



Introduction
At present, the Mitochondrial Free Radical Theory of Aging (MFRTA) is one of the most widely believed and supported theories of aging. As well as putting forward an explanation for aging it allows the explanation of inter and intra species differences. According to this theory, free radicals, essentially Reactive Oxygen Species (ROS) which are produced as by-products during normal metabolism inside mitochondria provoke the accumulation of oxidative damage. The accumulation of this oxidative damage is believed to disturb cellular homeostasis which, in turn, is responsible for the aging process. In spite of its attractiveness, MFRTA has received some recent criticism [1,2,3]. Indeed, some evidence indicates that free radicals are part of a complex network of cellular signaling, and not just toxic by-products of metabolism. Their relationship with aging may therefore be far from straightforward [4].
From MFRTA is it is possible to make three basic predictions: 1) long-lived individuals or species should produce fewer mitochondrial ROS (mtROS) than those which are short-lived, 2) a decrease in mtROS production will increase lifespan and 3) an increase in mtROS production will decrease lifespan. We can add one more prediction: if mtROS is controlling aging then both lifespan and ROS production are inherently linked. Most evidence in support of MFRTA comes from comparative biology and Dietary restriction (DR) studies, which have attempted to experimentally test the first prediction. It has been shown in several systems that isolated mitochondria from long-lived animals produce fewer mtROS than short-lived ones [5]. It is from these types of studies that a general ‘law' has been proposed, such that lower mtROS production results in a longer lifespan. However, two important exceptions to this law have recently beendescribed [6,7] Ames dwarf mutant mice are the longest-living mouse strain [8], but their mitochondria produce more free radicals than normal controls. Naked-mole rats are the longest-living rodents (Maximum Lifespan (MLS) = 28 years), yet they produce mtROS at the same rate as short-lived mice (MLS= 4 years). Paradoxically, naked-mole rats also have extraordinarily low levels of glutathione peroxidase [9], which could be responsible for the accumulation of unusually elevated levels of oxidative damage in proteins, lipids and nucleic acids [10].
Dietary Restriction (DR) is the only non-genetic treatment that has been shown clearly to increase MLS in most, if not all, species where it has been applied [11]. Since DR decreases mtROS production in isolated mitochondria, a cause and effect relationship has been proposed (reviewed in [12]). However as several different physiological parameters are also coordinately altered during DR, such as insulin signaling [13] and cellular autophagy [14], it is therefore not possible to attribute exclusively this effect on lifespan to simply the attenuation of mtROS production. Moreover, moderate exercise or protein restriction have also been shown to decrease free radical production in a similar way to DR, but do not increase MLS (reviewed in [2]).
In summary, MFRTA is currently mainly supported by indirect data which show a negative correlation between free radical production in isolated mitochondria and lifespan in several different model organisms. However, correlations can suggest but not demonstrate causality. In fact, the only definitive way to test MFRTA is to specifically decrease (or increase) mtROS production and to study the effect of such a modification on lifespan. In the present study we have employed a systematic testing of all basic predictions of MFRTA using such a strategy.
Drosophila melanogaster is an excellent model organism to study aging due to its short generation time and lifespan, the availability of the genome sequence and anenormous catalogue of genetic tools. In insects, as in mammals, there is a negative correlation between free radical production in isolated mitochondria and lifespan [15]. Thus, the extreme longevity of queen ants and bees is correlated with a resistance to oxidative stress [16,17]. Evidence from studies in Drosophila melanogasterstrongly supports MFRTA (reviewed in [18]). For example, oxygen tension modulates Drosophila lifespan and gene expression maps are similar in old and chronically hyperoxic flies [19]. Moreover, antioxidant therapies appear to be effective in delaying aging in Drosophila [20,21] , although some authors claim that the increase in lifespan is only produced in short-lived lines [22] or that it is not related to oxidative damage directly but through the activation of survival-signaling pathways [23]. However, there is also data from Drosophila studies that appears to contradict MFRTA. For example, DR has been shown to increase lifespan in Drosophila [24] without altering free radical production [25]. It is for these reasons that we have chosen Drosophila as model organism to test MFRTA.
We first studied free radical production in three independent wild-type strains ofD. melanogaster which show a substantial variation in longevity. The results of our study show that the longest-lived strain produces the fewest mtROS which is consistent with MFRTA. However, the longest-lived flies could have characteristics independent of mtROS that might confer the superior longevity. A more rigorous way to examine MFRTA is to test its second prediction by directly manipulating mtROS production. If MFRTA applies, individuals producing fewer mtROS should be long-lived. Unfortunately, the exact location and mechanism by which free radicals are produced in the electron transport chain (ETC) remains unclear. This means that any genetic modification to the ETC would most likely result in an increase in free radical production and therefore deleterious effects. However, nature provides some potential solutions to by-pass this problem. Fungi and plants modulate mitochondrial free radical levels through the expression of an enzyme named the Alternative Oxidase (AOX). AOX can by-pass the mitochondrial ETC at complexes III and IV, concomitantly decreasing mtROS generation [26]. Its expression has been shown to increase lifespan, at least in some fungi [27]. Our group has recently introduced a copy of the AOX gene from the urochordate Ciona intestinalis into human cells [28,29] and into Drosophila melanogaster [30]. AOX expression confers new physiological properties to cells and animals, such as resistance to ETC inhibitors and partial rescue of metabolic alterations caused by genetic disruption of the ETC complexes or their biosynthesis. We hypothesized that AOX expression would decrease mitochondrial free radical production in Drosophila and, if MFRTA is correct, that AOX expression should therefore also increase the lifespan of individuals expressing it. The third prediction was tested using a Drosophila mutant, dj-1β, which has previously been shown to have increased mitochondrial free radical production in aged flies, manifesting as a severe impairment of locomotive function [30]. If MFRTA is correct, dj-1β mutant flies should also be short-lived. Finally, we have selected flies with mitochondrial DNA from the OR long-lived background in DAH short-lived nuclear background (and vice versa), and we have measured both mtROS and lifespan. If MFRTA is correct both parameters should be inherently linked and therefore related such that an alteration of one parameter would translate into a direct effect on the other.
Results
Testing prediction #1: "Long-lived individuals should produce fewer mtROS"
In order to test the first prediction we investigated the relationship between levels of mitochondrial ROS production and lifespan in three different wild-type strains of Drosophila melanogaster (OR, CS and DAH).
Mitochondrial ROS Production in wild-type strains
mtROS production was measured in 10 day old flies using two different substrates to identify which ETC complex or complexes (if any) are implicated in variation of mtROS production. Using a (pyruvate + proline) substrate cocktail, significant differences were detected between groups (p < 0.001, Figure 1A). OR flies (both males and females) produced fewer mtROS than the other groups. CS males produced fewer mtROS than DAH males, whereas there were no differences between DAH females and CS flies. OR and DAH females produced significantly fewer mtROS than the corresponding males. Using SP3G as a substrate significant differences were also detected (p < 0.001; Figure 1A), but these were essentially a result of a lower mtROS production of OR males with respect to the other groups. Most of the mtROS production detected using S3PG as substrate is generated during the reverse transfer of electrons between the ubiquinone pool and complex I or by complex III [31]. In relation to aging, complex I seems to be more relevant than complex III (reviewed in [5]). Therefore, to study in detail the role of complex III in mtROS production, rotenone was added and experiments using S3PG were repeated. When rotenone was present, no significant differences were detected between groups (p = 0.05; Figure 1A). These results indicate that differences between groups are due to variation in ROS produced by complex I, but only when electrons flow in the forward direction. This is in accordance with most published data, supporting an instrumental role of complex I, but not complex III, in mediating variation of mtROS production related to longevity (e.g. [6,34]).
Mitochondrial oxygen consumption
Mitochondrial oxygen consumption was studied in parallel with ROS measurements in order to investigate whether differences in ROS production are related to overall oxygen consumption or to coupling. No differences were detected either in state 4 or state 3 respiration when pyruvate + proline was used as substrate (p > 0.05; Table 1). However, RCI was significantly different between the groups (p< 0.05), being consistently lower in males than females. Using S3PG (+rotenone) a similar trend was seen (Table 1); no significant differences were found in state 4 or state 3 respiration (p > 0.05), but OR males have a lower RCI (p < 0.05) males compared to other groups.
Lifespan studies in wild-type strains
The mean and MLS of females was found to be extended in comparison with males in all the strains studied (Figure 1B). OR males lived 51% longer than DAH males and 40% longer (p < 0.001) than CS males. Strikingly, OR males also produced only 53% of the mtROS produced by DAH males and 38% of that produced by CS males when pyruvate + proline was the substrate. OR females lived longer (p < 0.001) than DAH and CS females, although differences were much smaller (12% and 8% respectively). Our data are consistent with a direct (inverse) correlation between ROS production at complex I and lifespan in wild-type Drosophila strains. In order to confirm such a relationship, we looked at further possible correlations between different parameters associated with ROS generation, oxidative metabolism and MLS. The only significant correlation found was with mtROS production using pyruvate + proline as substrate (Supplementary Figure 1). Interestingly, antioxidants levels analyzed by qPCR (Supplementary Figure 2) negatively correlate with lifespan. This is in agreement with the idea that long-lived strains decrease the generation of damage rather than increase defense or repair in order to increase longevity.
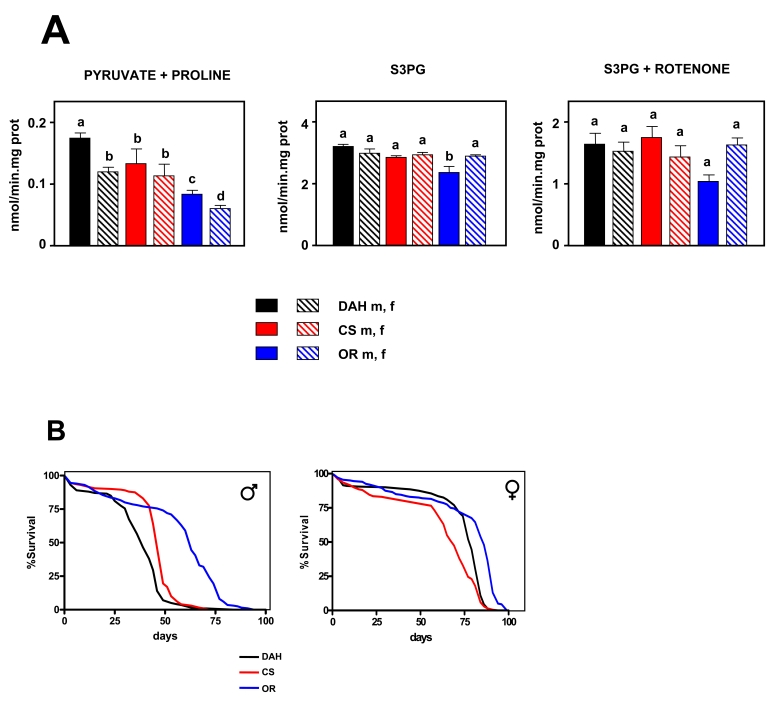
Figure 1. Mitochondrial ROS production versus lifespan in three wild type strains of Drosophila melanogaster.
(A) Rate of mtROS production (assayed as H2O2,
mean +
SEM). a, b, c and d indicate
statistically significant differences between groups (ANOVA, p <
0.05, n = 5-9 samples per group), m: male, f: female. (B) Survival
curves. Combined data from two independent experiments using 100
flies per group per experiment. Mean, maximum lifespans (d) were: DAH males
[39,49]; CS males (46, 53); OR males (63, 74); DAH females (79, 84); CS
females (69, 81); OR females (86, 91).
Table 1. Mitochondrial oxygen consumption (nmol O 2/min.mg prot) in three wild type strains of Drosophila melanogaster.
Results are presented as mean ±SEM. Number of independent samples in parentheses. Different letters (a, b) denote statically significant differences between groups. DAH = Dahomey, CS = Canton S, OR = Oregon R.
| DAH | CS | OR | ANOVA | ||||
| males | females | males | females | males | females | ||
| Pyruvate + Proline | |||||||
| State 4 | 30 ± 3 (6) | 29 ± 4 (6) | 28 ± 4 (6) | 25 ± 1 (6) | 36 ± 3 (6) | 23 ± 4 (6) | NS |
| State 3 | 313 ± 24 (6) | 390 ± 24 (6) | 366 ± 16 (6) | 393 ± 25 (6) | 316 ± 24 (6) | 324 ± 24 (6) | NS |
| RCI | 10.6 ± 1.1 (6)a | 15.2 ± 2.1 (6)b | 13.8 ± 1.1 (6)a | 15.7 ± 1 (6)b | 9 ± 1 (6)a | 14.9 ± 1.7 (6)b | p < 0.01 |
| sn-glycerol-3-Phosphate + rotenone | |||||||
| State 4 | 77 ± 10 (8) | 73 ± 9 (6) | 78 ± 10 (8) | 70 ± 7 (7) | 74 ± 11 (7) | 67 ± 6 (8) | NS |
| State 3 | 153 ± 19 (8) | 187 ± 25 (6) | 156 ± 32 (8) | 152 ± 19 (7) | 116 ± 15 (7) | 158 ± 18 (8) | NS |
| RCI | 2.2 ± 0.1 (8)a | 3 ± 0.1 (6)a | 2.3 ± 0.2 (8)a | 2.6 ± 0.2 (7)a | 1.9 ± 0.2(7)b | 2.6 ± 0.1 (8)a | p < 0.001 |
Table 2. Mitochondrial oxygen consumption (nmol O 2/min.mg prot) in in wild type flies (wt) and flies expressing (AOX/da-GAL4), or not expressing AOX (AOX/–).
AOX flies are from line F6. For equivalent data for line F24 see Supplementary Table 1. Results are presented as mean ± SEM. Number of independent samples in parentheses.
| wt | AOX/- | AOX/da-GAL4 | ANOVA | ||||
| males | females | males | females | males | females | ||
| Pyruvate + Proline | |||||||
| State 4 | 35 ± 7 (9) | 42 ± 9 (10) | 32 ± 7 (8) | 35 ± 6 (9) | 35 ± 5 (7) | 46 ± 5 (10) | NS |
| State 3 | 410 ± 45 (9) | 493 ± 48 (10) | 419 ± 41 (8) | 495 ± 40 (9) | 488 ± 54 (7) | 491±35 (10) | NS |
| RCI | 16 ± 5 (9) | 17 ± 5 (10) | 15 ± 3 (8) | 19 ± 4 (9) | 15 ± 2 (7) | 15 ± 1 (10) | NS |
| sn-glycerol-3-Phosphate + rotenone | |||||||
| State 4 | 65 ± 16 (6) | 88 ± 27 (6) | 132 ± 28 (8) | 112 ± 12 (7) | 98± 29 (5) | 104 ± 14 (8) | NS |
| State 3 | 255 ± 58 (8) | 273 ± 35 (6) | 254 ± 27 (8) | 288 ± 45 (7) | 241 ± 26 (5) | 276 ± 44 (8) | NS |
| RCI | 2.1 ± 0.2 (8) | 2.2 ± 0.4 (6) | 2.3 ± 0.7 (8) | 2.3 ± 0.4 (7) | 2.4 ± 0.7(5) | 2.9 ± 0.5 (8) | NS |
Testing prediction #2: "A decrease in mtROS production should increase MLS"
Expression of AOX in DAH background
In order to check the second prediction of MFRTA we expressed the alternative oxidase (AOX) of Ciona intestinalis in flies, after backcrossing to the DAH background for 11 generations (the flies expressing the daughterless-GAL4 driver were also backcrossed in the same conditions). AOX is able to regulate mtROS generation in plants and fungi, and its expression has been related to an increase in longevity in fungi [26,27]. Firstly, we performed some routine experiments to check the presence and functionality of AOX in vivo, in the backcrossed flies. We tested resistance to three different inhibitors of the ETC: 1) rotenone (Complex I), 2) antimycin A (Complex III) and 3) KCN (Complex IV). Flies expressing AOX showed an increased resistance to antimycin A and KCN compared to non-expressing flies (Figure 2A and Supplementary Figure 3). Differences in survival were observed after only 10 min of exposure to drugs inhibiting either complex III or IV. After 24 h of exposure only flies expressing AOX survived. However, no difference was observed when a complex I inhibitor (rotenone) was employed. In order to confirm that these observations were a result of AOX expression we tested the effect of the inhibitors also on mitochondrial bioenergetics and ROS production. Using isolated mitochondria we observed that AOX is able to support state 3 oxygen consumption in the presence of antimycin or KCN, but not in the presence of rotenone (Figure 2B). Moreover, AOX decreased mtROS production in the presence of complex III or IV inhibitors, but not in the presence of an inhibitor of complex I (Figure 2C). These data imply that AOX is expressed and is functional in vivo. Additionally AOX behaves as theoretically expected, e.g. AOX-expressing flies are resistant to blocks in complex III or IV, but not I.
Effects of AOX on mtROS production and oxygen consumption
Having established that AOX was functional in vivo, we studied mtROS production in isolated mitochondria in normal conditions (i.e. without inhibitors). The same experiments carried out in wild-type strains were repeated in AOX transgene-expressing and non-expres-sing flies from two independent transgenic lines and in wild type (DAH) controls. AOX was found to decrease mtROS production in 2-3 day old flies when either pyruvate + proline (by 32-34%) or S3PG (by 16-20%) was used as a substrate (Figure 3A, S4). When rotenone was also present in the assay medium AOX flies still produced fewer (27-37%) mtROS than controls with S3PG as substrate (Figure 3A). However; AOX did not modify oxygen consumption in state 3 nor state 4 (Tables 2, S1). We also studied the effects of AOX expression in aged flies. We repeated the same measurements in 30 day old males and 50 day old females, representing equivalent time points in normal male and female lifespan in the DAH background, but under which conditions more than 50% of flies are still alive, thus avoiding the selection of a sub-population.
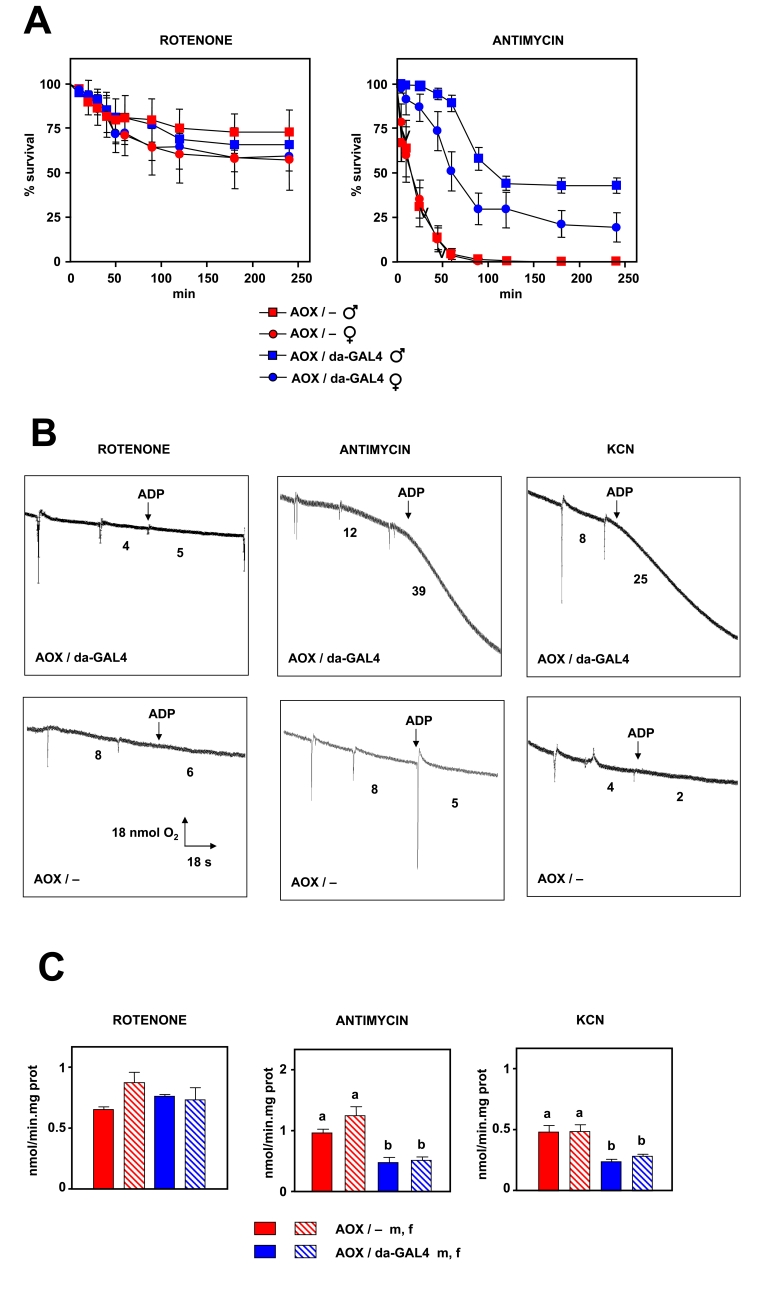
Figure 2. Effects of AOX expression on resistance to respiratory chain inhibitors. (A) Survival
after exposure to 3
mM rotenone or 3 mM antimycin A, of flies of strains and sexes indicated
(AOX / -, flies transgenic for UAS-AOX in absence of GAL4 driver; AOX /
da-GAL4, flies transgenic for AOX in presence of da-GAL4 driver). (B)
Representative oxygraph traces of mitochondrial suspensions (0.5 mg/ml in
state 3) in presence of inhibitors shown. Inferred oxygen consumption rates
(nmol/min) as indicated. Pyruvate+proline was used as substrate in all
experiments. (C) mtROS production (mean +
SEM) in presence of
inhibitors (at least 4 independent samples per experiment, a, b denote
significantly different groups, ANOVA, p < 0.05).
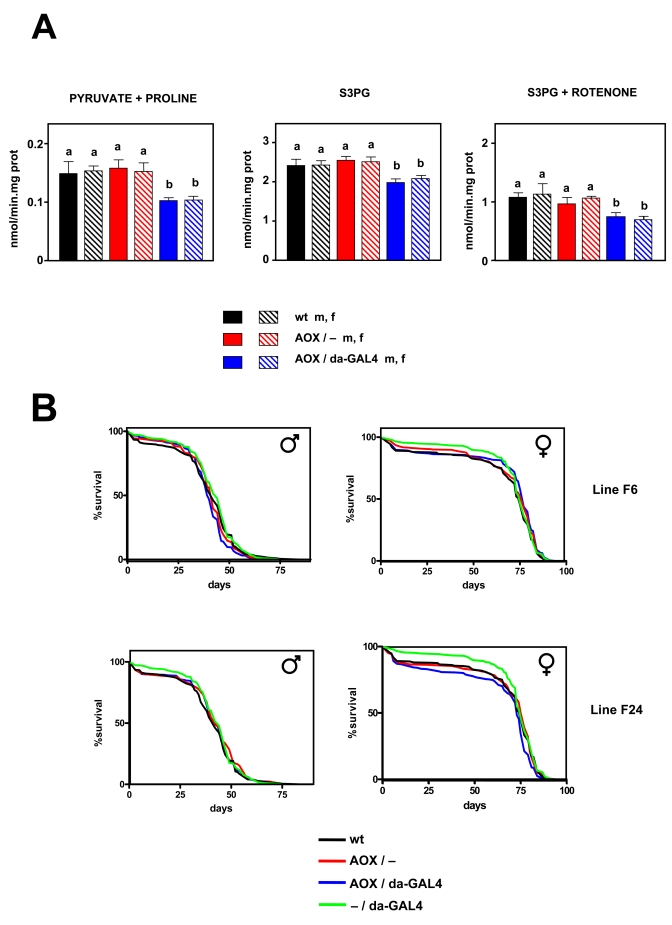
Figure 3. Effect of AOX expression on mtROS production and lifespan. (A) mtROS
production (mean +
SEM). a, b: statistically significant differences
between groups (ANOVA, p < 0.05, n=4-8 samples per group) m:
male, f: female. (B) Survival curves for wild
type (wt), AOX non-expressing (AOX / -), AOX expressing (AOX / da-GAL4 +),
and driver only (- / da-GAL4) flies, all in the DAH (w-)
background. Flies of AOX transgenic lines F6 and F24 as indicated. Combined
data from two independent experiments using 200 flies per group per
experiment. Mean, maximum life spans (d) were: wt males (42,51); wt
females (75, 82); - / daGAL4 males (44, 54); -
/ da-GAL4 females (75, 81), F6 AOX / - females (77, 82); F6 AOX / - males
(42,51); F6 AOX / da-GAL4 males (40,47); F6 AOX / da-GAL4 females (82,
51); F24 AOX / - males (42, 54); F24 AOX / - females (77, 81); F24 F24 AOX /
da-GAL4 males (42, 54); AOX / da-GAL4 females (73, 80).
At the ages studied, AOX also decreased mtROS production, both in the presence of KCN and absence of ETC inhibitors (Supplementary Figure 5). During aging mitochondrial oxygen consumption was strongly decreased and AOX was not able to compensate this decrease (Figure 4A, B). At the same time mtROS generation was increased, but AOX was able to negate the increase in such a way that mtROS production in old AOX-expressing flies was similar to that in young control flies (Figure 4C, D).
Lifespan and AOX
Lifespan was studied in the same two AOX transgenic- lines (F6 & F24). Both lines have an AOX insertion in an intergenic region, but on different chromosomes (2 and 3, respectively, [30]). Two independent experi-ments each with 200 flies per group were carried out. AOX did not significantly increase lifespan in any of the lines studied (Figure 3B). In males, AOX had a slightly deleterious effect on longevity in line F6 (MLS decreased by around 9%; p < 0.05), but none at all in line F24 (p > 0.05), whereas in females the opposite was found: no differences were observed in line F6 (p > 0.05), but it in line F24 a small decrease (1-3%, p < 0.05) was seen. In summary, AOX expression did not consistently or significantly modify lifespan in Drosophila.
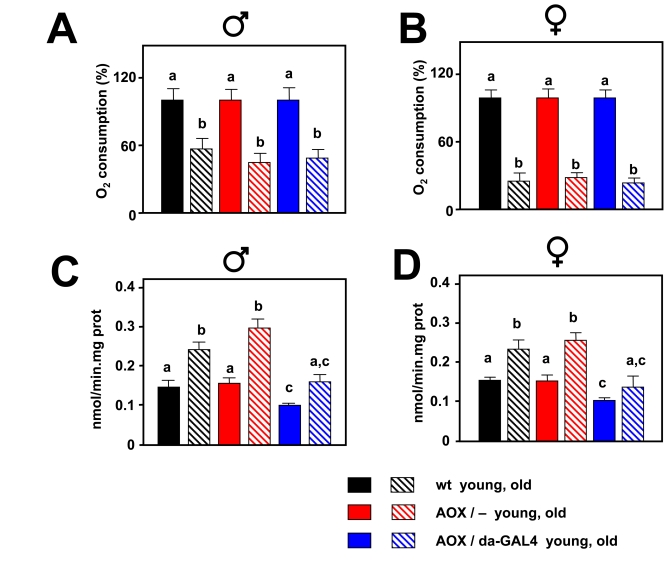
Figure 4. Effect of AOX on mitochondrial bioenergetics and mtROS during aging. Oxygen
consumption in state 3 (% of that in the young group) in 30 d old males (A)
and 50 d old females. (B) AOX expression is not able to compensate
the decrease in oxygen consumption associated with aging. mtROS generation
(nmol H2O2/min.mg.prot) in 30 d old males (C)
and 50 d old females (D). AOX expression diminishes mtROS production
in both young and aged flies, and compensates for the age-associated
increase. a, b and c denote statistically significant differences between
groups (ANOVA, p < 0.05, n = 4-10 samples per group).
Pyruvate+proline was used as substrate in all experiments. Plotted data are
means ± SEM.
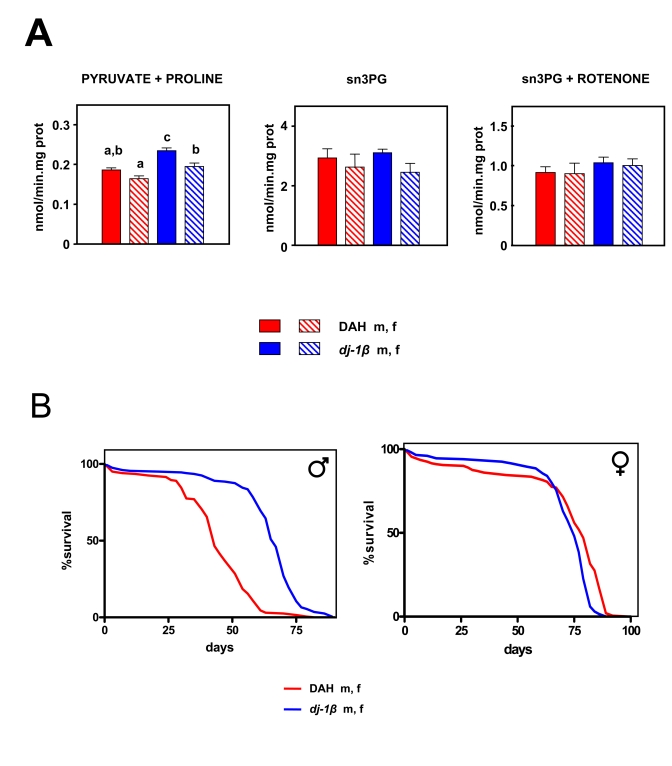
Figure 5. Effects of the dj-1β mutation on mtROS production and lifespan in Drosophila melanogaster. (A) mtROS production, assayed as H2O2,
(mean + SEM). a, b and c denote statistically significant differences between
groups (ANOVA, p<0.05, n = 4-6 samples per group), m: male, f: female. (B)
Survival curves. Combined data from two independent experiments using 100 flies per
group per experiment. Mean, maximum lifespans (d) were: DAH males (43, 58); DAH
females (75, 79); dj-1β mutant males (67, 75); dj-1β mutant females (79, 86).
Testing prediction #3: "An increase in mtROS production should decrease MLS"
Mitochondrial free radical production and oxygen consumption in dj-1β mutant flies
We measured mitochondrial free radical production in 10 day old dj-1β mutant flies using flies from the DAH background as controls. As expected, dj-1β mutant flies produced more mtROS than wild-type controls with pyruvate + proline as substrate (Figure 5A; p < 0.001), although no significant differences were observed when S3PG was used as a substrate (Figure 5A; p > 0.05). Differences in free radical production were not reflected in oxygen consumption (Table 3). Recently, we showed that dj-1β mutant flies produce more mtROS than wild-type flies at 3 weeks of age [30].In our previous report only pyruvate + prolinewas used as a substrate. Our present findings confirm these results and clarify the mechanism whereby the dj-1β mutation alters mtROS production. Only when electrons flow in the forward direction through complex I are differences detected between mutants and controls. Together, these data support the idea that dj-1β works as a peroxiredoxin [35]. When pyruvate + proline is used as substrate most of the ROS generated are directed to the mitochondrial matrix where dj-1β can exert its detoxifying action, whereas when SP3G is used as the substrate ROS production is split between the matrix and the inter-membrane space [31], decreasing the potential role of dj-1β in the detoxification process.
Lifespan of dj-1β mutant flies
In spite of increased levels of mtROS production dj-1β mutant flies were found to have a longer, not shorter lifespan than DAH flies of the corresponding sex: by 30% in males and 9% in females (Figure 5B, p < 0.001). Moreover, even after seven generations of backcrossing in to the DAH background (reducing background effects to a minimum) differences in lifespan between mutants and non-mutants for the dj-1β were maintained (Supplementary Figure 7A).
Table 3. Mitochondrial oxygen consumption (nmol O 2/min.mg prot) in DAH wild-type and dj-1β mutant.
| DAH | dj-1β | |||
| males (6) | females (7) | males (7) | females (10) | |
| State 4 | 33 ± 5 | 33 ± 8 | 38 ± 4 | 28 ± 5 |
| State 3 | 322 ± 16 | 396 ± 58 | 279 ± 37 | 281 ± 22 |
| RCI | 13± 2 | 12±1 | 9± 1 | 13 ± 2 |
Lifespan of dj-1β mutant flies
In spite of increased levels of mtROS production dj-1β mutant flies were found to have a longer, not shorter lifespan than DAH flies of the corresponding sex: by 30% in males and 9% in females (Figure 5B, p < 0.001). Moreover, even after seven generations of backcrossing in to the DAH background (reducing background effects to a minimum) differences in lifespan between mutants and non-mutants for the dj-1β were maintained (Supplementary Figure 7A).
Testing prediction #4: "An increase in mtROS production should decrease MLS"
Analysis of mitochondrial DNA
The nucleotide sequence of mitochondrial gene coI was analyzed in three different wild type strains of Drosophila melanogaster (OR, DAH and CS) as described in material and methods. We found 10 different polymorphic sites (Supplementary Table 3), although none of the modifications in the gene analyzed caused changes in the amino acid sequence of the protein (all of them were synonymous substitutions). The presence of so many polymorphisms indicates that they could play an important role in Drosophila physiology including aging and ROS production and so we investigated the question in more detail.
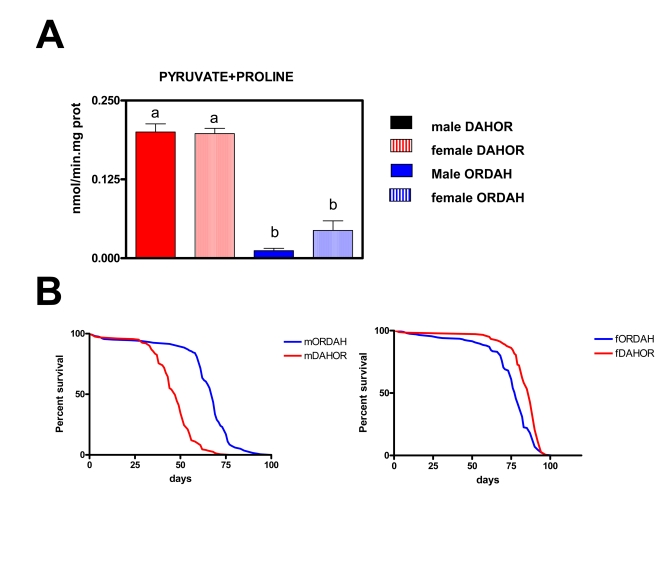
Figure 6. Effects of the changes on mtDNA content on mtROS production and lifespan in new wild type strains of Drosophila melanogaster (DAHOR and ORDAH).
(A) mtROS production, assayed as H2O2,
(mean + SEM). a and b denote statistically significant differences between
groups (ANOVA, p <0.05, n = 4-5 samples per group), m: male, f: female.
(B) Survival curves. Combined data from two independent experiments
using between 80-100 flies per group per experiment. Mean, maximum lifespans
(d) were: ORDAH males (68, 76); ORDAH FEMALES (78, 87); DAHOR males (47,59);
DAHOR females (87,92).
ROS production
Initially we wanted to know if mitochondrial ROS production was modulated by polymorphisms in mtDNA so we decided to create new strains of Drosophila melanogaster putting the mitochondrial genome of OR flies in a DAH nuclear background (and vice versa). We measured ROS production in 2/3 days old flies (Figure 6A). At this age we did not find any significant differences between males and females, this mirrors what is seen in the original DAH background where sex differences are only detected after 10 days. ROS production was lower (around 86%) in flies with an OR nuclear background independently of the DAH mitochondrial DNA background. In fact no differences in ROS were found between OR and ORDAH flies or between DAH and DAHOR flies (data not shown). This data clearly shows that mitochondrial DNA does not control free radical production or at least the free radical production related with longevity in these wild type Drosophila strains. Similar results were obtained when OR mtDNA was expressed in a CS nuclear background and vice versa (data not shown), this indicates that the phenomenon is not restricted to the DAH/OR strains.
Life span of DAHOR and ORDAH strains
Males and females answered differently to changes in the mitochondrial DNA composition. In the males (Figure 6B) the OR background resulted in a longer lifespan (45% mean and 29% MLS) independently of the mitochondrial DNA. This is in agreement with the lower levels of ROS generation in ORDAH flies. However, in females the results were totally opposite, mtDNA determines longevity independently of either nuclear DNA or levels of ROS generation. According to this females with OR mtDNA live longer than females with DAH mtDNA. Mean and maximum lifespan were 12% and 6% (p<0,001) respectively longer in DAHOR females than in ORDAH females in spite of this ROS production was 4.5-fold times higher in the DAHOR females (p<0.001).
Discussion
The MFRTA is one of themost widely invoked hypotheses accounting for aging, yet the evidence in support of it is almost entirely indirect. In this study we set out to test its predictions experimentally. Although we found a negative correlation between mtROS production and lifespan in 3 wild-type strains of Drosophila melanogaster (further support for low level of expression of antioxidants in long-lived individuals) lifespan was not modified as predicted, as a result of genetic manipulations designed to alter mtROS levels. Moreover, we were able to dissociate lifespan and ROS production in wild type strains through changes in the mitochondrial DNA.
A negative correlation between mtROS production in isolated mitochondria and lifespan in flies, mammals and birds [6,15,36] and under conditions of DR [12] has been previously reported. We found a similar relationship, with a long-lived strain (OR) producing fewer mtROS than short-lived strains (DAH and CS), without a major alteration in oxygen consumption. This was more pronounced in males, and held up in females only using a complex I-linked substrate mix. Our findings thus resolves contradictions of previous studies and emphasizes the importance of studying both sexes and using both complex I- and III-linked substrates. For example, Miwa et al. [25] found no correlation between longevity and mtROS in flies subjected to DR, but their study only looked at females using S3PG as a substrate. Conversely, Sohal et al. [15] did find a correlation using S3PG in males. However, all such correlations provide only indirect support for MFRTA. Interestingly, the expression of 4 antioxidants negatively correlates with lifespan. This supports the idea that long-lived species produce fewer mtROS and consequently need lower levels of antioxidants.
AOX in plants and fungi has been shown to decrease mtROS production when the cytochrome segment of the respiratory chain, but not complex I, is inhibited. When C. intestinalis AOX was expressed in Drosophila (this study and [30]) mtROS production was similarly diminished in the presence of antimycin or cyanide, but not rotenone. Furthermore, AOX expression sustained a substantial cyanide- or antimycin-resistant substrate oxidation in mitochondrial suspensions. AOX expression also decreased mtROS production under basal conditions, using either pyruvate + proline or S3PG (with or without rotenone) as a substrate. In plants, the ability of AOX to decrease mtROS production depends on its ability to keep the ubiquinone pool oxidized [37]. The same could apply in Drosophila, where semi-ubiquinone at complex I is considered a major site of mtROS generation [31]. We were unable to detect differences in oxygen consumption under basal conditions (i.e. without inhibitors) in AOX-expressing flies, suggesting that AOX does not exert a major effect on respiration in vivo. However, it is possible that AOX has a subtle effect on normal respiration that cannot be detected by polarography. The effect on mtROS production of AOX expression in the DAH background was to diminish it to levels similar to those of wild-type OR flies. In addition, AOX suppressed the age-associated increase in mtROS production, but not the age-associated decrease in substrate oxidation by isolated mitochondria [38,39]. Since AOX expression produced no significant effect on lifespan, this is consistent with the idea that a mitochondrial parameter other than ROS production could be a determinant of aging, as proposed e.g. by Trifunovic and Larsson [40]. Our data showing that mtDNA composition could regulate lifespan supports such an idea.
Previous attempts to test the MFRTA by disruption of complex II, both in Drosophila [41] and Caenorhabditis [42] are limited by the fact that this treatment clearly produces pleiotropic effects on energy metabolism and development. In contrast, the dj-1β mutation produces no negative effect on development or fecundity, its only known phenotypes being age-associated loss of locomotor function and a hypersensitivity to paraquat. Both of these have been attributed to the deficit of mitochondrial antioxidant capacity, as manifested by increased mtROS production compared with that of wild-type strains, which we confirmed in vitro, using flies of different ages. Indeed, the locomotor deficiency is corrected by AOX expression [30], which correlates with decreased mtROS production. Nevertheless, the dj-1β mutation did not result in shortened lifespan. Surprisingly, the opposite was observed, with the lifespan dj-1β flies comparable with that of wild-type OR flies. Moreover, even after backcrossing the dj-1β mutants for seven generations into a DAH background differences in longevity were still present. Surprising, the level of expression of antioxidants in dj-1β mutants is reduced when compared to controls. This indicates that compensation in antioxidant levels can not account for the long lifespan of dj-1β mutants. Even more surprising is the fact that correlation between mtROS and lifespan is lost when dj-1β flies are included, but the correlation between antioxidant levels and lifespan becomes more significant when dj-1β are included in the correlation (Supplementary Figure 8B,C). DAH background was used as a control for dj-1β (in experiments using either isogenic or non-isogenic lines) in order to keep the flies in experiments increasing (dj-1b mutants) or decreasing (AOX) ROS the same. For this reason, it may be argued that a decrease in lifespan may be found when dj-1β mutation is expressed in a long-lived background (e.g. OR). In any case, ROS themselves are bad predictors of lifespan as both OR and dj-1β are long-lived (compared to DAH) in spite of opposite levels of ROS.
One possible objection to our conclusion that AOX does not increase lifespan despite diminishing mtROS production would be that the enzyme may not be functional under normal physiological conditions. However, its ability to complement several mutations affecting cytochrome oxidase function, as well as the toxicity of cyanide and antimycin in vivo and the overproduction of ROSin vitro and in vivo caused by the dj-1β mutation [30] suggests otherwise. It can also be argued that an "exogenous" protein cannot increase the lifespan of the host organism. However, it has been previously shown that the expression of human UCP-2 increases Drosophila lifespan. And moreover, we and others have recently demonstrated that it is possible to increase Drosophila lifespan by expressing: NDI1 that as AOX is not encoded in the animal host genome [43,44]. Interestingly, the expression of NDI1 (a protein that can by-pass mitochondrial complex I) significantly increases lifespan without decreasing the basal rate of ROS production.
We also cannot exclude that AOX has other, undetected effects influencing lifespan, which over-ride those mediated through decreased mtROS production. Note, however, that AOX expression has only a minimal effect on the development or physiology of wild-type flies [30] and it does not alter the expression of major antioxidants (Supplementary Figure 6). Certainly, we cannot discard either that the expression of AOX in certain tissues (e.g. the nervous system) or during different life stages (e.g. in the last part of life) may have a different effect on longevity. A similar point could be made with regard to dj-1β mutant flies, i.e. that increased mtROS production in vitro and paraquat sensitivity in vivo do not reflect a systematic effect on mtROS levels in vivo under normal physiological conditions. Thus, the over-production of ROS in dj-1βmutant flies could be compensated by the alteration of another function of the protein. DJ-1 participates in RNA metabolism and transcription [45] so its effects on gene expression could compensate for the over-generation of ROS. However, all these caveats do not apply to our fourth experimental approach. Where mitochondrial DNA of OR is expressed in a DAH nuclear background (and vice versa) ROS production is not altered (it is totally determined by the nuclear background), but lifespan of females is significantly changed depending on the mitochondrial background. The fact that DAHOR females are a long-lived strain in spite of high levels of mtROS production is a strong argument against MFRTA. Moreover, this demonstrates that it is possible to separate lifespan and ROS production in wild type strains of Drosophila.
Drosophila melanogaster strains selected for long or short lifespan [46] exhibit differences in several physiological parameters (including mtROS production and the levels of antioxidant proteins). It has been suggested that longevity evolves through coordinated changes in multiple genes and biochemical pathways [47], which could accommodate our results by postulating that altered mtROS production or detoxification cannot have a material effect on lifespan without concomitant changes in other pathways, such as protein acetylation, insulin signaling or alterations in the degree of un-saturation of lipids in biological membranes. Moreover, a lifespan-increasing effect in one parameter, such as a decrease in mtROS production, could result in a compensatory change in another, such as the repair proteins of DNA, resulting in no net alteration in lifespan. Once time more the results of DAHOR and ORDAH females support such hypothesis.
Regardless of the molecular reasons, our findings indicate that mtROS production is not and cannot be the sole determinant of lifespan in Drosophila, strengthening similar conclusions arrived at recently in studies of naked mole rats [9,10], long-lived Ames dwarf mice [7] and C. elegans [48]. However, our findings are subject to two important caveats. First, the use of in vitro assays to measure mtROS production, which may not reflect the situation in vivo. And second, the assumption that AOX expression or the mutational downregulation of dj-1β does not produce pleiotropic or off-target effects that negate or over-ride effects on mtROS (see above).
The first caveat is one shared with the great majority (if not all) of studies supporting MFRTA. In fact, it is still not known if long-lived animals produce fewer free radicals than short-lived ones in vivo. Further, it has only been demonstrated that isolated mitochondria from long-lived animals produce fewer molecules of H2O2(data about superoxide are contradictory). Additionally, differences in ROS generation in isolated mitochondria are only observable under certain experimental conditions. For example, it is currently assumed that dietary restricted animals produce fewer mtROS than ad libitum-fed animals. However, differences are only observed when pyruvate (+malate) (or glutamate (+malate)) are used as a substrate [12]. On the other hand, the use of isolated mitochondria is required due to the lack of a sensitive method that would allow quantification of specific free radical species in cells (reviewed in [49]). As an example of this type of situation we could mention the work carried out in Brian Merry's laboratory. His laboratory had previously reported differences in ROS production between ad libitum and caloric restricted rats using isolated liver mitochondria [50]. However, no differences were observed in the same experimental model when intact hepatocytes were used for these measurements [51].
In summary, in order to test MFRTA we chose an original strategy of trying to modulate the generation of damage and not just increase antioxidant defense or repair mechanisms. With this we avoid the drawbacks and caveats of the latter approach [5]. In fact, even if our results do not reflect the situation in vivo, they are enormously relevant since they clearly dissociate -for the first time- levels of mtROS production in isolated mitochondria and longevity in Drosophila melanogaster, indicating that other mitochondrial factors such as the presence of polymorphisms in mitochondrial DNA may act as longevity regulators.
Materials and methods
Flies. Drosophila wild-type strains Dahomey (DAH), Canton S (CS) and Oregon R (OR) were obtained from stock-centers or collaborators. The dj-1βGE23381 mutant [31]: and AOX-transgenic lines F6 and F24 [30] were as described previously. Flies were maintained in a standard medium [30], collected using CO2 anesthesia within 24 h of eclosion, and then kept at a density of 20 flies per vial at 25 ºC in a controlled 12 h light:-dark cycle. Vials were changed every 2-3 days. We have created two new wild type strains of Drosophilamelanogaster backcrossing for eleven generations DAH virgin females with OR males and OR virgin females with DAH males. The new strains of Drosophila melanogaster are called DAHOR (flies with nuclear DAH DNA and mitochondrial OR DNA) and ORDAH (flies with nuclear OR DNA and mitochondrial DAH DNA).
Lifespan studies. Between 180 and 400 flies were used for each study. Each independent study was repeated twice: data were pooled and analysed together. Flies were collected within 24 h after eclosion using CO2 anaesthesia and kept at a density of 20 flies per vial at 25 ºC in a controlled 12 h light:-dark cycle. Every 2-3 days vials were changed and the number of dead flies was counted, from which mean and maximum lifespan (MLS, the last 10% of surviving flies) were calculated. Prism GraphPad software was utilized to build survival curves that were further analysed using the Kaplan Meier Log-Rank Test.
Mitochondrial biochemistry. Mitochondria were isolated according to Miwa et al.[31] with some minor modifications [30]. Mitochondrial respiration rates were measured by polarography using a Clark-type oxygen electrode as previously [30], in the absence or presence of KCN (100 μM), antimycin A (10 μM) or rotenone (5 μM). Mitochondrial ROS production was assayed according to the method described by [32] adapted to flies [30].
RNA quantification. Total RNA was extracted from 10 days old flies according to [30]. For cDNA synthesis, 13 μl reaction mixes containing 2 μg RNA, 1 μl DEPC 10 mM dNTP mix (Fermentas), 0.4μl Random Primers (0.5ug/μl Promega) and DEPC-treated water were incubated at 90°C for 3 min, then transferred to ice, where 4 μl 5x M-MuLV reaction buffer (Fermentas) and 1μl 40U/μl RNase inhibitor (Fermentas) were added. The reactions were mixed and incubated at 25°C for 10 min. On ice, 2μl of 20U/μl M-MuLV reverse transcriptase (Fermentas) was added, and the reaction was incubated for a further 10 min at 25°C, 1 h at 37°C and 70°C for 10 min. mRNA levels were analyzed by Q-RT-PCR. The transcript levels of RpL32, Catalase, Superoxide dismutase 1 and 2 and Glutathione Peroxidase were measured using primers pairs shown in supplementary Table 2. All RNA extractions were performed in triplicate, with each used as a template for three separate cDNA synthesis reactions which were then pooled. Each cDNA pool was itself analysed in triplicate. Expression of the target genes was measured relative to that of RpL32 (rp49), in order to normalize for sample and run to run variations. A series of 10-fold dilutions of an external standard was used in each run to produce a standard curve. Analytical reactions were performed using 20-fold diluted cDNA samples, in 25 μl reaction volume consisting of 2 μl of the cDNA template, 0.4μl of 20 μM forward and reverse primers, and 12.5ul of 2x MAXIMA SYBR GREEN Master Mix (Fermentas). The PCR program consisted of a 10 min pre-incubation at 95°C, 40 cycles of 35 secs denaturation at 95°C, 30 secs annealing at 60°C and 30 secs extension at 72°C. Melting curve analysis, consisting of a 15 secs denaturation step at 95°c followed by a 1 min annealing step at 60°C and a 0.3°C/s denaturation ramp to 95 °C, was performed after the amplification step to verify that only a single, specific extension product had been amplified. Data were extracted and analysed using Applied Biosystems StepOne software version 2.0.
Resistance to inhibitors of the ETC. To check the expression and activity of AOX in vivo experiments with a variety of ETC inhibitors were performed. 20 flies were kept (males and females separately) in fresh vials. To measure resistance to KCN, the drug was dissolved in water at varying concentrations and added directly to the food vial. Resistance to antimycin and rotenone was assayed essential as described by Fridell et al. [33]. In brief, 2-3 day old flies were starved for two hours in empty vials, following this flies were placed in vials containing Whatman paper (3 mm x 1 mm) impreg-nated with 5% (w/v) sucrose solution and the appropriate drug (3 mM antimycin or rotenone). Under these conditions without any drug, flies are able to survive more than 72 h so any effect before this time should be considered to be provoked by exposure to the drug. The proportion of flies surviving was recorded over 24 h.
Sequencing of Mitochondrial gene cytochrome c oxidase subunit I. Mitochondrial DNA was extracted using standard procedures from mitochondria isolated from around 150 flies according to Miwa et al. [31] High fidelity PCR using specific primers CoIF2 and CoIR5 (Supplementary Table 2) were used to amplify a 2.6 kb fragment containing the cytochrome c oxidase subunit I (CG34067, CoI). PCR products were purified using a Machary - Nagel PCR purification kit according to manufacturer's instructions. Products were sequenced using Big dye Terminator Chemistry 3.1v (Applied Biosystems) and a 3130 AB genetic analyser. AB sequencing analysis software was used for analysis of electropherograms.
Statistical analysis. Data were analysed using GraphPad Prism 4 and one-way ANOVA was used for statistical testing. When ANOVA was significant (p < 0.05) Newman-Keuls Multiple Comparison test was also used. Lifespan data were analysed using the Kaplan Meier Log-Rank Test. The statistically significant value was established as p < 0.05.
Acknowledgments
Our work is supported by funding from the Academy of Finland, Tampere Hospital Medical Research Fund, Juselius Foundation, the European Union and EMBO (long-term fellowship to AS). We thank Dr. J Chung for supplying the dj-1beta mutant stock.
Supplementary Materials
Conflicts of Interest
The authors of this manuscript have no conflict of interest to declare.
References
- 1. Linnane AW , Kios M and Vitetta L. Coenzyme Q(10)—its role as a prooxidant in the formation of superoxide anion/hydrogen peroxide and the regulation of the metabolome. Mitochondrion. 2007; 7: S51 -S61. [PubMed] .
- 2. Sanz A and Stefanatos R. The Mitochondrial Free Radical Theory of Aging: a critical view. Current Aging Science. 2008; 1: 10 -21. [PubMed] .
- 3. Perez VI , Bokov A , Van Remmen H , Mele J , Ran Q , Ikeno Y and Richardson A. Is the oxidative stress theory of aging dead. Biochim Biophys Acta. 2009; 1790: 1005 -1014. [PubMed] .
- 4. de Magalhaes JP and Church GM. Cells discover fire: employing reactive oxygen species in development and consequences for aging. Exp Gerontol. 2006; 41: 1 -10. [PubMed] .
- 5. Sanz A , Pamplona R and Barja G. Is the mitochondrial free radical theory of aging intact. Antioxid Redox Signal. 2006; 8: 582 -599. [PubMed] .
- 6. Lambert AJ , Boysen HM , Buckingham JA , Yang T , Podlutsky A , Austad SN , Kunz TH , Buffenstein R and Brand MD. Low rates of hydrogen peroxide production by isolated heart mitochondria associate with long maximum lifespan in vertebrate homeotherms. Aging Cell. 2007; 6: 607 -618. [PubMed] .
- 7. Csiszar A , Labinskyy N , Perez V , Recchia FA , Podlutsky A , Mukhopadhyay P , Losonczy G , Pacher P , Austad SN , Bartke A and Ungvari ZI. Endothelial function and vascular oxidative stress in long-lived GH/IGF-deficient Ames dwarf mice. Am J Physiol Heart Circ Physiol. 2008; 295: H1882 -1894. [PubMed] .
- 8. Bartke A , Wright JC , Mattisson JA , Ingram DK , Miller RA and Roth GS. Dietary restriction and lifespan. Science. 2002; 296: 319 [PubMed] .
- 9. Andziak B , O'Connor TP and Buffenstein R. Antioxidants do not explain the disparate longevity between mice and the longest-living rodent, the naked mole-rat. Mech Aging Dev. 2005; 126: 1206 -1212. [PubMed] .
- 10. Andziak B, O'Connor TP, Qi W, DeWaal EM, Pierce A, Chaudhuri AR, Van Remmen H, Buffenstein R. High oxidative damage levels in the longest-living rodent, the naked mole-rat. Aging Cell. 2006; 5: 463 -471. [PubMed] .
- 11. Weindruch R and Sohal RS. Seminars in medicine of the Beth Israel Deaconess Medical Center. Caloric intake and aging. N Engl J Med. 1997; 337: 986 -994. [PubMed] .
- 12. Gredilla R and Barja G. Minireview: the role of oxidative stress in relation to caloric restriction and longevity. Endocrinology. 2005; 146: 9:3713 -3719. .
- 13. Droge W Oxidative aging and insulin receptor signalling. J Gerontol A Biol Sci Med Sci. 2005; 60: 1378 -85. [PubMed] .
- 14. Wohlgemuth SE , Julian D , Akin DE , Fried J , Toscano K , Leeuwenburgh C and Dunn WA Jr. Autophagy in the heart and liver during normal aging and calorie restriction. Rejuvenation Res. 2007; 10: 281 -292. [PubMed] .
- 15. Sohal RS , Sohal BH and Orr WC. Mitochondrial superoxide and hydrogen peroxide generation, protein oxidative damage, and longevity in different species of flies. Free Rad Biol Med. 1995; 19: 499 -504. [PubMed] .
- 16. Parker JD , Parker KM , Sohal BH , Sohal RS and Keller L. Decreased expression of Cu-Zn superoxide dismutase 1 in ants with extreme lifespan. Proc Natl Acad Sci USA. 2004; 101: 3486 -3489. [PubMed] .
- 17. Haddad LS, Kelbert L, Hulbert AJ. Extended longevity of queen honey bees compared to workers is associated with peroxidation-resistant membranes. Exp Gerontol. 2007; 42: 601 -609. [PubMed] .
- 18. Muller FL , Lustgarten MS , Jang Y , Richardson A and Van Remmen H. Trends in oxidative aging theories. Free Rad Biol Med. 2007; 43: 477 -503. [PubMed] .
- 19. Landis GN , Abdueva D , Skvortsov J , Yang BE , Rabin J , Carrick S , Tavare S and Tower J. Similar gene expression patterns characterize aging and oxidative stress in Drosophila melanogaster. Proc Natl Acad Sci USA. 2004; 101: 7663 -7668. [PubMed] .
- 20. Parkes TL , Elia AJ , Dickinson D , Hilliker AJ , Phillips JP and Boulianne GL. Extension of Drosophila lifespan by overexpression of human SOD1 in motorneurons. Nat Genet. 1998; 19: 171 -174. [PubMed] .
- 21. Orr WC , Radyuk SN , Prabhudesai L , Toroser D , Benes JJ , Luchak JM , Mockett RJ , Rebrin I , Hubbard JG and Sohal RS. Overexpression of glutamate-cysteine ligase extends lifespan in Drosophila melanogaster. J Biol Chem. 2005; 280: 37331 -37338. [PubMed] .
- 22. Orr WC , Mockett RJ , Benes JJ and Sohal RS. Effects of overexpression of copper-zinc and manganese superoxide dismutases, catalase, and thioredoxin reductase genes on longevity in Drosophila melanogaster. J Biol Chem. 2003; 278: 26418 -26422. [PubMed] .
- 23. Curtis C , Landis GN , Folk D , Wehr NB , Hoe N , Waskar M , Abdueva D , Skvortsov D , Ford D , Luu A , Badrinath A , Levine RL , Bradley TJ , Tavare S and Tower J. Transcriptional profiling of MnSOD-mediated lifespan extension in Drosophila reveals a species-general network of aging and metabolic genes. Genome Biol. 2007; 8: R262 [PubMed] .
- 24. Mair W , Goymer P , Pletchers SD and Partridge L. Demography of dietary restriction and death in Drosophila. Science. 2003; 301: 1731 -1733. [PubMed] .
- 25. Miwa S , Riyahi K , Partridge L and Brand MD. Lack of correlation between mitochondrial reactive oxygen species production and lifespan in Drosophila. Ann N Y Acad Sci. 2004; 1019: 388 -391. [PubMed] .
- 26. Gredilla R , Grief J and Osiewacz HD. . Mitochondrial free radical generation and lifespan control in the fungal aging model Podospora anserina. Exp Gerontol. 2006; 41: 439 -447. [PubMed] .
- 27. Dufour E , Boulay J , Rincheval V and Sainsard-Chanet A. A causal link between respiration and senescence in Podospora anserina. Proc Natl Acad Sci USA. 2000; 97: 4138 -4143. [PubMed] .
- 28. Hakkaart FA , Dassa EP , Jacobs HT and Rustin P. Allotopic expression of a mitochondrial alternative oxidase confers cyanide resistance to human cell respiration. Embo Rep. 2006; 7: 341 -345. [PubMed] .
- 29. Dassa EP , Dufour E , Goncalves S , Paupe V , Hakkaart GAJ , Jacobs HT and Rustin P. Expressing the alternative oxidase complements cytochrome c oxidase deficiency in humans cells. EMBO Mol Med. 2009; 1: 30 -36. [PubMed] .
- 30. Fernandez-Ayala DJM , Sanz A , Vartiainen S , Kemppainen KK , Babusiak M , Mustalahti E , Costa R , Tuomela T , Zeviani M , Chung J , O'Dell KMC , Rustin P and Jacobs HT. Expression of the Ciona intestinalis alternative oxidase (AOX) in Drosophila complements defects in mitochondrial oxidative phosphorylation. Cell Metab. 2009; 9: 449 -460. [PubMed] .
- 31. Miwa S , St-Pierre J , Partridge L and Brand MD. Superoxide and hydrogen peroxide production by Drosophila mitochondria. Free Radic Biol Med. 2003; 35: 938 -948. [PubMed] .
- 32. Sanz A and Barja G. Academic Press New York Estimation of the rate of production of oxygen radicals at mitochondria. In Handbook of models for the study of human aging (M. Conn ed.). 2006; .
- 33. Fridell YW , Sanchez-Blanco A , Silvia BA and Helfand SL. Targeted expression of the human uncoupling protein 2 (hUCP2) to adult neurons extends lifespan in the fly. Cell Metab. 2005; 1: 145 -152. [PubMed] .
- 34. Lambert AJ , Buckingham JA , Boysen HM and Brand MD. Low complex I content explains the low hydrogen peroxide production rate of heart mitochondria from the long-lived pigeon, Columba livia. Aging Cell. 2010; 9: 78 -91. [PubMed] .
- 35. Andres-Mateos E , Perier C , Zhang L , Blanchard-Fillion B , Greco TM , Thomas B , Ko HS , Sasaki M , Ischiropoulos H , Przedborski S , Dawson TM and Dawson VL. DJ-1 gene deletion revels that DJ-1 is an atypical peroxiredoxin-like peroxidase. PNAS. 2008; 104: 14807 -14812. [PubMed] .
- 36. Ku HH and Sohal RS. Comparison of mitochondrial pro-oxidant generation and anti-oxidant defenses between rat and pigeon: possible basis of variation in longevity and metabolic potential. Mech Aging Dev. 1993; 72: 67 -76. [PubMed] .
- 37. Millenaar FF , Benschop JJ , Wagner AM and Lambers H. The role of the alternative oxidase in stabilizing the in vivo reduction state of the ubiquinone pool and the activation state of the alternative oxidase. Plant Physiol. 1998; 118: 599 -607. [PubMed] .
- 38. Ross RE Age-specific decrease in aerobic efficiency associated with increase in oxygen free radical production in Drosophila melanogaster. J Insect Physiol. 2000; 46: 1477 -1480. [PubMed] .
- 39. Melvin RG and Ballard JW. Intraspecific variation in survival and mitochondrial oxidative phosphorylation in wild-caught Drosophila simulans. Aging Cell. 2006; 5: 225 -233. [PubMed] .
- 40. Trifunovic A and Larsson NG. Mitochondrial dysfunction as a cause of aging. J Inter Med. 2008; 263: 167 -178. .
- 41. Walker DW , Hajek P , Muffat J , Knoepfle D , Cornelison S , Attardi G and Benzer S. Hypersensitivity to oxygen and shortened lifespan in a Drosophila mitochondrial complex II mutant. Proc Natl Acad Sci USA. 2006; 103: 16382 -16387. [PubMed] .
- 42. Ishii N , Ishii T and Hartman PS. The role of the electron transport gene SDHC on lifespan and cancer. Exp Gerontol. 2006; 41: 952 -956. [PubMed] .
- 43. Bahadorani S , Cho J , Lo JrT , Contreras H , Lawal HO , Krantz DE , Bradley TJ and Walker DW. Neuronal expression of a single-subunit yeast NADH-ubiquinone oxidoreductase (Ndi1) extends Drosophila lifespan. Aging Cell . 2010; 9: 191 -202. [PubMed] .
- 44. Sanz A , Soikkeli M , Portero-Otin M , Wilson A , Kemppainen E , McIlroy G , Ellilä S , Kemppainen KK , Tuomela T , Lakanmaa M , Kiviranta E , Stefanatos R , Dufour E , Hutz B , Naudi A , Jove M , Zeb A , Vartiainen S , Matsuno-Yagi A , Yagi T , Rustin P , Pamplona R and Jacobs HT. Expression of the yeast NADH dehydrogenase Ndi1 in Drosophila confers increased lifespan independently of dietary restriction. PNAS. 2010; In press .
- 45. Abeliovich A and Beal MF. Parkinsonism genes: culprits and clues. J Neurochemistry. 2006; 99: 1062 -1072. .
- 46. Arking R , Burde V , Graves K , Hari R , Feldman E , Zeevi A , Soliman S , Saraiya A , Buck S , Vettraino J , Sathrasala K , Wehr N and Levine RL. Forward and reverse selection for longevity in Drosophila is characterized by alteration of antioxidant gene expression and oxidative damage patterns. Exp Gerontol. 2000; 35: 167 -185. [PubMed] .
- 47. Barja G The gene cluster hypothesis of aging and longevity. Biogerontology. 2008; 9: 57 -66. [PubMed] .
- 48. Doonan R , McElwee JJ , Matthijssens F , Walker GA , Houthoofd K , Back P , Matscheski A , Vanfleteren JR and Gems D. Against the oxidative damage theory of aging: superoxide dismutase protect against oxidative stress but have little or not effect on lifespan in Caenorhabditis elegans. Genes Dev. 2008; 22: 3236 -3241. [PubMed] .
- 49. Zielonka J and Kalyaraman B. "ROS-generating mitochondrial DNA mutations can regulate tumor cell metastasis "—a critical commentary. Free Radic Biol Med. 2008; 45: 1217 -1219. [PubMed] .
- 50. Lambert AJ and Merry BJ. Effect of caloric restriction on mitochondrial reactive oxygen species production and bioenergetics: reversal by insulin. Am J Physiol Regul Integr Comp Physiol. 2004; 286: R71 -R79. [PubMed] .
- 51. Lambert AJ and Merry BJ. Lack of effect of caloric restriction on bioenergetics and reactive oxygen species production in intact rat hepatocytes. J Gerontol A Biol Sci Med Sci. 2005; 60: 175 -180. [PubMed] .