



Introduction
Peripheral arterial disease (PAD) is primarily seen as a manifestation of systemic atherosclerosis, presenting as chronic arterial occlusive disease in the lower extremities [1]. It is considered the third most prevalent manifestation of atherosclerotic vascular disease, with coronary artery disease (CAD) and stroke being the two leading presentations [2]. PAD can lead to intermittent claudication (IC), chronic limb-threatening ischemia (CLTI), limb functional impairment, and even life-threatening cardiovascular events [3, 4], and this mainly includes IC and CLTI. IC is clearly more common, characterized by pain in the calf, thigh, or buttock muscles that can be induced by repeated exercise and relieved by rest. CLTI is characterized by rest pain or tissue loss in the lower limbs. Compared to CLTI, the natural history of IC is benign, and few IC patients progress to CLTI. The development of PAD is influenced by various risk factors, such as smoking, hypertension, hyperlipidemia, and diabetes [5]. PAD is a widespread condition that affects more than 200 million individuals across the globe [6], and the prevalence of PAD ranges from 3% to 10%. However, based on data from older individuals, the prevalence among the elderly population can be as high as 15% to 20%. Furthermore, the current prevalence rates are even higher [7]. It has been found that the incidence of PAD increases with age, obesity, and diabetes [8]. PAD imposes a significant burden on affected individuals, families, and even the entire society. Therefore, it underscores the necessity of identifying risk factors associated with its development.
While the exact cause and underlying mechanisms of PAD remain incompletely understood, it is generally acknowledged to be a multifactorial disease with various contributing factors. The human gastrointestinal system provides a home to trillions of bacteria and archaea, which coexist with the human body in a mutually beneficial relationship that is crucial for human health [9]. Intestinal microbiota plays a role in various physiological functions, including maintaining metabolic stability, regulating immune responses, and resisting infections [10]. A growing body of research has established a link between the gut microbiota and the development and advancement of diseases, resulting in its acknowledgment as an endocrine organ [11]. Many studies and evidence suggest that the gut microbiota and its metabolites are associated with the formation and development of atherosclerosis (AS), including the formation and progression of atherosclerotic plaques, endothelial dysfunction, and thrombus formation [12, 13]. Atherosclerotic plaques themselves are a microbiota environment, containing microorganisms such as Streptococcus, Pseudomonas, Klebsiella, Veillonella, and Chlamydia pneumoniae [14]. A meta-analysis has revealed that shifts in the gut microbiota composition are linked to changes in the levels of bacterial metabolites, some of which have been shown to have atherosclerotic effects on endothelial cells [15]. In cross-sectional studies, it has been shown that symptomatic AS patients have higher abundances of Collinsella, Enterobacteriaceae, Streptococcaceae, and Klebsiella in their gut microbiota compared to healthy controls. On the other hand, the abundances of bacteria that produce short-chain fatty acids (SCFA) such as Faecalibacterium, Roseburia, and Ruminococcaceae are lower in symptomatic AS patients [16]. Additionally, there has been extensive research into the functions of specific metabolites originating from the gut microbiota, including trimethylamine N-oxide (TMAO) from carnitine metabolism, phenylacetylglutamine from phenylalanine metabolism, and bile acids from lipid metabolism, particularly in relation to AS. Most studies have been unable to establish a direct link between the composition of the gut microbiota and AS-related plaque vulnerability, rupture, or cardiovascular events. Currently, there is a lack of evidence for causal factors in this regard.
Although there have been rapid advancements in genomics, metabolomics, and other technologies that have revealed a strong correlation between gut microbiota and PAD, the exact causal relationship between them remains unclear [17, 18]. This method identifies and measures the causal impact of exposures on outcomes through the use of genetic variants as instrumental variables (IVs) [19, 20]. Because alleles are randomly distributed from parents to offspring, undergo independent assortment, and maintain genetic stability after birth, Mendelian randomization (MR), akin to randomized controlled trials (RCTs), can mitigate biases arising from conventional confounding factors (e.g., environmental influences, population traits, and dietary behaviors) and reverse causation [21, 22]. In this research, a two-sample MR analysis was conducted, leveraging summary statistics data from the Genome-wide Association Studies (GWAS) of the MiBioGen and FinnGen consortia, to explore the causal connection between gut microbiota and PAD.
Materials and Methods
Research design
We employed a two-sample MR design using GWAS meta-analysis data to furnish strong support for the causal link between gut microbiota and PAD. The study design and reporting of this study are in accordance with STROBE-MR guidelines [23]. To mitigate the impact of confounding variables on the outcomes, the MR approach must adhere to three essential assumptions (Figure 1). (1) Choose SNPs that exhibit significant associations with gut microbiota as IVs; (2) IVs should be independent, meaning they should not be correlated with other confounding factors like age and smoking; (3) IVs should solely be linked to the outcome via their connection with the exposure and must not affect the outcome through any alternative pathways [24].
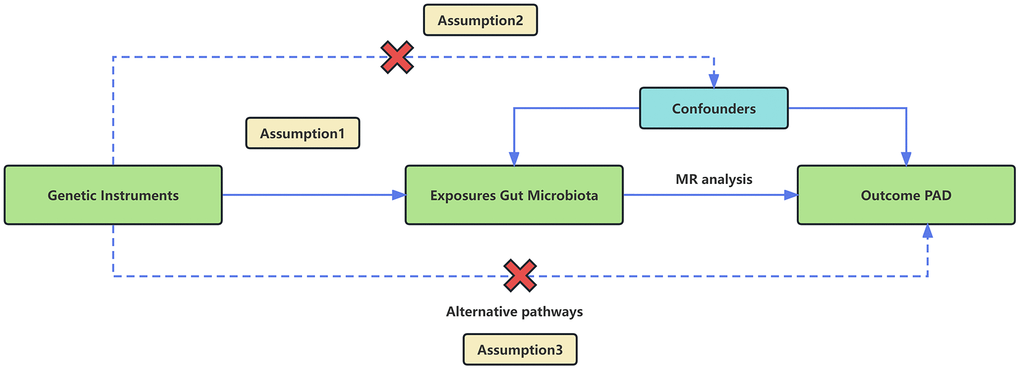
Figure 1. An overview of the MR analysis and its three primary assumptions.
Data source
Genetic information pertaining to the gut microbiota is derived from an extensive case-control GWAS meta-analysis carried out by the international MiBioGen consortium [25]. The study includes 25 cohorts from countries such as the United States, Italy, and South Korea, with a total of 18,340 participants. The primary objective is to pinpoint genetic loci that impact the gut microbiota’s relative abundance through the examination of 16S rRNA sequencing profiles of the participants. GWAS summary statistics pertaining to PAD were extracted from the FinnGen Consortium R6 release dataset, comprising 9,021 PAD cases and 244,907 individuals serving as controls [26].
Selection of IVs
Employ the subsequent quality control techniques to carefully choose suitable genetic IVs, thereby safeguarding the integrity and precision of causal inferences regarding the connection between gut microbiota composition and PAD risk. (1) In accordance with the typical practice observed in most MR investigations involving gut microbiota [27, 28], we established the significance threshold at P < 1.0×10^(-5) to detect an ample quantity of potential instrumental variables. This decision takes into account that the number of loci identified for gut microbiota is relatively limited [29]; (2) SNP variants associated with each bacterial taxonomic group are grouped together, and only independent SNPs are retained. The threshold for linkage disequilibrium (LD) aggregation is defined as r^2 < 0.001, and this aggregation is performed over a distance of 10,000 kb [30]; (3) In cases where palindrome SNPs are detected, allele frequency data is utilized to deduce the alleles on the forward strand; (4) An F-statistic threshold of >10 is set as the threshold for strong IVs. Otherwise, if the IVs are found to have a weak association with the exposure, they are considered to be excluded [31]. (5) To exclude SNPs linked to potential confounding factors like smoking and occupational exposure, we employed PhenoScanner to identify all SNPs meeting the specified criteria [32].
Reverse MR data
The data source for reverse MR corresponds to that used for forward MR. In this scenario, we regard PAD as the exposure and extract SNPs closely linked to PAD as the exposure (p < 5*10^-8). Similar to forward MR, we also conducted a selection process in reverse MR, which includes eliminating linkage disequilibrium, palindromic sequences, and weakly correlated variables, and excluding SNPs with confounding factors. We employed the significant genera identified from the forward MR analysis as our results and subsequently conduct a two-sample MR analysis to establish the causal connection between PAD and gut microbiota.
MR analysis
In this study, the primary analytical approach used to establish causal relationships was the inverse variance weighted (IVW) method [33]. This method calculates weighted averages based on the reciprocals of variances, under the assumption that all instrumental variables are valid. The results of this method are deemed the most accurate when there is neither heterogeneity nor horizontal pleiotropy. Furthermore, Supplementary Methods including MR-Egger regression, weighted median (WM), simple mode (SM), and weighted mode estimation are utilized alongside the IVW results [34]. MR-Egger has strong applicability for testing the hypothesis and can tolerate heterogeneity in more than 50% of the SNPs [35]. Weighted median is the median of the distribution function obtained by weighting all individual SNP effect values according to their weights. Robust estimation of weighted median can be obtained when at least 50% of the information comes from valid instrumental variables [36]. The principle of simple mode is to estimate causal effects directly using the effect estimate of the instrumental variable when there is only one instrumental variable available [37]. The weighted mode method is a commonly used statistical method for calculating the mode of a set of data. By considering the weight of each value, it can provide a more accurate description of the central tendency of the dataset [38].
Horizontal pleiotropy and heterogeneity evaluation
To evaluate the robustness of the findings, additional sensitivity analyses were carried out. Heterogeneity among the instrumental variables (IVs) in the IVW method was quantified using Cochran’s Q statistic and MR-Egger regression (p < 0.05 was regarded as potential heterogeneity in IVs) [39]. The MR-Egger intercept test was applied to detect the presence of horizontal pleiotropy. If the p-value is greater than 0.05, it suggests the absence of horizontal pleiotropy [40]. Additionally, a powerful method known as MR pleiotropy residual sum and outlier (MR-PRESSO) is used to detect and remove potential horizontal pleiotropy outliers, which could greatly impact the estimation results within the MR-PRESSO framework. For each SNP, the MR-PRESSO outlier test calculates a p-value for its horizontal pleiotropy significance, while the MR-PRESSO global test calculates a p-value for the overall level of horizontal pleiotropy. Based on the p-values from the MR-PRESSO outlier test, SNPs are sorted in ascending order and sequentially removed from the list. Each time a SNP is removed from the list, the remaining SNPs are subjected to the MR-PRESSO global test. This recursive process is repeated until the p-value from the global test becomes insignificant (P > 0.05). The remaining SNPs after removing the ones with horizontal pleiotropy are then used for subsequent MR analysis [41]. We employ the “leave-one-out” method to assess sensitivity. This involves systematically removing each instrumental SNP one at a time and then re-performing the IVW analysis to determine if the causal estimates might be biased or influenced by a single SNP [42]. The statistical analysis was conducted using R version 4.3.1, with the “TwoSampleMR” R software package, which is widely used for this type of analysis.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Results
Causal effect of gut microbiota on PAD
This study selected 211 relative abundances of intestinal microbiota from gut microbiota GWAS data involving 18,340 participants as the exposure variable for the research. Due to the negative results of heterogeneity and pleiotropy tests, the IVW analysis results were used as the primary reference indicator in the study. The MR analysis results indicated that there may have been associations between nine different intestinal microbiota (1 class, 1 order, 2 families, and 5 genera) and PAD (Figure 2). Additionally, the results of the other four computational methods were found in Table 1. Lastly, the results of the five MR models analyzing the association between all gut microbiota and PAD risk were found in Supplementary Table 1.
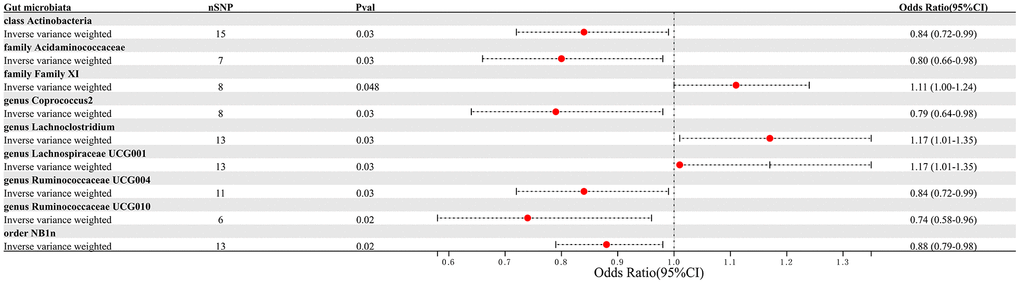
Figure 2. Forest plot summarizing the causal impact of gut microbiota composition on the risk of PAD, based on the IVW method.
Table 1. Statistical results of the four models other than IVW.
| Gut microbiota | Analytical methods | nSNP | P-value | Odds ratio (95%CI) |
| class Actinobacteria | ||||
| MR Egger | 15 | 0.45 | 1.20 (0.76-1.90) | |
| Weighted median | 15 | 0.35 | 0.90 (0.72-1.12) | |
| Simple mode | 15 | 0.22 | 0.76 (0.50-1.16) | |
| Weighted mode | 15 | 0.93 | 0.99 (0.74-1.31) | |
| family Acidaminococcaceae | ||||
| MR Egger | 7 | 0.33 | 0.72 (0.39-1.31) | |
| Weighted median | 7 | 0.05 | 0.77 (0.60-1.00) | |
| Simple mode | 7 | 0.19 | 0.74 (0.50-1.10) | |
| Weighted mode | 7 | 0.14 | 0.73 (0.52-1.04) | |
| family Family XI | ||||
| MR Egger | 8 | 0.21 | 1.63 (0.83-3.21) | |
| Weighted median | 8 | 0.07 | 1.14 (0.99-1.30) | |
| Simple mode | 8 | 0.19 | 1.20 (0.94-1.53) | |
| Weighted mode | 8 | 0.21 | 1.19 (0.93-1.53) | |
| genus Coprococcus2 | ||||
| MR Egger | 8 | 0.48 | 1.89 (0.36-10.01) | |
| Weighted median | 8 | 0.1 | 0.79 (0.60-1.04) | |
| Simple mode | 8 | 0.11 | 0.64 (0.39-1.04) | |
| Weighted mode | 8 | 0.1 | 0.65 (0.41-1.02) | |
| genus Lachnoclostridium | ||||
| MR Egger | 13 | 0.43 | 1.30 (0.70-2.42) | |
| Weighted median | 13 | 0.08 | 1.19 (0.98-1.44) | |
| Simple mode | 13 | 0.24 | 1.22 (0.89-1.68) | |
| Weighted mode | 13 | 0.31 | 1.19 (0.86-1.63) | |
| genus Lachnospiraceae UCG001 | ||||
| MR Egger | 13 | 0.43 | 1.30 (0.70-2.42) | |
| Weighted median | 13 | 0.08 | 1.19 (0.98-1.44) | |
| Simple mode | 13 | 0.24 | 1.22 (0.89-1.68) | |
| Weighted mode | 13 | 0.31 | 1.19 (0.86-1.63) | |
| genus Ruminococcaceae UCG004 | ||||
| MR Egger | 11 | 0.52 | 1.35 (0.56-3.27) | |
| Weighted median | 11 | 0.3 | 0.89 (0.71-1.11) | |
| Simple mode | 11 | 0.67 | 0.92 (0.63-1.35) | |
| Weighted mode | 11 | 0.72 | 0.93 (0.63-1.37) | |
| genus Ruminococcaceae UCG010 | ||||
| MR Egger | 6 | 0.32 | 0.64 (0.30-1.37) | |
| Weighted median | 6 | 0.02 | 0.68 (0.50-0.93) | |
| Simple mode | 6 | 0.09 | 0.64 (0.42-0.97) | |
| Weighted mode | 6 | 0.09 | 0.66 (0.45-0.98) | |
| order NB1n | ||||
| MR Egger | 13 | 0.56 | 0.87 (0.56-1.36) | |
| Weighted median | 13 | 0.02 | 0.85 (0.74-0.98) | |
| Simple mode | 13 | 0.2 | 0.86 (0.69-1.07) | |
| Weighted mode | 13 | 0.2 | 0.86 (0.69-1.07) |
We identified 3 microbiota taxonomic groups that were positively correlated with PAD. These included the family Family XI (OR=1.11, CI 1.00-1.24, P=0.048), the genus Lachnoclostridium (OR=1.24, CI 1.02-1.50, P=0.033), and the genus Lachnospiraceae UCG001 (OR=1.17, CI 1.01-1.35, P=0.031). Additionally, we found 6 microbiota taxonomic groups that were negatively correlated with PAD. These included the class Actinobacteria (OR=0.84, CI 0.72-0.99, P=0.034), the family Acidaminococcaceae (OR=0.80, CI 0.66-0.98, P=0.029), the genus Coprococcus2 (OR=0.79, CI 0.64-0.98, P=0.029), the genus Ruminococcaceae UCG004 (OR=0.84, CI 0.72-0.99, P=0.032), the genus Ruminococcaceae UCG010 (OR=0.74, CI 0.58-0.96, P=0.022), and the order NB1n (OR=0.88, CI 0.79-0.98, P=0.02). We presented these statistical results using scatter plots (Figure 3) and forest plots (Figure 4) to illustrate the causal effects of each SNP of the gut microbiota on PAD risk.
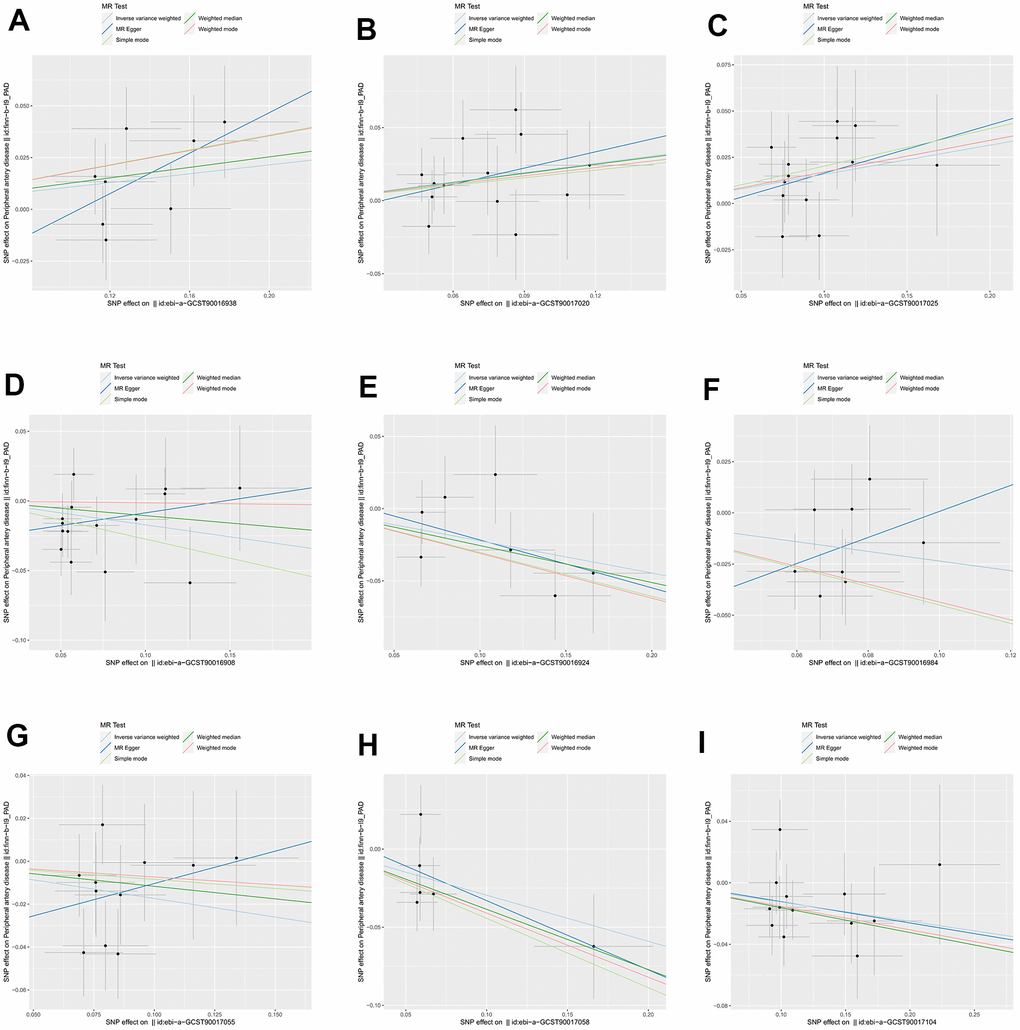
Figure 3. The scatter plots for association between gut microbiota and PAD. (A) family Family XI; (B) genus Lachnoclostridium; (C) genus Lachnospiraceae; (D) class Actinobacteria; (E) family Acidaminococcaceae; (F) genus Coprococcus2; (G) genus Ruminococcaceae UCG004; (H) genus Ruminococcaceae UCG010; (I) order NB1n. Note: SNP effects were plotted into lines for the inverse-variance weighted test (light blue line), MR-Egger (blue line), weighted median (green line), Simple mode (light green line) and Weighted mode (red line). The slope of the line corresponded to the causal estimation.
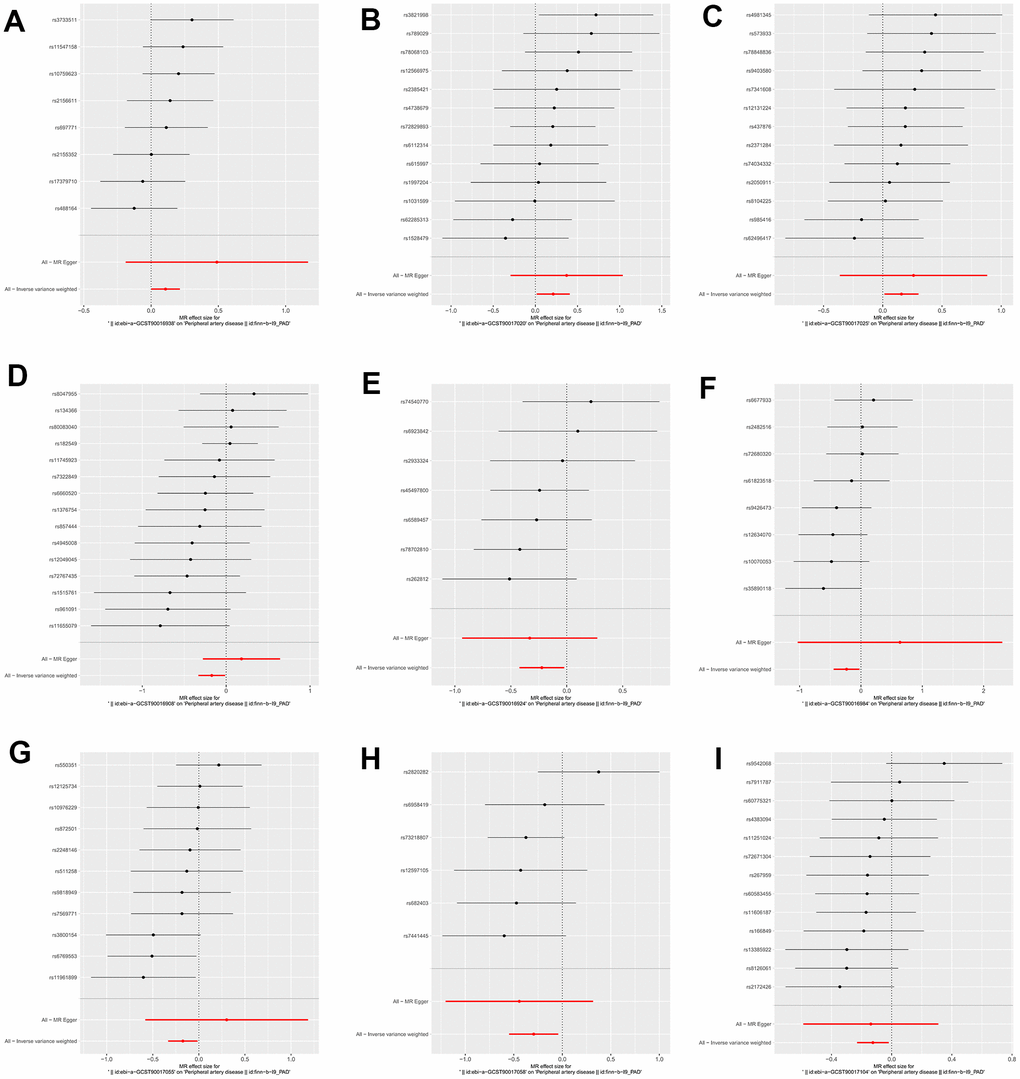
Figure 4. The forest plots for the association between gut microbiota and PAD. (A) family Family XI; (B) genus Lachnoclostridium; (C) genus Lachnospiraceae; (D) class Actinobacteria; (E) family Acidaminococcaceae; (F) genus Coprococcus2; (G) genus Ruminococcaceae UCG004; (H) genus Ruminococcaceae UCG010; (I) order NB1n.
To assess heterogeneity in the results, a Cochran’s Q test with a distribution set at 10,000 for both the IVW and MR-Egger regression analyses was conducted. The test results indicated the absence of heterogeneity in the SNPs for each microbiota taxonomic group. Additionally, the symmetry of the funnel plot suggested that there was no heterogeneity in the study (Supplementary Figure 1). The P-values of the intercept term in the MR-Egger regression consistently exceeded 0.05, signifying the absence of horizontal pleiotropy. In addition, in the MR-PRESSO analysis, if the global test p-value for the associated microbiota community is greater than 0.05, it indicates the absence of horizontal pleiotropy (Table 2). The leave-one-out test results confirmed the robustness of the MR findings in the study (Supplementary Figure 2).
Table 2. Heterogeneity and level of pleiotropy testing for 9 specific microbiota communities.
| Gut microbiota | nsnp | Q_inverse.variance.weighted | Q_pval_inverse.variance.weighted | Q_MR.egger | Q_pval_MR.egger | Egger_intercept | Egger_intercept_pval | MR-PRESSO_global_test_p |
| family Family XI | 8 | 6.51 | 0.48 | 5.26 | 0.51 | -0.05 | 0.31 | 0.523 |
| genus Lachnoclostridium | 13 | 9.01 | 0.7 | 8.77 | 0.64 | -0.01 | 0.63 | 0.735 |
| genus Lachnospiraceae UCG001 | 13 | 7.15 | 0.85 | 7.04 | 0.8 | -0.01 | 0.75 | 0.825 |
| class Actinobacteria | 15 | 12.48 | 0.57 | 9.89 | 0.7 | -0.03 | 0.13 | 0.676 |
| family Acidaminococcaceae | 7 | 4.9 | 0.56 | 4.76 | 0.45 | 0.01 | 0.72 | 0.588 |
| genus Coprococcus2 | 8 | 6.36 | 0.5 | 5.3 | 0.51 | -0.06 | 0.34 | 0.53 |
| genus Ruminococcaceae UCG004 | 11 | 9.58 | 0.48 | 8.43 | 0.49 | -0.04 | 0.31 | 0.49 |
| genus Ruminococcaceae UCG010 | 6 | 6.04 | 0.3 | 5.8 | 0.21 | 0.01 | 0.7 | 0.376 |
| order NB1n | 13 | 10.31 | 0.59 | 10.31 | 0.5 | 0 | 0.95 | 0.528 |
Reverse-direction MR analyses
We employed the IVW method to perform a reverse MR analysis, aiming to examine the causal association between 9 specific gut microbiota taxa and PAD. Three SNPs that were closely associated with PAD were identified. As shown in Table 3, none of the gut microbiota taxa categories, including class Actinobacteria (P=0.31), family Acidaminococcaceae (P=0.88), family Family XI (P=0.99), genus Coprococcus2 (P=0.93), genus Lachnoclostridium (P=0.14), genus Lachnospiraceae UCG001 (P=0.77), genus Ruminococcaceae UCG004 (P=0.66), genus Ruminococcaceae UCG010 (P=0.53), and order NB1n (P=0.72), showed a significant reverse causal relationship with PAD. The reliability of our findings was validated through the utilization of the MR-Egger regression method and the Cochran’s Q test.
Table 3. Reverse MR analysis results for 9 specific microbiota communities.
| Gut microbiota | OR (95%CI) | Pval | Q_pval_inverse.variance.weighted | Q_pval_MR.egger | Egger_intercept (pval) |
| class Actinobacteria | 0.95 (0.87-1.05) | 0.31 | 0.6 | 0.78 | 0.09 (0.51) |
| family Acidaminococcaceae | 0.99 (0.89-1.10) | 0.88 | 0.99 | 0.89 | -0.01 (0.96) |
| family Family XI | 1.00 (0.79-1.28) | 0.99 | 0.27 | 0.46 | -0.3 (0.38) |
| genus Coprococcus2 | 1.00 (0.89-1.11) | 0.93 | 0.88 | 0.78 | -0.05 (0.74) |
| genus Lachnoclostridium | 1.08 (0.98-1.19) | 0.14 | 0.3 | 0.12 | -0.02 (0.92) |
| genus Lachnospiraceae UCG001 | 0.98 (0.85-1.13) | 0.77 | 0.22 | 0.52 | -0.18 (0.35) |
| genus Ruminococcaceae UCG004 | 0.95 (0.77-1.18) | 0.66 | 0.05 | 0.01 | 0.02 (0.96) |
| genus Ruminococcaceae UCG010 | 0.97 (0.87-1.07) | 0.53 | 0.97 | 0.82 | 0 (0.98) |
Discussion
Main findings and interpretation
For the first time, this study evaluated the possible causal connection between gut microbiota and PAD utilizing bidirectional MR. We identified specific gut microbiota communities strongly linked to PAD at various taxonomic levels, including phylum, order, family, and genus. Among them, family Family XI, genus Lachnoclostridium, and genus Lachnospiraceae UCG001 were identified as potential risk factors for PAD, while class Actinobacteria, family Acidaminococcaceae, genus Coprococcus2, genus Ruminococcaceae UCG004, genus Ruminococcaceae UCG010, and order NB1n were identified as potential protective factors for PAD. Sensitivity analysis indicated the absence of heterogeneity or horizontal pleiotropy, affirming that our MR analysis remains unaffected by confounding factors. Furthermore, the “leave-one-out” analysis validated the study’s robustness.
The gut microbiota, as the most complex microbiota ecosystem in the human body, regulates a wide range of physiological processes [43]. Simultaneously, progress in high-throughput DNA sequencing technology has furnished us with a more profound comprehension of the gut microbiota’s composition and functionalities. Generally, the gut microbiota community comprises a diverse assortment of bacteria present in specific proportions. These bacteria interact, constrain, and depend on each other, establishing a qualitative and quantitative ecological balance [44]. Gut dysbiosis increases intestinal permeability by inhibiting tight junction proteins, allowing translocation of lipopolysaccharides into the circulation [45]. The lipopolysaccharides derived from gut dysbiosis bind to Toll-like receptors (TLRs) and activate downstream immune responses [46]. Lipopolysaccharides bind to the TLR4 complex and its co-receptor, cluster of differentiation 14 (CD14). Upregulation of TLRs initiates the inflammation-driven process of atherosclerosis [47]. The interaction between lipopolysaccharides and TLR4 activates the MYD88 and NFκB pathways, thereby enhancing the synthesis of pro-inflammatory cytokines such as IL-6, IL-1, IL-27, and TNF-α. These inflammatory cytokines play a role in the development of atherosclerosis and cardiovascular diseases [48].
The primary characteristic of PAD is AS. A growing body of evidence underscores the pivotal role of the gut microbiota in the initiation and advancement of AS, rendering it a novel target for the prevention and management of AS [49]. During the early 21st century, numerous investigations documented the presence of bacterial DNA from diverse species within atherosclerotic plaques, representing the initial evidence of a connection between the microbiota and AS [50, 51]. The gut microbiota can serve as a circulating factor that promotes or prevents the formation of AS by generating metabolites from dietary sources [52, 53]. Gut dysbiosis, which signifies the perturbation of gut microbiota linked to various diseases, was also highlighted. Another study revealed that individuals with AS display an ecological imbalance characterized by an overabundance of Actinobacteria in their gut compared to a healthy control group, a phenomenon associated with butyrate production [54]. Furthermore, changes in the composition of gut microbiota and the presence of its byproducts, including short-chain fatty acids (SCFAs) and bile acids (BAs), are linked to the risk of AS development [55, 56].
Particular bacterial species within the gut are correlated with the progression of PAD. At the phylum level, significant alterations in the gut microbiota composition are observed in PAD patients compared to the control group [57]. In this study, we have identified and confirmed 9 specific gut microbiota communities that have a potential causal relationship with PAD. Dysbiosis has the potential to compromise the integrity of the intestinal barrier, resulting in the release of detrimental metabolites like lipopolysaccharides (LPS) and other bacterial constituents such as peptidoglycans into the bloodstream. This, in turn, can incite an inflammatory reaction and contribute to the progression of AS [58, 59]. A study has established a positive correlation between the presence of Lachnoclostridium and the production of inflammatory cytokines and LPS in the serum [60]. Trimethylamine (TMA) is a small-molecule byproduct resulting from the microbiota metabolism of choline, carnitine, and phosphatidylcholine within the gut [61, 62]. Within the liver, TMA undergoes conversion into trimethylamine-N-oxide (TMAO) through the action of flavin-containing monooxygenase 3 [63, 64]. A growing body of evidence supports the notion that plasma TMAO serves as an independent risk factor for AS. Research has shown that Lachnoclostridium strains can effectively convert choline into TMA. When co-administered with choline to ApoE-/- mice, these strains can increase the levels of TMAO in the serum and promote the formation of AS [65]. Ruminococcaceae generates acetate and butyrate, serving as the primary energy source for intestinal epithelial cells while concurrently suppressing the signaling pathways of pro-inflammatory cytokines [66]. The anti-atherosclerotic effects of butyrate have been extensively confirmed [67, 68]. Butyrate can reduce cholesterol levels in the blood by converting cholesterol to coprostanol [69]. In addition, Coprococcus can also produce butyrate [70]. In AS disease, the significant reduction of Acidaminocccaceae, decreased synthesis of SCFAs, and increased TMAO levels can lead to an increase in atherogenic lipoproteins in the blood, contributing to the development of AS [71].
Limitation
However, this study also has limitations: (1) Human behavior is intricate, and while comprehending the genetic predisposition to a disease can aid in its prevention to a certain degree [72], environmental factors also contribute to the disease’s onset [73]. MR can only partially address confounding factors, including environmental influences. (2) The study’s outcome data are based on European and American populations, and further validation is required to ascertain whether the results are representative of the broader population. (3) While confirming a causal link between gut microbiota and PAD, the precise underlying mechanisms remain elusive, necessitating additional research.
Conclusions
The study obtained data from GWAS databases and employed a two-sample bidirectional MR approach to validate the potential causal connection between gut microbiota and PAD. These findings offer novel insights into the pathogenesis of PAD and potential treatment strategies. Subsequent studies should aim to delve deeper into the mechanisms through which these microbiota communities impact PAD and investigate potential therapeutic approaches targeting the gut microbiota. Additionally, gut microbiota may serve as a novel important predictive biomarker for PAD.
Author Contributions
LGB, YX, and FWJ proposed the research design. SHS and YX conducted literature search, publication screening, and data extraction. SHS carried out the data analysis. WFF and LXY conducted manuscript revisions. The authors collectively contributed to and endorsed the final version of the article.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
The study was funded by the National Natural Science Foundation of China (82274528); Construction Task Book for the Three-Year Action Plan for Accelerating the Inheritance and Innovative Development of Traditional Chinese Medicine in Shanghai (2021-2023) (ZY(2021-2023)-0211); Shanghai Municipal Health Commission Scientific Research Programme Mission Statement (202240228); Special Youth Project for Clinical Research of Shanghai Municipal Health Commission (20234Y0162); Clinical Research Talent Training Program of Shanghai University of Traditional Chinese Medicine Affiliated Hospital (2023LCRC06); Four Bright Foundations of Shuguang Hospital Affiliated to Shanghai University of Traditional Chinese Medicine (SGKJ-202301); Shanghai Pudong New Area Health and Family Planning Commission’s Pudong Famous TCM Cultivation Program project (PWRZM2020-18).
References
- 1. Cook IO, Chung J. Contemporary Medical Management of Peripheral Arterial Disease. Cardiovasc Drugs Ther. 2023. [Epub ahead of print]. https://doi.org/10.1007/s10557-023-07516-2 [PubMed]
- 2. Fowkes FG, Rudan D, Rudan I, Aboyans V, Denenberg JO, McDermott MM, Norman PE, Sampson UK, Williams LJ, Mensah GA, Criqui MH. Comparison of global estimates of prevalence and risk factors for peripheral artery disease in 2000 and 2010: a systematic review and analysis. Lancet. 2013; 382:1329–40. https://doi.org/10.1016/S0140-6736(13)61249-0 [PubMed]
- 3. Hiatt WR, Armstrong EJ, Larson CJ, Brass EP. Pathogenesis of the limb manifestations and exercise limitations in peripheral artery disease. Circ Res. 2015; 116:1527–39. https://doi.org/10.1161/CIRCRESAHA.116.303566 [PubMed]
- 4. Hamburg NM, Balady GJ. Exercise rehabilitation in peripheral artery disease: functional impact and mechanisms of benefits. Circulation. 2011; 123:87–97. https://doi.org/10.1161/CIRCULATIONAHA.109.881888 [PubMed]
- 5. Rando MM, Biscetti F, Masciocchi C, Savino M, Nicolazzi MA, Nardella E, Cecchini AL, Rossini E, Massetti M, Gasbarrini A, Flex A. Impact of COVID-19 pandemic on patients affected by peripheral arterial disease: an Italian single-center study. Eur Rev Med Pharmacol Sci. 2023; 27:10144–55. https://doi.org/10.26355/eurrev_202310_34194 [PubMed]
- 6. Song P, Rudan D, Zhu Y, Fowkes FJ, Rahimi K, Fowkes FG, Rudan I. Global, regional, and national prevalence and risk factors for peripheral artery disease in 2015: an updated systematic review and analysis. Lancet Glob Health. 2019; 7:e1020–30. https://doi.org/10.1016/S2214-109X(19)30255-4 [PubMed]
- 7. Nosova EV, Conte MS, Grenon SM. Advancing beyond the “heart-healthy diet” for peripheral arterial disease. J Vasc Surg. 2015; 61:265–74. https://doi.org/10.1016/j.jvs.2014.10.022 [PubMed]
- 8. Qamar SUR, Spahić L, Benolić L, Zivanovic M, Filipović N. Treatment of Peripheral Artery Disease Using Injectable Biomaterials and Drug-Coated Balloons: Safety and Efficacy Perspective. Pharmaceutics. 2023; 15:1813. https://doi.org/10.3390/pharmaceutics15071813 [PubMed]
- 9. Krga I, Glibetic M. 2.16—Gut microbiota in health and diseases. In: Glibetic M, editor. Comprehensive gut microbiota. Oxford: Elsevier; 2022; 182–98.
- 10. Adak A, Khan MR. An insight into gut microbiota and its functionalities. Cell Mol Life Sci. 2019; 76:473–93. https://doi.org/10.1007/s00018-018-2943-4 [PubMed]
- 11. Busnelli M, Manzini S, Chiesa G. The Gut Microbiota Affects Host Pathophysiology as an Endocrine Organ: A Focus on Cardiovascular Disease. Nutrients. 2019; 12:79. https://doi.org/10.3390/nu12010079 [PubMed]
- 12. Huang JJ, Guo X. [Research update on the association between gut microbiota and its metabolites with atherosclerosis]. Zhonghua Xin Xue Guan Bing Za Zhi. 2023; 51:548–54. https://doi.org/10.3760/cma.j.cn112148-20221227-01016 [PubMed]
- 13. Belli M, Barone L, Longo S, Prandi FR, Lecis D, Mollace R, Margonato D, Muscoli S, Sergi D, Federici M, Barillà F. Gut Microbiota Composition and Cardiovascular Disease: A Potential New Therapeutic Target? Int J Mol Sci. 2023; 24:11971. https://doi.org/10.3390/ijms241511971 [PubMed]
- 14. Lanter BB, Sauer K, Davies DG. Bacteria present in carotid arterial plaques are found as biofilm deposits which may contribute to enhanced risk of plaque rupture. mBio. 2014; 5:e01206–14. https://doi.org/10.1128/mBio.01206-14 [PubMed]
- 15. Choroszy M, Litwinowicz K, Bednarz R, Roleder T, Lerman A, Toya T, Kamiński K, Sawicka-Śmiarowska E, Niemira M, Sobieszczańska B. Human Gut Microbiota in Coronary Artery Disease: A Systematic Review and Meta-Analysis. Metabolites. 2022; 12:1165. https://doi.org/10.3390/metabo12121165 [PubMed]
- 16. Liu H, Chen X, Hu X, Niu H, Tian R, Wang H, Pang H, Jiang L, Qiu B, Chen X, Zhang Y, Ma Y, Tang S, et al. Alterations in the gut microbiome and metabolism with coronary artery disease severity. Microbiome. 2019; 7:68. https://doi.org/10.1186/s40168-019-0683-9 [PubMed]
- 17. Cheng X, Han X, Zhou L, Sun Y, Zhou Q, Lin X, Gao Z, Wang J, Zhao W. Cabernet sauvignon dry red wine ameliorates atherosclerosis in mice by regulating inflammation and endothelial function, activating AMPK phosphorylation, and modulating gut microbiota. Food Res Int. 2023; 169:112942. https://doi.org/10.1016/j.foodres.2023.112942 [PubMed]
- 18. Chen H, Ye B, Su W, Song Y, Sun PL, Zhou X, Zhang G. The causal role of gut microbiota in susceptibility and severity of COVID-19: A bidirectional Mendelian randomization study. J Med Virol. 2023; 95:e28880. https://doi.org/10.1002/jmv.28880 [PubMed]
- 19. Bowden J, Holmes MV. Meta-analysis and Mendelian randomization: A review. Res Synth Methods. 2019; 10:486–96. https://doi.org/10.1002/jrsm.1346 [PubMed]
- 20. Birney E. Mendelian Randomization. Cold Spring Harb Perspect Med. 2022; 12:a041302. https://doi.org/10.1101/cshperspect.a041302 [PubMed]
- 21. Didelez V, Sheehan N. Mendelian randomization as an instrumental variable approach to causal inference. Stat Methods Med Res. 2007; 16:309–30. https://doi.org/10.1177/0962280206077743 [PubMed]
- 22. Lawlor DA, Harbord RM, Sterne JA, Timpson N, Davey Smith G. Mendelian randomization: using genes as instruments for making causal inferences in epidemiology. Stat Med. 2008; 27:1133–63. https://doi.org/10.1002/sim.3034 [PubMed]
- 23. Skrivankova VW, Richmond RC, Woolf BAR, Yarmolinsky J, Davies NM, Swanson SA, VanderWeele TJ, Higgins JP, Timpson NJ, Dimou N, Langenberg C, Golub RM, Loder EW, et al. Strengthening the Reporting of Observational Studies in Epidemiology Using Mendelian Randomization: The STROBE-MR Statement. JAMA. 2021; 326:1614–21. https://doi.org/10.1001/jama.2021.18236 [PubMed]
- 24. Davey Smith G, Hemani G. Mendelian randomization: genetic anchors for causal inference in epidemiological studies. Hum Mol Genet. 2014; 23:R89–98. https://doi.org/10.1093/hmg/ddu328 [PubMed]
- 25. Kurilshikov A, Medina-Gomez C, Bacigalupe R, Radjabzadeh D, Wang J, Demirkan A, Le Roy CI, Raygoza Garay JA, Finnicum CT, Liu X, Zhernakova DV, Bonder MJ, Hansen TH, et al. Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat Genet. 2021; 53:156–65. https://doi.org/10.1038/s41588-020-00763-1 [PubMed]
- 26. FinnGen_Consortium. FinnGen data. https://www.finngen.fi/.
- 27. Yu XH, Yang YQ, Cao RR, Bo L, Lei SF. The causal role of gut microbiota in development of osteoarthritis. Osteoarthritis Cartilage. 2021; 29:1741–50. https://doi.org/10.1016/j.joca.2021.08.003 [PubMed]
- 28. Xiang K, Wang P, Xu Z, Hu YQ, He YS, Chen Y, Feng YT, Yin KJ, Huang JX, Wang J, Wu ZD, Yang XK, Wang DG, et al. Causal Effects of Gut Microbiome on Systemic Lupus Erythematosus: A Two-Sample Mendelian Randomization Study. Front Immunol. 2021; 12:667097. https://doi.org/10.3389/fimmu.2021.667097 [PubMed]
- 29. Cao J, Wang N, Luo Y, Ma C, Chen Z, Chenzhao C, Zhang F, Qi X, Xiong W. A cause-effect relationship between Graves’ disease and the gut microbiome contributes to the thyroid-gut axis: A bidirectional two-sample Mendelian randomization study. Front Immunol. 2023; 14:977587. https://doi.org/10.3389/fimmu.2023.977587 [PubMed]
- 30. Hemani G, Zheng J, Elsworth B, Wade KH, Haberland V, Baird D, Laurin C, Burgess S, Bowden J, Langdon R, Tan VY, Yarmolinsky J, Shihab HA, et al. The MR-Base platform supports systematic causal inference across the human phenome. Elife. 2018; 7:e34408. https://doi.org/10.7554/eLife.34408 [PubMed]
- 31. Zhuang Z, Yu C, Guo Y, Bian Z, Yang L, Millwood IY, Walters RG, Chen Y, Xu Q, Zou M, Chen J, Chen Z, Lv J, et al. Metabolic Signatures of Genetically Elevated Vitamin D Among Chinese: Observational and Mendelian Randomization Study. J Clin Endocrinol Metab. 2021; 106:e3249–60. https://doi.org/10.1210/clinem/dgab097 [PubMed]
- 32. Kamat MA, Blackshaw JA, Young R, Surendran P, Burgess S, Danesh J, Butterworth AS, Staley JR. PhenoScanner V2: an expanded tool for searching human genotype-phenotype associations. Bioinformatics. 2019; 35:4851–53. https://doi.org/10.1093/bioinformatics/btz469 [PubMed]
- 33. Bowden J, Del Greco M F, Minelli C, Davey Smith G, Sheehan N, Thompson J. A framework for the investigation of pleiotropy in two-sample summary data Mendelian randomization. Stat Med. 2017; 36:1783–802. https://doi.org/10.1002/sim.7221 [PubMed]
- 34. Bowden J, Davey Smith G, Burgess S. Mendelian randomization with invalid instruments: effect estimation and bias detection through Egger regression. Int J Epidemiol. 2015; 44:512–25. https://doi.org/10.1093/ije/dyv080 [PubMed]
- 35. Papadimitriou N, Dimou N, Tsilidis KK, Banbury B, Martin RM, Lewis SJ, Kazmi N, Robinson TM, Albanes D, Aleksandrova K, Berndt SI, Timothy Bishop D, Brenner H, et al. Physical activity and risks of breast and colorectal cancer: a Mendelian randomisation analysis. Nat Commun. 2020; 11:597. https://doi.org/10.1038/s41467-020-14389-8 [PubMed]
- 36. Zheng J, Baird D, Borges MC, Bowden J, Hemani G, Haycock P, Evans DM, Smith GD. Recent Developments in Mendelian Randomization Studies. Curr Epidemiol Rep. 2017; 4:330–45. https://doi.org/10.1007/s40471-017-0128-6 [PubMed]
- 37. Guo HY, Wang W, Peng H, Yuan H. Bidirectional two-sample Mendelian randomization study of causality between rheumatoid arthritis and myocardial infarction. Front Immunol. 2022; 13:1017444. https://doi.org/10.3389/fimmu.2022.1017444 [PubMed]
- 38. Dai Z, Xu W, Ding R, Peng X, Shen X, Song J, Du P, Wang Z, Liu Y. Two-sample Mendelian randomization analysis evaluates causal associations between inflammatory bowel disease and osteoporosis. Front Public Health. 2023; 11:1151837. https://doi.org/10.3389/fpubh.2023.1151837 [PubMed]
- 39. Bowden J, Spiller W, Del Greco M F, Sheehan N, Thompson J, Minelli C, Davey Smith G. Improving the visualization, interpretation and analysis of two-sample summary data Mendelian randomization via the Radial plot and Radial regression. Int J Epidemiol. 2018; 47:2100. https://doi.org/10.1093/ije/dyy265 [PubMed]
- 40. Carter AR, Sanderson E, Hammerton G, Richmond RC, Davey Smith G, Heron J, Taylor AE, Davies NM, Howe LD. Mendelian randomisation for mediation analysis: current methods and challenges for implementation. Eur J Epidemiol. 2021; 36:465–78. https://doi.org/10.1007/s10654-021-00757-1 [PubMed]
- 41. Verbanck M, Chen CY, Neale B, Do R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat Genet. 2018; 50:693–8. https://doi.org/10.1038/s41588-018-0099-7 [PubMed] Erratum in: Nat Genet. 2018; 50:1196 https://doi.org/10.1038/s41588-018-0164-2 [PubMed]
- 42. Gronau QF, Wagenmakers EJ. Limitations of Bayesian Leave-One-Out Cross-Validation for Model Selection. Comput Brain Behav. 2019; 2:1–11. https://doi.org/10.1007/s42113-018-0011-7 [PubMed]
- 43. Fan Y, Pedersen O. Gut microbiota in human metabolic health and disease. Nat Rev Microbiol. 2021; 19:55–71. https://doi.org/10.1038/s41579-020-0433-9 [PubMed]
- 44. Li T, Shao W, Wang Y, Zhou R, Yun Z, He Y, Wu Y. A two-sample mendelian randomization analysis investigates associations between gut microbiota and infertility. Sci Rep. 2023; 13:11426. https://doi.org/10.1038/s41598-023-38624-6 [PubMed]
- 45. Cani PD, Bibiloni R, Knauf C, Waget A, Neyrinck AM, Delzenne NM, Burcelin R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes. 2008; 57:1470–81. https://doi.org/10.2337/db07-1403 [PubMed]
- 46. Chacón MR, Lozano-Bartolomé J, Portero-Otín M, Rodríguez MM, Xifra G, Puig J, Blasco G, Ricart W, Chaves FJ, Fernández-Real JM. The gut mycobiome composition is linked to carotid atherosclerosis. Benef Microbes. 2018; 9:185–98. https://doi.org/10.3920/BM2017.0029 [PubMed]
- 47. Xu XH, Shah PK, Faure E, Equils O, Thomas L, Fishbein MC, Luthringer D, Xu XP, Rajavashisth TB, Yano J, Kaul S, Arditi M. Toll-like receptor-4 is expressed by macrophages in murine and human lipid-rich atherosclerotic plaques and upregulated by oxidized LDL. Circulation. 2001; 104:3103–8. https://doi.org/10.1161/hc5001.100631 [PubMed]
- 48. Guzzo C, Ayer A, Basta S, Banfield BW, Gee K. IL-27 enhances LPS-induced proinflammatory cytokine production via upregulation of TLR4 expression and signaling in human monocytes. J Immunol. 2012; 188:864–73. https://doi.org/10.4049/jimmunol.1101912 [PubMed]
- 49. Al Samarraie A, Pichette M, Rousseau G. Role of the Gut Microbiome in the Development of Atherosclerotic Cardiovascular Disease. Int J Mol Sci. 2023; 24:5420. https://doi.org/10.3390/ijms24065420 [PubMed]
- 50. Haraszthy VI, Zambon JJ, Trevisan M, Zeid M, Genco RJ. Identification of periodontal pathogens in atheromatous plaques. J Periodontol. 2000; 71:1554–60. https://doi.org/10.1902/jop.2000.71.10.1554 [PubMed]
- 51. Ott SJ, El Mokhtari NE, Musfeldt M, Hellmig S, Freitag S, Rehman A, Kühbacher T, Nikolaus S, Namsolleck P, Blaut M, Hampe J, Sahly H, Reinecke A, et al. Detection of diverse bacterial signatures in atherosclerotic lesions of patients with coronary heart disease. Circulation. 2006; 113:929–37. https://doi.org/10.1161/CIRCULATIONAHA.105.579979 [PubMed]
- 52. Brown JM, Hazen SL. The gut microbial endocrine organ: bacterially derived signals driving cardiometabolic diseases. Annu Rev Med. 2015; 66:343–59. https://doi.org/10.1146/annurev-med-060513-093205 [PubMed]
- 53. Gregory JC, Buffa JA, Org E, Wang Z, Levison BS, Zhu W, Wagner MA, Bennett BJ, Li L, DiDonato JA, Lusis AJ, Hazen SL. Transmission of atherosclerosis susceptibility with gut microbial transplantation. J Biol Chem. 2015; 290:5647–60. https://doi.org/10.1074/jbc.M114.618249 [PubMed]
- 54. Karlsson FH, Fåk F, Nookaew I, Tremaroli V, Fagerberg B, Petranovic D, Bäckhed F, Nielsen J. Symptomatic atherosclerosis is associated with an altered gut metagenome. Nat Commun. 2012; 3:1245. https://doi.org/10.1038/ncomms2266 [PubMed]
- 55. El-Far M, Durand M, Turcotte I, Larouche-Anctil E, Sylla M, Zaidan S, Chartrand-Lefebvre C, Bunet R, Ramani H, Sadouni M, Boldeanu I, Chamberland A, Lesage S, et al. Upregulated IL-32 Expression And Reduced Gut Short Chain Fatty Acid Caproic Acid in People Living With HIV With Subclinical Atherosclerosis. Front Immunol. 2021; 12:664371. https://doi.org/10.3389/fimmu.2021.664371 [PubMed]
- 56. Bai Z, Liu Y, Zhao Y, Yan R, Yang L, Ma H, Wang J, Wang T, Li Y, Zhang G, Zhang X, Jia S, Wang H. Aspirin ameliorates atherosclerotic immuno-inflammation through regulating the Treg/Th17 axis and CD39-CD73 adenosine signaling via remodeling the gut microbiota in ApoE-/- mice. Int Immunopharmacol. 2023; 120:110296. https://doi.org/10.1016/j.intimp.2023.110296 [PubMed]
- 57. Zhang HJ, Wang Y, Jiang JD. Drug-gut Microbiome Interaction in Atherosclerosis Therapeutics. Curr Drug Metab. 2023; 24:482–92. https://doi.org/10.2174/1389200224666230410094806 [PubMed]
- 58. Lau K, Srivatsav V, Rizwan A, Nashed A, Liu R, Shen R, Akhtar M. Bridging the Gap between Gut Microbial Dysbiosis and Cardiovascular Diseases. Nutrients. 2017; 9:859. https://doi.org/10.3390/nu9080859 [PubMed]
- 59. Feng Q, Chen WD, Wang YD. Gut Microbiota: An Integral Moderator in Health and Disease. Front Microbiol. 2018; 9:151. https://doi.org/10.3389/fmicb.2018.00151 [PubMed]
- 60. Nastasi C, Candela M, Bonefeld CM, Geisler C, Hansen M, Krejsgaard T, Biagi E, Andersen MH, Brigidi P, Ødum N, Litman T, Woetmann A. The effect of short-chain fatty acids on human monocyte-derived dendritic cells. Sci Rep. 2015; 5:16148. https://doi.org/10.1038/srep16148 [PubMed]
- 61. Zhu Y, Jameson E, Crosatti M, Schäfer H, Rajakumar K, Bugg TD, Chen Y. Carnitine metabolism to trimethylamine by an unusual Rieske-type oxygenase from human microbiota. Proc Natl Acad Sci USA. 2014; 111:4268–73. https://doi.org/10.1073/pnas.1316569111 [PubMed]
- 62. Koeth RA, Levison BS, Culley MK, Buffa JA, Wang Z, Gregory JC, Org E, Wu Y, Li L, Smith JD, Tang WH, DiDonato JA, Lusis AJ, Hazen SL. γ-Butyrobetaine is a proatherogenic intermediate in gut microbial metabolism of L-carnitine to TMAO. Cell Metab. 2014; 20:799–812. https://doi.org/10.1016/j.cmet.2014.10.006 [PubMed]
- 63. Wang Z, Klipfell E, Bennett BJ, Koeth R, Levison BS, Dugar B, Feldstein AE, Britt EB, Fu X, Chung YM, Wu Y, Schauer P, Smith JD, et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature. 2011; 472:57–63. https://doi.org/10.1038/nature09922 [PubMed]
- 64. Serino M, Blasco-Baque V, Nicolas S, Burcelin R. Far from the eyes, close to the heart: dysbiosis of gut microbiota and cardiovascular consequences. Curr Cardiol Rep. 2014; 16:540. https://doi.org/10.1007/s11886-014-0540-1 [PubMed]
- 65. Cai YY, Huang FQ, Lao X, Lu Y, Gao X, Alolga RN, Yin K, Zhou X, Wang Y, Liu B, Shang J, Qi LW, Li J. Integrated metagenomics identifies a crucial role for trimethylamine-producing Lachnoclostridium in promoting atherosclerosis. NPJ Biofilms Microbiomes. 2022; 8:11. https://doi.org/10.1038/s41522-022-00273-4 [PubMed]
- 66. Morgan XC, Tickle TL, Sokol H, Gevers D, Devaney KL, Ward DV, Reyes JA, Shah SA, LeLeiko N, Snapper SB, Bousvaros A, Korzenik J, Sands BE, et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012; 13:R79. https://doi.org/10.1186/gb-2012-13-9-r79 [PubMed]
- 67. Bultman SJ. Bacterial butyrate prevents atherosclerosis. Nat Microbiol. 2018; 3:1332–3. https://doi.org/10.1038/s41564-018-0299-z [PubMed] Erratum in: Nat Microbiol. 2019; 4:375. https://doi.org/10.1038/s41564-018-0348-7 [PubMed]
- 68. Tayyeb JZ, Popeijus HE, Mensink RP, Konings MCJM, Mulders KHR, Plat J. The effects of short-chain fatty acids on the transcription and secretion of apolipoprotein A-I in human hepatocytes in vitro. J Cell Biochem. 2019; 120:17219–27. https://doi.org/10.1002/jcb.28982 [PubMed]
- 69. Vacca M, Celano G, Calabrese FM, Portincasa P, Gobbetti M, De Angelis M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms. 2020; 8:573. https://doi.org/10.3390/microorganisms8040573 [PubMed]
- 70. Altemani F, Barrett HL, Gomez-Arango L, Josh P, David McIntyre H, Callaway LK, Morrison M, Tyson GW, Dekker Nitert M. Pregnant women who develop preeclampsia have lower abundance of the butyrate-producer Coprococcus in their gut microbiota. Pregnancy Hypertens. 2021; 23:211–9. https://doi.org/10.1016/j.preghy.2021.01.002 [PubMed]
- 71. Drapkina OM, Ashniev GA, Zlobovskaya OA, Yafarova AA, Dementeva EV, Kaburova AN, Meshkov IO, Sheptulina AF, Kiselev AR, Kontsevaya AV, Zhamalov LM, Koretskiy SN, Pokrovskaya MS, et al. Diversities in the Gut Microbial Patterns in Patients with Atherosclerotic Cardiovascular Diseases and Certain Heart Failure Phenotypes. Biomedicines. 2022; 10:2762. https://doi.org/10.3390/biomedicines10112762 [PubMed]
- 72. Agustí A, Melén E, DeMeo DL, Breyer-Kohansal R, Faner R. Pathogenesis of chronic obstructive pulmonary disease: understanding the contributions of gene-environment interactions across the lifespan. Lancet Respir Med. 2022; 10:512–24. https://doi.org/10.1016/S2213-2600(21)00555-5 [PubMed]
- 73. Meisel SF, Beeken RJ, van Jaarsveld CH, Wardle J. Genetic susceptibility testing and readiness to control weight: Results from a randomized controlled trial. Obesity (Silver Spring). 2015; 23:305–12. https://doi.org/10.1002/oby.20958 [PubMed]