



Dedication
This review is dedicated to the memory and honor of Dr. Mikhail Blagosklonny, a preeminent scientist in oncology, the biology of aging, longevity and personalized medicine. I am deeply grateful to Mikhail Vladimirovich for supporting my research in the field personalized approaches to the endocrinology of aging, specifically using nonhuman primate models under mild to moderate stress. My previous work was published in Aging in Volume 2, Issue 11. I hope that this new article, which reviews literature on individual differences in age-related neuroendocrine dysregulation as a foundation for stress vulnerability, accelerated aging, and age-related pathology, will be a worthy tribute to Dr. Mikhail Blagosklonny's memory.
Introduction
With aging, the frequency of stress-related diseases, including mental, metabolic, endocrine, cognitive, cardiovascular, and neurodegenerative diseases increases dramatically [1–9]. Despite this generalized phenomenon, there are notable individual differences in both vulnerability and resilience to stress and stress-related pathologies [4, 8, 10–12]. In particular, in spite of the abundance of ongoing studies in stress research, the mechanisms underlying individual vulnerability to stress and age-related diseases remain unclear. It is well known that the most important adaptive neuroendocrine systems, the hypothalamic-pituitary-adrenal (HPA) axis and hypothalamic-pituitary-thyroid (HPT) axis, are essential modulators of both the endocrine and behavioral adaptation of the organism to changing environmental factors, and their dysfunction can contribute to the development of various stress-dependent socially significant diseases. The review focuses on individual differences in age-related neuroendocrine dysregulation as a mechanistic basis for stress vulnerability, accelerated aging, and age-related pathology.
The HPA axis, a key adaptive neuroendocrine axis, reacts to all environment stimuli in a non-specific way, activating the secretion of corticotropin-releasing hormone (CRH) and arginine vasopressin (AVP) by the hypothalamic paraventricular nucleus (PVN), which in turn activate the secretion of corticotropin (ACTH) by corticotrophs of the anterior pituitary. ACTH stimulates corticosteroid secretion, mainly cortisol (CORT) and dehydroepiandrosterone (DHEA) with DHEA sulfate (DHEAS) in humans and nonhuman primates, and corticosterone in rodents. Corticosteroids act on specific receptors present in most peripheral tissues and the brain and trigger the metabolic, immune, neuromodulatory, and behavioral changes needed to cope with the impact of the stressors [10–14]. While adequate activation of the HPA axis during acute stress is critical for survival, inadequate activation (too high, prolonged, unpredictable, etc.) leads to hypercortisolemia, which is known to play a pathophysiological role in the development of various stress-related diseases, including reproductive, immune, metabolic, cardiovascular, psychiatric, and neurodegenerative [8, 10–14].
Activation of the HPT axis occurs mainly in response to a decrease in ambient temperature, physical and mental stress and other stimuli. The main component of this axis is the PVN of the hypothalamus, where thyrotropin-releasing hormone (TRH) is produced in specific parvocellular neurons and secreted into the pituitary portal system. The interaction of TRH with specific receptors on thyrotrophs of the anterior pituitary gland induces the release of thyroid-stimulating hormone (thyrotropin, TSH) into the circulation. TSH interacting with receptors on thyroid gland cells stimulates the uptake of molecular iodine from the blood and the synthesis and secretion of thyroid hormones, thyroxine (T4) and triiodothyronine (T3). Through specific cell receptors, thyroid hormones mediate the physiological effects of the HPT axis and regulate the function of this axis at the level of the pituitary and hypothalamus by a negative feedback mechanism. The HPT axis regulates diverse physiological processes such as regulation of lipid metabolism and glucose homeostasis, thermogenesis, cellular repair mechanisms, normal growth, development and differentiation of nerve cells, reproduction, homeostasis of the cardiovascular system [2, 7, 8, 15–17]. Thyroid dysfunction is considered to be associated with type 2 diabetes mellitus [18–20], metabolic syndrome [20–21], fatty liver disease [17, 22], depression [3, 23]. According to the majority of experimental data, the activity of the HPT axis decreases in response to stress exposure [24–26]. The inhibitory effect of stress on the function of the HPT axis is apparently based on the inhibitory effect of increased glucocorticoid concentrations, which can be realized at the level of pituitary thyrotrophs [25–26], the hypothalamic paraventricular nucleus [24–26], and at the level of peripheral conversion of T4 to T3 [27].
The functions of the endocrine systems generally deteriorate during aging both in basal conditions and in response to stress exposure or their specific activation, increasing the risk of age-related disorders [8, 10, 28–33]. For example, it has been found that hypercortisolemia and increased vulnerability to stress exposure are characteristic of age-related dysfunctions of the HPA axis [8, 11, 32–33]. Circulating concentrations of DHEAS peak at about age 25 and then decline gradually with age, falling to childhood concentrations by age 80 in most adults, reflecting a gradual reduction in the size of the zona reticularis [34–35]. The majority of older individuals with elevated TSH concentrations have normal free T4 concentrations, a combination of thyroid test results is known as subclinical hypothyroidism [36]. The insulin resistance and hyperinsulinemia are characteristic for age-related dysfunctions of the pancreatic islet apparatus [5, 8, 30, 37] and are usually associated with thyroid and adrenal dysfunctions [8, 18–20].
However, the type and magnitude of deterioration are not similar in all individuals and appear to be associated with different behavioral characteristics. For example, it has been found that trajectory of age-related decline of plasma DHEA (DHEAS) concentrations vary significantly among individuals, and in population studies, DHEAS concentrations are higher in men than women [8, 38]. A higher activity of the HPA axis has been noted in elderly individuals with depression and in nonhuman primates that exhibit depression and anxiety-like behavior [8, 11–12, 39, 40]. The data presented a potential risk of revealed reduced adaptive capacity the HPA axis under constant lighting in rhesus monkeys for the development of stress-related pathology and accelerated aging, especially in the depression-like/anxiety-like compromised animals [41]. Konstandi and Johnson [42] discussed the data on the activation of the HPA axis during stress and aging that can lead to disruption of hepatic cytochrome P450 (CYP)-dependent drug metabolism with pronounced inter-individual variations in the effectiveness and side effects of standard treatment protocols. The article by Degroote et al. [43] focuses on the study of the relationship between individual differences in the daily activity of the HPA axis in middle-aged and elderly people and coronary heart disease. A growing body of work also points to the regulatory influence of steroid and thyroid hormones on lifespan through modulation of key cellular pathways associated with the main hallmarks of aging and longevity [15, 44–45]. The majority of clinical studies indicate a significant predominance of age-related dysfunctions of HPA and HPT axes in women compared to men and in females compared to male nonhuman primates [1, 8–9, 12, 46], which indicates the importance of sex hormones, and especially estrogens, in age-related impairment of stress reactivity of these neuroendocrine systems.
Тhus, age-related dysfunctions of the neuroendocrine axes form the pathophysiological conditions for the development of age-related pathology. Individual differences in the characteristics and intensity of these age-related endocrine dysfunctions may underlie differences in vulnerability to age-related pathology [40]. Therefore, study of individual features of aging of the key endocrine adaptive systems is important to identify individuals with increased vulnerability to stress and accelerated aging and to develop a personalized approach to the prevention and treatment of stress-related pathology in the elderly. Biomarkers are needed both to identify individuals with increased vulnerability to stress and age-related pathologies and for understanding the mechanisms of individual differences. Dynamic responses (stress tests, circadian profiles, tests with the administration of CRH, AVP, ACTH, TRH, TSH, etc.) are more informative than basal hormone levels as biomarkers of increased vulnerability to stress and age-related pathology [8, 10–11, 31–33].
Since most studies on this problem were carried out in experiments on nocturnal rodents [28, 30, 47, 48], which differ significantly from humans in the functioning of HPA axis, and individual features of age-related changes were assessed, as a rule, only for one of the endocrine systems, mainly HPA axis, it is of considerable interest to simultaneously study the age-related features of the functioning of various parts of the endocrine system in an experiment on an adequate experimental model. Nonhuman primates, by virtue of their genetic, physiological, and behavioral similarities with humans, appear to be the optimal translational model for such research [35, 49–59].
One of the most promising experimental approaches to studying individual characteristics of the aging endocrine system is apparently the study of age-related features of the endocrine system in individuals who differ in the characteristics of higher nervous activity, in particular in behavior under conditions of mild/moderate stress [11, 31, 33, 37, 38, 51, 55, 60–63]. For example, the behavior of rhesus monkeys in an enclosure is characterized by social behavior and groups of animals with huddling and non-huddling behavior are distinguished; cortisol levels were elevated significantly in high huddlers compared with low huddlers and non-huddlers [53]. In addition, it has been demonstrated that in the marmoset monkey the serotonergic and regional brain volume correlates with trait anxiety and high-anxious marmosets showed reduced amygdala serotonin levels, and smaller volumes in a closely connected prefrontal region and the dorsal anterior cingulate cortex [51]. These findings highlight behavioral and neural similarities between trait-like anxiety in monkeys and humans. Low- and high-anxious animals were identified by behavioral responses to a human intruder [51]. Studies by other authors made it possible to distinguish animals (female rhesus monkeys) with standard control (SB) and depression-like and/or anxiety-like behavior (DAB) by studying the behavior of monkeys housed individually [31, 37, 38, 41]. During the 2-week period immediately following the transfer of monkeys from their usual residence, in groups, into the individual metabolic cages, they exhibited considerable orientation and aggressive defensive unconditioned reflexes in response to new living conditions, as well as to the procedure of bleeding. A training procedure with monkeys during 4-week after transporting animals in metabolic cages was sufficient for the elimination of these reflexes in animals with SB, but not in animals with depression-like and anxiety behavior. Behavior following the adaptation period, i.e., during experimentation was distinctly different between groups, and it was considered by the authors to be each animal's individual response to mild stress induced by the procedural stress [31, 38, 41]. The data on the polymorphism of the behavioral response of nonhuman primates to mild stress and their spontaneous anxious and depressive behavior are confirmed by data from other authors [53, 61, 64].
This review presents data on the study of age-related features in functioning of the key adaptive neuroendocrine systems, HPA and HPT axes, in individuals that differ in adaptive behavior with an emphasis on experimental studies in nonhuman primates. This review may be useful for researchers and clinicians interested in the problems of identifying individuals with increased vulnerability to stress and accelerated aging and developing a personalized approach to the prevention and treatment of stress-related pathology in the elderly.
Hypothalamic – pituitary - adrenal axis
An increasing number of studies, both clinical [45, 62, 65–68] and experimental [11, 33, 38, 48, 53, 60, 61, 69], indicate an association between the characteristics of an individual’s HPA axis response to stress and the characteristics of his adaptive behavior. However, most of these studies concerned mainly the young period of individuals' lives and did not study the mechanisms of age-related changes in HPA axis function depending on behavioral characteristics [60, 61, 66, 68, 69]. In addition, most of them assessed HPA axis response to stress without taking into account the circadian rhythm of HPA axis activity under basal conditions. At the same time, a number of publications noted the phenomenon of dependence of stress reactivity of the HPA axis on the time of day [10, 11, 45, 70–72]. In addition, many authors believe that age-related disturbances of the circadian rhythms of the HPA axis play an important role in the development of age-related diseases [11, 71, 73, 74]. Individual differences in the regulation of the HPA axis via a negative feedback mechanism based on mineralocorticoid receptors (MRs) have also been noted in humans and primates, showing a strong time-of-day dependence [31, 70, 75].
In the context of studying the age-related features of the HPA axis in individuals with various adaptive behavioral characteristics, the results of systematic studies conducted at the Adler Primatology Center using special technologies are of considerable interest. Their approach to the problem of the relationship between behavior and endocrine system was based on observations of nonhuman primates (female rhesus monkeys), usually living in enclosures or cages designed for group housing, and for the experiments moved in individual metabolic cages. It turned out that after a 4-week adaptation period, the animals behave differently. The specifics of keeping animals, their physical condition and behavior, which underlie the formation of a particular behavioral group, were described in detail earlier [31, 33, 38].
Three main types of primate behavior can be distinguished: standard (SB, healthy, control), depression-like and anxiety-like (DAB), and aggressive [31, 33, 38, 41]. Since individual adaptive behavior of animals is quite stable and characterizes the features of the higher nervous activity of individuals, in the regulation of which the endocrine system plays an important role, the authors attempted to establish a relationship between individual characteristics of stress behavior and the functioning of the endocrine system. It turned out that all the above-mentioned types of behavior differ, first of all, in the functioning of the HPA axis - a key adaptive neuroendocrine axis, the dysfunction of which plays a central role in the pathophysiology of stress and age-related pathologies.
First, they identified differences in the functioning of HPA axis in these behavioral types at aging under basal conditions, namely: in the secretion of DHEAS, the cortisol/DHEAS (CORT/DHEAS) molar ratio, the CORT plasma levels in the evening, and the amplitude of the circadian rhythm of CORT. Thus, the minimum values of DHEAS concentration and the maximum values of the CORT/DHEAS molar ratio were observed in old females with DAB behavior. Statistically significant differences were also noted between animals of different behavioral groups in relation to cortisol concentrations in the evening. It was significantly higher at 21:00, and the amplitude of the CORT circadian rhythm was correspondingly lower in animals with DAB compared to animals with other types of behavior [38]. A decrease in basal DHEA (DHEAS) levels and an increase in the cortisol/DHEAS ratio are characteristic of aging humans and primates [8, 10, 35, 76, 77].
They also hypothesized that different behavioral patterns in female rhesus monkeys may differ in the HPA axis response to stress exposure. In developing experimental approaches to assessing possible individual differences in HPA axis response to stressors, they relied on the results of their previous studies on the circadian rhythm of the HPA axis response to acute stress exposure (ASE, restraint for 2 hours) in young monkeys without taking into account their behavioral characteristics [11, 72, 78]. They demonstrated a significantly higher increase in ACTH and CORT levels in young female rhesus monkeys when the stress was applied at 15:00 then at 09:00. At the same time, they showed a smoothing of the circadian rhythm of plasma ACTH and CORT in old female monkeys and pronounced age-related differences in the stress reactivity of HPA axis at 15:00 with a lower reaction in old animals [11, 72, 78].
Based on the obtained data that the response of the HPA axis to ASE depends on the time of day, an attempt was made to identify possible individual differences in the stress reactivity of the HPA axis when exposed to stress at 15:00. It turned out, no differences in ACTH and CORT response to ASE were detected between young animals with DAB and SB [33], but intergroup differences were found in them in ACTH response to CRH and AVP tests, with a higher response in young DAB animals [79, 80]. These phenomena are illustrated in Figures 1, 2, respectively. These data indicate that, despite the absence of intergroup differences in HPA axis response to ASE, there are intergroup latent differences in the functioning of the HPA axis in young animals with DAB.
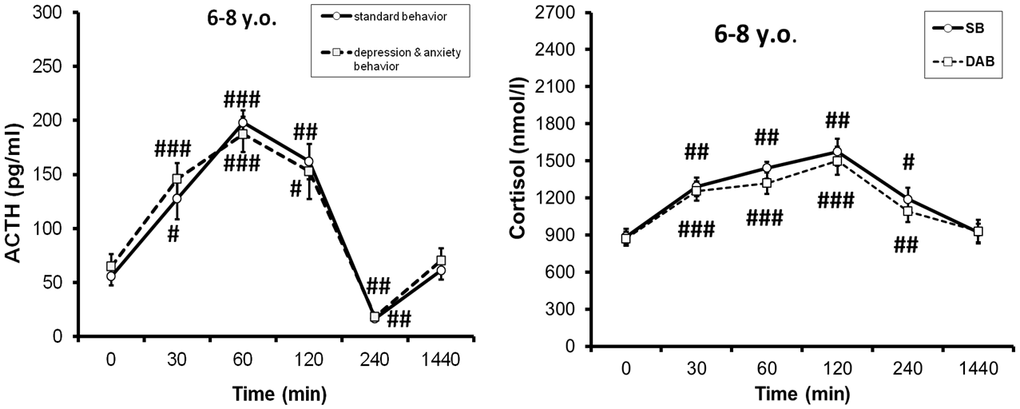
Figure 1. Dynamics of ACTH and cortisol concentrations (mean±S.E.M) in peripheral blood plasma of young female rhesus monkeys with different types of behavior in response to acute stress exposure (restraint for 2 hours, begun at 15:00h). # p<0.05, ## p<0.01, ### p<0.001– vs. basal values (0 min). Adapted with permission in [33]. Copyright. Elsevier.
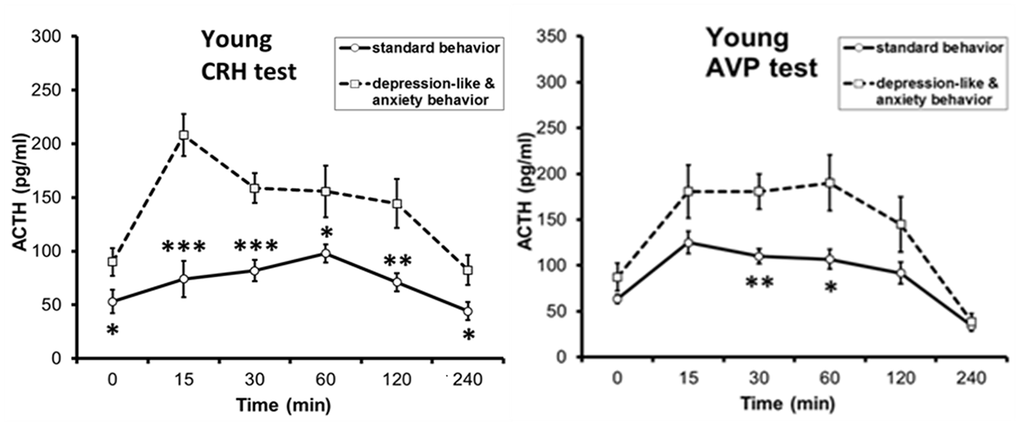
Figure 2. Dynamics of plasma ACTH level in response to functional tests with administration of CRH and AVP to young female rhesus monkeys with different types of behavior (mean±S.E.M; begun at 15:00h). *p < 0.05; **p < 0. 01; **p < 0.001 – vs. relative values in animals with DAB.
Unlike young animals, pronounced intergroup differences in ACTH response to ASE were revealed in old female rhesus monkeys with higher response in animals with DAB [33]. The identified intergroup differences in the ACTH response to ASE in old animals are apparently due to the age-related differences in the ACTH response to ASE in animals with DAB and SB. Indeed, as the experimental data showed, old animals with SB are characterized by a marked decrease in the ACTH response to ASE compared to young animals. At the same time, for old animals with DAB, the ACTH response to ASE was mainly not significantly changed or increased compared to young animals with DAB [33]. In contrast to ACTH, the rise in CORT levels in response to ASE in old DAB animals was lower than in old SB animals, presumably due to compensatory desensitization of corticocytes following central sensitization of the HPA axis [33]. Figure 3 shows the differences in ACTH and CORT responses to ASE in aged female rhesus monkeys with SB and DAB. However, the CORT/DHEAS molar ratio was higher than in SB animals, i.e., DAB females developed relative hypercortisolemia [33]. Signs of increased secretory activity of corticotrophs of the anterior pituitary gland were observed in pathomorphological material from the majority of elderly cadavers (men over 70 years of age) [81].
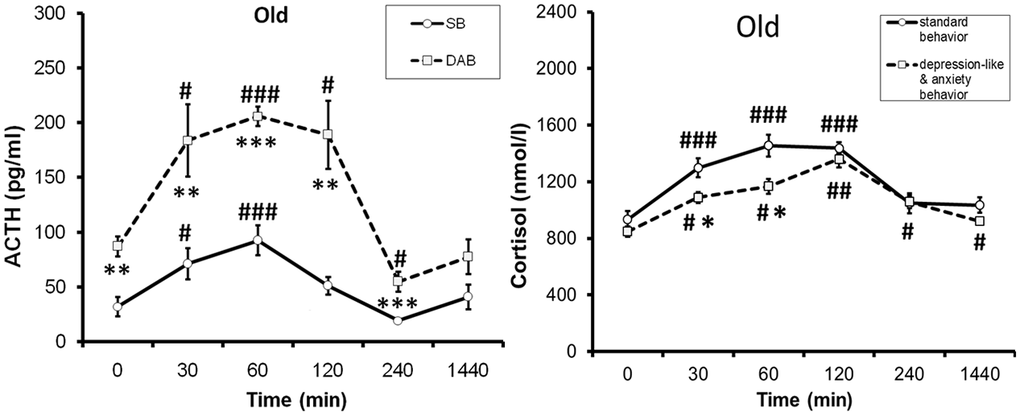
Figure 3. Dynamics of ACTH and cortisol concentrations (mean±S.E.M) in peripheral blood plasma of old female rhesus monkeys with different types of behavior in response to acute stress exposure (restraint for 2 hours, begun at 15:00). # p < 0.05, ## p < 0.01, ### p < 0.001– vs. basal values (0 min). *p < 0.05; **p < 0. 01; **p < 0.001 – vs. relative values in animals with SB. Adapted with permission in [33]. Copyright. Elsevier.
What are the mechanisms of intergroup differences in ACTH response to acute psycho-emotional stress in old female Macaca mulatta? In order to answer this question, a study was initiated to investigate the age-related features of the ACTH response to functional tests with CRH and AVP in female rhesus monkeys depending on their behavioral characteristics. As already mentioned above, intergroup differences were found in reaction of ACTH to CRH or AVP tests in young mature animals with higher response in DAB animals. It was further found that with aging in all animals, regardless of behavior, the ACTH response to the CRH test does not undergo significant changes and remains higher in old animals with DAB compared to old animals with SB [79]. At the same time, the ACTH response to the AVP test had a character similar to age-related changes in the ACTH response to the ASE, i.e., it was associated with behavioral characteristics. It decreased with aging in animals with SB, but increased in animals with DAB [10, 80, 82]. The presented experimental data are illustrated in Figure 4. Research with parallel determination of ACTH and AVP levels under stress conditions (restraint for 2 hours or insulin-induced hypoglycemia), as well as with the vasopressin V1b receptor antagonist (SSR 149415, Nelivaptan, “Axon”, Netherlands) administration allowed to conclude that the excessive ACTH response to stress exposure in old animals with DAB may be due to excessive activation of vasopressin V1b receptors on pituitary corticotrophs, and the use of antagonists of these receptors is promising for its prevention [80, 82].
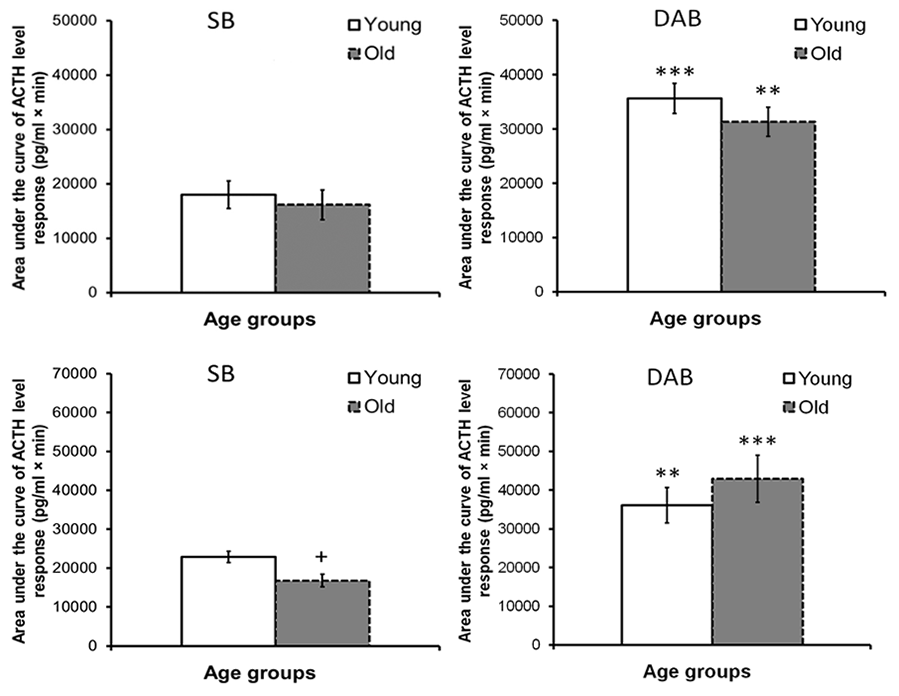
Figure 4. The area under the curves of ACTH level response to functional tests with administration of CRH (at top) and AVP (at bottom) in young and old female rhesus monkeys with different types of behavior (mean±S.E.M; begun at 15:00h). **p < 0.01; **p < 0.001 – vs. relative values in animals with SB; + p <0.05 – age-related differences.
Since the search for individual differences in the HPA axis response to ASE in female rhesus monkeys was conducted in the afternoon and evening time, the period of greatest sensitivity of the HPA axis to ASE [72, 78] and minimal circadian activity of the HPA axis [11, 70, 73, 76, 83], it could also be assumed that, with aging, intergroup differences in the regulation of the HPA axis by the negative feedback mechanism at the level of mineralocorticoid receptors (MR) are possible. Indeed, the functional test with Fludrocortisone (FlUD, agonist of MR) revealed the existence of pronounced intergroup differences in HPA axis response to FlUD, with lower sensitivity in old animals with DAB. The differences were mainly due to the slower rate of onset of the inhibitory effect of FLUD on CORT secretion compared to placebo in DAB-treated than SB-treated animals [31]. The consequence of intergroup differences in HPA axis response to FLUD was the revealed intergroup differences in the functioning of the HPA axis in old animals under basal conditions during the period of minimal circadian activity of the HPA axis (18:00–22:00) with higher levels of ACTH and CORT in animals with DAB, i.e. absolute hypercortisolemia develops in individuals with DAB [31].
The data obtained in the experiment on nonhuman primates are in good agreement with the results of a number of clinical studies, which detected MR transcripts in postmortem brains of depressed patients in lower amounts in the hippocampus and prefrontal cortex compared to non-depressed individuals [75, 84, 85, 86]. It has also been found that MR in the hippocampus is activated in patients with depression during a course of antidepressants [75, 84, 86], and stimulation of MR with FLUD decreases the secretion of CORT and improves cognitive function in individuals with depression [87]. As is known, the hippocampal MR plays a leading role in controlling the activity of the HPA axis in the evening in humans and nonhuman primates [14, 85, 88]. It is also possible that female rhesus monkeys with DAB contain a polymorphic form of the MR gene, which is more vulnerable to stress and the aging process. For instance, NR3C2 is found in a number of healthy people, as well as in patients with depression and other mental illnesses [85, 86]. In contrast to the results of the FLUD test, no significant intergroup differences were found in sensitivity of the HPA axis to dexamethasone, an agonist of glucocorticoid receptors, in old female rhesus monkeys with DAB and SB [31].
It should be emphasized that a number of changes in the functioning of the HPA axis during aging in primates with DAB described above are in good agreement with the results of studies of HPA axis function in adult people and animals exposed to severe early life stress (ELS). As shown by studies in humans and animals underwent childhood trauma, ELS induces long-term hyperactivity of the HPA axis, which is associated with changes in DNA methylation of various genes involved in the regulation of HPA axis activity, in particular, GR, FKBP51, CRH, AVP, pituitary proopiomelanocortin [66, 85, 89, 90], induce depression and other psychiatric disorders in humans [66, 85, 89, 91], promote to the development of mood disorders in nonhuman primates during adolescence [92, 93]. A stimulating effect of maternal deprivation experienced by female rhesus monkeys in neonatal period on the development of DAB behavior, as well as dysfunctions in the HPA axis in adulthood and aging, has been revealed [94].
Thus, it was revealed that there are pronounced age-related differences in the functioning of the HPA axis in monkeys with DAB compared to SB under basal conditions and under stress, such as smoothing of the circadian rhythm of CORT with an increase in its concentration at night, increased tone of the vasopressinergic system, impairment negative feedback regulation based on mineralocorticoid receptors with an increase in basal levels of CORT in the evening (18:00-22:00) in old animals with DAB. The latter were also characterized by a higher ACTH response and the CORT/DHEAS molar ratio to ASE, and a weakening of the sensitivity of the adrenal cortex to ACTH. In turn, hypercortisolemia, including relative (increase CORT/DHEAS molar ratio in response to ASE), which develops during aging in animals with DAB, can be an important pathophysiological factor in the development of dysfunctions of the HPT axis, the pancreatic islet apparatus, disorders of carbohydrate and lipid homeostasis, and contribute to the development of various aging-associated diseases, including mental, diabetes 2 type, neurodegenerative and others [8, 11–14, 40, 45, 77, 95]. The identified age-related disturbances in circadian rhythmicity in various aspects of HPA axis functioning in animals with DAB may be important for adequate adaptation to environmental stress, because almost all physiological effects of glucocorticoids from regulation of metabolic and immune signaling to effects on cognitive processes and host behavior, are exposed to similar rhythms showing a strong dependence on time of day and the close relationship between HPA axis activity and the circadian system [11, 70, 72, 73, 76, 83]. In addition, elevated CORT levels are negatively correlated with a decrease in life expectancy in humans [44] and survival in a wild primate population [96].
Hypothalamic – pituitary - thyroid axis
The results of experimental studies of the features of the functioning of the HPT axis in the same female rhesus monkeys, in which individual and age-related features of the activity of the HPA axis were studied (see section above), made it possible to identify a number of significant individual differences in HPT axis functioning, which largely correlated with individual and age-related differences in the activity of HPA axis. Thus, thyrotropin (TSH) and free fractions of triiodothyronine (T3) and thyroxine (T4) in female rhesus monkeys exhibit circadian profiles in the circulation that are negatively correlated with the circadian profile of ACTH and CORT. Unlike ACTH and CORT plasma profiles, the minimum values of TSH and free fraction of T4 in young rhesus monkeys were detected in the morning hours (09:00) and the maximum values were detected at 22:00-03:00 [41, 54, 97]. In humans, the concentration of TRH, TSH, T3, and T4 demonstrated a nocturnal peak around 02:00–04:00 and the minimum during daytime [98–101].
The circadian rhythm of TSH in old rhesus monkeys, regardless of behavioral features, retained the same profile as in young female rhesus monkeys [54, 97]. At the same time, pronounced intergroup differences were revealed in the characteristics of age-related changes in the concentration of thyroid hormones. Thus, aging does not alter the levels of circulating T3 but decreases circulating levels of T4 in rhesus monkeys [102, 103, 104, 105, 106] and female western lowland gorilla (Gorilla gorilla gorilla) [104] or leads to a simultaneous decrease in both fractions of thyroid hormones as in females, as in males cynomolgus monkeys (Macaca facsicularis) probably due to the combination of altered sensitivity of the thyroid gland to TSH and decreased secretory activity of the thyroid gland [106]. Concentration of free T4 remained without significant changes with aging in female rhesus monkeys with SB, but statistically significantly decreased in old animals with DAB at 9:00, 15:00, and 22:00 [97]. The levels of free T3 fraction during aging in animals with SB statistically significantly decreased at 15:00, 22:00, and 03:00, but not at 9:00, and remained without significant changes in old animals with DAB [97]. The discussed age differences in the characteristics of thyroid hormone circadian rhythms in nonhuman primates with DAB and SB behavior are original and are illustrated in Figure 5.
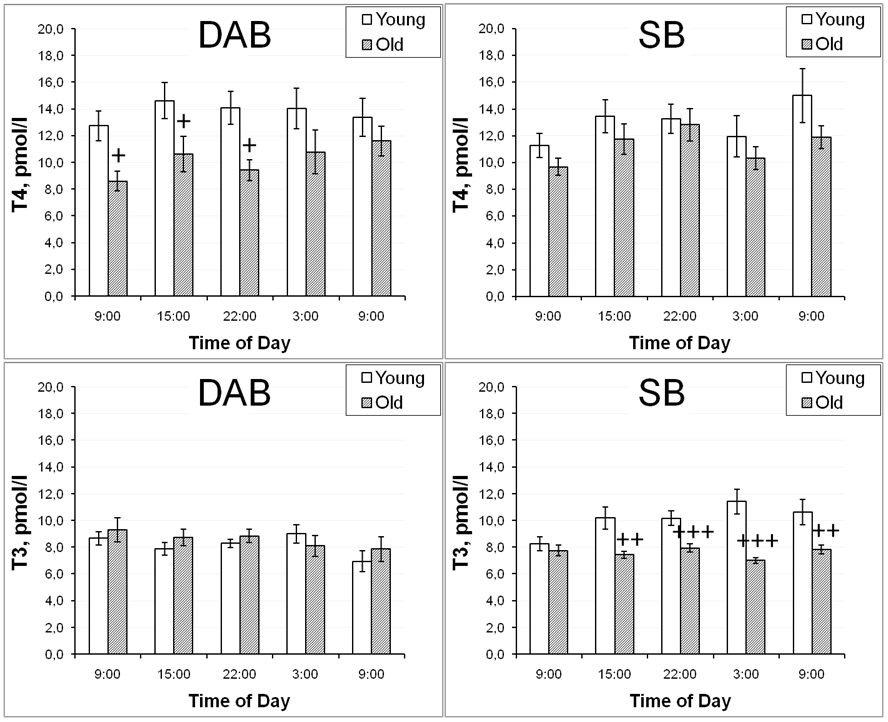
Figure 5. Dynamics of free T4 and free T3 levels in peripheral blood plasma of young and old female rhesus monkeys with various types of behavior at different time of day (mean±S.E.M.). + p<0.05, ++ p<0.01, +++ p<0.0001 vs. relative values in old animals.
A decrease in serum T3 levels with aging has been repeatedly observed in healthy people of both sexes [8, 101, 107]. A number of clinical studies have also noted a decrease in T4 secretion with age [18, 108, 109]. In addition, it was noted that in elderly individual subclinical hypothyroidism (increased TSH concentrations and normal levels of thyroid hormones in the blood serum) is more common than overt hypothyroidism [110–112].
The differences in the direction of age-related changes in thyroid hormone concentrations in primates with SB with a decrease in free T3, as well as the results of clinical studies that found a marked decrease in the plasma T3 levels of healthy aging people, are apparently due to a marked change in the activity of peripheral iodothyronine deiodinases. It is known that iodothyronine deiodinases in peripheral tissues and brain (iodothyronine deiodinase-1 and iodothyronine deiodinase-2) convert T4 to T3 and iodothyronine deiodinase-3 convert T3 to inactive reverse T3 (rT3) [98, 113]. It is possible that with aging in monkeys with SB, the activity of deiodinases is impaired in the afternoon and nighttime, which leads to a marked decrease in the concentration of free T3 not in the morning hours, but during the day and at night. Indeed, there is evidence in the literature that aging reduces the activity of iodothyronine deiodinase-1 and increases iodothyronine deiodinase-3 activity [114, 115].
In contrast to thyroid hormones, TSH levels tended to increase with age in rhesus monkeys of both behavioral groups (SB and DAB) [116]. Most clinical data also suggest that TSH levels tend to increase with age [110, 117, 118, 119, 120]. It should also be noted that there were statistically significant intergroup differences in basal TSH values in young animals with DAB and SB, with lower values in animals with DAB in the absence of significant intergroup differences in the concentration of thyroid hormones [116]. These intergroup differences in TSH levels in young monkeys may be due to a lower concentration of TRH receptors on the pituitary thyrotrophs in females with DAB, which in turn may be due to a decreased TRH secretion in the PVN of the hypothalamus. Thus, a marked decrease in mRNA expression for TRH was detected in pathological material from patients with depression [121].
The rhesus monkeys with SB and DAB differed not only in age-related features of the functioning of the HPT axis under basal conditions, but also in their response to the administration of specific activators of thyroid function (TRH and TSH). Thus, for animals with DAB, more pronounced age-related disturbances in the HPT axis response to the functional test with TRH were revealed [116]. They were characterized by a statistically significant decrease in the magnitude of the rise in the concentration of free T4 and a pronounced increase in the magnitude of the rise in the concentration of TSH in the absence of significant age-related changes in the response of free T4 and TSH in monkeys with SB. In contrast to the free T4 and TSH secretion, no significant age-related differences in the response of free T3 to this test neither in animals with DAB, nor in animals with SB were found [116]. The opposite changes in the T4 and TSH reactions in response to the TRH test in old monkeys with DAB indicated a possible damage to the cells of the thyroid gland itself, including a weakening of sensitivity of thyroid thyrocytes to TSH. Indeed, an additional functional test with TSH administration to female rhesus monkeys with SB and DAB of both age groups revealed a lower secretory response of the thyroid gland in old animals with DAB compared to young animals with the same behavioral type. At the same time, in animals with SB, no significant age-related differences were found in the response of thyroid hormones to the TSH test [97]. Figure 6 shows age-related differences in the response of free T4 to TSH administration in female rhesus monkeys with DAB and SB. These data suggested that with aging, DAB females develop primary dysfunction of thyrocytes, leading to a decrease in the secretion of free T4 and, through a negative feedback mechanism, to the activation of the hypothalamic-pituitary axis. Similar age-related changes in the functioning of the HPT axis were also noted by other authors in an experiment on the cynomolgus monkeys of different ages without taking into account the peculiarities of their adaptive behavior [103].
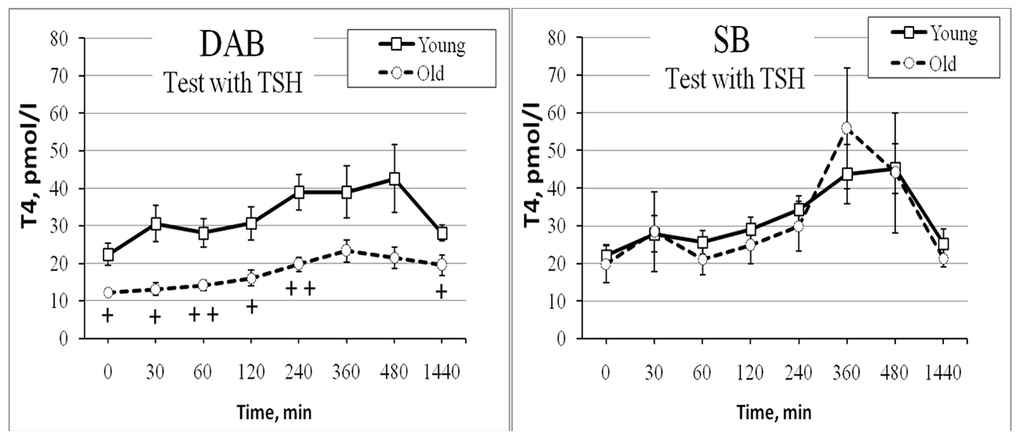
Figure 6. Dynamics of free T4 level in peripheral blood plasma of old female rhesus monkeys with different types of behavior in response to administration of TSH at 09:00h (mean±S.E.M.). + p < 0.05; ++ p <0.01– vs. relative values in young animals.
The revealed differences in the functioning of HPA and HPT axes in old rhesus monkeys with DAB compared to healthy SB monkeys give reason to believe that there are differences in age-related changes of metabolic processes in these behavioral groups, since, as already indicated above (see sections: Introduction, Hypothalamic-pituitary-adrenal axis), thyroid and steroid hormones regulate lipid and carbohydrate metabolism, and also affect the function of the islet apparatus of the pancreas [8, 17–22, 29, 45, 77, 95, 97, 122–125]. Indeed, a positive correlation was found between thyroid activity (plasma free T4 level) and body mass index in young monkeys and a negative correlation in old ones, regardless of behavioral characteristics [116]. An increase in triglyceride concentrations with age was observed in both DAB and SB female rhesus monkeys, regardless of body weight [79, 97]. However, age-related changes in triglyceride concentrations in overweight animals were behaviorally dependent: in old animals with DAB, triglyceride levels were lower than in old animals with SB [79, 97]. More pronounced indicators of insulin resistance and impaired insulin secretion were also noted in old female rhesus monkeys with DAB and overweight compared to old monkeys with SB and overweight [37]. Apparently, a deficiency of compensatory insulin secretion in old DAB monkeys with overweight, which is known to stimulate the synthesis of triglycerides and fatty acids, underlies the identified phenomenon with triglyceride levels. In turn, triglycerides and fatty acids are used by cells as energy material in severe insulin deficiency [19]. It should also be noted that, along with more pronounced disorders in age-related changes in the functions of neuroendocrine systems and the metabolism of lipids and glucose, the female rhesus monkeys with DAB showed the greatest age-related disturbances (decrease) in the activity of the antioxidant enzyme defense system and the activation of lipid peroxidation process, one of the well-known biomarkers of acceleration aging and age-related pathology [33, 126].
The results of experimental studies on nonhuman primates are in good agreement with the clinical trial data [108, 111, 112, 115, 127, 128]. Thus, it was demonstrated that patients with overt hypothyroidism have a higher chance of developing type 2 diabetes [18, 115, 125, 129, 130]. Subclinical hypothyroidism increases insulin resistance in normoglycemic people [131]. Obesity is an important factor not only in insulin resistance of various tissues, but also in the long-term increase in the secretory capacity of the β-cells of the pancreas [127]. A clear correlation between body mass index and β-cell mass was shown in humans [128]. In addition, it has been repeatedly noted that dysfunctions of both the HPT axis and the pancreas are closely associated with obesity [19, 122, 127, 132, 133]. At the same time, the association between low thyroid function and an increased risk of diabetes and cardiovascular pathology was noted [16, 20, 111, 112, 115, 132].
Thus, with aging changes in the functioning of the HPT axis develop in primates, the features and severity of which depend on the psycho-physiological characteristics of the individual, with more pronounced disturbances in animals with DAB. Old animals with DAB develop hypothyroxinaemia in the morning, evening, and night time in the absence of significant changes in the concentration of free T3. At the same time old animals with SB develop a decrease in plasma T3 during the day and night hours, a period of increased circadian thyroid activity. Age-related changes in the HPT axis function in primates are accompanied by pronounced changes in lipid metabolism, glucose homeostasis, and insulin secretion, correlating with the characteristics of adaptive behavior and body weight of the individual. More pronounced metabolic disturbances are characteristic of overweight animals with DAB. The results of studies in nonhuman primates may underlie the inconsistency of the results of clinical studies examining the function of the HPT regardless of the adaptive behavior of individuals (their psychophysiological characteristics), who did or did not observe a decline in thyroid function with aging and they are consistent with the results of clinical studies that have revealed different trajectories of metabolic, cognitive, and mental aging [2–4, 6, 12] and in light of new data on the regulatory influence of thyroid hormones on lifespan through modulation of key cellular pathways associated with longevity and the main hallmarks of aging [15]. The data on the decrease in the level of free T3 in the blood plasma of old monkeys with SB not in the morning hours, but in the afternoon - night time may have important practical significance, since they indicate the prospects of the procedure for collecting blood samples in the diagnosis of thyroid dysfunction in the elderly not only at 9:00, but also at other times of the day, for example, at 15:00 or 22:00.
In conclusion, changes in the demographic situation with an increase in the proportion of elderly and senile people, and an expansion of the range of stressful influences have caused a sharp increase in the incidence of age-related socially significant diseases (psychiatric, metabolic, endocrine, cognitive, cardiovascular, neurodegenerative) in the etiopathogenesis of which an important role is given to dysfunctions of HPA and HPT axes, the most important adaptive neuroendocrine axes. Clinical studies demonstrate the existence of distinct trajectories of cognitive and metabolic aging. Experimental studies highlight the importance of steroid and thyroid hormones in regulating key cellular pathways associated with longevity and the main hallmarks of aging [15]. Of significant interest in this regard are also physiological studies conducted in experiments on primates, the best translational model, aimed at finding biomarkers of increased vulnerability and resistance of the body to stressful influences and identifying the characteristics of stress exposure in old and senile age. Among them, studies on the study of individual characteristics of changes in the function of HPA and HPA axes and a number of metabolic indicators in primates at different age periods using medical functional tests are quite informative and of significant scientific and practical interest. The use of diagnostic tests has revealed a number of indicators that can be used to identify individuals predisposed to the development of age-related pathology at a young age: DAB behavior under mild stress, a history of severe stress in early childhood, an increased ACTH response to CRH and AVP, reduced basal TSH levels, excess body weight, and signs of impaired glucose tolerance. A deeper understanding of the age-related changes in neuroendocrine function and their role in aging pathways may provide information on new strategies to promote healthy aging, including modulation of thyroid and adrenal hormones and personalized endocrine optimization. Monitoring the behavior of individuals, as well as the functions of key adaptive endocrine systems, is promising for the early diagnosis of age-related pathology, its prevention and personalized treatment.
Conflicts of Interest
The author declares no conflicts of interest.
References
- 1. Franks KH, Bransby L, Saling MM, Pase MP. Association of Stress with Risk of Dementia and Mild Cognitive Impairment: A Systematic Review and Meta-Analysis. J Alzheimers Dis. 2021; 82:1573–90. https://doi.org/10.3233/JAD-210094 [PubMed]
- 2. Lu Y, Ye S, Gu Y, Xia Q, Hou L. Central and Peripheral Sensitivity to Thyroid Hormones Correlate to Metabolically Obesity Phenotypes in Chinese Euthyroid Adults: A Cross-Sectional Study. Diabetes Metab Res Rev. 2024; 40:e3849. https://doi.org/10.1002/dmrr.3849 [PubMed]
- 3. Ludgate S, McGowan A, Moran C, Attanasio R, Žarković M, Nagy EV, Negro R, Papini E, Adler Cohen C, Akarsu E, Alevizaki M, Ayvaz G, Bednarczuk T, et al. The use of thyroid hormones in the treatment of euthyroid patients with treatment-resistant depression. Data from a survey of 5695 European national endocrine professional organization members. Front Endocrinol (Lausanne). 2025; 16:1665720. https://doi.org/10.3389/fendo.2025.1665720 [PubMed]
- 4. Rodrigues EA, Djiberou Mahamadou AJ, Moreno S. The impact of lifestyle factors on trajectories of cognitive subtypes in the older adult population. Sci Rep. 2025; 15:31744. https://doi.org/10.1038/s41598-025-91171-0 [PubMed]
- 5. Stewart AJ, Tuncay E, Pitt SJ, Rainbow RD. Editorial: Insulin resistance and cardiovascular disease. Front Endocrinol (Lausanne). 2023; 14:1266173. https://doi.org/10.3389/fendo.2023.1266173 [PubMed]
- 6. Rao PP, Mishra S. Genetic convergence in brain aging and neurodegeneration: from cellular mechanisms to therapeutic targets. J Neurogenet. 2025; 39:140–59. https://doi.org/10.1080/01677063.2025.2571127 [PubMed]
- 7. Yan X, Xu P, Sun X. Circadian rhythm disruptions: A possible link of bipolar disorder and endocrine comorbidities. Front Psychiatry. 2023; 13:1065754. https://doi.org/10.3389/fpsyt.2022.1065754 [PubMed]
- 8. Cappola AR, Auchus RJ, El-Hajj Fuleihan G, Handelsman DJ, Kalyani RR, McClung M, Stuenkel CA, Thorner MO, Verbalis JG. Hormones and Aging: An Endocrine Society Scientific Statement. J Clin Endocrinol Metab. 2023; 108:1835–74. https://doi.org/10.1210/clinem/dgad225 [PubMed]
- 9. Liu Q, He H, Yang J, Feng X, Zhao F, Lyu J. Changes in the global burden of depression from 1990 to 2017: Findings from the Global Burden of Disease study. J Psychiatr Res. 2020; 126:134–40. https://doi.org/10.1016/j.jpsychires.2019.08.002 [PubMed]
- 10. Goncharova ND. Stress responsiveness of the hypothalamic-pituitary-adrenal axis: age-related features of the vasopressinergic regulation. Front Endocrinol (Lausanne). 2013; 4:26. https://doi.org/10.3389/fendo.2013.00026 [PubMed]
- 11. Goncharova ND. The HPA Axis under Stress and Aging: Individual Vulnerability is Associated with Behavioral Patterns and Exposure Time. Bioessays. 2020; 42:e2000007. https://doi.org/10.1002/bies.202000007 [PubMed]
- 12. Pereira-Figueiredo I, Umeoka EHL. Stress: Influences and determinants of psychopathology. Encyclopedia. 2024; 4:1026–43. https://doi.org/10.3390/encyclopedia4020066
- 13. Herman JP, Nawreen N, Smail MA, Cotella EM. Brain mechanisms of HPA axis regulation: neurocircuitry and feedback in context Richard Kvetnansky lecture. Stress. 2020; 23:617–32. https://doi.org/10.1080/10253890.2020.1859475 [PubMed]
- 14. Gupta A. Role of Stress and Hormones of the Hypothalamic-Pituitary-Adrenal (HPA) Axis in Aging. In: Models, Molecules and Mechanisms in Biogerontology, Rath P, ed. (Singapore: Springer), pp. 241–63. https://doi.org/10.1007/978-981-13-3585-3_12
- 15. Mazza AD. Thyroid Hormones and Aging: Modulators of Mitochondrial Health, Metabolic Flexibility, and Longevity Pathways. Horm Metab Res. 2026; 58:50–5. https://doi.org/10.1055/a-2698-0521 [PubMed]
- 16. Neves JS, Fontes-Carvalho R, Borges-Canha M, Leite AR, von Hafe M, Vale C, Martins S, Guimarães JT, Carvalho D, Leite-Moreira A, Azevedo A. Association of thyroid function, within the euthyroid range, with cardiovascular risk: The EPIPorto study. Front Endocrinol (Lausanne). 2022; 13:1067801. https://doi.org/10.3389/fendo.2022.1067801 [PubMed]
- 17. Zhang X, Li R, Chen Y, Dai Y, Chen L, Qin L, Cheng X, Lu Y. The Role of Thyroid Hormones and Autoantibodies in Metabolic Dysfunction Associated Fatty Liver Disease: TgAb May Be a Potential Protective Factor. Front Endocrinol (Lausanne). 2020; 11:598836. https://doi.org/10.3389/fendo.2020.598836 [PubMed]
- 18. Ogbonna SU, Ezeani IU. Risk Factors of Thyroid Dysfunction in Patients With Type 2 Diabetes Mellitus. Front Endocrinol (Lausanne). 2019; 10:440. https://doi.org/10.3389/fendo.2019.00440 [PubMed]
- 19. Biondi B, Kahaly GJ, Robertson RP. Thyroid Dysfunction and Diabetes Mellitus: Two Closely Associated Disorders. Endocr Rev. 2019; 40:789–824. https://doi.org/10.1210/er.2018-00163 [PubMed]
- 20. Laclaustra M, Moreno-Franco B, Lou-Bonafonte JM, Mateo-Gallego R, Casasnovas JA, Guallar-Castillon P, Cenarro A, Civeira F. Impaired Sensitivity to Thyroid Hormones Is Associated With Diabetes and Metabolic Syndrome. Diabetes Care. 2019; 42:303–10. https://doi.org/10.2337/dc18-1410 [PubMed]
- 21. Teixeira PF, Dos Santos PB, Pazos-Moura CC. The role of thyroid hormone in metabolism and metabolic syndrome. Ther Adv Endocrinol Metab. 2020; 11:2042018820917869. https://doi.org/10.1177/2042018820917869 [PubMed]
- 22. Borges-Canha M, Neves JS, Mendonça F, Silva MM, Costa C, Cabral PM, Guerreiro V, Lourenço R, Meira P, Salazar D, Ferreira MJ, Pedro J, Leite A, et al. Thyroid Function and the Risk of Non-Alcoholic Fatty Liver Disease in Morbid Obesity. Front Endocrinol (Lausanne). 2020; 11:572128. https://doi.org/10.3389/fendo.2020.572128 [PubMed]
- 23. Tang R, Wang J, Yang L, Ding X, Zhong Y, Pan J, Yang H, Mu L, Chen X, Chen Z. Subclinical Hypothyroidism and Depression: A Systematic Review and Meta-Analysis. Front Endocrinol (Lausanne). 2019; 10:340. https://doi.org/10.3389/fendo.2019.00340 [PubMed]
- 24. Helmreich DL, Parfitt DB, Lu XY, Akil H, Watson SJ. Relation between the hypothalamic-pituitary-thyroid (HPT) axis and the hypothalamic-pituitary-adrenal (HPA) axis during repeated stress. Neuroendocrinology. 2005; 81:183–92. https://doi.org/10.1159/000087001 [PubMed]
- 25. Sun Q, Liu A, Ma Y, Wang A, Guo X, Teng W, Jiang Y. Effects of forced swimming stress on thyroid function, pituitary thyroid-stimulating hormone and hypothalamus thyrotropin releasing hormone expression in adrenalectomy Wistar rats. Exp Ther Med. 2016; 12:3167–74. https://doi.org/10.3892/etm.2016.3790 [PubMed]
- 26. Feng G, Kang C, Yuan J, Zhang Y, Wei Y, Xu L, Zhou F, Fan X, Yang J. Neuroendocrine abnormalities associated with untreated first episode patients with major depressive disorder and bipolar disorder. Psychoneuroendocrinology. 2019; 107:119–23. https://doi.org/10.1016/j.psyneuen.2019.05.013 [PubMed]
- 27. Bianco AC, Nunes MT, Hell NS, Maciel RM. The role of glucocorticoids in the stress-induced reduction of extrathyroidal 3,5,3'-triiodothyronine generation in rats. Endocrinology. 1987; 120:1033–8. https://doi.org/10.1210/endo-120-3-1033 [PubMed]
- 28. Solas M, Aisa B, Mugueta MC, Del Río J, Tordera RM, Ramírez MJ. Interactions between age, stress and insulin on cognition: implications for Alzheimer's disease. Neuropsychopharmacology. 2010; 35:1664–73. https://doi.org/10.1038/npp.2010.13 [PubMed]
- 29. Abiri B, Ahmadi AR, Mahdavi M, Hosseinpanah F, Amouzegar A, Valizadeh M. Association between different metabolic phenotypes and the development of hypothyroidism: 9 years follow-up of Tehran thyroid study. Front Endocrinol (Lausanne). 2023; 14:1134983. https://doi.org/10.3389/fendo.2023.1134983 [PubMed]
- 30. Snyder JT, Darko C, Sharma RB, Alonso LC. Endoplasmic Reticulum Stress Induced Proliferation Remains Intact in Aging Mouse β-Cells. Front Endocrinol (Lausanne). 2021; 12:734079. https://doi.org/10.3389/fendo.2021.734079 [PubMed]
- 31. Goncharova N, Chigarova O, Rudenko N, Oganyan T. Glucocorticoid Negative Feedback in Regulation of the Hypothalamic-Pituitary-Adrenal Axis in Rhesus Monkeys With Various Types of Adaptive Behavior: Individual and Age-Related Differences. Front Endocrinol (Lausanne). 2019; 10:24. https://doi.org/10.3389/fendo.2019.00024 [PubMed]
- 32. Oikonomakos I, Siow R, Bornstein SR, Steenblock C. The Role of Growth Hormone-Releasing Hormone and the Hypothalamic-Pituitary-Somatotropic Axis in Aging: Potential Therapeutic Applications and Risks. Horm Metab Res. 2025; 57:662–7. https://doi.org/10.1055/a-2641-1207 [PubMed]
- 33. Goncharova ND, Oganyan TE. Age-related differences in stress responsiveness of the hypothalamic-pituitary-adrenal axis of nonhuman primates with various types of adaptive behavior. Gen Comp Endocrinol. 2018; 258:163–72. https://doi.org/10.1016/j.ygcen.2017.08.007 [PubMed]
- 34. Auchus RJ, Rainey WE. Adrenarche - physiology, biochemistry and human disease. Clin Endocrinol (Oxf). 2004; 60:288–96. https://doi.org/10.1046/j.1365-2265.2003.01858.x [PubMed]
- 35. Goncharova ND. [The hormonal function of the adrenals and gonads in man and monkeys during aging]. Zh Evol Biokhim Fiziol. 1997; 33:44–51. [PubMed]
- 36. Stott DJ, Rodondi N, Kearney PM, Ford I, Westendorp RG, Mooijaart SP, Sattar N, Aubert CE, Aujesky D, Bauer DC, Baumgartner C, Blum MR, Browne JP, et al., and TRUST Study Group. Thyroid Hormone Therapy for Older Adults with Subclinical Hypothyroidism. N Engl J Med. 2017; 376:2534–44. https://doi.org/10.1056/NEJMoa1603825 [PubMed]
- 37. Goncharova ND, Ivanova LG, Oganyan TE, Vengerin AA. Features of endocrine function of the pancreas with aging in nonhuman primates with various types of adaptive behavior. Advances in Gerontology. 2019; 9:389–95. https://doi.org/10.1134/S2079057019040076
- 38. Goncharova ND, Marenin VY, Oganyan TE. Aging of the hypothalamic-pituitary-adrenal axis in nonhuman primates with depression-like and aggressive behavior. Aging (Albany NY). 2010; 2:854–66. https://doi.org/10.18632/aging.100227 [PubMed]
- 39. Belvederi Murri M, Pariante C, Mondelli V, Masotti M, Atti AR, Mellacqua Z, Antonioli M, Ghio L, Menchetti M, Zanetidou S, Innamorati M, Amore M. HPA axis and aging in depression: systematic review and meta-analysis. Psychoneuroendocrinology. 2014; 41:46–62. https://doi.org/10.1016/j.psyneuen.2013.12.004 [PubMed]
- 40. Goncharova N, Bowden D, Johnson E. Editorial: The HPA axis and aging: individual features, age-related pathology. Front Endocrinol (Lausanne). 2023; 14:1222033. https://doi.org/10.3389/fendo.2023.1222033 [PubMed]
- 41. Goncharova N, Chigarova O, Oganyan T. Age-related and individual features of the HPA axis stress responsiveness under constant light in nonhuman primates. Front Endocrinol (Lausanne). 2023; 13:1051882. https://doi.org/10.3389/fendo.2022.1051882 [PubMed]
- 42. Konstandi M, Johnson EO. Age-related modifications in CYP-dependent drug metabolism: role of stress. Front Endocrinol (Lausanne). 2023; 14:1143835. https://doi.org/10.3389/fendo.2023.1143835 [PubMed]
- 43. Degroote C, von Känel R, Thomas L, Zuccarella-Hackl C, Messerli-Bürgy N, Saner H, Wiest R, Wirtz PH. Lower diurnal HPA-axis activity in male hypertensive and coronary heart disease patients predicts future CHD risk. Front Endocrinol (Lausanne). 2023; 14:1080938. https://doi.org/10.3389/fendo.2023.1080938 [PubMed]
- 44. Lai JC, Lee DY. Network cultivation, diurnal cortisol and biological ageing: The rejuvenation hypothesis. Med Hypotheses. 2019; 122:1–4. https://doi.org/10.1016/j.mehy.2018.09.041 [PubMed]
- 45. Yiallouris A, Tsioutis C, Agapidaki E, Zafeiri M, Agouridis AP, Ntourakis D, Johnson EO. Adrenal Aging and Its Implications on Stress Responsiveness in Humans. Front Endocrinol (Lausanne). 2019; 10:54. https://doi.org/10.3389/fendo.2019.00054 [PubMed]
- 46. Lopez M, Seidl A, Phillips KA. Cortisol levels across the lifespan in common marmosets (Callithrix jacchus). Am J Primatol. 2024; 86:e23597. https://doi.org/10.1002/ajp.23597 [PubMed]
- 47. Fink LH, Anastasio NC, Fox RG, Rice KC, Moeller FG, Cunningham KA. Individual Differences in Impulsive Action Reflect Variation in the Cortical Serotonin 5-HT2A Receptor System. Neuropsychopharmacology. 2015; 40:1957–68. https://doi.org/10.1038/npp.2015.46 [PubMed]
- 48. Balsevich G, Baumann V, Uribe A, Chen A, Schmidt MV. Prenatal Exposure to Maternal Obesity Alters Anxiety and Stress Coping Behaviors in Aged Mice. Neuroendocrinology. 2016; 103:354–68. https://doi.org/10.1159/000439087 [PubMed]
- 49. Colman RJ. Non-human primates as a model for aging. Biochim Biophys Acta Mol Basis Dis. 2018; 1864:2733–41. https://doi.org/10.1016/j.bbadis.2017.07.008 [PubMed]
- 50. Havel PJ, Kievit P, Comuzzie AG, Bremer AA. Use and Importance of Nonhuman Primates in Metabolic Disease Research: Current State of the Field. ILAR J. 2017; 58:251–68. https://doi.org/10.1093/ilar/ilx031 [PubMed]
- 51. Mikheenko Y, Shiba Y, Sawiak S, Braesicke K, Cockcroft G, Clarke H, Roberts AC. Serotonergic, brain volume and attentional correlates of trait anxiety in primates. Neuropsychopharmacology. 2015; 40:1395–404. https://doi.org/10.1038/npp.2014.324 [PubMed]
- 52. Schneider ML, Moore CF, Ahlers EO, Barnhart TE, Christian BT, DeJesus OT, Engle JW, Holden JE, Larson JA, Moirano JM, Murali D, Nickles RJ, Resch LM, Converse AK. PET Measures of D1, D2, and DAT Binding Are Associated With Heightened Tactile Responsivity in Rhesus Macaques: Implications for Sensory Processing Disorder. Front Integr Neurosci. 2019; 13:29. https://doi.org/10.3389/fnint.2019.00029 [PubMed]
- 53. Qin D, Rizak J, Chu X, Li Z, Yang S, Lü L, Yang L, Yang Q, Yang B, Pan L, Yin Y, Chen L, Feng X, Hu X. A spontaneous depressive pattern in adult female rhesus macaques. Sci Rep. 2015; 5:11267. https://doi.org/10.1038/srep11267 [PubMed]
- 54. Epelbaum J, Terrien J. Mini-review: Aging of the neuroendocrine system: Insights from nonhuman primate models. Prog Neuropsychopharmacol Biol Psychiatry. 2020; 100:109854. https://doi.org/10.1016/j.pnpbp.2019.109854 [PubMed]
- 55. Kalin NH, Shelton SE. Nonhuman primate models to study anxiety, emotion regulation, and psychopathology. Ann N Y Acad Sci. 2003; 1008:189–200. https://doi.org/10.1196/annals.1301.021 [PubMed]
- 56. Verdier JM, Acquatella I, Lautier C, Devau G, Trouche S, Lasbleiz C, Mestre-Francés N. Lessons from the analysis of nonhuman primates for understanding human aging and neurodegenerative diseases. Front Neurosci. 2015; 9:64. https://doi.org/10.3389/fnins.2015.00064 [PubMed]
- 57. Marvanová M, Ménager J, Bezard E, Bontrop RE, Pradier L, Wong G. Microarray analysis of nonhuman primates: validation of experimental models in neurological disorders. FASEB J. 2003; 17:929–31. https://doi.org/10.1096/fj.02-0681fje [PubMed]
- 58. Anderson TJ, Watowich MM, Chiou KL, Goldman EA, Peterson S, Anderson JA, Snyder-Mackler N, Carbone L, Kohama SG, Sterner KN. Patterns of transcriptomic aging in the hippocampus of rhesus macaques highlight midlife transitions. Geroscience. 2026; 48:2913–29. https://doi.org/10.1007/s11357-025-01834-z [PubMed]
- 59. Van Someren EJ, Riemersma-Van Der Lek RF. Live to the rhythm, slave to the rhythm. Sleep Med Rev. 2007; 11:465–84. https://doi.org/10.1016/j.smrv.2007.07.003 [PubMed]
- 60. Linden JB, Capitanio JP, McCowan B, Isbell LA. Coping style and cortisol levels in infancy predict hair cortisol following new group formation in captive rhesus macaques (Macaca mulatta). Am J Primatol. 2018; 80:e22938. https://doi.org/10.1002/ajp.22938 [PubMed]
- 61. Zijlmans DG, Meijer L, Vernes MK, Wubben JA, Hofman L, Louwerse AL, Sterck EH, Langermans JA, Stammes MA. Effect of Housing Conditions on Cortisol and Body Fat Levels in Female Rhesus Macaques. Biology (Basel). 2021; 10:744. https://doi.org/10.3390/biology10080744 [PubMed]
- 62. Richer MJ, Grenier S, Lupien S, Plusquellec P. Increasing stress resilience in older adults through a 6-week prevention program: effects on coping strategies, anxiety symptoms, and cortisol levels. Front Psychol. 2025; 15:1499609. https://doi.org/10.3389/fpsyg.2024.1499609 [PubMed]
- 63. Maestripieri D, Georgiev AV. What cortisol can tell us about the costs of sociality and reproduction among free-ranging rhesus macaque females on Cayo Santiago. Am J Primatol. 2016; 78:92–105. https://doi.org/10.1002/ajp.22368 [PubMed]
- 64. Suomi SJ. Risk, resilience, and gene x environment interactions in rhesus monkeys. Ann N Y Acad Sci. 2006; 1094:52–62. https://doi.org/10.1196/annals.1376.006 [PubMed]
- 65. Lupien SJ, Juster RP, Raymond C, Marin MF. The effects of chronic stress on the human brain: From neurotoxicity, to vulnerability, to opportunity. Front Neuroendocrinol. 2018; 49:91–105. https://doi.org/10.1016/j.yfrne.2018.02.001 [PubMed]
- 66. de Kloet ER, Joëls M. The cortisol switch between vulnerability and resilience. Mol Psychiatry. 2024; 29:20–34. https://doi.org/10.1038/s41380-022-01934-8 [PubMed]
- 67. Ancelin ML, Scali J, Norton J, Ritchie K, Dupuy AM, Chaudieu I, Ryan J. Heterogeneity in HPA axis dysregulation and serotonergic vulnerability to depression. Psychoneuroendocrinology. 2017; 77:90–4. https://doi.org/10.1016/j.psyneuen.2016.11.016 [PubMed]
- 68. Grebe NM, Del Giudice M, Emery Thompson M, Nickels N, Ponzi D, Zilioli S, Maestripieri D, Gangestad SW. Testosterone, cortisol, and status-striving personality features: A review and empirical evaluation of the Dual Hormone hypothesis. Horm Behav. 2019; 109:25–37. https://doi.org/10.1016/j.yhbeh.2019.01.006 [PubMed]
- 69. Retana-Márquez S, Bonilla-Jaime H, Vázquez-Palacios G, Martínez-García R, Velázquez-Moctezuma J. Changes in masculine sexual behavior, corticosterone and testosterone in response to acute and chronic stress in male rats. Horm Behav. 2003; 44:327–37. https://doi.org/10.1016/j.yhbeh.2003.04.001 [PubMed]
- 70. Rao R, Androulakis IP. The physiological significance of the circadian dynamics of the HPA axis: Interplay between circadian rhythms, allostasis and stress resilience. Horm Behav. 2019; 110:77–89. https://doi.org/10.1016/j.yhbeh.2019.02.018 [PubMed]
- 71. Nicolaides NC, Charmandari E, Kino T, Chrousos GP. Stress-Related and Circadian Secretion and Target Tissue Actions of Glucocorticoids: Impact on Health. Front Endocrinol (Lausanne). 2017; 8:70. https://doi.org/10.3389/fendo.2017.00070 [PubMed]
- 72. Goncharova ND, Oganyan TE, Smelkova SA. Effect of aging on stress reactivity of the adrenal cortex in laboratory primates. Dependence on the time of day. Bull Exp Biol Med. 2006; 141:368–71. https://doi.org/10.1007/s10517-006-0173-5 [PubMed]
- 73. Hood S, Amir S. The aging clock: circadian rhythms and later life. J Clin Invest. 2017; 127:437–46. https://doi.org/10.1172/JCI90328 [PubMed]
- 74. Mattis J, Sehgal A. Circadian Rhythms, Sleep, and Disorders of Aging. Trends Endocrinol Metab. 2016; 27:192–203. https://doi.org/10.1016/j.tem.2016.02.003 [PubMed]
- 75. Berardelli R, Karamouzis I, D'Angelo V, Zichi C, Fussotto B, Giordano R, Ghigo E, Arvat E. Role of mineralocorticoid receptors on the hypothalamus-pituitary-adrenal axis in humans. Endocrine. 2013; 43:51–8. https://doi.org/10.1007/s12020-012-9750-8 [PubMed]
- 76. Gaffey AE, Bergeman CS, Clark LA, Wirth MM. Aging and the HPA axis: Stress and resilience in older adults. Neurosci Biobehav Rev. 2016; 68:928–45. https://doi.org/10.1016/j.neubiorev.2016.05.036 [PubMed]
- 77. Gupta D, Morley JE. Hypothalamic-pituitary-adrenal (HPA) axis and aging. Compr Physiol. 2014; 4:1495–510. https://doi.org/10.1002/cphy.c130049 [PubMed]
- 78. Goncharova ND, Shmaliy AV, Marenin VY, Smelkova SA, Lapin BA. Circadian and age-related changes in stress responsiveness of the adrenal cortex and erythrocyte antioxidant enzymes in female rhesus monkeys. J Med Primatol. 2008; 37:229–38. https://doi.org/10.1111/j.1600-0684.2007.00278.x [PubMed]
- 79. Goncharova ND. (2019). Aging, stress responsiveness of neuroendocrine systems and insulin resistance in nonhuman primates with depression-like and anxiety-like phenotype. In: Productive Longevity: Problems and Forecasts. Proceedings of the All-Russian Forum with International Participation, Zhdanov RI, Mamaev VV. eds. (Moscow: OOO ”Astra-Poligrafiya”), pp. 19–21.
- 80. Goncharova ND, Oganyan TE, Marenin VY, Vengerin AA. Age-specific and individual features of vasopressinergic regulation of the hypothalamic-pituitary-adrenal system in primates. Bull Exp Biol Med. 2015; 158:804–6. https://doi.org/10.1007/s10517-015-2866-0 [PubMed]
- 81. Čukuranović-Kokoris J, Kundalić B, Pavlović M, Ugrenović S. Pituitary cells in man during aging: An immunohistological and morphometric study. J Med Biochem. 2025; 44:203–10. https://doi.org/10.5937/jomb0-54605 [PubMed]
- 82. Goncharova ND, Chigarova OA, Oganyan TE. Effect of Vasopressin V1b Receptor Blockade on Activity of the Hypothalamic-Pituitary-Adrenal Axis in Old Monkeys with Depression-Like and Anxious Behavior Subjected to Stress or Injected with Vasopressin. Bull Exp Biol Med. 2018; 166:86–91. https://doi.org/10.1007/s10517-018-4294-4 [PubMed]
- 83. Oster H, Challet E, Ott V, Arvat E, de Kloet ER, Dijk DJ, Lightman S, Vgontzas A, Van Cauter E. The Functional and Clinical Significance of the 24-Hour Rhythm of Circulating Glucocorticoids. Endocr Rev. 2017; 38:3–45. https://doi.org/10.1210/er.2015-1080 [PubMed]
- 84. Wingenfeld K, Otte C. Mineralocorticoid receptor function and cognition in health and disease. Psychoneuroendocrinology. 2019; 105:25–35. https://doi.org/10.1016/j.psyneuen.2018.09.010 [PubMed]
- 85. Faye C, Mcgowan JC, Denny CA, David DJ. Neurobiological Mechanisms of Stress Resilience and Implications for the Aged Population. Curr Neuropharmacol. 2018; 16:234–70. https://doi.org/10.2174/1570159X15666170818095105 [PubMed]
- 86. Keller J, Gomez R, Williams G, Lembke A, Lazzeroni L, Murphy GM
Jr , Schatzberg AF. HPA axis in major depression: cortisol, clinical symptomatology and genetic variation predict cognition. Mol Psychiatry. 2017; 22:527–36. https://doi.org/10.1038/mp.2016.120 [PubMed] - 87. Otte C, Wingenfeld K, Kuehl LK, Kaczmarczyk M, Richter S, Quante A, Regen F, Bajbouj M, Zimmermann-Viehoff F, Wiedemann K, Hinkelmann K. Mineralocorticoid receptor stimulation improves cognitive function and decreases cortisol secretion in depressed patients and healthy individuals. Neuropsychopharmacology. 2015; 40:386–93. https://doi.org/10.1038/npp.2014.181 [PubMed]
- 88. Gjerstad JK, Lightman SL, Spiga F. Role of glucocorticoid negative feedback in the regulation of HPA axis pulsatility. Stress. 2018; 21:403–16. https://doi.org/10.1080/10253890.2018.1470238 [PubMed]
- 89. Miller AL, Lumeng JC. Pathways of Association from Stress to Obesity in Early Childhood. Obesity (Silver Spring). 2018; 26:1117–24. https://doi.org/10.1002/oby.22155 [PubMed]
- 90. Gugula A, Sambak P, Trenk A, Drabik S, Nogaj A, Soltys Z, Gundlach AL, Blasiak A. Early-life adversity alters adult nucleus incertus neurons: implications for neuronal mechanisms of increased stress and compulsive behavior vulnerability. Neuropsychopharmacology. 2025; 50:1406–19. https://doi.org/10.1038/s41386-025-02089-0 [PubMed]
- 91. Vazquez CR, Becker LJ, Kuo CC, Cariello SA, Hamdan AN, Al-Hasani R, Maloney SE, McCall JG. Maternal separation disrupts noradrenergic control of adult coping behaviors. Neuropsychopharmacology. 2025; 50:1795–806. https://doi.org/10.1038/s41386-025-02201-4 [PubMed]
- 92. Law AJ, Pei Q, Walker M, Gordon-Andrews H, Weickert CS, Feldon J, Pryce CR, Harrison PJ. Early parental deprivation in the marmoset monkey produces long-term changes in hippocampal expression of genes involved in synaptic plasticity and implicated in mood disorder. Neuropsychopharmacology. 2009; 34:1381–94. https://doi.org/10.1038/npp.2008.106 [PubMed]
- 93. Wakeford AG, Nye JA, Morin EL, Mun J, Meyer JS, Goodman M, Howell LL, Sanchez MM. Alterations in adolescent brain serotonin (5HT)1A, 5HT2A, and dopamine (D)2 receptor systems in a nonhuman primate model of early life adversity. Neuropsychopharmacology. 2024; 49:1227–35. https://doi.org/10.1038/s41386-023-01784-0 [PubMed]
- 94. Goncharova ND. Individual life history, behavior, and biomarkers of ageing. JNHA. 2009; 13:S215. https://doi.org/10.1007/s12603-009-0095-9
- 95. van den Beld AW, Kaufman JM, Zillikens MC, Lamberts SW, Egan JM, van der Lely AJ. The physiology of endocrine systems with ageing. Lancet Diabetes Endocrinol. 2018; 6:647–58. https://doi.org/10.1016/S2213-8587(18)30026-3 [PubMed]
- 96. Rakotoniaina JH, Kappeler PM, Kaesler E, Hämäläinen AM, Kirschbaum C, Kraus C. Hair cortisol concentrations correlate negatively with survival in a wild primate population. BMC Ecol. 2017; 17:30. https://doi.org/10.1186/s12898-017-0140-1 [PubMed]
- 97. Goncharova ND, Chigarova OA, Ermolaeva AM, Oganyan TE, Timoshenko NV, Karaseva DM. Neuroendocrine systems during aging under constant lighting. In: Proceedings of the International Conference: New technologies in medicine, biology, pharmacology and ecology NT + M`25, Gurzuf. 2025; 331–5. https://doi.org/10.47501/978-5-6044060-5-2.331-335
- 98. Philippe J, Dibner C. Thyroid circadian timing: roles in physiology and thyroid malignancies. J Biol Rhythms. 2015; 30:76–83. https://doi.org/10.1177/0748730414557634 [PubMed]
- 99. Kalsbeek A, Fliers E. Daily regulation of hormone profiles. Handb Exp Pharmacol. 2013: 185–226. https://doi.org/10.1007/978-3-642-25950-0_8 [PubMed]
- 100. Roelfsema F, Pijl H, Kok P, Endert E, Fliers E, Biermasz NR, Pereira AM, Veldhuis JD. Thyrotropin secretion in healthy subjects is robust and independent of age and gender, and only weakly dependent on body mass index. J Clin Endocrinol Metab. 2014; 99:570–8. https://doi.org/10.1210/jc.2013-2858 [PubMed]
- 101. Veldhuis JD. Changes in pituitary function with ageing and implications for patient care. Nat Rev Endocrinol. 2013; 9:205–15. https://doi.org/10.1038/nrendo.2013.38 [PubMed]
- 102. Roth GS, Handy AM, Mattison JA, Tilmont EM, Ingram DK, Lane MA. Effects of dietary caloric restriction and aging on thyroid hormones of rhesus monkeys. Horm Metab Res. 2002; 34:378–82. https://doi.org/10.1055/s-2002-33469 [PubMed]
- 103. Yoshida T, Sato M, Ohtoh K, Cho F, Honjo S. Effects of aging on the in vivo release of thyrotropin (TSH), triiodothyronine, and thyroxine induced by TSH-releasing hormone in the cynomolgus monkey (Macaca fascicularis). Endocrinology. 1989; 124:1287–93. https://doi.org/10.1210/endo-124-3-1287 [PubMed]
- 104. Lair S, Crawshaw GJ, Mehren KG, Perrone MA. Diagnosis of hypothyroidism in a western lowland gorilla (Gorilla gorilla gorilla) using human thyroid-stimulating hormone assay. J Zoo Wildl Med. 1999; 30:537–40. [PubMed]
- 105. Ozpinar A, Golub MS, Poppenga RH, Blount BC, Gillespie JR. Thyroid status of female rhesus monkeys and preliminary information on impact of perchlorate administration. Lab Anim. 2011; 45:209–14. https://doi.org/10.1258/la.2011.010047 [PubMed]
- 106. Sciurba JD, Hayes JM, Nouraie SM, Wilson HM, Fortman JD, Halliday LC. Reference Intervals for Total T4 and Free T4 in Cynomolgus Macaques (Macaca fascicularis) and Rhesus Macaques (Macaca mulatta). J Am Assoc Lab Anim Sci. 2021; 60:380–7. https://doi.org/10.30802/AALAS-JAALAS-20-000126 [PubMed]
- 107. Waring AC, Arnold AM, Newman AB, Bùzková P, Hirsch C, Cappola AR. Longitudinal changes in thyroid function in the oldest old and survival: the cardiovascular health study all-stars study. J Clin Endocrinol Metab. 2012; 97:3944–50. https://doi.org/10.1210/jc.2012-2481 [PubMed]
- 108. Calsolaro V, Niccolai F, Pasqualetti G, Calabrese AM, Polini A, Okoye C, Magno S, Caraccio N, Monzani F. Overt and Subclinical Hypothyroidism in the Elderly: When to Treat? Front Endocrinol (Lausanne). 2019; 10:177. https://doi.org/10.3389/fendo.2019.00177 [PubMed]
- 109. Mele C, Mai S, Cena T, Pagano L, Scacchi M, Biondi B, Aimaretti G, Marzullo P. The pattern of TSH and fT4 levels across different BMI ranges in a large cohort of euthyroid patients with obesity. Front Endocrinol (Lausanne). 2022; 13:1029376. https://doi.org/10.3389/fendo.2022.1029376 [PubMed]
- 110. Bremner AP, Feddema P, Leedman PJ, Brown SJ, Beilby JP, Lim EM, Wilson SG, O'Leary PC, Walsh JP. Age-related changes in thyroid function: a longitudinal study of a community-based cohort. J Clin Endocrinol Metab. 2012; 97:1554–62. https://doi.org/10.1210/jc.2011-3020 [PubMed]
- 111. Chen J, Zhou W, Pan F, Cui W, Li M, Hu Y. Age-related change in thyroid-stimulating hormone: a cross-sectional study in healthy euthyroid population. Endocr J. 2018; 65:1075–82. https://doi.org/10.1507/endocrj.EJ18-0113 [PubMed]
- 112. Pasqualetti G, Tognini S, Polini A, Caraccio N, Monzani F. Is subclinical hypothyroidism a cardiovascular risk factor in the elderly? J Clin Endocrinol Metab. 2013; 98:2256–66. https://doi.org/10.1210/jc.2012-3818 [PubMed]
- 113. Mullur R, Liu YY, Brent GA. Thyroid hormone regulation of metabolism. Physiol Rev. 2014; 94:355–82. https://doi.org/10.1152/physrev.00030.2013 [PubMed]
- 114. Visser WE, Bombardieri CR, Zevenbergen C, Barnhoorn S, Ottaviani A, van der Pluijm I, Brandt R, Kaptein E, van Heerebeek R, van Toor H, Garinis GA, Peeters RP, Medici M, et al. Tissue-Specific Suppression of Thyroid Hormone Signaling in Various Mouse Models of Aging. PLoS One. 2016; 11:e0149941. https://doi.org/10.1371/journal.pone.0149941 [PubMed]
- 115. Chaker L, Cappola AR, Mooijaart SP, Peeters RP. Clinical aspects of thyroid function during ageing. Lancet Diabetes Endocrinol. 2018; 6:733–42. https://doi.org/10.1016/S2213-8587(18)30028-7 [PubMed]
- 116. Goncharova ND, Ermolaeva AM, Chigarova OA, Oganyan TE, Ivanova LG, Timoshenko NV. Individual Features of the Hypothalamic-Pituitary-Thyroid Axis Functioning during Aging in Non-Human Primates. Bull Exp Biol Med. 2023; 175:497–502. https://doi.org/10.1007/s10517-023-05894-z [PubMed]
- 117. Wang D, Yu S, Ma C, Li H, Qiu L, Cheng X, Guo X, Yin Y, Li D, Wang Z, Hu Y, Lu S, Yang G, Liu H. Reference intervals for thyroid-stimulating hormone, free thyroxine, and free triiodothyronine in elderly Chinese persons. Clin Chem Lab Med. 2019; 57:1044–52. https://doi.org/10.1515/cclm-2018-1099 [PubMed]
- 118. Rakov H, De Angelis M, Renko K, Hönes GS, Zwanziger D, Moeller LC, Schramm KW, Führer D. Aging Is Associated with Low Thyroid State and Organ-Specific Sensitivity to Thyroxine. Thyroid. 2019; 29:1723–33. https://doi.org/10.1089/thy.2018.0377 [PubMed]
- 119. Jonklaas J, Razvi S. Reference intervals in the diagnosis of thyroid dysfunction: treating patients not numbers. Lancet Diabetes Endocrinol. 2019; 7:473–83. https://doi.org/10.1016/S2213-8587(18)30371-1 [PubMed]
- 120. Xing D, Liu D, Li R, Zhou Q, Xu J. Factors influencing the reference interval of thyroid-stimulating hormone in healthy adults: A systematic review and meta-analysis. Clin Endocrinol (Oxf). 2021; 95:378–89. https://doi.org/10.1111/cen.14454 [PubMed]
- 121. Fliers E, Alkemade A, Wiersinga WM, Swaab DF. Hypothalamic thyroid hormone feedback in health and disease. Prog Brain Res. 2006; 153:189–207. https://doi.org/10.1016/S0079-6123(06)53011-0 [PubMed]
- 122. Biondi B. Subclinical Hypothyroidism in Patients with Obesity and Metabolic Syndrome: A Narrative Review. Nutrients. 2023; 16:87. https://doi.org/10.3390/nu16010087 [PubMed]
- 123. Li Y, Chai Y, Liu X, Wang X, Meng X, Tang M, Zhang L, Zhang H. The non-high-density lipoprotein cholesterol to high-density lipoprotein cholesterol ratio (NHHR) is associated with thyroid hormones and thyroid hormone sensitivity indices: a cross-sectional study. Lipids Health Dis. 2024; 23:310. https://doi.org/10.1186/s12944-024-02292-w [PubMed]
- 124. Patrizio A, Ferrari SM, Elia G, Ragusa F, Balestri E, Botrini C, Rugani L, Mazzi V, Antonelli A, Fallahi P, Benvenga S. Hypothyroidism and metabolic cardiovascular disease. Front Endocrinol (Lausanne). 2024; 15:1408684. https://doi.org/10.3389/fendo.2024.1408684 [PubMed]
- 125. Kim HK, Song J. Hypothyroidism and Diabetes-Related Dementia: Focused on Neuronal Dysfunction, Insulin Resistance, and Dyslipidemia. Int J Mol Sci. 2022; 23:2982. https://doi.org/10.3390/ijms23062982 [PubMed]
- 126. Goncharova ND, Yu Marenin V, Vengerin AA. Age-related changes in the reliability of antioxidant enzyme defense in monkeys with different types of adaptive behavior. Curr Aging Sci. 2013; 6:163–9. https://doi.org/10.2174/18746098112059990004 [PubMed]
- 127. Ashcroft FM, Rorsman P. Diabetes mellitus and the β cell: the last ten years. Cell. 2012 Mar 16;148:1160–71. https://doi.org/10.1016/j.cell.2012.02.010 [PubMed]
- 128. Saisho Y, Butler AE, Manesso E, Elashoff D, Rizza RA, Butler PC. β-cell mass and turnover in humans: effects of obesity and aging. Diabetes Care. 2013; 36:111–7. https://doi.org/10.2337/dc12-0421 [PubMed]
- 129. Chang CH, Yeh YC, Shih SR, Lin JW, Chuang LM, Caffrey JL, Tu YK. Association between thyroid dysfunction and dysglycaemia: a prospective cohort study. Diabet Med. 2017; 34:1584–90. https://doi.org/10.1111/dme.13420 [PubMed]
- 130. Roa Dueñas OH, Van der Burgh AC, Ittermann T, Ligthart S, Ikram MA, Peeters R, Chaker L. Thyroid Function and the Risk of Prediabetes and Type 2 Diabetes. J Clin Endocrinol Metab. 2022; 107:1789–98. https://doi.org/10.1210/clinem/dgac006 [PubMed]
- 131. Khan SH, Fazal N, Ijaz A, Manzoor SM, Asif N, Rafi T, Yasir M, Niazi NK. Insulin Resistance and Glucose Levels in Subjects with Subclinical Hypothyroidism. J Coll Physicians Surg Pak. 2017; 27:329–33. [PubMed]
- 132. Fang X, Hu R, Fei S, Ding Z, Zhao J, Shang J. Associations between cardiovascular health and low thyroid function among US adults: a population-based study. Front Endocrinol (Lausanne). 2024; 15:1437386. https://doi.org/10.3389/fendo.2024.1437386 [PubMed]
- 133. Santini F, Marzullo P, Rotondi M, Ceccarini G, Pagano L, Ippolito S, Chiovato L, Biondi B. Mechanisms in endocrinology: the crosstalk between thyroid gland and adipose tissue: signal integration in health and disease. Eur J Endocrinol. 2014; 171:R137–52. https://doi.org/10.1530/EJE-14-0067 [PubMed]