Abstract
To understand the role of mitochondrial uncoupling protein (UCP) in regulating insulin signaling and glucose homeostasis, we created transgenicDrosophila lines with targeted UCP expression in insulin producing cells (IPCs). Increased UCP activity in IPCs results in decreased steady state Ca2+ levels in IPCs as well as decreased PI3K activity and increased FoxO nuclear localization in periphery. This reduced systemic insulin signaling is accompanied by a mild hyperglycemia and extended life span. To test the hypothesis that ATP-sensitive potassium (KATP) channels may link changes in metabolic activity (e.g., glucose mediated ATP production or UCP-mediated ATP reduction) with insulin secretion, we characterized the effects of glucose and a specific KATP channel blocker, glibenclamide on membrane potential in adult IPCs. Exposure to glucose depolarizes membrane potential of IPCs and this effect is mimicked with glibenclamide, suggesting that KATP channels contribute to the mechanism whereby IPCs sense changes in circulating sugar. Further, as demonstrated in mammalian β-pancreatic cells, high glucose initiates a robust Ca2+ influx in adult IPCs. The presence of functional KATP channels in adult IPCs is further substantiated by in situ hybridization detecting the transcript for the sulfonylurea receptor (Sur) subunit of the KATP channel in those cells. Quantitative expression analysis demon-strates a reduction in transcripts for both Sur and the inward rectifying potassium channel (Kir) subunits when IPCs are partially ablated. In summary, we have demonstrated a role for UCP in adult Drosophila IPCs in influencing systemic insulin signaling and longevity by a mechanism that may involve KATP channels.
Introduction
Mammalian mitochondrial uncoupling
proteins (UCPs) have been shown to be involved in energy metabolism, β-pancreatic cell function, and aging [1-6]. Located
in the inner membrane of mitochondria, these carriers allow leakage of protons into the matrix,thereby disrupting the proton gradient generated by the respiratory
electron transport chain and effectively uncoupling substrate oxidation from
ATP phosphorylation. In β-pancreatic cells, insulin secretion depends upon
detection of changes in the ATP levels generated by mitochondrial oxidative
phosphorylation [7,8]. ed
levels of glucose in the β-pancreatic cells cause an increase in the ATP/ADP
ratio, leading to closure of the ATP-dependent potassium (KATP)
channel, plasma membrane depolarization, opening of the voltage-gated calcium
channel, calcium influx, and insulin secretion. Consistent with this paradigm,
elevated UCP2 activity in β-pancreatic
cells that should lead to a decrease in the ATP/ADP ratio has been shown to
have a negative effect on glucose-stimulated insulin secretion [2,4,9].
Additional studies including the finding that UCP2 activity is stimulated by
glucotoxicity and lipotoxicity in diabetic animal models have established a
crucial role for UCP2 in the regulation of insulin secretion and β-cell function [10].
Glucose homeostasis is maintained in a remarkably
conserved manner between mammals and fruit flies. Analogous to the
insulin-secreting β-pancreatic cells and glucagon-secreting pancreatic
islet a-cells that act in opposition to maintain glucose
homeostasis in mammals, fruit flies possess neurosecretory insulin-like
peptide-producing cells (IPCs) in the pars intercerebralis of the brain
and adipokinetic hormone (AKH)-producing corpora cardiaca (CC) cells that
function in glucose-sensing [11,12].
Genetic ablation of IPCs in the brain mimics a diabetic phenotype, with
increased sugar levels in larval and adult hemolymph associated with growth
retardation, developmental delay and reduced fecundity [12,13].
Conversely, targeted ablation of the AKH-producing CC cells renders larvae and
adults hypoglycemic [11,14]. SevenDrosophila insulin-like peptides (DILPs) have been identified with five
of them (DILPs 1-5) showing high homology with their mammalian counterparts [15] whereas a
single Drosophila AKH peptide has been documented [16,17]. While
genetic studies have indicated that adult IPCs are likely the primary endocrine
tissue responsible for DILP secretion and signaling, functional evidence as to
how adult IPCs sense circulating sugar is lacking and the underlying molecular
mechanism(s) responsible for glucose sensitivity are unknown.
In this study, we demonstrate that
targeted expression of exogenous UCP in adult IPCs results in attenuated
systemic insulin signaling, a mild hyperglycemia and a significant life span
extension. To test whether the mechanism for Drosophila insulin
secretion, like that found in the b-pancreatic cells, also involves KATP
channels, we show that adult IPCs respond to glucose and sulfonylurea KATP
channel blocker glibenclamide with membrane depolarization whereas adjacent
non-IPCs show no discernable response to either agent. Furthermore, we have
detected a robust Ca2+ influx in IPCs in response to glucose
exposure. These electrophysiological recordings are further supported by in
situ hybridization detecting transcripts for the sulfonylurea receptor
(SUR) subunit of the KATP channel in adult IPCs. Taken together, we
have provided strong evidence suggesting that the mechanism for the release of Drosophila
ILPs, as for insulin secretion in mammalian β-pancreatic cells,
involves KATP channels.
Results
UCP expression in adult
IPCs does not engender any measurable damage in these neurons
While Drosophila
IPCs have been shown to regulate DILPs action, the mechanism whereby this is
achieved is not understood [12]. We
hypothesized that similar molecular events regulating insulin secretion in
mammalian β-pancreatic cells may also be involved in glucose sensing and
secretion of DILPs in fruit flies. We therefore predicted that increased
mitochondrial uncoupling in IPCs should alter the intracellular events leading
to secretion of DILPs via modulation of intracellular ATP/ADP ratio [18]. Adult Drosophila
IPCs consist of a distinct cluster of 14 medial neurosecretory cells in the parsintercerebralis of the brain and can be readily visualized by targeted
GFP expression with an IPC-specific, dilp2 promoter (SI Figure 1A-1B) [12]. Using the
UAS-Gal4 system, we targeted two mammalian UCPs, mUCP1 and hUCP2, to fly IPCs.
The UAS-hucp2 construct has previously been demonstrated to increase
physiological mitochondrial uncoupling activity when targeted to the adult fly
nervous system [6]. Indirect
immunofluorescence studies with an anti-mUCP1 antibody confirmed targeted
expression of this protein specifically in the IPCs (SI Figure 1C-1D). We next
confirmed that constitutive UCP expression in IPCs did not cause death or
discernable damage to the IPCs. IPC neurons from flies coexpressing mUCP1 and
GFP under the control of the dilp2-Gal4 driver were morphologically
indistinguishable from those of control flies expressing only GFP in the IPCs
(SI Figure 1B, 1E1, and 1E2). An intact function of IPCs in these flies was
further substantiated by finding normal levels of mRNA expression in fly heads
for two of the three DILPs (DILP2 and DILP5) that are selectively expressed in
the IPCs (SI Figure 4).
Increased UCP activity
in adult IPCs modulates molecular events associated with insulin signaling pathways
We investigated whether
increased UCP activity in IPCs affects intracellular changes known to be
associated with mammalian insulin release. We began by measuring intracellular
Ca2+ flux in IPCs. In order to do this, we targeted a fluorescent Ca2+-binding
indicator "camgaroo" (Cg-2) to adult IPCs and tested initially whether these
cells responded to food intake with an influx of Ca2+as has been shown for larval Drosophila CCs in
culture and for mammalian β-pancreatic cells [7,11,19]. In
adult flies expressing Cg-2 in IPCs that have fasted on water alone, IPCs
showed a very low level of Cg-2 fluorescence (SI Figure 2B). Following
re-feeding with glucose or trehalose, two circulating insect sugars, a three-fold
increase in Ca2+-dependent fluorescence intensity was measured (SI
Figure 2A, 2C-2D). These results demonstrate the ability of adult IPCs to
sense extracellular nutrient conditions to increase intracellular Ca2+ concentration,
a key step in mammalian insulin release. Having established that nutrient
conditions increase Ca2+ concentration in adult IPCs in vivo, we next
asked whether increased UCP activity in these cells affects intracellular Ca2+ levels,
using the same Cg-2 reporter fly line. Under normal growth conditions, we
found a 21-51% decrease in steady-state Ca2+ levels in IPCs of adult
brains isolated from two different mUCP1, Cg-2 co-expressing fly lines as
compared to control flies (Figure 1). Thus, constitutive UCP expression in
IPCs results in a decreased steady state intracellular Ca2+ flux, a
physiological condition consistent with a lowered ATP/ADP ratio and a negative
regulation of insulin release.
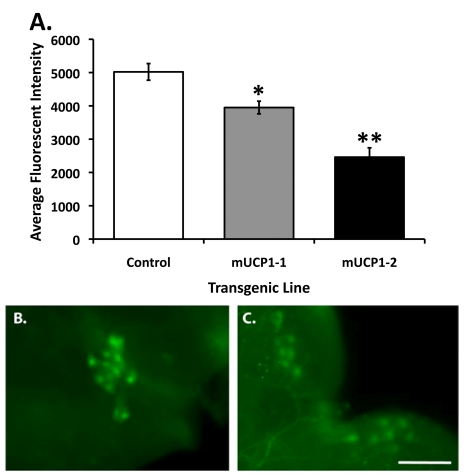
Figure 1. Constitutive UCP expression in adult IPCs results in decreased steady state intracellular Ca 2+ levels. (A) Under normal growth conditions, the Cg-2 protein in the
adult IPCs produces fluorescent signals reflecting a steady state Ca2+
flux whereas a 21-51% decrease in average fluorescent intensity is detected
in adult IPCs of two dilp2-Gal4/UAS-mucp1, UAS-cg-2 co-expressing
fly lines (mUCP1-1 and mUCP1-2), consistent with attenuated insulin
action. Each bar represents mean +
S.E.M. *p=0.0004, **p<0.0001
(Student's t-test). Shown are
average fluorescent signals from IPCs of 10 brains of each line in a
representative experiment (see Materials and Methods). Reproducible
results were obtained in three independent experiments. Representative
images of IPCs from a control adult brain isolated from a dilp2-Gal4/UAS-cg-2 fly line (B)
and an adult brain isolated from a dilp2-Gal4/UAS-mucp1, UAS-cg-2 fly line (C)
are shown. Images were taken with a 40X objective. Scale bar, 100 μm.
To understand how systemic
insulin signaling may be influenced by increased UCP activity in adult IPCs, we
measured the activity of PI-3' kinase, an integral part of the insulin
signaling cascade [20].
Responding to the activated insulin receptor, PI-3' kinase generates the second
messenger phosphatidylinositol-3,4,5-P3 which in turn interacts with the pleckstrin
homology (PH) domain found in several proteins involved in the PI-3' kinase
signaling and recruits them to the plasma membrane. Therefore, membrane
localization of PH containing molecules indicates increased PI-3' kinase
activity. A reporter construct consisting of PH-tagged GFP under the control
of the ubiquitous tubulin promoter (tGPH) enables monitoring of PI-3' kinase
activity downstream of insulin receptor activation [21]. Consistent
with the decreased Ca2+ levels measured in adult IPCs as the result of
increased UCP activity, we detected reduced membrane localization and increased
cytoplasmically retained tGPH reporter protein in the insulin-responsive
abdominal fat body of adult flies co-expressing tGPH and mUCP1 or hUCP2 under
normal feeding conditions (Figure 2B and 2C). This is in sharp contrast to a
predominant plasma membrane accumulation of the tGPH protein in fat body of
control flies (Figure 2A), suggesting that systemic insulin signaling is
attenuated when UCP is expressed in adult IPCs. Another functional measure of
peripheral insulin signaling is the sub-cellular localization of the Drosophila
homologue of the mammalian forkhead Box O (FoxO) transcription factor, known to
be involved in regulating the insulin signaling pathway [22]. During
normal insulin signaling FoxO is phosphorylated and found in the cytoplasm.
However, under conditions of decreased insulin signaling, FoxO remains unphosphorylated
and localizes predominantly to the nucleus [22,23]. Thus,
sub-cellular localization of FoxO reflects the status of insulin signaling in
the periphery. Immunohistochemical analysis of frozen sections of fly heads
with an anti-FoxO antibody [22] allowed us
to track the sub-cellular localization of dFoxO in the pericerebral fat body of
adult UCP-expressing flies. We found a significant increase in nuclear dFoxO
staining of pericerebral fat body cells isolated from UCP-expressing flies as
compared to controls (Figure 2D-2F). Taken together, evaluation of PI-3'
kinase activity and dFoxO sub-cellular localization, two independent methods
for assessing systemic insulin signaling, confirms that targeted UCP expression
in adult fly IPCs reduces systemic insulin signaling. Having established at
the cellular level attenuated insulin signaling, we next asked whether expression
of UCP in adult IPCs also regulates levels of circulating sugars. When
measuring both circulating glucose and trehalose levels in the hemolymph under
fasting conditions, we found an up to a 29% increase in circulating sugars in
adult flies expressing UCP in their adult IPCs, compared to control flies (Figure 2M). This finding demonstrates that at the whole animal level, UCP expression
in adult IPCs moderately disrupts glucose homeostasis likely as a consequence
of decreased secretion of some of the DILPs.
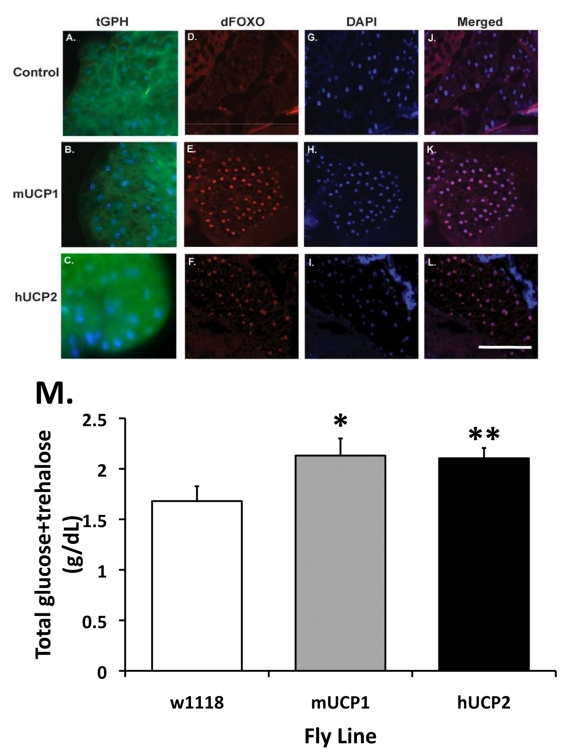
Figure 2. Normal and reduced systemic insulin signaling as reflected by the cellular localization of the PH-tagged GFP reporter protein (tGPH) and dFoxO in fat body cells. (A-C)
Increased UCP activities in the adult IPCs attenuate systemic insulin
signaling events. In control flies, under normal growth conditions and a
full strength PI-3' kinase activity, tGPH is predominantly located at the
plasma membrane of each fat body cell (A). (B-C) UCP
expression in adult IPCs results in a diffused, cytoplasmic distribution of
the tGPH protein. Control: dilp2-Gal4, tGPH; mUCP1: dilp2-Gal4/UAS-mucp1, tGPH; hUCP2: dilp2-Gal4/UAS-hucp2, tGPH. (D-F)
Increased accumulation of dFoxO in the nucleus of pericerebral fat body
cells in adult dilp2-Gal4/UAS-mucp1 and dilp2-Gal4/UAS-hucp2 flies indicates
reduced insulin signaling. Cryosections of adult heads were stained with
an α-dFoxO antibody
followed by Alexa 568-conjugated secondary antibodies. A strong nuclear
staining of the dFoxO protein was observed in the pericerebral fat body in
both dilp2-Gal4/UAS-mucp1 (mUCP1, Panel
E) and dilp2-Gal4/UAS-hucp2 (hUCP2, Panel
F) flies but not in dilp2-Gal4/w1118 (Control, Panel
D) flies. All sections were counter stained with DAPI (Panels G-I) to
locate the nucleus of each cell. Merged images of anti-FoxO staining and
DAPI are shown in Panels J-L. (M) Elevated levels of fasting
circulating sugars are measured in adult dilp2-Gal4/UAS-mucp1 and dilp2-Gal4/UAS-hucp2 flies. An
average of 29% increase in circulating sugars measured in 14-day-old dilp2-Gal4/UAS-mucp1 (mUCP1) and dilp2-Gal4/UAS-hucp2 (hUCP2) females
as compared to control dilp2-Gal4/w1118 (w1118)
females. Each bar represents mean +
SEM. N=5-7, *p= 0.046, **p=
0.05 (Student's t test).
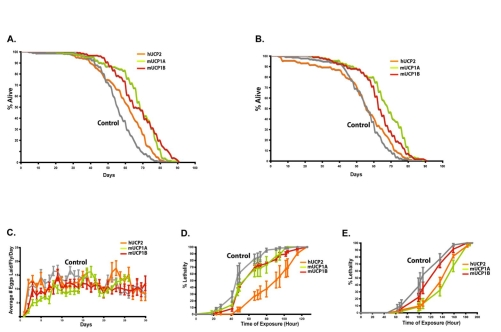
Figure 3. Targeted UCP expression in the adult IPCs extends life span and renders flies stress resistant with normal fecundity.
Survivorship curves for female
(A) and male (B) flies are shown. Two independent trials
were performed and results from one trial are shown for one hUCP2 and two
mUCP1 lines. Gray lines are control dilp2-Gal4/w1118 flies; two
mUCP1 expressing dilp2-Gal4/UAS-mucp1 lines, mUCP1A
and mUCP1B are green and red lines, respectively; and orange lines are dilp2-Gal4/UAS-hucp2. Panel A
(females) median life spans are 55 for controls and 71, 63, and 65 for
three UCP transgenic lines. Panel B (male) median life spans are 57 for
controls and 70, 59, and 59 for three UCP transgenic lines. Log rank
analysis shows an average of 19% increase in median life span in female
with targeted UCP expression in the adult IPCs and an average of 10%
increase in male (see SI Supplementary Table 1). (C) Female fecundity is similar
for flies expressing mUCP1 or hUCP2 in the adult IPCs and control flies.
Average number of eggs per day for 20 individual females was determined
from daily counts of eggs produced from single mated pairs [6]. (D)
Transgenic flies with targeted UCP expression in the adult IPCs are
resistant to oxidative stress. Survival during administration of paraquat
(20 mM paraquat in 5% sucrose) shows that 10-day-old flies expressing mUCP1
or hUCP2 in the IPCs are more resistant than controls. (E) mUCP1 or
hUCP2-expressing flies are resistant to starvation. Ten-day-old dilp2-Gal4/UAS-mucp1 and
dilp2-Gal4/UAS-hucp2 flies were
placed in vials containing water soaked filters and the number of dead
flies was counted at noted time intervals. Three independent assays were
performed and a representative experiment is shown for females. All values
are presented as mean ± S.E.M. Similar differences were seen for males and
females. Each experiment included 8-10 vials with 20 flies in each vial,
total of 160-200 flies for each condition.
Transgenic flies
expressing UCP in IPCs are long-lived and stress resistant with unique
alterations in dilp3 expression
By targeting UCP expression to the adult
IPCs, we have demonstrated a systemic reduction of insulin signaling
activities. Reduced insulin/IGF-1 signaling has been shown to be a conserved
mechanism for life span extension in multiple model
organisms [24]. To assess
the impact of our transgenic system on longevity, we performed survivorship
studies and found that expression of mUCP1 or hUCP2 in adult IPCs extended life
span. Using three different transgenic lines in two independent trials, we
observed an average increase in median life span in females of 19% (Figure 3A),
and of 10% in males as compared to genetically matched controls (Figure 3B)
(SI Supplementary Table 1). Significantly, this extended
longevity does not come with any measurable physiological costs. As shown in
Figure 3C, average egg laying activities were comparable between control and
UCP expressing flies, consistent with the view that endocrine manipulations for
life span extension are not necessarily associated with compromises in
reproduction [24]. In addition, we show that the
long-lived UCP transgenic flies are also resistant to oxidative stress (Figure 3D) and starvation (Figure 3E). Finally, we monitored the physical activity of
these transgenic flies and found comparable levels of spontaneous activity
between flies having constitutive UCP expression in their IPCs and control
flies (data not shown).
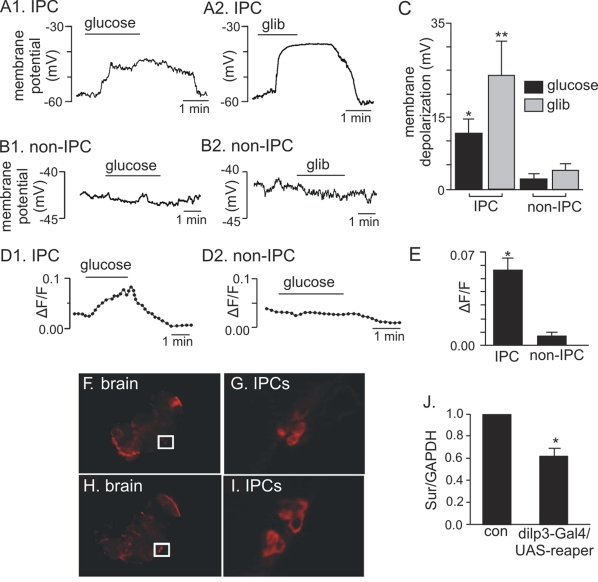
Figure 4. Electrophysiological, Ca 2+ influx, and expression evidence of functional KATP channels in adult IPCs. (A1-C)
Membrane depolarization of adult IPCs in response to glucose and
glibenclamide. (A1) Trace of membrane potential from an adult IPC
in the whole brain preparation shows that exposure to high glucose (80 mM)
evoked a reversible membrane depolarization. (A2) Trace of membrane
potential from an adult IPC shows that exposure to a commonly known KATP
channel blocker, glibenclamide (glib, 20 μM) also evoked a reversible
membrane depolarization. (B1 and B2) traces of membrane
potential from adult non-IPCs show that these cells do not respond to
glucose or glibenclamide. (C) Average membrane potential response
to glucose and glibenclamide of IPCs (N=5) and non-IPCs (N=3). Glucose (*)
and glibenclamide (**) significantly increased membrane potential of IPCs
as compared to non-IPCs, *p and **p <0.05 (Student's t test). Each
bar represents mean +
S.E.M. (D1-E) Glucose-dependent Ca2+
influx measured in adult IPCs. (D1) Normalized fluorescence trace (∆F/F) (see
Materials and Methods) recorded from an acutely dissociated adult IPC shows
that exposure to glucose (80 mM) increased fluorescence intensity, thus
indicating an increase in intracellular Ca2+. (D2)
Normalized fluorescence trace (∆F/F) recorded from an acutely
dissociated non-IPC shows that glucose does not increase intracellular Ca2+
in these cells. (E). An average of normalized fluorescence
intensity in response to glucose demonstrates a significant increase in Ca2+
influx recorded from IPCs (N=6) as compared to non-IPCs (N=3). *P= 0.007
(Student's t test). Each
bar represents mean +
S.E.M. (FI) In situ hybridization
of whole mount adult brains demonstrates dSur expression in
IPCs (F-G) when probed with anti-sense dSur probes and dilp2 expression (HI)
when probed with anti-sense dilp2 probes. The
IPCs marked in squares are shown in panel G for dSur signals and
panel I for dilp2
signals.
(J) Quantitative real-time PCR analysis reveals an average of 33%
reduction in dSur transcripts
when the IPCs are partially ablated using an IPC-specific driver, dilp3-Gal4 to drive the
expression of a pro-apoptotic gene, reaper. The
housekeeping gene GAPDH was used as the reference gene. Each bar
represents mean +
S.E.M. N=5, *p<0.001 (Student's t test).
Control: dilp3-Gal4/w1118.
To understand how energy
utilization in these flies was affected, we measured body composition of two
major energy stores: glycogen and lipid. We found an average of 26% increase
in glycogen storage in flies having UCP expression targeted to their IPCs (SI Figure 3A). On the other hand, in contrast to the previous report of a slight
elevation (10%) of lipid triglyceride content in animals with ablated IPCs [13], targeted
UCP expression in IPCs of adult flies does not result in a noticeable increase
in lipid storage (SI Figure 3B). UCP-induced reduction of insulin secretion,
as evidenced by a decrease in Ca2+ flux and
attenuation of insulin signaling in peripheral fat body, may also have an
effect on insulin production. Interestingly, when the transcript level of the
three DILPs found in the IPCs (DILP2, DILP3, and DILP5) was measured, we
detected a selective decrease in only dilp3 mRNA levels in flies
expressing mUCP1 or hUCP2 in IPCs whereas both the dilp2 and dilp5
mRNA level remained unchanged from control levels (SI Figure 4).
Electrophysiological and
expression evidence for KATP channels in adult Drosophila IPCs
The key component in regulating Ca2+ influx
into the mammalian β-pancreatic cells leading to insulin release is
membrane depolarization initiated by inhibition of KATP
channels [25,26].
Prompted by our studies demonstrating a decreased Ca2+ flux in
adult IPCs with increased UCP activity, we asked whether KATP
channels are involved in regulating insulin secretion in adult fly IPCs.
Whole cell currentclamp recordings from adult IPCs labeled with GFP in the whole brain
preparation showed that exposure to glucose depolarized membrane potential
from -69 ± 8 mV to -57 ± 9 mV (Figure 4A1). In the pars intercerebralis
of the fly brain, glucose-sensitivity appears to be a unique property of IPCs
since nearby non-IPCs did not respond to glucose (Figure 4B1). The effects of
glucose on IPCs were mimicked by the sulfonylurea glibenclamide with membrane
depolarization from -56 ± 7 mV to -30 ± 3 mV (Figure 4A2). Conversely,
non-IPCs did not respond to glibenclamide (Figure 4B2). As summarized in Figure 4C, a significant net membrane depolarization of 12 ± 4 mV and 23 ± 9 mV
specifically in adult IPCs in response to glucose and glibenclamide,
respectively was recorded suggesting that KATP channels contribute to
glucose sensing in these cells. In addition, preliminary voltage clamp
recordings from IPCs suggest that glibenclamide inhibited ~ 30% of total
outward current, further suggesting functional KATP
channels in IPCs (Mohammad Shahidullah, Yih-Woei Fridell, and Irwin B. Levitan,
unpublished results). To determine if exposure to glucose also initiates Ca2+ influx
into adult IPCs, we loaded acutely dissociated adult IPCs with a Ca2+-sensitive
fluorescent dye (rhod-3) and measured Ca2+ influx upon exposure to
glucose. Consistent with our electrophysiological results, glucose evoked a
significant Ca2+ influx into adult IPCs (Figure 4D1) whereas non-IPCs
did not show a Ca2+ response to glucose (Figure 4D2). Finally,
quantitative measure-ment demonstrated a large increase in Ca2+ influx
in response to glucose that was specific to brain IPCs (Figure 4E). To further
support our electrophysiological results indicating the presence of functional
sulfonylurea-sensitive KATP channels in adult IPCs, we investigated the expression
of the Sur subunit of the KATP channel in these cells. In situ hybridization
of whole mount adult brains reveals IPC-specific dSur expression with an
anti-sense dSur probe (Figure 4F-4G), but not with a sense dSur
probe (data not shown). In parallel experiments with an anti-sense dilp2
probe, neurons located in the same position in parsintercerebralis
stained positive for dilp2 transcript, confirming IPC-specific dSur
expression (Figure 4H-4I). Consistent with this result is the quantitative, real-time
expression analysis detecting an average reduction of 33% in dSur transcript
in the heads of adult flies having partially ablated IPCs [27] (Figure 4J). And finally, when the expression status of the other subunit of the KATP
channel, Kir was quantified in the heads of adult flies with partially ablated
IPCs, an average of 27% reduction in dKir transcript was seen
(SI Supplementary Figure 5) [28].
Discussion
In this report, we have
demonstrated that increased UCP activity in adult fly IPCs modulates systemic
insulin signaling as measured by both molecular events in the insulin signaling
pathway and glucose homeostasis at the organismal level. As expected,
attenuated insulin signaling measured in dilp2-Gal4/UAS-mucp1 anddilp2-Gal4/UAS-hucp2 flies is also associated with a significant
life span extension in females and a moderate increase in males [24]. In two
peripheral insulin responsive tissues, abdominal and pericerebral fat body, we
have demonstrated a reduction in the insulin signaling cascade that is
evidenced by decreased PI-3' kinase activity and increased nuclear accumulation
of the dFoxO transcription factor. Both events are associated with reduced
insulin signaling predicting decreased circulating DILPs. Consistent with this
notion we show that UCP expression in adult IPCs also leads to moderate fasting
hyperglycemia. While we do not currently know which of the three DILPs
produced in the IPCs may be affected in its secretion, we have shown that only
the mRNA expression of dilp3 is significantly reduced as a consequence
of increased UCP activity in adult IPCs. This is in marked contrast to
previous reports where a decrease specific to dilp2 transcripts resulted
from modulating insulin signaling through manipulation of the activity of JNK,
dFoxO, or Dmp53 [23, 29, 30].
Our findings suggest that not only does increased mitochondrial uncoupling in
adult IPCs attenuate insulin signaling by potentially decreasing DILPs
secretion, but that it may also be involved in a novel transcriptional
regulation of dilp3. Thus, modulation of mitochondrial uncoupling in
IPCs with our UCP transgenic model system has the potential to uncover novel
molecular targets controlled by the neuroendocrine axis that are involved in
energy metabolism, glucose homeostasis, and longevity.
A UCP2 activity in
mammalian β-pancreatic cells has been implicated in the development of
Type II diabetes by altering the ATP/ADP ratio, causing the KATPchannel to stay open and leading to decreased insulin
secretion [4, 5, 9, 31].
To further strengthen our transgenic system as a genetic model for diabetes, we
have taken electrophysiological approaches to understand the mechanism whereby
increased UCP activity modulates the release of DILPs in Drosophila
adult IPCs. Whole cell current clamp experiments with intact adult IPCs
recorded depolarized membrane potential in the presence of extracellular
glucose. Importantly, these effects were mimicked in the presence of the
sulfonylurea glibenclamide, a pharmacological blocker of KATPchannels. Under similar nutrient conditions, our
real-time Ca2+ imaging studies have demonstrated a significant Ca2+
influx. These studies strongly suggest that a conserved cascade of
intracellular events leading to β-cell insulin secretion, namely closure
of KATP channels and opening of voltage-gated calcium
channels, may also function in adult fly IPCs. A previous report has noted the
absence of expression of both Sur and Kir in the Drosophila
larval IPCs [11]. In that
report, the expression status of those two genes in the adult IPCs was not
determined [11]. In
addition to the strong electrophysiological data presented here, we have also
demonstrated an expression pattern for both Sur and Kir by in
situ hybridization and real-time quantitative RT-PCR that is consistent
with the notion of functional presence of KATPchannels in adult IPCs.
The Drosophila SUR subunit of the
KATP channel was first identified as a major component of
AKH secretion in cultured larval CC cells [11]. More
recently, in Drosophila heart, SUR has been shown to protect against
hypoxic stress, electrical pacing induced heart failure, and the flock house
virus [27, 32]. Here, in further support of the growing evidence
that KATP channels have evolved to maintain a homeostatic
function during both glucose sensing and infection resistance, we present
evidence for a potentially important role for KATP
channels in the secretion of the DILPs in adult fly IPCs. Complementing an
already significant body of knowledge on the opposing functions of Drosophila
IPCs and CC cells in maintaining glucose homeostasis in larval stages [11,12], we have
identified molecular events involved in the release of DILPs by adult IPCs,
further demonstrating a conserved mechanism for glucose sensing between fruit
flies and mammals.
Our studies have revealed
the physiological impact of increased mitochondrial uncoupling in the Drosophila
adult IPCs and the utility of such genetic manipulation to model metabolic
disorders such as type II diabetes. Because aging is one of the risk factors
for type II diabetes, it is vitally important to develop model systems to
understand the parameters involved in insulin regulation during adulthood. We
show that the genetically malleable model system of adult Drosophila is
well suited for the study of insulin regulation and it is our hope that a
better understanding of insulin secretion by adult fly IPCs will stimulate the
development of interventions in the fly that are likely to be relevant to human
disease.
Materials and Methods
Generation of mucp1
transgenic flies and double transgenic lines; maintenance of fly stocks.
The
full-length mouse ucp1 cDNA was reverse transcribed and amplified from the
mouse brown adipose tissue RNA (gift of Dr. Leslie Kozak, Pennington Biomedical
Research Center). Subsequent injection of the sequence-verified UAS-mucp1
construct resulted in several germ line transformants. Five independent
transgenic lines were then back-crossed to the w1118 stock
for 10 generations to achieve the same genetic background as in UAS-hucp2
flies [6]. Consistent
with our hUCP2 studies, two out of five independent UAS-mucp1 lines (UAS-mucp1A
and UAS-mucp1B) with the highest mitochondrial protein expression (data
not shown) also engender the most severe developmental lethality when driven by
the ubiquitous actin-Gal4 driver (data not shown) [6]. Except for
life span, stress resistance and egg laying studies where both transgenic lines
were examined, only UAS-mucp1A flies were included in the rest of the
studies described here.
Double transgenic flies
carrying UAS-mucp1, tGPH, or UAS-hucp2, tGPH insertions were
generated through meiotic recombination. Putative double transgenics selected
based on eye color were further confirmed by genomic PCR analysis to ensure the
presence of both inserts. Similar strategies were used to create dilp2-Gal4,
UAS-mucp1 flies. These flies were then crossed to UAS-cg-2 lines (gift
of Dr. R Davis, Baylor College of Medicine) for Ca2+
measurements. Despite our repeated attempts, no dilp2-Gal4, UAS-hucp2
flies were obtained. To partially ablate IPCs, the dilp3-Gal4 driver
was used to drive the expression of the pro-apoptotic gene, reaper in
IPCs.
The constitutive dilp2-Gal4
driver line was kindly provided by Dr. E. Rulifson (UCSF) and the tGPH line by
Dr. B. Edgar (Fred Hutchinson Cancer ResearchCenter, Seattle, WA). The dilp3-Gal4
driver line was obtained from Dr. M. Tatar (Brown University). The UAS-reaper
line (5824) was obtained from Bloomington Stock Center. All fly stocks were
maintained in a humidified, temperature-controlledincubator with
12h on/off light cycle at 25°C on standard corn meal/yeast/sucrose/agar diet [6].
Whole brain preparation
and electrophysiology.
Heads were collected from 10-day old dilp2-Gal4/UAS-GFP
flies and their brains isolated for digestion in 0.5% Trypsin in HBSS for 7
minutes at room temperature. Whole brains were stored on poly-L-lysine coated
cover slips in hemolymph-like (HL) solution [33] at room
temperature until individual cover slips were transferred into recording
chamber mounted on a fluorescence microscope (Zeiss Axioskop FS) and
continuously perfused with a buffer solution containing in mM: 3 KCl, 101 NaCl,
1 CaCl2, 4 MgCl2, 1.25 NaH2PO4, 20.7 NaHCO3, 35 sucrose, 5 glucose, bubbled with 5% CO2 balance
O2, pH 7.3. Individual IPCs were differentiated from
nearby non-IPCs by their bright GFP fluorescence; whole cell patch clamp
recordings were made in both cell types. High glucose (80 mM) media was made
iso-osmotic by eliminating sucrose and decreasing NaCl. Whole cell current
clamp recordings were made at room temperature using patch electrodes (4-7
mΩ) and Axopatch 200B amplifier (Molecular Devises). Internal pipette
solution contains in mM: 120 KCH3SO3, 4 NaCl, 1 MgCl2, 0.5 CaCl2, 10
HEPES, 10 EGTA, 0.3 GTP-Tris, pH 7.2. Membrane potential was recorded and
analyzed using a Digidata 1322A digitizer and pCLAMP 10 software. All chemicals
were purchased from Sigma.
Acute dissociation preparation and
real time Ca
2+ imaging.
To dissociate the cells, dilp2-Gal4/UAS-GFP
brains were treated as described for the whole brain preparation but incubated
in 0.5%Trypsin for 15 min. Trypsin was then inactivated by adding 5% FBS and
the tissue fragments were pelleted and resuspended in modified HL solution
containing in mM: 5 KCl, 108 NaCl, 2 CaCl2, 8.2 MgCl2, 1 NaH2PO4, 4 NaHCO3, 5
HEPES, 10 sucrose, 5 glucose, pH adjusted to 7.3 with NaOH. The tissue was
then titurated and plated on poly-L-lysine coated cover slips at a cell density
of ~ 15 brains per cover slip. Cells were allowed to settle for at least 2h at
room temperature before loading with the cell permeable Ca2+-dye
rhod-3 AM according to manufacturer's instructions (Molecular Probes, Cat. Nr.
R10145). Individual cover slips were submerged in a chamber mounted on a
fluorescence microscope (NIKON TE 200) and cells visualized with Nomarski
optics with a 40x oil immersion objective; fluorescently labeled IPCs and
non-fluorescent control cells were chosen for imaging.
NIS Elements 3.20 software
was used to acquire fluorescence images at 0.1 Hz with DsRed filter set (Chroma
42005). High glucose (80 mM) media was made iso-osmotic by eliminating sucrose
and decreasing NaCl. The response to glucose was normalized by dividing the
glucose-induced change in fluorescence (∆F=Fglucose-Fcontrol)
by control fluorescence (∆F/F). At the end of each experiment, cell
viability was assessed by responsiveness to high Ca2+ media
(data not shown).
Quantitative real-time
expression PCR analysis
. Total RNA was isolated from heads of 10-day
oldfemales
using the TRIzol method (Invitrogen), and subsequent cDNAand QPCR
experiments were performed as described previously [23]. Two-four
independent RNA preparations with triplicates in each QPCR experiment were used
to derive the mean ratios of target gene expression against the reference gene
GAPDH. The followingprimers were used: dilp2-F,
5'-AGCAAGCCTTTGTCCTTCATCTC-3';dilp2-R, 5'-ACACCATACTCGCACCTCGTTG-3'; dilp3-F,
5'-AGAGAACTTTGGACCCCGTGAA-3'; dilp3-R, 5'- TGAACCGAACTATCACTCAACAGTCT-3';
dilp5-F, 5'-GAGGCACCT TGGGCCTATTC-3'; dilp5-R, 5'-CATGTGGTGAGATTCGGAGCAA-3';
dSur-F, 5'-GAGCAGGCGACGACAAA-3', dSur-R,
5'-GCCCTC GTATCGCAGACTAAC-3';
dKir-F, 5'-CAGGACAA AGAGCACCAAGGAG-3',
dKir-R, 5'- CCAGATGA AGAACAAATCAGAGCC-3';
GAPDH-F, 5'-GAC GAAATCAAGGCTAAG GTCG-3'; GAPDH-R,5'-AATGGGTGTCGCTGAAGAAGTC-3'.
In situ
hybridization of adult IPCs.
In situ
hybridization of adult brains containing the IPCs was performed as previously
described (personal communication with Dr. E Ruflison, UCSF) [11].
To detect dSur expression, digoxigenin labeled dSur probes were generated by
subcloning the cDNA clone SD08664 (Open Biosystems, CA) into a pBluescript
vector. T3- and T7-mediated in vitro transcription was performed to
generate sense and anti-sense RNA probes. To use as a positive control,
IPC-specific dilp2 probes were generated in parallel reactions. The
indirect TSA System (PerkinElmer, Wellesley, PA) was used for signal
amplification.
Immunofluorescence
staining, immunohistochemistry, and fluorescence microscopy.
Whole mount fluores-cence experiments with dilp2-Gal4/UAS-mucp1,
UAS-GFP lines were performed to show the integrity of IPCs in the presence of
mUCP1 expression [23]. To detect
mUCP1 expression in the IPCs of the transgenic dilp2-Gal4/UAS-mucp1
flies, adult brains were dissected and fixed in 4% paraformaldehyde in PBS.
The immunofluorescence procedure with an anti-mUCP1 antibody (1:1000, UCP12-A,
Alpha Diagnostic) was performed as previously described [30]. For dFoxO
subcellular localization, 10-day old adult fly headswere fixed in
fresh 4% paraformaldehyde, embedded in tissue freezing medium (TFM, Triangle
Biomedical), frozen, cutat 10 μM and mounted on SuperFrost Plus slides (Fisher
Scientific). Slides were washed to remove TFM and stainedwith an
anti-dFoxO (1:500, rabbit antiserum) (kindly provided by Dr. O. Puig,
University of Helsinki, Finland) following the procedure described by Bauer,
et. al. [23]. All
imageswere taken using a Zeiss Axiovision Z1 fluorescentmicroscope with
identical magnification and exposure time for both control and experimental
samples (Thornwood, NY). For tGPH subcellular localization, adult abdominal
fat body tissues were dissectedinto PBS and visualized usingApoTome optics
(Zeiss Axiovision Z1 fluorescentmicroscope). Intra-cellular localization of the
fluorescent reporter tGPH in fat body cells under different treatments and
genetic backgrounds was scored by involving two people where one person
performed the staining and the other read the slides, which have been
numerically "blinded" by the first person.
Measuring steady-state Ca
2+
concentration in adult IPCs
. To measure the steady-state intracellular Ca2+
concentration in the adult IPCs with the Ca2+sensor Cg-2, flies were fasted 12-16 hours and then
placed in vials containing 10% glucose or 10% trehalose soaked filters for 30
minutes. Fly heads were then collected and fixed in 4% formaldehyde/PBS, and
brains containing IPCs were dissected for fluorescent measurements. Ca2+-dependent,
fluorescent images with identical magni-fication and exposure time for both
control and experimental samples were collected by using Axio-vision Apotome
microscopy equipped with a CCD camera and quantification of signals was
achieved using the Axiovision softwaresuite, Version 4.5 (Zeiss, Inc.). Arbitrary
fluorescent units representing the signal intensity were both generated and
analyzed using the Axiovision software [23].
Typically, 8-10 brains/treatment were included in each
experiment. Three independent experiments were performed. Average signals
under each treatment were achieved by quantifying signal intensity of each IPC
before calculating the average signal intensity (~112-140 cells/treatment).
Hemolymph collection and
carbohydrate measurements.
Adult hemolymph was extracted by capillary action
after a small puncture to the head capsule near the ocelli. Flies were reared on standard corn meal/ yeast/ sucrose/agar diet and
fasted for 12 hours on 2% agar prior to hemolymph collection. In each
experiment, triplicates of ~40 female flies were used to obtain ~1 ul of
hemolymph from each sample. Multiple [5-7] experiments were performed. The
amount of circulating glucose was measured using the Infinity Glucose Reagent
(Sigma) and porcine kidney trehalase (Sigma) was added to convert trehalose to
glucose as previously described [11,12].
Glycogen and
triglyceride body composition determination.
Whole body
homogenates from 10-day-old female flies were prepared as described [34]. For each
assay, triplicates of 20 μl of homogenate for eachsample were
included. Glycogen content was calculated by subtractingthe total glucose
composition without amyloglucosidase digestionfrom the total glucose
composition after amyloglucosidase digestion [34]. For,
triglyceride measurements, fly homogenates were similarly prepared and
subjected to analysis using the triacylglycerolhydrolysis kit (335-UV, Sigma).
Three independent experiments were performed. All results were normalized with
fresh fly weight measured immediately before homogenization.
Life span and stress
resistance studies.
To perform life span studies, homozygous virgins
bearing UAS-mucp1A, UAS-mucp1B, or UAS-hucp2 transgene and
control w1118 female virgins were crossed to homozygous dilp2-Gal4
driver males. The progeny from these crosses were maintained on standard corn
meal/yeast/sucrose/agar diet and passed to fresh vials every other day [6].
Both starvation and
paraquat resistance assays were conducted as described previously [6]. Briefly,
10-day-old dilp2-Gal4/UAS-mucp1, dilp2-Gal4/UAS-hucp2
and dilp2-Gal4/w1118flies
were placed in vials containing filter paper soaked in water (starvation assay)
or a solution of 20 mM paraquat and 5% sucrose (paraquat assay) and the number
of dead flies counted every 8-14 hours. Three independent experiments were
performed. Each experiment used eight to ten vials with 20 males or 20 females
in each vial (160-200 males and 160-200 females per experiment).
Female fecundity.
Female
fecundity was determined from daily counts of eggs produced by 20 individual
females in single mating pairs of dilp2-Gal4/UAS-mucp1, dilp2-Gal4/UAS-hucp2
and dilp2-Gal4/w1118flies fed with regular
yeast/sucrose/agar food. The flies were passed to new vials every day and the
number of eggs laid was counted and recorded for the first 24 days of the adult
life [6].
Statistical analysis.
Statistical
analysis for independent life span trials was performed using log-rank test
(StatView). Results for all other assays were analyzed using paired Student's t
test.
Supplementary Materials
Life spans of mUCP1 and hUCP2 expressing and control flies.
Trial 1 and Trial 2 are two independent life span experiments.
Genotype: mUCP1A and mUCP1B are two independent dilp2-Gal4/UAS-mucp1
transgenic lines; hUCP2 is dilp2-Gal4/UAS-hucp2; w1118 is dilp2-Gal4/w1118.
Median life spans are calculated by StatView. % increase is
calculated as the percent change between the w1118 flies and
UCP expressing flies. Chi-square and probability (p values)
are calculated by log-rank test (StatView). Maximum life span
is calculated as the mean life span of flies remaining at 10%
survivorship. N= number of flies in each life span trial.
Exogenous expression of the mUCP1 protein in the adult IPCs does not engender any morphological abnormalities. (A-B) Fluorescent images of adult IPCs in the
pars intercerebralis at low (A) and high (B) magnification via
GFP expression of the dilp2-Gal4/UAS-GFP flies. (C-D) Immunofluorescent
staining with an anti-mUCP1 antibody demonstrating the localization
of the mUCP1 protein in the IPCs of the dilp2-Gal4/UAS-mucp1
flies (D) but not in the IPCs of the control dilp2-Gal4/w1118
flies (C). (E1-E2) Fluorescent images of a representative adult
brain isolated from dilp2-Gal4/UAS-mucp1, UAS-GFP flies.
Images of the same brain on two different focus planes were
taken to show a total of 14 IPCs, indistinguishable from those
of a control dilp2-Gal4/UAS-GFP brain (B). Image A was taken
with a 20X objective whereas images B-E2 were taken with a 40X
objective. Scale bars, 100 μm.
Changes in intracellular Ca 2+ concentration in adult IPCs in response to nutrient
conditions. (A) A three-fold Increase in fluorescent intensity
is measured in adult IPCs producing ″camgaroo″ (Cg-2) in response
to glucose or trehalose. Each bar represents mean + S.E.M.
(N=3 independent experiments with 8-10 brains analyzed in each
experiment). *P < 0.001. (Student's t test). (B-D) Representative
images of brains of dilp2-Gal4/UAS-cg-2 flies following 16 hour-fasting
(B) and refeeding with 10% glucose. (C) or 10% trehalose
(D) for 30 minutes demonstrate an increase of Ca2+-dependent
fluorescence in adult IPCs located in the pars intercerebralis.
All images were taken with a 40X objective. Scale bar, 100 μm.
Body composition analysis of adult flies expressing mUCP1 or hUCP2 in IPCs. (A) An average
of 26% increase in glycogen storage is the result of IPC-specific
UCP expression. (B) IPC-specific UCP expression does not significantly
alter total triglyceride content of the fly. Control: dilp2-Gal4/w1118;
mUCP1: dilp2-Gal4/UASmucp1; hUCP2: dilp2-Gal4/UAS-hucp2.
Each bar represents mean + S.E.M. N=3. *P=0.005; **p=0.016 (Student's t test).
Quantitative real-time RT-PCR
analysis reveals a dramatic decrease (70%) in dilp3 expression
in dilp2-Gal4/UAS-mucp1 (mUCP1) flies as compared to Control
dilp3-Gal4/w1118 flies. The changes in transcript levels for
dilp2 and dilp5 as the result of UCP expression in IPCs are
not statistically significant. A 30% decrease in dilp3 expression
in dilp2-Gal4/UAS-hucp2 (hUCP2) females is the average of two
independent experiments. The housekeeping gene GAPDH was used
as a reference gene. Each bar represents mean + S.E.M except
for the dilp3/GAPDH value measured in dilp2-Gal4/UAS-hucp2 females
where two independent experiments were performed. N=4 independent
experiments with 4 separate RNA preparations. In each experiment,
each sample was measured in triplicate. *P<0.01 (Student's t test).
Quantitative real-time PCR analysis
reveals an average of 27% decrease in Kir expression when
IPCs are partially ablated using an IPC-specific dilp3-Gal4
driver for the expression of a pro-apoptotic gene, reaper.
The housekeeping gene GAPDH was used as a reference gene.
Each bar represents mean + S.E.M. N=4. *P=0.018 (Student's t test). Control: dilp3-Gal4/w1118.
Acknowledgments
The authors are indebted to
Drs. Mohammad Shahidullah and Irwin B. Levitan (University of Pennsylvania) for
performing the initial electrophysiological experiments on the adult IPCs. We
thank Dr. R Davis (Baylor College of Medicine), Dr. M. Tatar (Brown
University), and Dr. E. Rulifson (UCSF) for the kind gift of fly stocks; Dr. O.
Puig for the dFoxO antibody. This work was supported by grants from the NIA to
Y-WCF (AG21068, AG31086) and SLH (AG16667, AG24353, and AG25277). S.L.H. is
also supported by the Ellison Medical Foundation and Glenn Research Foundation.
S.L.H. is an Ellison Medical Research Foundation Senior Investigator.
Conflicts of Interest
The authors in this manuscript have no conflict of
interests to declare.
References
-
1.
Zhang
CY
, Baffy
G
, Perret
P
, Krauss
S
, Peroni
O
, Grujic
D
, Hagen
T
, Vidal-Puig
AJ
, Boss
O
and Kim
YB.
Uncoupling protein-2 negatively regulates insulin secretion and is a major link between obesity, beta cell dysfunction, and type 2 diabetes.
Cell.
2001;
105:
745
-755.
[PubMed]
.
-
2.
Chan
CB
, De
Leo D
, Joseph
JW
, McQuaid
TS
, Ha
XF
, Xu
F
, Tsushima
RG
, Pennefather
PS
, Salapatek
AM
and Wheeler
MB.
Increased uncoupling protein-2 levels in beta-cells are associated with impaired glucose-stimulated insulin secretion: mechanism of action.
Diabetes.
2001;
50:
1302
-1310.
[PubMed]
.
-
3.
Joseph
JW
, Koshkin
V
, Zhang
CY
, Wang
J
, Lowell
BB
, Chan
CB
and Wheeler
MB.
Uncoupling protein 2 knockout mice have enhanced insulin secretory capacity after a high-fat diet.
Diabetes.
2002;
51:
3211
-3219.
[PubMed]
.
-
4.
Krauss
S
, Zhang
CY
, Scorrano
L
, Dalgaard
LT
, St-Pierre
J
, Grey
ST
and Lowell
BB.
Superoxide-mediated activation of un- coupling protein 2 causes pancreatic beta cell dysfunction.
J Clin Invest.
2003;
112:
1831
-1842.
[PubMed]
.
-
5.
Affourtit
C
and Brand
MD.
Uncoupling protein-2 contributes significantly to high mitochondrial proton leak in INS-1E insulinoma cells and attenuates glucose-stimulated insulin secretion.
Biochem J.
2008;
409:
199
-204.
[PubMed]
.
-
6.
Fridell
YW
, Sanchez-Blanco
A
, Silvia
BA
and Helfand
SL.
Targeted expression of the human uncoupling protein 2 (hUCP2) to adult neurons extends life span in the fly.
Cell Metab.
2005;
1:
145
-152.
[PubMed]
.
-
7.
Ashcroft
FM
K(ATP) channels and insulin secretion: a key role in health and disease.
Biochem Soc Trans.
2006;
34:
243
-246.
[PubMed]
.
-
8.
Maechler
P
and Wollheim
CB.
Mitochondrial function in normal and diabetic beta-cells.
Nature.
2001;
414:
807
-812.
[PubMed]
.
-
9.
Joseph
JW
, Koshkin
V
, Saleh
MC
, Sivitz
WI
, Zhang
CY
, Lowell
BB
, Chan
CB
and Wheeler
MB.
Free fatty acid-induced beta-cell defects are dependent on uncoupling protein 2 expression.
J Biol Chem.
2004;
279:
51049
-51056.
[PubMed]
.
-
10.
Krauss
S
, Zhang
CY
and Lowell
BB.
The mitochondrial uncoupling-protein homologues.
Nat Rev Mol Cell Biol.
2005;
6:
248
-261.
[PubMed]
.
-
11.
Kim
SK
and Rulifson
EJ.
Conserved mechanisms of glucose sensing and regulation by Drosophila corpora cardiaca cells.
Nature.
2004;
431:
316
-320.
[PubMed]
.
-
12.
Rulifson
EJ
, Kim
SK
and Nusse
R.
Ablation of insulin-producing neurons in flies: growth and diabetic phenotypes.
Science.
2002;
296:
1118
-1120.
[PubMed]
.
-
13.
Broughton
SJ
, Piper
MD
, Ikeya
T
, Bass
TM
, Jacobson
J
, Driege
Y
, Martinez
P
, Hafen
E
, Withers
DJ
and Leevers
SJ.
Longer lifespan, altered metabolism, and stress resistance in Drosophila from ablation of cells making insulin-like ligands.
Proc Natl Acad Sci U S A.
2005;
102:
3105
-3110.
[PubMed]
.
-
14.
Lee
G
and Park
JH.
Hemolymph sugar homeostasis and starvation-induced hyperactivity affected by genetic manipulations of the adipokinetic hormone-encoding gene in Drosophila melanogaster.
Genetics.
2004;
167:
311
-323.
[PubMed]
.
-
15.
Ikeya
T
, Galic
M
, Belawat
P
, Nairz
K
and Hafen
E.
Nutrient-dependent expression of insulin-like peptides from neuroendocrine cells in the CNS contributes to growth regulation in Drosophila.
Curr Biol.
2002;
12:
1293
-1300.
[PubMed]
.
-
16.
Schaffer
MH
, Noyes
BE
, Slaughter
CA
, Thorne
GC
and Gaskell
SJ.
The fruitfly Drosophila melanogaster contains a novel charged adipokinetic-hormone-family peptide.
Biochem J.
1990;
269:
315
-320.
[PubMed]
.
-
17.
Noyes
BE
, Katz
FN
and Schaffer
MH.
Identification and expression of the Drosophila adipokinetic hormone gene.
Mol Cell Endocrinol.
1995;
109:
133
-141.
[PubMed]
.
-
18.
Affourtit
C
and Brand
MD.
On the role of uncoupling protein-2 in pancreatic beta cells.
Biochim Biophys Acta.
2008;
1777:
973
-979.
[PubMed]
.
-
19.
Yu
D
, Baird
GS
, Tsien
RY
and Davis
RL.
Detection of calcium transients in Drosophila mushroom body neurons with camgaroo reporters.
J Neurosci.
2003;
23:
64
-72.
[PubMed]
.
-
20.
Rameh
LE
and Cantley
LC.
The role of phosphoinositide 3-kinase lipid products in cell function.
J Biol Chem.
1999;
274:
8347
-8350.
[PubMed]
.
-
21.
Britton
JS
, Lockwood
WK
, Li
L
, Cohen
SM
and Edgar
BA.
Drosophila's insulin/PI3-kinase pathway coordinates cellular metabolism with nutritional conditions.
Dev Cell.
2002;
2:
239
-249.
[PubMed]
.
-
22.
Puig
O
, Marr
MT
, Ruhf
ML
and Tjian
R.
Control of cell number by Drosophila FOXO: downstream and feedback regulation of the insulin receptor pathway.
Genes Dev.
2003;
17:
2006
-2020.
[PubMed]
.
-
23.
Bauer
JH
, Chang
C
, Morris
SN
, Hozier
S
, Andersen
S
, Waitzman
JS
and Helfand
SL.
Expression of dominant-negative Dmp53 in the adult fly brain inhibits insulin signaling.
Proc Natl Acad Sci U S A.
2007;
104:
13355
-13360.
[PubMed]
.
-
24.
Tatar
M
, Bartke
A
and Antebi
A.
The endocrine regulation of aging by insulin-like signals.
Science.
2003;
299:
1346
-1351.
[PubMed]
.
-
25.
Koster
JC
, Permutt
MA
and Nichols
CG.
Diabetes and insulin secretion: the ATP-sensitive K+ channel (K ATP) connection.
Diabetes.
2005;
54:
3065
-3072.
[PubMed]
.
-
26.
Ashcroft
FM
and Rorsman
P.
Molecular defects in insulin secretion in type-2 diabetes.
Rev Endocr Metab Disord.
2004;
5:
135
-142.
[PubMed]
.
-
27.
Akasaka
T
, Klinedinst
S
, Ocorr
K
, Bustamante
EL
, Kim
SK
and Bodmer
R.
The ATP-sensitive potassium (KATP) channel-encoded dSUR gene is required for Drosophila heart function and is regulated by tinman.
Proc Natl Acad Sci U S A.
2006;
103:
11999
-12004.
[PubMed]
.
-
28.
Doring
F
, Wischmeyer
E
, Kuhnlein
RP
, Jackle
H
and Karschin
A.
Inwardly rectifying K+ (Kir) channels in Drosophila. A crucial role of cellular milieu factors Kir channel function.
J Biol Chem.
2002;
277:
25554
-25561.
[PubMed]
.
-
29.
Richard
DS
, Rybczynski
R
, Wilson
TG
, Wang
Y
, Wayne
ML
, Zhou
Y
, Partridge
L
and Harshman
LG.
Insulin signaling is necessary for vitellogenesis in Drosophila melanogaster independent of the roles of juvenile hormone and ecdysteroids: female sterility of the chico1 insulin signaling mutation is autonomous to the ovary.
J Insect Physiol.
2005;
51:
455
-464.
[PubMed]
.
-
30.
Hwangbo
DS
, Gersham
B
, Tu
MP
, Palmer
M
and Tatar
M.
Drosophila dFOXO controls lifespan and regulates insulin signalling in brain and fat body.
Nature.
2004;
429:
562
-566.
[PubMed]
.
-
31.
Lowell
BB
and Shulman
GI.
Mitochondrial dysfunction and type 2 diabetes.
Science.
2005;
307:
384
-387.
[PubMed]
.
-
32.
Croker
B
, Crozat
K
, Berger
M
, Xia
Y
, Sovath
S
, Schaffer
L
, Eleftherianos
I
, Imler
JL
and Beutler
B.
ATP-sensitive potassium channels mediate survival during infection in mammals and insects.
Nat Genet.
2007;
39:
1453
-1460.
[PubMed]
.
-
33.
Feng
Y
, Ueda
A
and Wu
CF.
A modified minimal hemolymph-like solution, HL3.1, for physiological recordings at the neuromuscular junctions of normal and mutant Drosophila larvae.
J Neurogenet.
2004;
18:
377
-402.
[PubMed]
.
-
34.
Sanchez-Blanco
A
, Fridell
YW
and Helfand
SL.
Involvement of Drosophila uncoupling protein 5 in metabolism and aging.
Genetics.
2006;
172:
1699
-1710.
[PubMed]
.