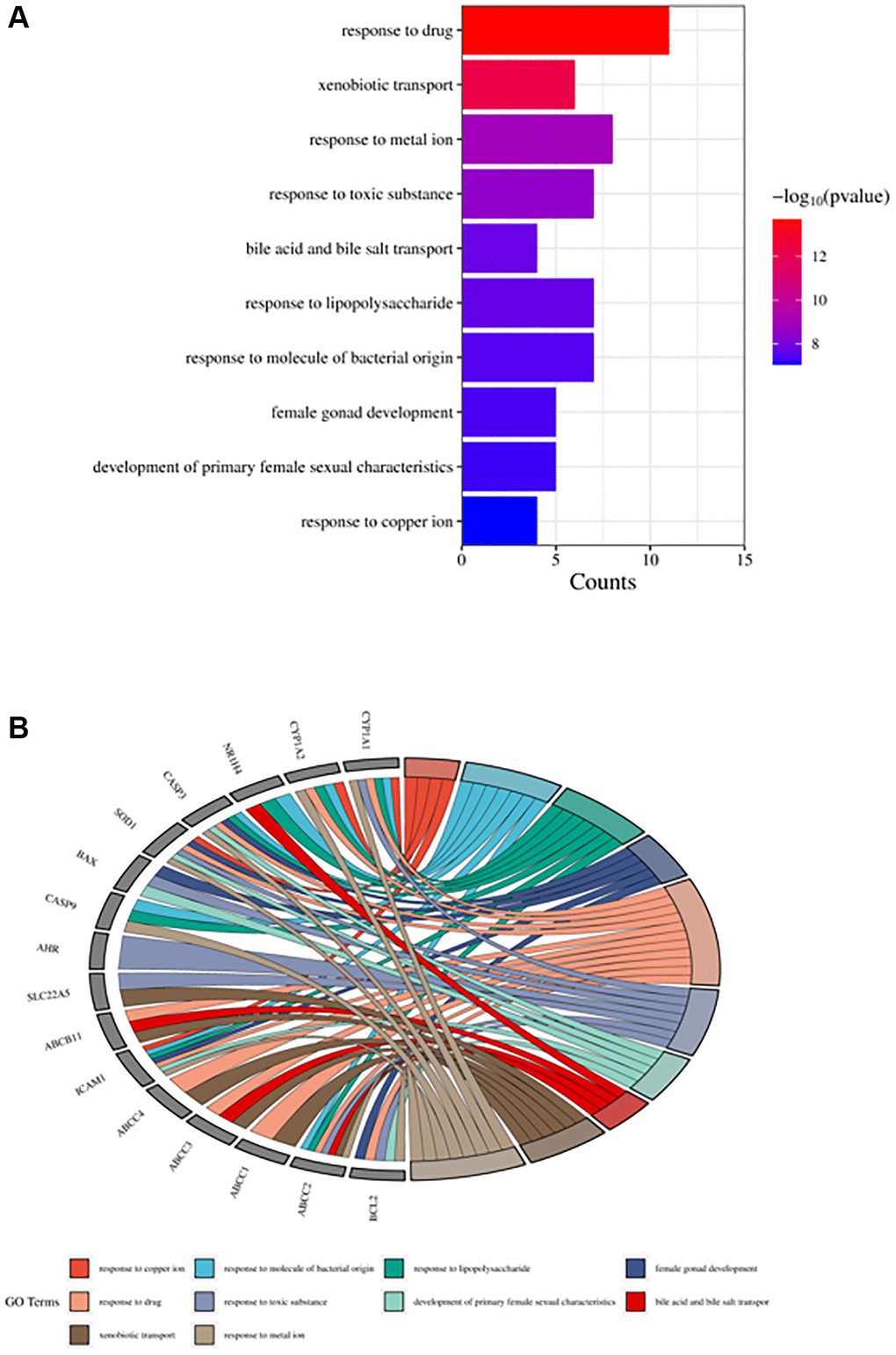
Figure 4. Top ten significant biological process (BP) entries. (A) GO enrichment analysis of therapeutic targets for biological processes. (B) Relationship between the therapeutic targets and biological processes.
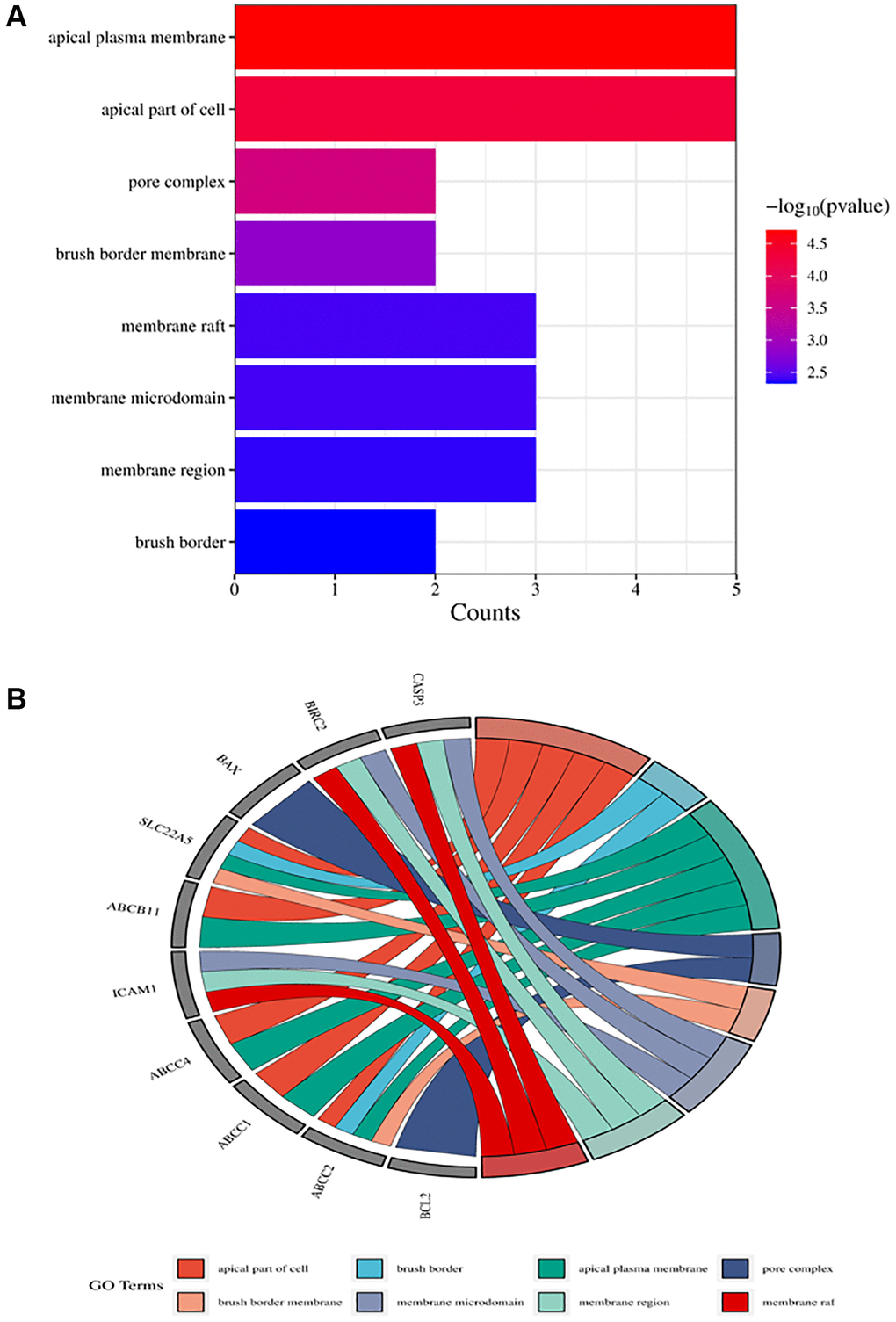
Figure 5. Top ten significant cell component (CC) entries. (A) GO enrichment analysis of therapeutic targets for cell component. (B) Relationship between the therapeutic targets and cell components.
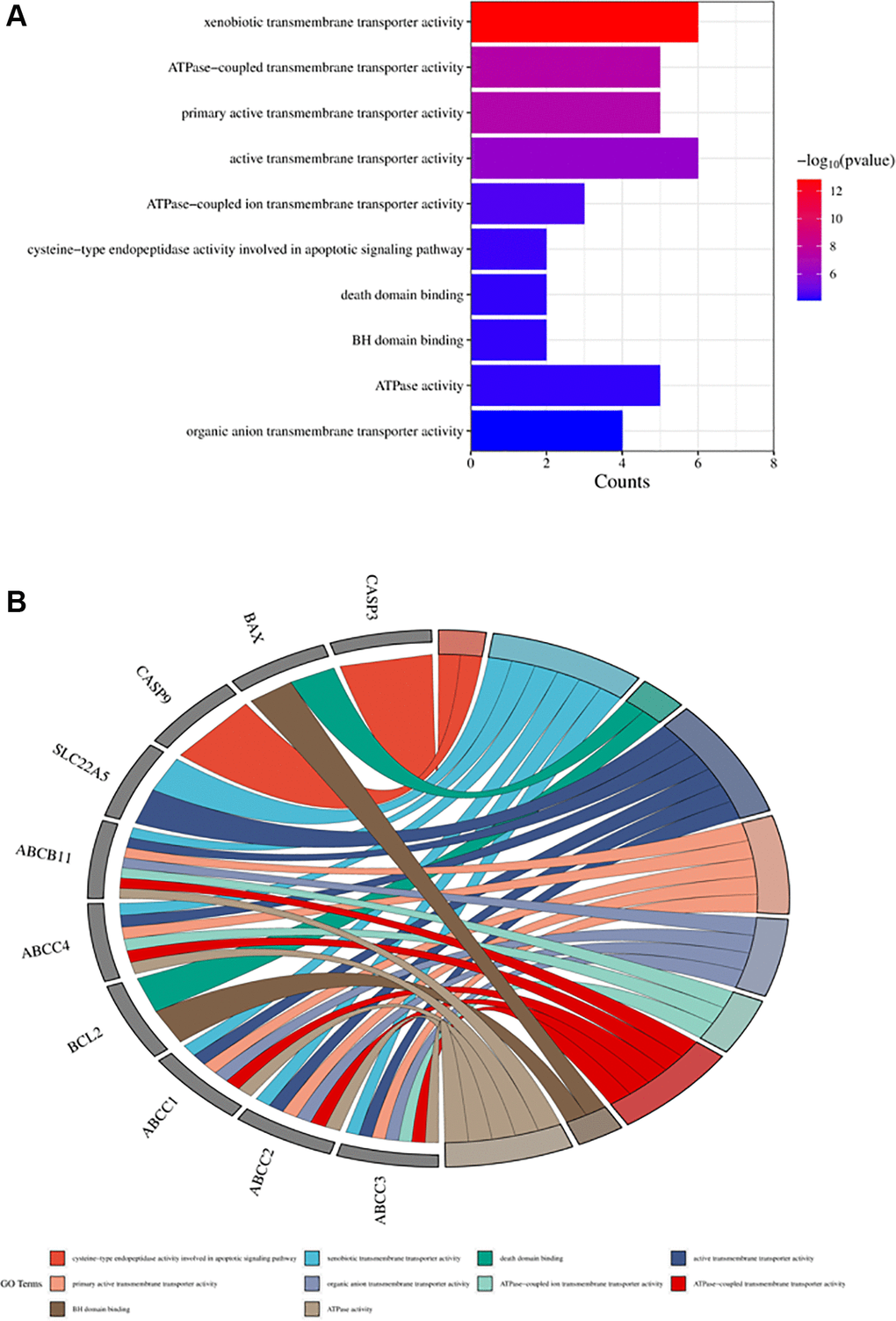
Figure 6. Top ten significant molecular function (MF) entries. (A) GO enrichment analysis of therapeutic targets for molecular function. (B) Relationship between the therapeutic targets and molecular functions.
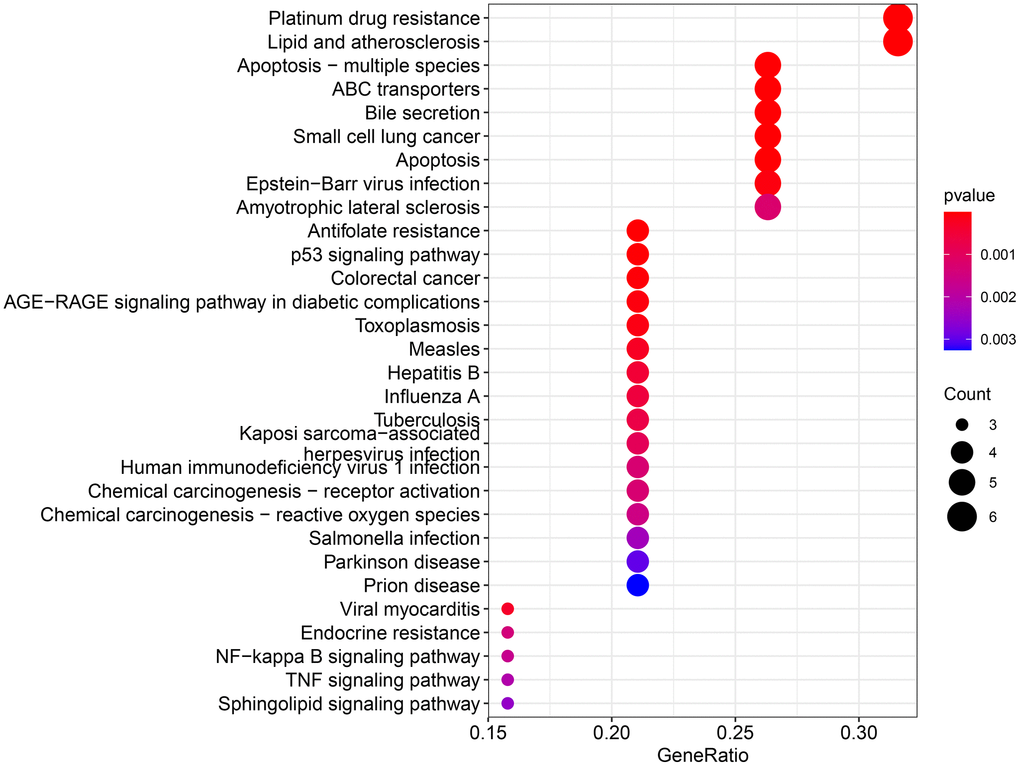
Figure 7. KEGG enrichment analysis for therapeutic targets. The larger the bubble, the more targets are enriched in that pathway, and the redder the color; the smaller the P value, the more meaningful the corresponding signaling pathway.
Molecular docking simulation
Molecular docking of the identified hub genes was performed using AutoDock software. The molecular docking results were visualized using PyMOL software, and the docking results of the four hub targets are shown in Figure 8. The specific details of the docking results are shown in Supplementary Table 4. A binding free energy<−4 kcal/mol when one of the targets was docked with the small molecule of angelicin indicated that the target could freely bind to angelicin. According to molecular docking results, angelicin could bind freely to the hub targets, and the hub targets BAX, BRIC2, BCL2, and Casp9 may be effective targets by which angelicin affects osteosarcoma cells.
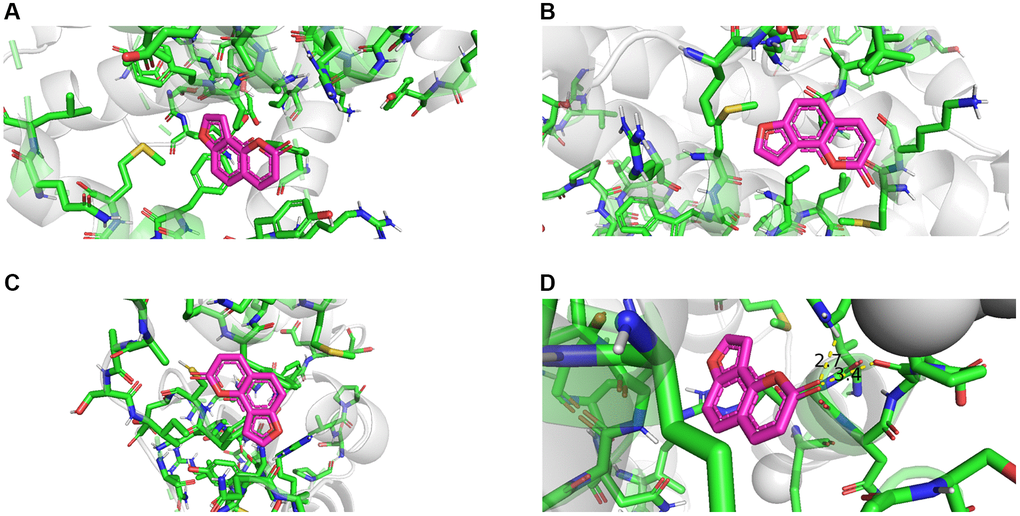
Figure 8. Molecular docking results of angelicin and hub genes. (A) Bcl 2 free energy: −6.2 kcal/mol. (B) BAX free energy: −5.7 kcal/mol. (C) BIRC 2 free energy: −5.3 kcal/mol. (D) Casp9 free energy: -5.8 kcal/mol).
Angelicin inhibits the proliferation of MG63 OS cells in vitro
To investigate the effect of angelicin on the proliferation of OS cells, we used a CCK8 assay to determine MG63 cell viability after treatment with different concentrations of angelicin. The differences in cell viability among the 24 h, 48 h and 72 h groups were statistically significant (P < 0.0001). The differences in cell viability between the 50 μmol/L and 200 μmol/L groups were statistically significant (P < 0.0001) (Figure 9). The viability of MG63 cells gradually decreased as the angelicin concentration increased and the culture time increased. These results indicated that angelicin inhibited the proliferation of MG63 cells in a dose-dependent and time-dependent manner.
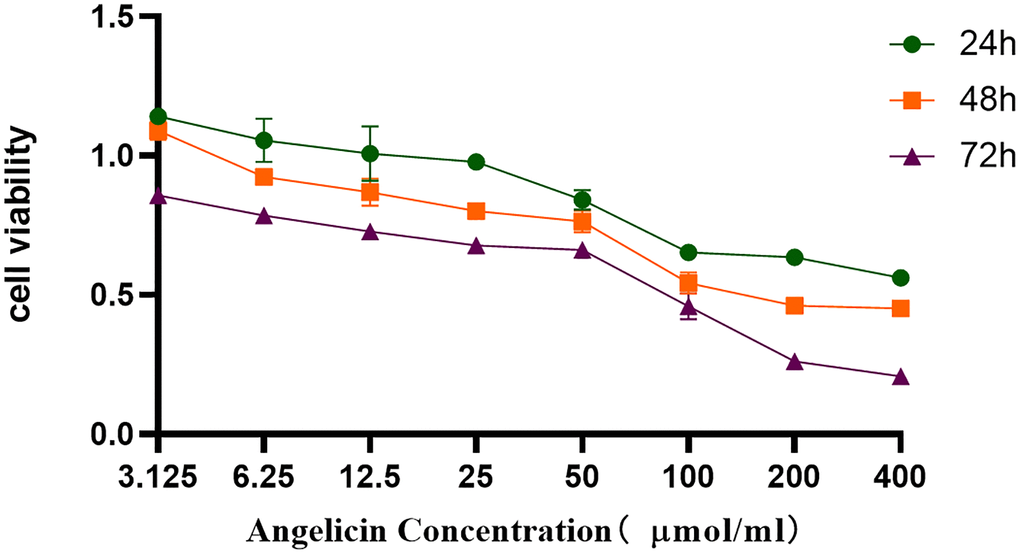
Figure 9. CCK8 assay. Angelicin inhibited the proliferation of osteosarcoma MG63 cells. The viability of osteosarcoma MG63 cells was measured at different time points (24 h, 48 h, or 72 h) of culture with media containing different angelicin concentrations (3.125–400 μmol/ml), and the results are shown as the mean ± standard deviation.
Angelicin induces the apoptosis of MG63 OS cells in vitro
We used flow cytometry to analyze the apoptosis of MG63 cells after treatment with different concentrations of angelicin. The apoptosis rates of the angelicin-treated groups were statistically significantly different compared with that of the control group (P < 0.05). The apoptosis rate of OS MG63 cells gradually increased with increasing angelicin concentrations (Figure 10). These results suggest that angelicin promotes the apoptosis of OS MG63 cells in a dose-dependent manner.
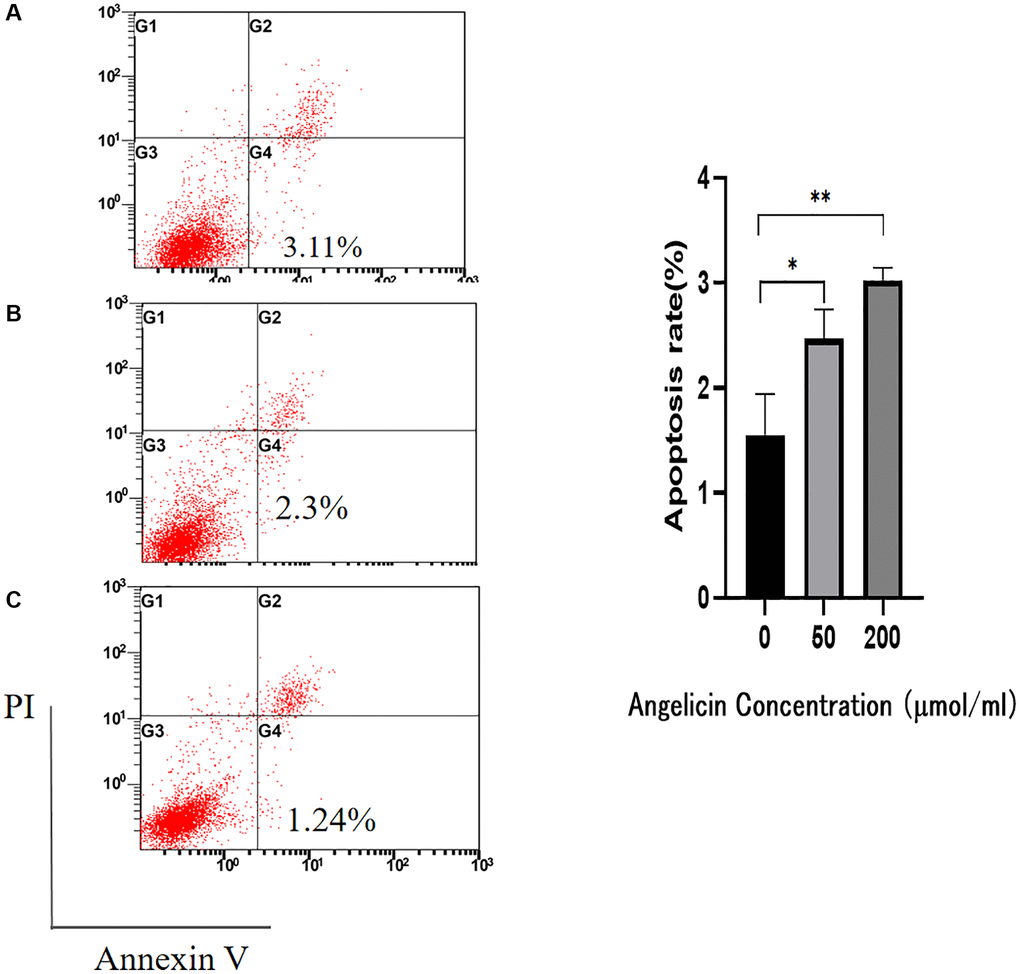
Figure 10. Flow cytometry. Angelicin promotes apoptosis in osteosarcoma MG63 cells. The apoptosis rate of osteosarcoma MG63 cells was measured after 24 h of culture with media containing different concentrations of angelicin (A) 0 μmol/ml, (B) 50 μmol/ml, and (C) 200 μmol/ml). The results of quantitative analysis are shown as the mean ± S.D. of three independent experiments. *Significant difference compared to the control group: p < 0.05, **p < 0.01.
Angelicin inhibits the migration of OS MG63 cells in vitro
We performed the wound healing assay to evaluate the migration rate of MG63 cells, and the results showed that the differences in the wound healing rate among the different concentration groups were statistically significant (P < 0.05). The wound healing rate of OS cells gradually decreased with increasing angelicin concentration (Figure 11), which indicates that angelicin inhibited the migration of OS MG63 cells in a dose-dependent manner.

Figure 11. Wound healing experiment. Angelicin inhibits the migration of osteosarcoma MG63 cells. MG63 osteosarcoma cell wound healing results at different time points (0 h, 24 h, or 48 h) of culture with media containing different angelicin concentrations (0 μmol/ml, 50 μmol, or 200 μmol/ml). The results of quantitative analysis are shown as the mean ± S.D. of three independent experiments. *Significant difference between the two groups: p < 0.05, ****p < 0.0001.
RT-PCR experiments
To measure the mRNA expression of pivotal targets in OS MG63 cells after treatment with different concentrations of angelicin, we performed RT-PCR experiments, and the results demonstrated that the expression of BAX and Casp9 gradually increased, while the expression of the antiapoptotic proteins Bcl-2 and BIRC2 gradually decreased with increasing angelicin concentration. The differences in mRNA expression among the different concentration groups were statistically significant (P < 0.05), and the results are shown in Figure 12.
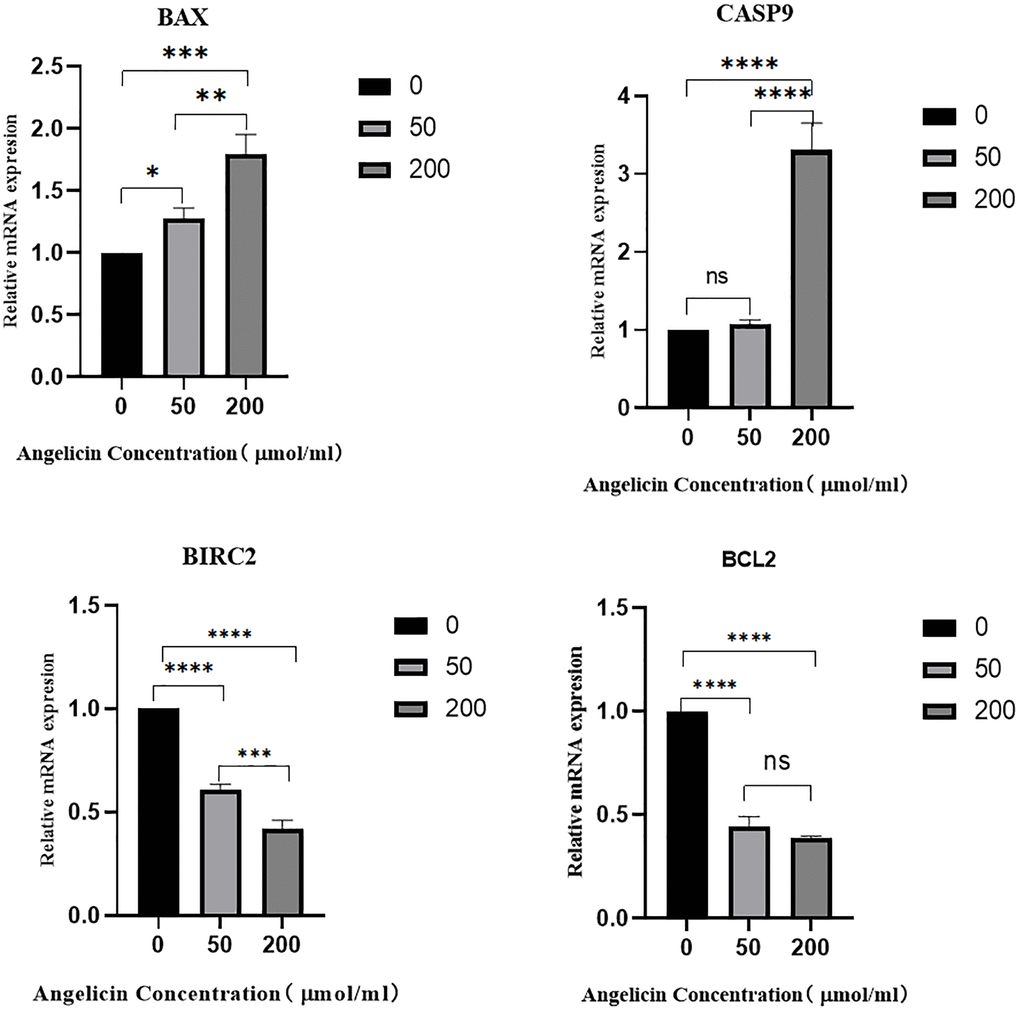
Figure 12. The effect of angelicin on the expression of hub target genes in osteosarcoma MG63 cells. The mRNA expression of the hub genes in osteosarcoma MG63 cells was measured after 24 h of culture with media containing different concentrations (0 μmol/ml, 50 μmol/ml, or 200 μmol/ml) of angelicin; (A) BAX, (B) Casp9, (C) BIRC2, (D) Bcl2; ns (No significance): No statistically significant difference between the two groups. When P < 0.05, *the difference between the two groups is statistically significant. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
Discussion
Osteosarcoma is the most common malignant tumor, accounting for approximately 55% of all bone tumors [15]. OS is more common in children and adolescents and poses great risks to human health [1, 16]. The current clinical treatment is so-called “sandwich therapy”, which consists of neoadjuvant chemotherapy, radical resection surgery and adjuvant chemotherapy [4, 17]. The 5-year overall survival rate of primary OS patients who receive classical treatment is approximately 60–70% [6]; however, OS has a high probability of local recurrence and distant metastasis [18]. Approximately 15–20% of OS patients have distant metastases when they are first diagnosed [19], and pulmonary metastasis is the most common metastatic event [20]. The 5-year survival rate decreases to less than 15% once distant metastasis occurs [3]. Although new treatments and pathological mechanisms are being explored, no significant progress has been made over the past several decades [6]. In this research, we showed that angelicin exerts an inhibitory effect on osteosarcoma cells by suppressing proliferation and migration and promoting apoptosis. Angelicin is one of the furanocoumarins found in many Chinese herbal medicines, such as Angelica archangelica and Paeoniae Radix Alba [9, 21]. Existing studies have shown that angelicin has antitumor effects on many types of malignancies, such as liver cancer and lung cancer [9, 10]. Inspired by the above research, we first investigated the role of angelicin in OS.
In this study, we intersected angelicin therapeutic targets and OS-related targets that were identified in several databases and found potential therapeutic targets through which angelicin functions in the treatment of OS. Furthermore, we found the molecular hub targets of angelicin. Then, we screened the hub targets, including Casp9, BIRC2, BAX, and BCL2, of which BCL2 and Casp9 are proapoptotic proteins, and BAX and BIRC2 are antiapoptotic proteins [22]. All four hub target genes are closely associated with apoptosis. Moreover, the GO and KEGG enrichment results showed that the potential targets of angelicin in the treatment of OS were enriched in multiple signaling pathways. In addition, we found that the hub targets are involved in the transduction of multiple signaling pathways. For example, BAX is involved in the signaling of apoptosis, apoptosis-multiple species, and the p53 signaling pathway. BCL2 is involved in signaling pathways such as apoptosis, apoptosis-multiple species, the p53 signaling pathway, and the NF-κB signaling pathway. BIRC2 is involved in apoptosis, apoptosis-multiple species, the NF-κB signaling pathway, and the TNF signaling pathway. Numerous studies have demonstrated that multiple signaling pathways are involved in regulating the multiple biological behaviors of osteosarcoma cells [23–27]. The hub targets connect multiple signaling pathways to form an interactive signaling network, through which Angelica affects the biological behaviors of osteosarcoma. When the NF-κB signaling pathway is inhibited, the biological behaviors of osteosarcoma, such as proliferation, migration, and apoptosis, are suppressed [25, 28, 29]. When p53 is activated, it inhibits the proliferation and migration of osteosarcoma cells and promotes the apoptosis of osteosarcoma cells [27, 30, 31]. We found that angelicin can inhibit the proliferation and migration of osteosarcoma by modulating the signaling network formed by connections of hub targets. Based on these results, we further explored the effect of angelicin on OS. Since the hub targets are all apoptosis-related proteins and are enriched in several apoptosis-related signaling pathways according to the KEGG results, we focused on verifying the effect of angelicin on the apoptosis of osteosarcoma cells. The results showed that angelicin promotes OS cell apoptosis in a dose-dependent manner. The in vitro results further confirmed the network pharmacological results. It was also demonstrated that angelicin inhibits osteosarcoma cell proliferation and migration in a dose-dependent manner and promotes osteosarcoma cell apoptosis in a dose-dependent manner.
The network pharmacology, molecular docking, and in vitro experiments described above show that angelicin inhibits OS cell proliferation and migration and promotes OS cell apoptosis upregulating Casp9 and Bcl-2 gene expression and downregulating BAX and BIRC2 gene expression, thereby regulating apoptosis in multiple species, apoptosis, the P53 signaling pathway, the NF-κB signaling pathway, and the TNF signaling pathway to form a signaling pathway network.
Through pharmacological and molecular docking assays, we discovered the regulatory effect of angelicin on osteosarcoma cells. Based on these results, we verified with in vitro experiments that angelicin inhibits osteosarcoma cell proliferation and migration and promotes osteosarcoma cell apoptosis. We are the first to observe these results. Network pharmacology is an efficient tool that is used for the investigation of the mechanisms of action of drugs. However, it is clear that the present study still has some limitations. This study did not examine the specific signaling pathways that are regulated by angelicin in osteosarcoma cells, but we have predicted which pathways are targeted by angelicin via network pharmacology; molecular biological validation is our next step. In addition, in vivo experiments were not conducted in this study, and the inhibitory effect of angelicin on osteosarcoma in vivo was not explored; we will pursue these experiments in future work. Third, the exploration of the loci of apoptotic proteins on which angelicin acts is another focus of our future work.
Conclusion
We found that via the regulation of multiple signaling pathways, angelicin inhibited osteosarcoma cell proliferation in a time- and dose-dependent manner, inhibited osteosarcoma cell migration in a dose-dependent manner, and promoted osteosarcoma cell apoptosis in a dose-dependent manner. Angelicin could be an alternative medicine for the treatment of osteosarcoma, and it still needs to be explored in depth.
Abbreviations
RT-PCR: reverse transcription-polymerase chain reaction; BAX: BCL2-associated X protein; BCL-2: B-cell lymphoma-2; BIRC 2: baculoviral IAP repeat-containing protein 2; Casp9: caspase 9; OS: osteosarcoma; NIH: National Institutes of Health; BP: biological process; CC: cellular component; MF: molecular function; GO: Gene Ontology; KEGG: Kyoto Encyclopedia of Genes and Genomes; PPI: protein-protein interaction; CCK8: Cell Counting Kit-8; DMSO: dimethyl sulfoxide; MEM: minimum essential medium; PI: propidium iodide; FITC: fluorescein isothiocyanate; P53: tumor antigen p53; NF-κB: NF-kappa-B; TNF: tumor necrosis factor; MEM: modified Eagle’s medium; FBS: fetal bovine serum(FBS).
Author Contributions
Yu Jin designed this study. Yafang Zhang, Junqiang Wei, Mingze Song, Xiangyu Xiao and Yange Zhang conducted this study and analyzed the data. Yafang Zhang and Junqiang Wei drafted the manuscript. Yu Jin revised this study. Y Zhang, J Wei and Y Zhang did experiments.
Acknowledgments
We are very grateful for the contributions of the Pubchem database, symMap database,the DisGeNET database and the GeneCards database that provide information on cancer research, as well as all colleagues involved in the study.
Conflicts of Interest
The authors have declared that no competing interest exists.
Funding
This study was supported by the 2020 Chengde Science and Technology Research and Development Program Project (202006A163).
References
- 1. Ritter J, Bielack SS. Osteosarcoma. Ann Oncol. 2010 (Suppl 7); 21:vii320–5. https://doi.org/10.1093/annonc/mdq276 [PubMed]
- 2. Eaton BR, Schwarz R, Vatner R, Yeh B, Claude L, Indelicato DJ, Laack N. Osteosarcoma. Pediatr Blood Cancer. 2021 (Suppl 2); 68:e28352. https://doi.org/10.1002/pbc.28352 [PubMed]
- 3. Lindsey BA, Markel JE, Kleinerman ES. Osteosarcoma Overview. Rheumatol Ther. 2017; 4:25–43. https://doi.org/10.1007/s40744-016-0050-2 [PubMed]
- 4. Harrison DJ, Geller DS, Gill JD, Lewis VO, Gorlick R. Current and future therapeutic approaches for osteosarcoma. Expert Rev Anticancer Ther. 2018; 18:39–50. https://doi.org/10.1080/14737140.2018.1413939 [PubMed]
- 5. Kansara M, Teng MW, Smyth MJ, Thomas DM. Translational biology of osteosarcoma. Nat Rev Cancer. 2014; 14:722–35. https://doi.org/10.1038/nrc3838 [PubMed]
- 6. Chen C, Xie L, Ren T, Huang Y, Xu J, Guo W. Immunotherapy for osteosarcoma: Fundamental mechanism, rationale, and recent breakthroughs. Cancer Lett. 2021; 500:1–10. https://doi.org/10.1016/j.canlet.2020.12.024 [PubMed]
- 7. Sayles LC, Breese MR, Koehne AL, Leung SG, Lee AG, Liu HY, Spillinger A, Shah AT, Tanasa B, Straessler K, Hazard FK, Spunt SL, Marina N, et al. Genome-Informed Targeted Therapy for Osteosarcoma. Cancer Discov. 2019; 9:46–63. https://doi.org/10.1158/2159-8290.CD-17-1152 [PubMed]
- 8. Harris MA, Hawkins CJ. Recent and Ongoing Research into Metastatic Osteosarcoma Treatments. Int J Mol Sci. 2022; 23:3817. https://doi.org/10.3390/ijms23073817 [PubMed]
- 9. Wang F, Li J, Li R, Pan G, Bai M, Huang Q. Angelicin inhibits liver cancer growth in vitro and in vivo. Mol Med Rep. 2017; 16:5441–9. https://doi.org/10.3892/mmr.2017.7219 [PubMed]
- 10. Li G, He Y, Yao J, Huang C, Song X, Deng Y, Xie S, Ren J, Jin M, Liu H. Angelicin inhibits human lung carcinoma A549 cell growth and migration through regulating JNK and ERK pathways. Oncol Rep. 2016; 36:3504–12. https://doi.org/10.3892/or.2016.5166 [PubMed]
- 11. Min KJ, Um HJ, Seo SU, Woo SM, Kim S, Park JW, Lee HS, Kim SH, Choi YH, Lee TJ, Kwon TK. Angelicin potentiates TRAIL-induced apoptosis in renal carcinoma Caki cells through activation of caspase 3 and down-regulation of c-FLIP expression. Drug Dev Res. 2018; 79:3–10. https://doi.org/10.1002/ddr.21414 [PubMed]
- 12. Liu F, Sun GQ, Gao HY, Li RS, Soromou LW, Chen N, Deng YH, Feng HH. Angelicin regulates LPS-induced inflammation via inhibiting MAPK/NF-κB pathways. J Surg Res. 2013; 185:300–9. https://doi.org/10.1016/j.jss.2013.05.083 [PubMed]
- 13. Luo TT, Lu Y, Yan SK, Xiao X, Rong XL, Guo J. Network Pharmacology in Research of Chinese Medicine Formula: Methodology, Application and Prospective. Chin J Integr Med. 2020; 26:72–80. https://doi.org/10.1007/s11655-019-3064-0 [PubMed]
- 14. Zhou Z, Chen B, Chen S, Lin M, Chen Y, Jin S, Chen W, Zhang Y. Applications of Network Pharmacology in Traditional Chinese Medicine Research. Evid Based Complement Alternat Med. 2020; 2020:1646905. https://doi.org/10.1155/2020/1646905 [PubMed]
- 15. Li HB, Chen JK, Su ZX, Jin QL, Deng LW, Huang G, Shen JN. Cordycepin augments the chemosensitivity of osteosarcoma to cisplatin by activating AMPK and suppressing the AKT signaling pathway. Cancer Cell Int. 2021; 21:706. https://doi.org/10.1186/s12935-021-02411-y [PubMed]
- 16. Simpson E, Brown HL. Understanding osteosarcomas. JAAPA. 2018; 31:15–9. https://doi.org/10.1097/01.JAA.0000541477.24116.8d [PubMed]
- 17. Rothzerg E, Pfaff AL, Koks S. Innovative approaches for treatment of osteosarcoma. Exp Biol Med (Maywood). 2022; 247:310–6. https://doi.org/10.1177/15353702211067718 [PubMed]
- 18. Bloomingdale P, Meregalli C, Pollard K, Canta A, Chiorazzi A, Fumagalli G, Monza L, Pozzi E, Alberti P, Ballarini E, Oggioni N, Carlson L, Liu W, et al. Systems Pharmacology Modeling Identifies a Novel Treatment Strategy for Bortezomib-Induced Neuropathic Pain. Front Pharmacol. 2022; 12:817236. https://doi.org/10.3389/fphar.2021.817236 [PubMed]
- 19. Wang Z, Li H, Yan J, Liu Y. Flavonoid compound breviscapine suppresses human osteosarcoma Saos-2 progression property and induces apoptosis by regulating mitochondria-dependent pathway. J Biochem Mol Toxicol. 2021; 35:e22633. https://doi.org/10.1002/jbt.22633 [PubMed]
- 20. Meazza C, Scanagatta P. Metastatic osteosarcoma: a challenging multidisciplinary treatment. Expert Rev Anticancer Ther. 2016; 16:543–56. https://doi.org/10.1586/14737140.2016.1168697 [PubMed]
- 21. Lapalikar GV, Taylor MC, Warden AC, Scott C, Russell RJ, Oakeshott JG. F420H2-dependent degradation of aflatoxin and other furanocoumarins is widespread throughout the actinomycetales. PLoS One. 2012; 7:e30114. https://doi.org/10.1371/journal.pone.0030114 [PubMed]
- 22. Shan HJ, Gu WX, Duan G, Chen HL. Fat mass and obesity associated (FTO)-mediated N6-methyladenosine modification of Krüppel-like factor 3 (KLF3) promotes osteosarcoma progression. Bioengineered. 2022; 13:8038–50. https://doi.org/10.1080/21655979.2022.2051785 [PubMed]
- 23. Chen YQ, Yang TQ, Zhou B, Yang MX, Feng HJ, Wang YL. HOXA5 overexpression promotes osteosarcoma cell apoptosis through the p53 and p38α MAPK pathway. Gene. 2019; 689:18–23. https://doi.org/10.1016/j.gene.2018.11.081 [PubMed]
- 24. Zhou JL, Deng S, Fang HS, Yu G, Peng H. Hsa-let-7g promotes osteosarcoma by reducing HOXB1 to activate NF-kB pathway. Biomed Pharmacother. 2019; 109:2335–41. https://doi.org/10.1016/j.biopha.2018.11.026 [PubMed]
- 25. Hu Z, Li L, Lan W, Wei X, Wen X, Wu P, Zhang X, Xi X, Li Y, Wu L, Li W, Liao X. Enrichment of Wee1/CDC2 and NF-κB Signaling Pathway Constituents Mutually Contributes to CDDP Resistance in Human Osteosarcoma. Cancer Res Treat. 2022; 54:277–93. https://doi.org/10.4143/crt.2021.320 [PubMed]
- 26. Chen H, Pan R, Li H, Zhang W, Ren C, Lu Q, Chen H, Zhang X, Nie Y. CHRDL2 promotes osteosarcoma cell proliferation and metastasis through the BMP-9/PI3K/AKT pathway. Cell Biol Int. 2021; 45:623–32. https://doi.org/10.1002/cbin.11507 [PubMed]
- 27. Wu C, Li Z, Feng G, Wang L, Xie J, Jin Y, Wang L, Liu S. Tumor suppressing role of serum-derived exosomal microRNA-15a in osteosarcoma cells through the GATA binding protein 2/murine double minute 2 axis and the p53 signaling pathway. Bioengineered. 2021; 12:8378–95. https://doi.org/10.1080/21655979.2021.1987092 [PubMed]
- 28. Li R, Shi Y, Zhao S, Shi T, Zhang G. NF-κB signaling and integrin-β1 inhibition attenuates osteosarcoma metastasis via increased cell apoptosis. Int J Biol Macromol. 2019; 123:1035–43. https://doi.org/10.1016/j.ijbiomac.2018.11.003 [PubMed]
- 29. Kumar A, Kaur S, Dhiman S, Singh PP, Bhatia G, Thakur S, Tuli HS, Sharma U, Kumar S, Almutary AG, Alnuqaydan AM, Hussain A, Haque S, et al. Targeting Akt/NF-κB/p53 Pathway and Apoptosis Inducing Potential of 1,2-Benzenedicarboxylic Acid, Bis (2-Methyl Propyl) Ester Isolated from Onosma bracteata Wall. against Human Osteosarcoma (MG-63) Cells. Molecules. 2022; 27:3478. https://doi.org/10.3390/molecules27113478 [PubMed]
- 30. Zhou F, Dou X, Li C. CKB affects human osteosarcoma progression by regulating the p53 pathway. Am J Cancer Res. 2022; 12:4652–65. [PubMed]
- 31. Zhao L, Li J, Sun ZB, Sun C, Yu ZH, Guo X. Saikosaponin D inhibits proliferation of human osteosarcoma cells via the p53 signaling pathway. Exp Ther Med. 2019; 17:488–94. https://doi.org/10.3892/etm.2018.6969 [PubMed]