



Introduction
Squamous cell carcinoma of the oral cavity (OSCC) is a frequently encountered malignancy, notable for its high rate of recurrence. Despite recent progress in diagnosis and treatment, the 5-year overall survival (OS) has only marginally improved to 50–60%. However, the prognosis remains dismal for patients with advanced stage disease, with a survival rate of only 30–40% [1–3]. In locally advanced OSCC, the main cause of death is local and distant tumor recurrence. Therefore, it is crucial to diagnose recurrence early in advanced OSCC. Identifying molecular markers that predict disease prognosis and recurrence would be beneficial in this context.
Receptor tyrosine protein kinase KDR, also known as VEGFR2, is a key player in cell proliferation. It belongs to the EGFR family of receptor tyrosine kinases, which play a crucial role in regulating cell growth and division. KDR is often overexpressed in cancer cells, leading to uncontrolled cell growth and tumor development. Therefore, understanding the mechanisms of KDR regulation is crucial for developing effective cancer treatments [4]. KDR amplification has been detected in several malignant tumors [5–8]. In addition, the genetic analysis of OSCC and lymph node metastasis samples has revealed that the KDR gains, due to both amplifications and polysomes, were prevalent in the OSCC samples [9, 10]. There is also evidence that the heterodimerization of KDR causes the autophosphorylation of tyrosine residues within the cytoplasmic domain of the heterodimer, which subsequently initiates various signaling pathways that promote tumorigenesis [11]. Therefore, the treatment against KDR has been recognized as a reliable approach in multiple cancers, particularly breast cancer [11–13]. It has been observed that trastuzumab resistance occurs in patients who have lost KDR [14–16]. In addition, despite the identification of targetable KDR aberrations in OSCC, certain OSCC cell lines exhibited low levels of KDR expression [16]. Given the contradictory findings, it is crucial to reevaluate the role of KDR in OSCC, which was the objective of the present study.
Endogenous non-coding RNAs of a length of nucleotides, known as micro-RNAs, have the ability to regulate the expression of target genes. This regulation occurs through complementary pairing with the target gene’s 3′-untranslated region (3′UTR), leading to either mRNA degradation or translational inhibition [17]. It plays a vital role in various biological processes, including cancer metastasis and cell apoptosis [18]. Despite the increasing focus on micro-RNAs and their regulation of gene expression, research on KDR regulation in OSCC remains limited. However, miR-424-5p is currently being extensively studied as it is known to play complex roles in various cancers [17–19]. A study has reported the role of miR-424-5p in OSCC based on bioinformation analysis. According to the report, miR-424-5p may regulate ERBB4 [20]. However, there is no additional supporting evidence regarding the function of miR-424-5p in OSCC. Therefore, the current study also aims to explore the potential role of miRNAs in the development and progression of OSCC.
The aim of our research is to use bioinformatics analysis to explore the regulation to the expression of KDR in OSCC by miR-424-5p and lay a foundation of the further research value.
Materials and Methods
Bioinformatics analysis
The miRNA expression data were retrieved from the GSE115000 dataset, which is stored in the GEO database (https://www.ncbi.nlm.nih.gov/geo/). To identify miRNAs that were differentially expressed (DEmiRNAs), the SVA package was used for batch correction. In order to identify differentially expressed mRNAs (DEmiRNAs), the edgeR package (criteria: |logFC| >2.5, padj < 0.05) was used to compare differences between the normal group and the tumor group. Additionally, mRNA profiles were obtained for both normal samples (n = 44) and oral squamous cell carcinoma (OSCC) samples (n = 520) from the TCGA-GA clinical database (https://portal.gdc.cancer.gov/). A volcano plot was generated using the R package ‘cluster profiler’ to visualize the expression patterns of the DEmRNAs. The edgeR package was also used to analyze DEmRNAs between the normal and tumor groups, with criteria of a change in expression greater than 1 log (|logFC| > 1) and a significant p-value of less than 0.05 (padj < 0.05). The binding sites of these DEmiRNAs on their target mRNAs were predicted using miRDB, TargetScan, and microT databases. Kaplan-Meier analysis was conducted to assess survival based on the expression levels of these identified DEmiRNAs and their target mRNAs. Furthermore, potential upstream miRNAs that regulate KDR expression were predicted using data from miRDB, TargetScan, and microT databases.
Cell lines and culture
The OSCC cell lines HN4, HN6, HB96, and CAL27, as well as the normal human oral epithelial keratinocyte cell line HIOEC, were kindly provided by Shanghai Ninth People’s Hospital, which is affiliated with Shanghai Jiaotong University School of Medicine. Cells were grown in Dulbecco’s modified Eagle medium (DMEM; Sigma, St. Louis, MO, USA) containing 10% fetal bovine serum (Gibco, Carlsbad, CA, USA). The cells were incubated at 37°C in a 5% CO2 atmosphere. Cells from passages 2 to 4 were used for the experiments.
Cell transfection
To investigate the impact of miR-424-5p and KDR, we seeded cells in 12-well plates at a density of 2 × 105/well. We obtained the necessary reagents, including normal control (NC) mimics, miR-424-5p mimics, NC-overexpression vectors, and KDR-overexpression vectors, from Genomeditech (Shanghai, China). Using Lipofectamine 3000 (Thermo Fisher Scientific Inc., Waltham, MA, USA), we transfected these into GA cell lines according to the manufacturer’s instructions. Additionally, we obtained the mimic miR-424-5p sequence and si-KDR from Sangon Technologies (Shanghai, China), which we then inserted into plasmids. The transfection concentrations were 100 nM for mimic NC and miR-424-5p mimic, 50 nM for si-NC and si-KDR, and 0.5 μg/well for plasmid of oe-NC and oe-KDR.
RNA extraction and qRT-PCR analysis
The manufacturer’s instructions were followed to obtain total RNA using the Axygen RNA Miniprep Kit (Axygen, USA) two days after transfection. For each sample, 1000 ng of total RNA was used in the reverse transcription and subsequent Rt-qPCR procedures. These procedures were based on previous related research [11]. The reactions were carried out using the Light Cycler96 Real-Time PCR System (Roche, Ltd., Basel, Switzerland). The human primer sets used were as follows: human KDR: forward, 5-ggtgatctttgggatcctca-3 and reverse, 5-accttcgccttcctcagctc-3; human GAPDH: forward, 5-ccagcccatcatcctgcct-3 and reverse, 5-cctgcgcacaccaccgatcttg-3.
Colony formation assay
The colony formation assay was used to estimate cell proliferation. After 48 h of transfection, 1 × 103 cells were seeded on 6-well plates and incubated at 37°C for 9 days. The cell colonies were fixed with 5% paraformaldehyde, and 10 min later, the cell colonies were stained with 0.5% crystal violet. The number of colonies was calculated with the ImageJ Software.
Statistical analysis
GraphPad Prism 6.0 (Graphpad Software Inc., San Diego, CA, USA) was used for statistical analysis. All values are presented as the mean ± standard deviation. Differences between the experimental and control groups were evaluated using Student t-test. Results for multiple group comparisons were analyzed using Scheffe’s test and one-way analysis of variance (ANOVA) with the SPSS 22.0 software (SPSS Inc., Chicago, IL, USA). Pearson χ2 test was used to analyze the relationship between expression of miR-424-5p and KDR. Differences were determined to be significant at P < 0.05, P < 0.01, or P < 0.001, as indicated for each experiment.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on request.
Results
Association of high KDR expression with poor prognosis of OSCC
Analysis of mRNA expression data in the TGCA-GA dataset demonstrated that higher KDR expression was associated with lower survival in patients (Figure 1A, 1B). Survival analysis according to tumor grade showed poor OS in patients with high KDR expression (Figure 1B). The risk analysis indicated a higher risk rate of mortality of patient with higher KDR expression (Figure 1C). Moreover, qRT-PCR and western blot results showed that the OSCC cells had higher KDR expression than the HOK cells (Figure 1D).
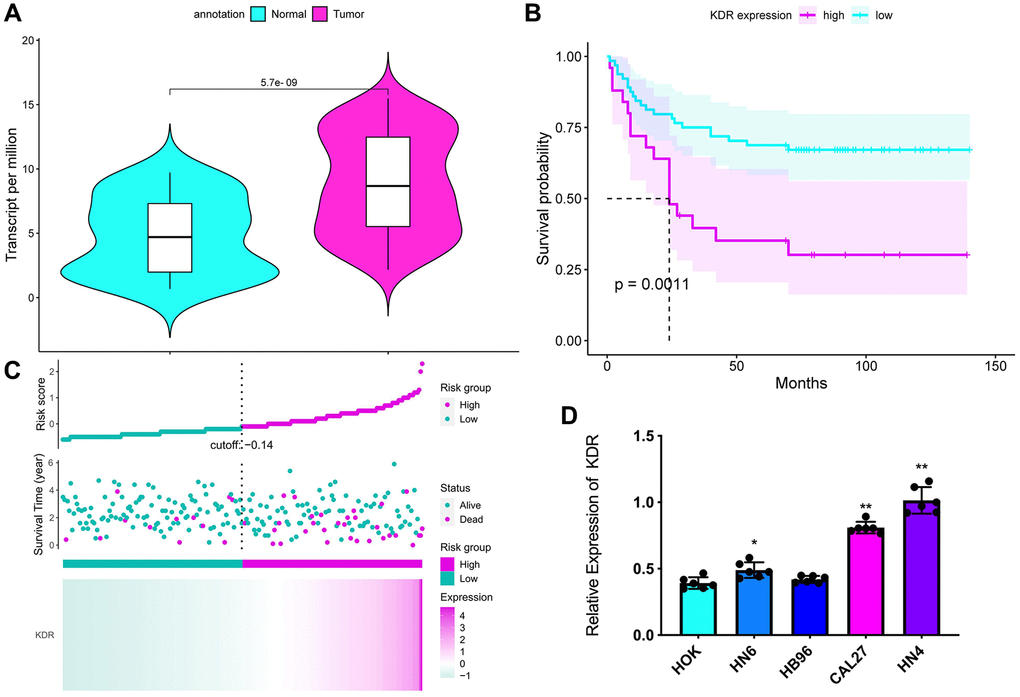
Figure 1. Expression of KDR and its association with survival in OSCC. (A) KDR expression data from the TCGA-OSCC dataset. (B) Survival curves based on KDR expression data and tumor grade from the TCGA-OSCC dataset. (C) Lasso Cox regression analysis of core circadian clock genes in OSCC. (D) mRNA expression of KDR in the normal cell line HOK and the OSCC cell lines HN6, HB96, CAL27, and HN4. (n = 3 in each group) *P < 0.05 **P < 0.01.
Suppression in OSCC cell proliferation and immigration by down-regulating KDR expression
Downregulation of mRNA expression of KDR was performed by transfection of shRNA 1-3 on CAL27 cells (Figure 2A). Downregulation of KDR resulted in a decreasing in immigration (Figure 2B), colony formation (Figure 2C), proliferation (Figure 2D).
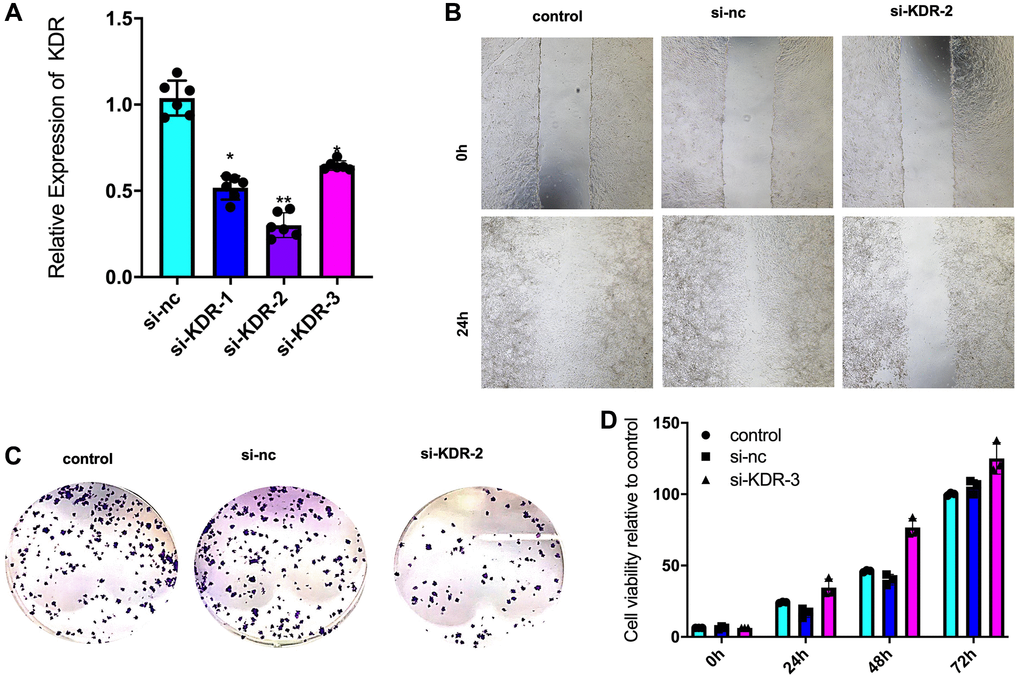
Figure 2. KDR upregulation-induced proliferation, viability, and invasion of OSCC cells. (A) mRNA downregulation of KDR in the CAL27 cell line mediated by the si-KDR 1-3 plasmid (n = 3 in each group). (B) Immigration of CAL27 cells determined by the wound healing assay (n = 3 in each group). (C) Proliferation of CAL27 cells determined by the colony formation assay (n = 3 in each group). (D) Proliferation of CAL27 cells determined by the MTT assay (n = 3 in each group). *P < 0.05 **P < 0.01.
KDR as a target gene of miR-424-5p
A total of 19349 DEmiRNAs and DEmRNA that were differentially expressed between the OSCC and normal tissue samples were identified (Figure 3A). From the downregulated DEmiRNAs, the potential upstream miRNAs of KDR were predicted using the miRDB, TargetScan, and microT databases, and from the 76 upregulated miRNAs that overlapped between these three databases, miR-424-5p was selected (Figure 3A). In agreement with these findings, miR-424-5p expression was lower in the OSCC cell lines than in the normal cell line (Figure 3B) and indicated a worse prognosis (Figure 3C). Moreover, qRT-PCR and western blot results showed that the OSCC cells had lower miR-424-5p expression than the HOK cells (Figure 3D). TargetScan was used to explore the binding regions between miR-424-5p and KDR (Figure 3E). According to the results of the dual luciferase reporter gene assay, the miR-424-5p mimic effectively downregulated luciferase activity in KDR-WT cells (Figure 3F). These results confirm that KDR gene expression was downregulated by miR-424-5p in OSCC cells.
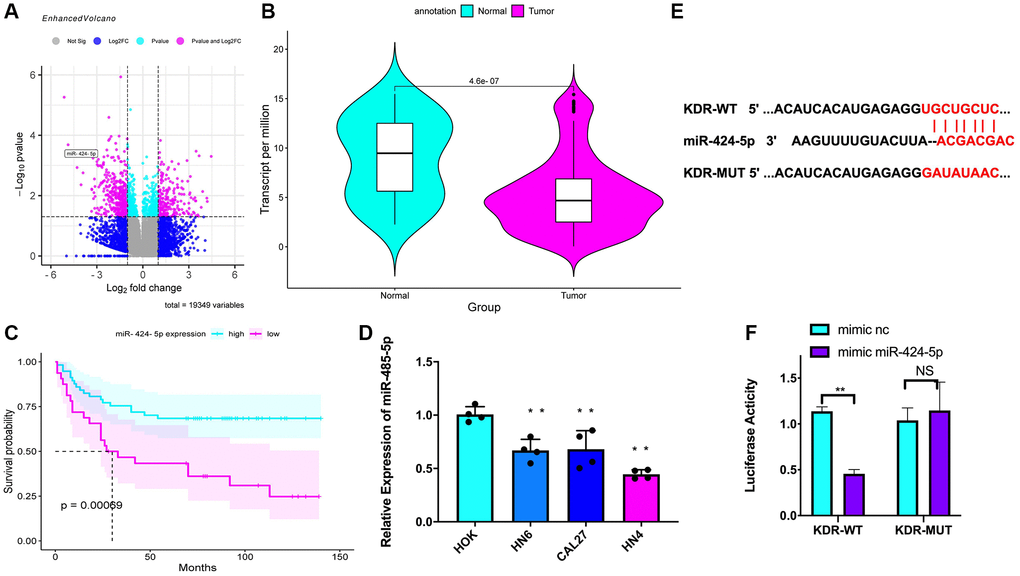
Figure 3. miR-424-5p-induced downregulation of KDR. (A) A volcano map of the differentially expressed miRNAs and mRNAs (n = 19349) identified between the normal samples and the OSCC samples based on data from the GEO dataset. (B) miR-424-5p expression data from the TCGA-OSCC dataset. (C) Survival curves based on miR-424-5p expression data and tumor grade from the TCGA-OSCC dataset. (D) Expression of miR-424-5p in the HOK cell line and OSCC cell lines (n = 4 in each group). (E) TargetScan predicts the binding site between miR-424-5p and promoter of KDR. (F) Dual luciferase reporter gene assay showing the targeted binding of miR-424-5p and KDR. *P < 0.05 **P < 0.01.
Inhibition of KDR-induced OSCC cell proliferation by miR-424-5p overexpression
Transfection of CAL27 cells with the miR-424-5p mimic resulted in significant downregulation of KDR expression (Figure 4A). Further investigation was conducted to explore the effects of miR-424-5p expression on KDR in cell functions. Compared with control group, the results of immigration (Figure 4B), colony formation (Figure 4C), invasion (Figure 4D) assays showed that the overexpress KDR induces proliferation, cell viability ability and mimic miR-424-5p reverses the function of KDR. Our results demonstrated that miR-424-5p inhibits biological function.
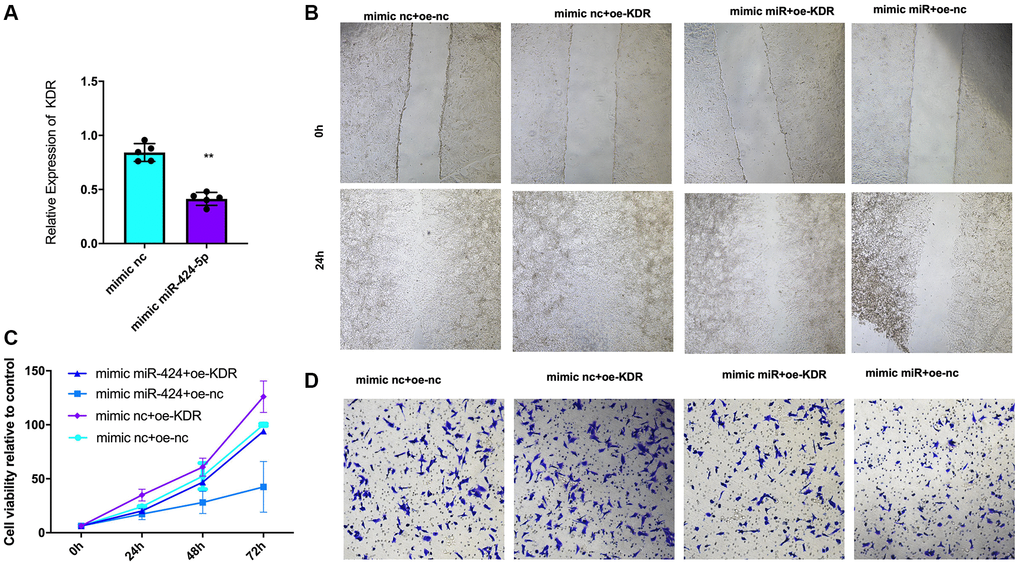
Figure 4. Effect of miR-424-5p on the proliferation, invasion, and viability of OSCC cells induced by KDR overexpression. (A) KDR expression in CAL27 cells with mimic-nc or mimic-miR-424 plasmid. (B) Immigration of CAL27 cells determined by the wound healing assay (n = 3 in each group). (C) The MTT assay was used to determine the cell viability at 24 h, 48 h, and 72 h. (n = 3 in each group). (D) Invasion of CAL27 cells determined by the wound healing assay (n = 3 in each group). *P < 0.05 **P < 0.01.
Discussion
Our research demonstrates that upregulation of KDR promotes metastasis and proliferation in OSCC. Additionally, our findings show that miR-424-5p has a regulatory effect on KDR expression in OSCC cells.
KDR is well known as a biomarker of breast cancer and has shown promising effects as a treatment target in clinical trials [14, 15, 20–22]. Recently, amplification and overexpression of KDR was reported in a subset of OSCC specimens [16, 23, 24]. However, the relationship between KDR expression and OSCC prognosis has not been explored based on clinical data. Our findings fill in this gap by analyzing relevant TCGA-GA mRNA datasets to demonstrate that there are significant differences in KDR expression between normal tissue and OSCC tissue. Additionally, the role of KDR in OSCC was confirmed by Kaplan-Meier survival analysis based on the TCGA-OSCC dataset, as higher KDR expression was associated with poorer survival. Moreover, overexpression of KDR by means of plasmid transfection demonstrated that KDR significantly induces the proliferation, migration, and invasion abilities of OSCC cells.
In this study, using bioinformatics analysis, we identified miR-424-5p as one of the miRNAs involved in the upstream regulation of KDR expression. We also found that miR-424-5p had target binding sites on KDR and a relatively high expression level in OSCC. Our findings further demonstrate that miR-424-5p could suppress the proliferation and metastasis of OSCC cells by inhibiting KDR expression. In accordance with our findings, miR-424-5p has been found to play a regulatory role in other cancers too [25, 26]. For example, Wen et al. reported that miR-424-5p suppresses lung adenocarcinoma by binding with SNHG17 [27], and in renal cancer, miR-424-5p was found to regulate MMP14 expression by binding with circPTCH1 [28]. On the contrary, miR-424-5p was found to elevate the expression of SRC, an oncogene, in ovarian cancer [29]. In this study, we were able to confirm that miR-424-5p functions in OSCC cells as an KDR regulator at the cellular level by reversing its effects on the proliferation of OSCC cells. As an exosomal miRNA derived from tumors, miR-424-5p could be a potential biomarker for the diagnosis of OSCC in the early stage. Based on these findings, in the future, we would like to explore the potential of the miR-424-5p/KDR axis in the treatment of OSCC. It may give us a promising hint that the regulation of KDR could be released by the miR-424-5p and for the micro-RNA related drugs.
To conclude, our findings reveal KDR and miR-424-5p as potential biomarkers for both diagnosis and prognosis prediction, and as therapeutic targets for improving the outcome of OSCC. Thus, future research on their potential in the diagnosis and treatment of OSCC is warranted.
Author Contributions
Junqing Zhang performed all the bioinformatic analysis and wrote the manuscript. Zhangui Tang supervised the study design and revised the manuscript. Guanghui Bao supervised the whole process of this research and takes responsibility for the quality of raw data.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Funding
No funding was used for this paper.
References
- 1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin. 2019; 69:7–34. https://doi.org/10.3322/caac.21551 [PubMed]
- 2. Chinn SB, Myers JN. Oral Cavity Carcinoma: Current Management, Controversies, and Future Directions. J Clin Oncol. 2015; 33:3269–76. https://doi.org/10.1200/JCO.2015.61.2929 [PubMed]
- 3. Singhvi HR, Malik A, Chaturvedi P. The Role of Chronic Mucosal Trauma in Oral Cancer: A Review of Literature. Indian J Med Paediatr Oncol. 2017; 38:44–50. https://doi.org/10.4103/0971-5851.203510 [PubMed]
- 4. Yarden Y, Sliwkowski MX. Untangling the ErbB signalling network. Nat Rev Mol Cell Biol. 2001; 2:127–37. https://doi.org/10.1038/35052073 [PubMed]
- 5. Pu T, Guo P, Qiu Y, Chen S, Yang L, Sun L, Ye F, Bu H. Quantitative real-time polymerase chain reaction is an alternative method for the detection of HER-2 amplification in formalin-fixed paraffin-embedded breast cancer samples. Int J Clin Exp Pathol. 2015; 8:10565–74. [PubMed]
- 6. Nathanson DR, Nash GM, Chen B, Gerald W, Paty PB. Detection of HER-2/neu gene amplification in breast cancer using a novel polymerase chain reaction/ligase detection reaction technique. J Am Coll Surg. 2003; 197:419–25. https://doi.org/10.1016/S1072-7515(03)00431-9 [PubMed]
- 7. Qiu X, Lin Q, Ning Z, Qian X, Li P, Ye L, Xie S. Quantitative bioanalytical assay for the human epidermal growth factor receptor (HER) inhibitor dacomitinib in rat plasma by UPLC-MS/MS. J Pharm Biomed Anal. 2019; 166:66–70. https://doi.org/10.1016/j.jpba.2018.12.041 [PubMed]
- 8. Bofin AM, Ytterhus B, Martin C, O'Leary JJ, Hagmar BM. Detection and quantitation of HER-2 gene amplification and protein expression in breast carcinoma. Am J Clin Pathol. 2004; 122:110–9. https://doi.org/10.1309/8A2D-JFT0-7NE6-EWHE [PubMed]
- 9. Khan AJ, King BL, Smith BD, Smith GL, DiGiovanna MP, Carter D, Haffty BG. Characterization of the HER-2/neu oncogene by immunohistochemical and fluorescence in situ hybridization analysis in oral and oropharyngeal squamous cell carcinoma. Clin Cancer Res. 2002; 8:540–8. [PubMed]
- 10. Martín-Ezquerra G, Salgado R, Toll A, Gilaberte M, Baró T, Alameda Quitllet F, Yébenes M, Solé F, Garcia-Muret M, Espinet B, Pujol RM. Multiple genetic copy number alterations in oral squamous cell carcinoma: study of MYC, TP53, CCDN1, EGFR and ERBB2 status in primary and metastatic tumours. Br J Dermatol. 2010; 163:1028–35. https://doi.org/10.1111/j.1365-2133.2010.09947.x [PubMed]
- 11. Klapper LN, Waterman H, Sela M, Yarden Y. Tumor-inhibitory antibodies to HER-2/ErbB-2 may act by recruiting c-Cbl and enhancing ubiquitination of HER-2. Cancer Res. 2000; 60:3384–8. [PubMed]
- 12. Slamon DJ, Leyland-Jones B, Shak S, Fuchs H, Paton V, Bajamonde A, Fleming T, Eiermann W, Wolter J, Pegram M, Baselga J, Norton L. Use of chemotherapy plus a monoclonal antibody against HER2 for metastatic breast cancer that overexpresses HER2. N Engl J Med. 2001; 344:783–92. https://doi.org/10.1056/NEJM200103153441101 [PubMed]
- 13. Mann M, Sheng H, Shao J, Williams CS, Pisacane PI, Sliwkowski MX, DuBois RN. Targeting cyclooxygenase 2 and HER-2/neu pathways inhibits colorectal carcinoma growth. Gastroenterology. 2001; 120:1713–9. https://doi.org/10.1053/gast.2001.24844 [PubMed]
- 14. Makiyama A, Sukawa Y, Kashiwada T, Kawada J, Hosokawa A, Horie Y, Tsuji A, Moriwaki T, Tanioka H, Shinozaki K, Uchino K, Yasui H, Tsukuda H, et al. Randomized, Phase II Study of Trastuzumab Beyond Progression in Patients With HER2-Positive Advanced Gastric or Gastroesophageal Junction Cancer: WJOG7112G (T-ACT Study). J Clin Oncol. 2020; 38:1919–27. https://doi.org/10.1200/JCO.19.03077 [PubMed]
- 15. Ignatov T, Gorbunow F, Eggemann H, Ortmann O, Ignatov A. Loss of HER2 after HER2-targeted treatment. Breast Cancer Res Treat. 2019; 175:401–8. https://doi.org/10.1007/s10549-019-05173-4 [PubMed]
- 16. Birkeland AC, Yanik M, Tillman BN, Scott MV, Foltin SK, Mann JE, Michmerhuizen NL, Ludwig ML, Sandelski MM, Komarck CM, Carey TE, Prince ME, Bradford CR, et al. Identification of Targetable ERBB2 Aberrations in Head and Neck Squamous Cell Carcinoma. JAMA Otolaryngol Head Neck Surg. 2016; 142:559–67. https://doi.org/10.1001/jamaoto.2016.0335 [PubMed]
- 17. Li W, Zheng Y, Mao B, Wang F, Zhong Y, Cheng D. SNHG17 upregulates WLS expression to accelerate lung adenocarcinoma progression by sponging miR-485-5p. Biochem Biophys Res Commun. 2020; 533:1435–41. https://doi.org/10.1016/j.bbrc.2020.09.130 [PubMed]
- 18. Liu H, Hu G, Wang Z, Liu Q, Zhang J, Chen Y, Huang Y, Xue W, Xu Y, Zhai W. circPTCH1 promotes invasion and metastasis in renal cell carcinoma via regulating miR-485-5p/MMP14 axis. Theranostics. 2020; 10:10791–807. https://doi.org/10.7150/thno.47239 [PubMed]
- 19. Yang Y, Liu J, Qian X, Li Y, Wang Y, Xu X. miR-485-5p improves the progression of ovarian cancer by targeting SRC in vitro and in vivo. Neoplasma. 2020; 67:1022–31. https://doi.org/10.4149/neo_2020_190927N979 [PubMed]
- 20. Ratovitski EA. Phospho-ΔNp63α/microRNA network modulates epigenetic regulatory enzymes in squamous cell carcinomas. Cell Cycle. 2014; 13:749–61. https://doi.org/10.4161/cc.27676 [PubMed]
- 21. Zhou Z, Chen X, Chen X, Qin A, Mao Y, Pang Y, Yu S, Zhang S. PP121 suppresses RANKL-Induced osteoclast formation in vitro and LPS-Induced bone resorption in vivo. Exp Cell Res. 2020; 388:111857. https://doi.org/10.1016/j.yexcr.2020.111857 [PubMed]
- 22. Wang X, Xiao H, Wu D, Zhang D, Zhang Z. miR-335-5p Regulates Cell Cycle and Metastasis in Lung Adenocarcinoma by Targeting CCNB2. Onco Targets Ther. 2020; 13:6255–63. https://doi.org/10.2147/OTT.S245136 [PubMed]
- 23. Liu O, Xu J, Wang F, Jin W, Zanvit P, Wang D, Goldberg N, Cain A, Guo N, Han Y, Bynum A, Ma G, Wang S, et al. Adipose-mesenchymal stromal cells suppress experimental Sjögren syndrome by IL-33-driven expansion of ST2+ regulatory T cells. iScience. 2021; 24:102446. https://doi.org/10.1016/j.isci.2021.102446 [PubMed]
- 24. Coronel R, Bernabeu-Zornoza A, Palmer C, Muñiz-Moreno M, Zambrano A, Cano E, Liste I. Role of Amyloid Precursor Protein (APP) and Its Derivatives in the Biology and Cell Fate Specification of Neural Stem Cells. Mol Neurobiol. 2018; 55:7107–17. https://doi.org/10.1007/s12035-018-0914-2 [PubMed]
- 25. Slamon D, Eiermann W, Robert N, Pienkowski T, Martin M, Press M, Mackey J, Glaspy J, Chan A, Pawlicki M, Pinter T, Valero V, Liu MC, et al, and Breast Cancer International Research Group. Adjuvant trastuzumab in HER2-positive breast cancer. N Engl J Med. 2011; 365:1273–83. https://doi.org/10.1056/NEJMoa0910383 [PubMed]
- 26. von Minckwitz G, Procter M, de Azambuja E, Zardavas D, Benyunes M, Viale G, Suter T, Arahmani A, Rouchet N, Clark E, Knott A, Lang I, Levy C, et al, and APHINITY Steering Committee and Investigators. Adjuvant Pertuzumab and Trastuzumab in Early HER2-Positive Breast Cancer. N Engl J Med. 2017; 377:122–31. https://doi.org/10.1056/NEJMoa1703643 [PubMed]
- 27. Ghanemi M, Pourshohod A, Ghaffari MA, Kheirollah A, Amin M, Zeinali M, Jamalan M. Specific Targeting of HER2-Positive Head and Neck Squamous Cell Carcinoma Line HN5 by Idarubicin-ZHER2 Affibody Conjugate. Curr Cancer Drug Targets. 2019; 19:65–73. https://doi.org/10.2174/1568009617666170427105417 [PubMed]
- 28. De Pauw I, Lardon F, Van den Bossche J, Baysal H, Fransen E, Deschoolmeester V, Pauwels P, Peeters M, Vermorken JB, Wouters A. Simultaneous targeting of EGFR, HER2, and HER4 by afatinib overcomes intrinsic and acquired cetuximab resistance in head and neck squamous cell carcinoma cell lines. Mol Oncol. 2018; 12:830–54. https://doi.org/10.1002/1878-0261.12197 [PubMed]
- 29. Jin X, Chen Y, Chen H, Fei S, Chen D, Cai X, Liu L, Lin B, Su H, Zhao L, Su M, Pan H, Shen L, et al. Evaluation of Tumor-Derived Exosomal miRNA as Potential Diagnostic Biomarkers for Early-Stage Non-Small Cell Lung Cancer Using Next-Generation Sequencing. Clin Cancer Res. 2017; 23:5311–9. https://doi.org/10.1158/1078-0432.CCR-17-0577 [PubMed]