



Introduction
Senescent cells express myriad inflammatory cytokines and chemokines, a phenomenon termed the senescence-associated secretory phenotype (SASP) [1-3]. The SASP connects cellular senescence with age-related decline in tissue function, and specific SASP proteins act as systemic factors to promote aging. For instance, age-dependent increases in the levels of the chemokine CCL11 [4] and β2-microglobulin [5] impair hippo-campal neurogenesis and cognitive function, and C1q-mediated activation of Wnt signaling impairs muscle repair by inducing dysfunction in skeletal muscle satellite cells [6]. Clearance of senescent cells delays the induction of various geriatric pathologies, supporting the notion that the SASP promotes aging in a non-cell-autonomous fashion [7, 8]. Although very few anti-aging functions of the SASP have been reported, the pathway has been shown to serve as a surveillance system for cellular senescence [1, 9]. For example, oncogene-induced senescent hepatocytes secrete SASP-like inflammatory cytokines that attract attention from immune cells. These pre-malignant senescent cells are usually eliminated by the CD4-positive (CD4+) T cell-mediated adaptive immune response, but impairment of this system by CD4+ T cell deficiency results in the development of hepatocellular carcinoma. However, little is known about the disruption of tissue homeostasis by the cellular SASP and its influence on aging at the organismal level.
Aging of adult tissue stem cells also contributes to loss of tissue homeostasis, e.g., dysregulation of hematopoiesis due to a decline in the lymphopoietic potential of hematopoietic stem/progenitor cells (HSCs) [10, 11]; reduced muscle repair capacity due to diminished activity of skeletal muscle satellite cells [12-14]; and loss of multipotency (i.e., the potential to differentiate into osteogenic, adipogenic, chondrogenic, neurogenic, and myogenic lineages) of mesenchymal stem/stromal cells (MSCs). Previous studies showed that regenerative efficacy is greater in young MSCs than in old MSCs [15-17]. Moreover, the expression of growth factors declines with age [18, 19], and secretory factors exert different effects on MSC function in young vs. old MSCs [16, 20]. These reports suggest that age-dependent changes in secretory properties are induced in MSCs, although to date the MSC SASP remains uncharacterized.
MSCs are a source of various humoral factors, including vascular endothelial growth factor (VEGF), insulin-like growth factor-1 (IGF-1), and hepatocyte growth factor (HGF) [21]. The effects of some of these factors have been determined. For instance, TNF-α-induced protein 6 (TSG-6), an anti-inflammatory protein secreted from transplanted MSCs, exerts a beneficial effect in the mouse model of myocardial infarction [22]. Combined administration of monocyte chemoattractant protein-1 (MCP-1) and the ectodomain of sialic acid-binding Ig-like lectin-9 (Siglec-9), which are both secreted from dental pulp-derived MSCs, induces an anti-inflammatory environment, promoting functional recovery after spinal cord injury [23]. However, the secretory factors responsible for the regenerative effects of transplanted MSCs remain largely unknown.
In this study, we identified growth differentiation factor 6 [Gdf6; also known as bone morphogenetic protein (Bmp)-13 and cartilage-derived morphogenetic protein (CDMP-2)] as a regenerative factor secreted by young MSCs. Administration of Gdf6 restored differentiation potential of aged MSCs in vitro and exerted beneficial effects on some age-associated pathologies in mice.
Results
Characterization of the age-related phenotypes of old MSCs
First, we compared the phenotypes of young (2–3 months) and old (> 18 months) MSCs. To exclude the influence of contaminating differentiated cells, such as lineage-restricted osteoblasts and adipocyte progenitors, we used fluorescence-activated cell sorting (FACS) to isolate MSC fractions (CD45-, CD31-, TER119-, Pdgfra+, and Sca-1+) from young and old murine bone marrow (BM) [24, 25]. We confirmed upregulation of the representative cellular senescence markers p21 (also known as Cdk1na) and senescence-associated β-galactosidase activity (SA-β-gal) (Supplementary Fig. 1). Consistent with previous reports, both differentiation potential and secretory profiles differed between young and old MSCs [19]. Specifically, the osteogenic and adipogenic potential of MSCs declined with age (Fig. 1A). In addition, old MSCs expressed higher levels of inflammatory factors, including representative SASP factors such as interleukin 6, Groa, and Gmcsf, and also exhibited alterations in the expression levels of certain growth factors (Fig. 1B–C).
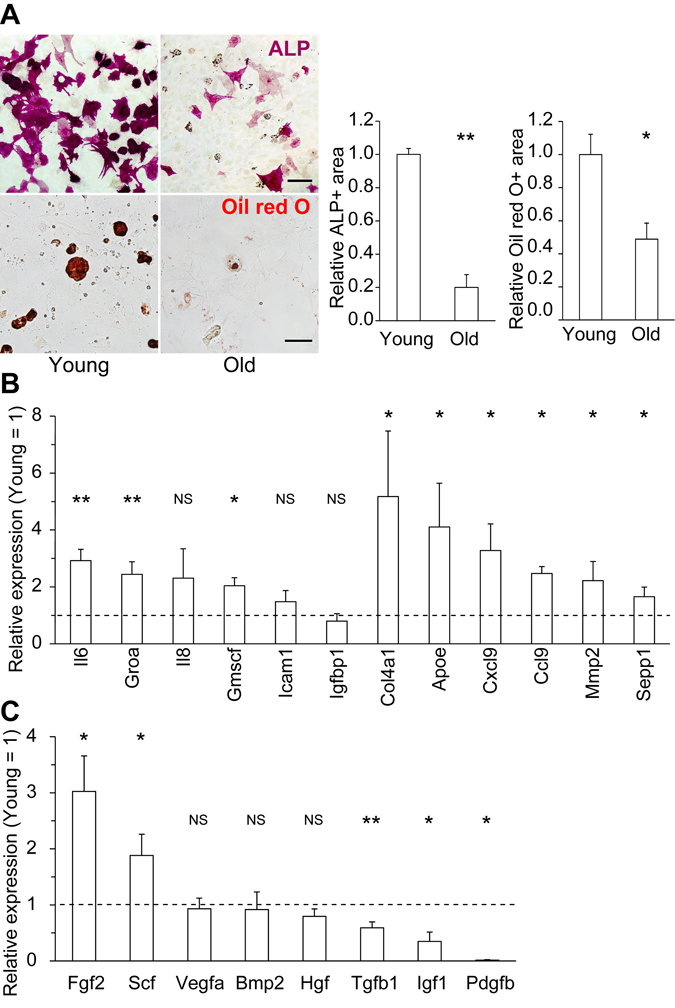
Figure 1. Changes in the differentiation potential and secretory profiles of old MSCs (A) Representative immunocytochemical images of young and old MSCs differentiated into ALP+ osteoblasts (upper) and Oil red O+ fat droplet-producing adipocytes (lower). Both the osteogenic and adipogenic potentials of old MSCs were reduced (n ≥ 3). (B) Expression levels of representative SASP factors (Il6, Groa, Il8, Gmscf, Icam1, and Igfbp1) and major secretory factors in old MSCs, relative to those in young MSCs, determined by qPCR (n ≥ 3). (C) qPCR of representative growth factors (n ≥ 3). Scale bars: 50 μm. Results are expressed as means ± standard error of the mean (SEM). NS (not significant), p > 0.05; *p < 0.05; **p < 0.01.
miR-17 overexpression restores the differentiation potential of old MSCs
To identify genes capable of restoring the regenerative capacity of old MSCs, we compared the gene expression profiles of young and old MSCs using microarrays and microRNA (miRNA) quantitative PCR (qPCR) arrays (Supplementary Tables 1 and 2). These analyses identified several candidate genes that were highly expressed in young MSCs and downregulated with age, including the miR-17 family members miR-17, -18a, -19a/b, -20a/b, -25, -93, -92a, -106a/b, and -363 (Supplementary Table 3). Previously, we showed that miR-17 and its paralogs miR-106a/b are responsible for the neurogenic differentiation potential of neural stem/progenitor cells (NSCs) during early developmental stages, and that overexpression of miR-17 restores the neuropotency of gliogenic NSCs without altering the epigenetic status of glial genes [26, 27].
Although miR-17 is known to exert both pro- and anti-osteogenic effects on osteoblast differentiation of MSCs [28-30], the role of miR-17/106 in regulating the differentiation potential and secretory phenotype of undifferentiated MSCs remains unclear. To address this issue, we transduced old MSCs with miR-17 lentivirus, which integrates the transgene into the genome of the infected cell and continuously overexpresses EGFP and miR-17 under the control of the human elongation factor-1α promoter, and cultured them for 1 week, and retrospectively evaluated differentiation potential by measuring the area that stained positive for alkaline phosphatase (ALP) in osteoblasts and Oil red O in adipocytes. In cells overexpressing miR-17, osteogenic and adipogenic potential was significantly restored (Fig. 2A–B).
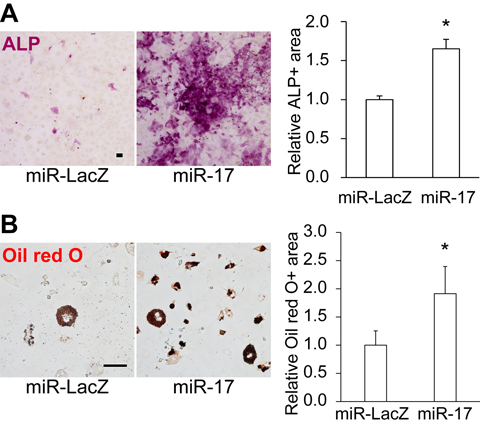
Figure 2. Overexpression of miR-17 restores the differentiation potential of old MSCs Overexpression of miR-17 restored the (A) osteogenic and (B) adipogenic potential of old MSCs (n = 3). Scale bars: 100 μm in A, 50 μm in B. Results are expressed as means ± SEM. *p < 0.05.
Transplantation of miR-17-overexpressing old MSCs reverses the age-related decline in lymphopoiesis
To determine whether miR-17 overexpression could restore the regenerative efficacy of old MSCs in vivo, we intravenously injected miR-17-overexpressing old MSCs into old mice. The transplanted GFP+ cells were detected in the lung, but not in BM (Fig. 3A and Supplementary Fig. 2), consistent with previous studies showing that most intravenously injected cultured MSCs engraft in this organ [22, 31]. Transplantation of miR-17-overexpressing, but not control miR-LacZ-expressing, old MSCs reversed the age-dependent reduction in lymphocyte number in peripheral blood (PB) (Fig. 3B and Supplementary Fig. 3). Because we detected no GFP+ transplanted cells in BM, we hypothesized that lymphopoiesis was rescued by a secretory factor derived from the transplanted MSCs.
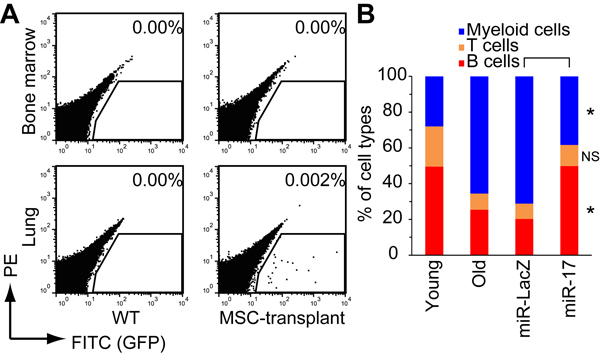
Figure 3. Restoration of lymphopoiesis by transplantation of miR-17-expressing old MSCs (A) EGFP+ transplanted cells engrafted in the lungs, but not in bone marrow. (B) Percentages of PB cell types were analyzed by FACS 16 weeks after transplantation (n = 4). NS, p > 0.05; *p < 0.05.
Gdf6 restores the osteogenic potential of old MSCs
To identify the MSC-derived secretory factors responsible for restoring lymphopoiesis, we used microarrays to compare the gene expression profiles of young, old, and miR-17-overexpressing old MSCs. Genes whose expression was downregulated in old MSCs but restored by miR-17 overexpression were subjected to Gene Ontology analysis, revealing 13 candidate secretory factors (Fig. 4, Supplementary Table 4, and also see Methods).
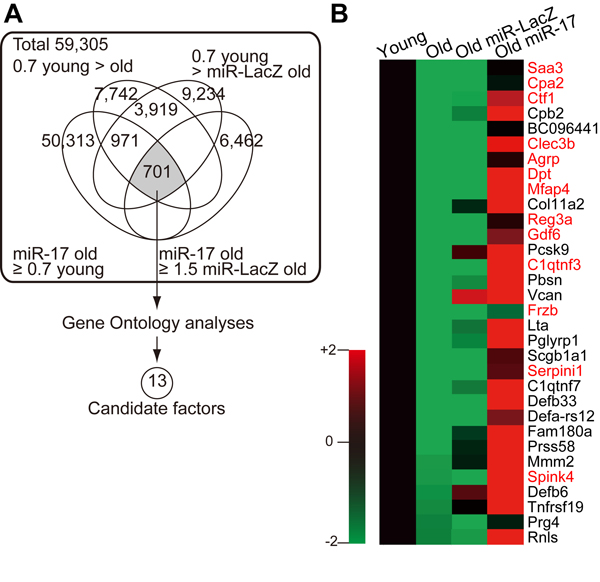
Figure 4. Age- and miR-17-dependent expression of MSC-secreted factors (A) Schematic describing the strategy for identifying candidate factors based on microarray data. (B) Heatmap of the expression levels of secretory factors downregulated in old MSCs. The 13 candidate factors are shown in red.
We functionally analyzed these 13 candidate factors in MSC osteoblast differentiation assays. For this purpose, each factor was expressed in 293T cells, and the conditioned medium (CM) was collected. Differentiation of old MSCs into osteoblasts was induced by osteogenic differentiation media supplemented with or without 50% CM. Among the 13 candidate factors, only Gdf6 promoted osteogenic differentiation in old MSCs (Fig. 5A). Next, we investigated the dependency of Gdf6 expression on miR-17. To this end, we utilized RNA decoys, referred to as “tough decoy” RNAs (TuDs), to efficiently and stably suppress the activity of miR-17 and its paralogs miR-106a/b (Fig. 5B) [26, 32]. Repression of miR-17/106 induced downregulation of Gdf6, suggesting that expression of Gdf6 is regulated by miR-17 in MSCs (Fig. 5C).
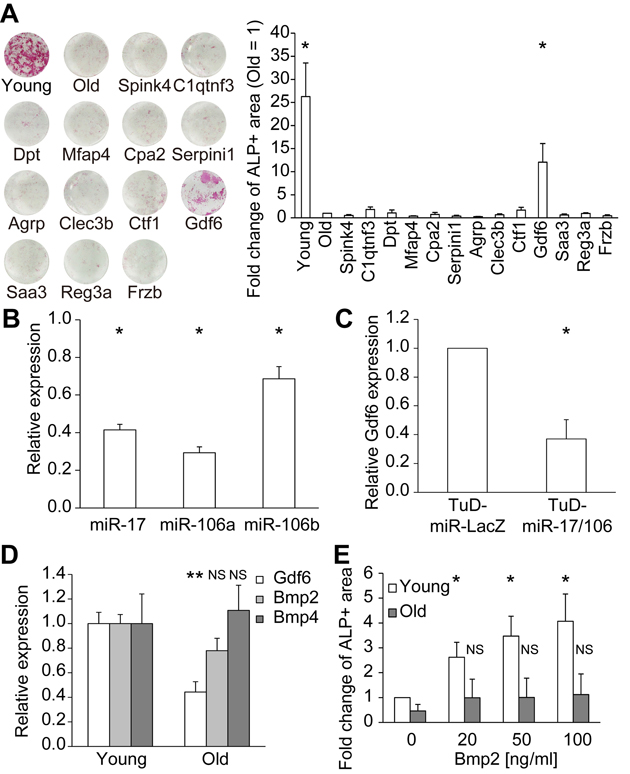
Figure 5. Identification of Gdf6 as an osteogenic factor derived from young MSCs (A) Representative images of ALP staining of differentiated MSCs. ALP activity was quantified as the percentage of ALP+ area and compared with that of old MSCs (n = 4). (B) TuD-miR-17/106 repressed miR-17/106 (n = 3). (C) Repression of miR-17/106 induced downregulation of Gdf6 (n = 3). (D) qPCR of Gdf6 and Bmp2/4 in young and old MSCs (n = 8). (E) Administration of recombinant Bmp2 promoted osteogenic differentiation in young but not old MSCs (n = 4). Results are expressed as means ± SEM. NS, p > 0.05; *p < 0.05.
Gdf6, which belongs to the TGF-β superfamily, is required for proper formation of eyes [33], skull [34], and certain bones and joints in the limbs [35]. Because Gdf6 and Bmp2/4 both bind to the same BMP receptors (ALK3/6 and BMPRII/ActRIIA) and signal through Smad1/5/8 [36], we compared the expression levels of Gdf6 and Bmp2/4 in young and old MSCs. Expression of Gdf6 was downregulated in old MSCs, in concordance with the diminished osteogenic potential of these cells, whereas Bmp2/4 expression remained unchanged over the course of MSC aging (Fig. 5D). Moreover, recombinant human BMP2 promoted osteogenic differentiation in a dose-dependent manner only in young MSCs, whereas BMP2-mediated osteogenic competence was suppressed in old MSCs (Fig. 5E). We confirmed that the expression levels of BMP receptors did not differ between young and old MSCs. They were approximately stable after supplementation with GDF6, BMP2, and a combination of both (Supplementary Fig. 4). There is a report about the divergent activities of BMP2 and Gdf6, despite them having similar biochemical receptor-binding affinities [37]. These results suggest that Gdf6 enhances the osteogenic potential of old MSCs independently of BMP2/4.
Upregulation of Gdf6 ameliorates geriatric pathologies in vivo
Although the results presented above clearly demonstrate that Gdf6 was able to restore the differentiation potential of old MSCs, it remained unclear whether Gdf6 was the critical secretory factor responsible for restoring lymphopoiesis. To clarify this issue, we overexpressed human GDF6 (hGDF6) in 20-month-old mice by intraperitoneal lentivirus injection, and then examined the percentage of each blood cell type in PB. This method proved useful for investigating the long-term in vivo effects of hGDF6. We confirmed that peritoneal cells and subcutaneous adipose tissue, but not PB cells, were mainly infected with lentivirus (Supplementary Fig. 5). These lentivirus-infected cells secreted hGDF6 into blood and upregulated the plasma level at least over 16 weeks (Supplementary Fig. 6). The plasma level of Gdf6 increased with age and was further elevated by overexpression of hGDF6 (Supplementary Fig. 7). As in the MSC transplantation experiments, hGDF6 overexpression restored B- and T-cell populations to the levels observed in young mice (Fig. 6A and Supplementary Fig. 8).
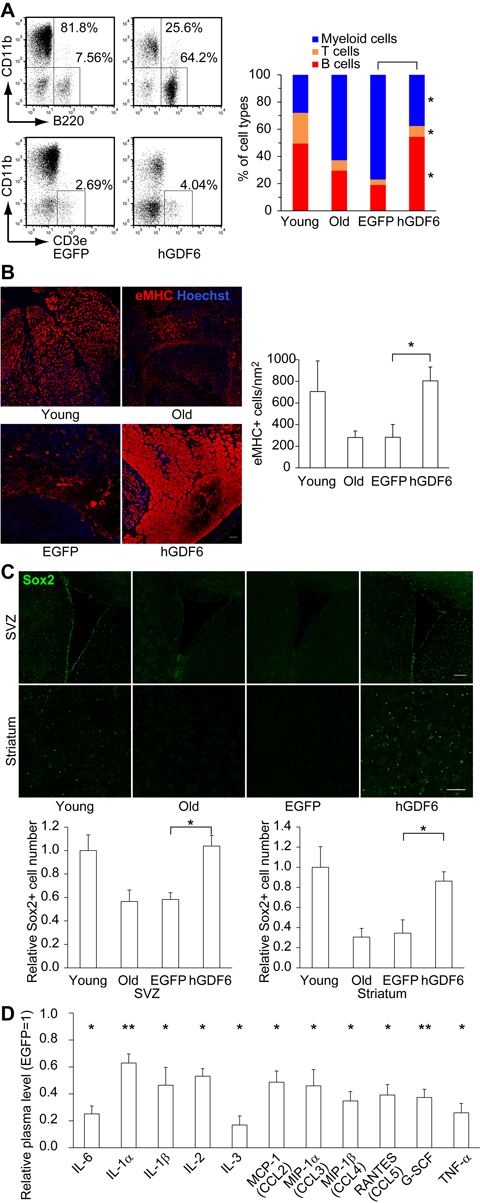
Figure 6. Lentiviral transduction of Gdf6 ameliorated age-related pathologies (A) Representative FACS plots showing the percentages of B- (B220+), T- (CD3e+), and myeloid (CD11b+) cells in PB 16 weeks after transduction. (B) Embryonic myosin heavy chain+ (eMHC+) regenerating myoblasts and myotubes 5 days after CTX injury were examined by immunohistochemistry. Scale bar: 100 μm. (C) The number of Sox2+ neural progenitor cells was elevated in hGDF6-transduced mouse brains. Scale bars: 100 μm. (D) Relative plasma levels of inflammatory factors (n ≥ 4). Results are expressed as means ± SEM. *p < 0.05, **p < 0.01.
In addition, we examined the influence of hGDF6 overexpression on other geriatric pathologies. To investigate the effect on muscle repair capacity, we injured the tibialis anterior muscle by focal cardiotoxin (CTX) injection 5 days before sacrifice, and then monitored the formation of embryonic myosin heavy chain-positive (eMHC+) myoblasts. In addition to mononuclear myoblasts, large numbers of eMHC+ myotubes were observed in hGDF6-overexpressing mice, as demonstrated by an increase in the eMHC+ area (Fig. 6B and Supplementary Fig. 9A). These results suggest that continuous exposure to high doses of hGDF6 accelerates muscle regeneration. Moreover, in hGDF6-overexpressing old mice, Sox2-positive (Sox2+) neural progenitors were distributed in a wide brain region that included the subventricular zone (SVZ) and striatum concomitant with improvements in the cerebrovascular network; by contrast, relatively few Sox2+ cells were detected in the brains of old control mice (Fig. 6C and Supplementary Figs. 9B–C, 10). The number of Dcx+ neuroblasts was also increased in hGDF6-overexpressing old mouse brain (Supplementary Fig. 11). We also observed broad expression of Gdf6 both in young and old mouse brains, whereas little is known about its physiological functions in the brain (Supplementary Fig. 12). Furthermore, the plasma levels of various inflammatory factors were reduced in hGDF6-overexpressing mice (Fig. 6D). Taken together, these results indicate that Gdf6 is a regenerative factor secreted from MSCs.
Discussion
Studies of age-dependent changes in secretory profiles have focused on the upregulation of diverse inflammatory factors, collectively termed the SASP. However, many secretory factors, including homeostatic growth factors, are downregulated with age. In this study, we discovered that old MSCs also exhibit age-related changes in secretory profiles, and identified Gdf6 as a regenerative factor whose expression declines with age. These results suggest that both up- and downregulation of secretory factors contribute to the development of age-related cellular dysfunction.
The major inducer of SASP is the NF-κB signaling pathway, which is activated by both genotoxic and environmental stress [38]. However, the regulatory mechanisms underlying the induction of the SASP remain largely obscure. Recently, GATA4 was identified as a novel activator of NF-κ B [39]. Selective autophagy of GATA4 is inhibited by the DNA damage response, leading to NF-κ B-mediated activation of various inflammatory factors. Likewise, it remains unclear how homeostatic factors are downregulated during senescence. Here, we showed that a subset of secretory factors are regulated by miR-17, which is also downregulated with age, suggesting that changes in the expression levels of certain miRNAs play important roles in age-related changes in secretory profiles, and also that modulation of miRNA expression enables control of the SASP.
GDF11, another member of the TGF-β superfamily, was recently identified as a regenerative factor that ameliorates age-related cardiac hypertrophy [40], slows vascular deterioration, and promotes brain neurogenesis [41] and skeletal muscle repair [42]. Despite the fact that the two factors signal via different BMP receptors and SMAD pathways [43], the regenerative effects of GDF6 observed in this study were similar to those reported previously for GDF11. Elucidation of the mechanisms underlying their effects on geriatric disorders in various tissues should improve our understanding of the aging at the organismal level.
Mutations in the GDF6 gene lead to the Klippel-Feil syndrome [44], microphthalmia, anophthalmia [33], and age-related macular degeneration (AMD) [45]. Formation of aberrant vasculature induced by GDF6 mutations has been proposed as a cause of AMD and anomalies in ocular development [33, 45, 46]. The promotion of neurogenesis and neuronal activity induced by administration of GDF11 is presumably mediated by an increase in blood flow in the brain [41]. Consistent with this, we observed improvements in the cerebrovascular network in hGDF6- overexpressing mice. These observations suggest that regeneration of vascular networks is one mechanism underlying the regenerative effects of GDF6 and GDF11 in various tissues. Smad1/5-mediated signaling functions as an activator of angiogenesis, whereas Smad2/3 signaling has a repressor effect [47], indicating that GDF6 is more suitable than GDF11 for regeneration mediated by vascular remodeling. Indeed, GDF11 can also induce the opposite effect. The active form of the GDF11 protein, which is a TGF-β family member, is processed to form biologically active carboxy-terminal dimers whose levels increase with age, inhibiting muscle regeneration [48]. Similarly, Smad1/5 signaling can inhibit endothelial cell functions [49, 50]. Moreover, context-dependent modifications of BMP/TGF-β signals by diverse cofactors further complicate the pleiotropic functions of these proteins [51-53].
Chronic inflammation accelerates systemic aging [54]. In this study, we found that plasma levels of various inflammatory factors were downregulated in the hGDF6-overexpressing old mice. Attenuation of chronic inflammation might reduce the severity of geriatric disorders or prevent them altogether. Therefore, an understanding of the mechanisms underlying the regenerative effects of GDF6 in various tissues could lead to the development of novel therapeutic agents for the treatment of age-related disorders.
Methods
Cell preparation and culture
Primary MSCs (CD45−, CD31−, TER119−, Pdgfra+, and Sca-1+) were isolated from young (2–3 months) and old (> 18 months) mice and cultured as previously described [24, 25], except that the ilium was used in addition to the femur and tibia. Functional screening of the 13 secretory factors was performed using conventional BM-derived adherent stromal cells without cell sorting.
MSC differentiation assay
Induction of osteoblast and adipocyte differentiation was performed using the Human Mesenchymal Stem Cell Osteogenic/Adipogenic Differentiation BulletKit Medium (Lonza). Differentiated osteoblasts were stained with a Leukocyte Alkaline Phosphatase Kit (Sigma-Aldrich), and adipocyte-produced fat droplets were detected by Oil red O staining. The rate of differentiation of MSCs was evaluated using the Hybrid Cell Count software of the BZ-X700 microscope (KEYENCE) and ImageJ (http://imagej.nih.gov/ij/).
Lentivirus preparation
Lentiviruses were produced by transient transfection of human embryonic kidney 293T cells with lentiviral constructs (provided by Dr. H. Miyoshi, Keio University, and RIKEN BRC) containing the gene of interest. Sequences of TuDs were described in a previous study [26].
Mice
Young (2–3 months) and old (> 18 months) C57BL/6JJcl mice were used. Animal care and experiments were performed according to the guidelines of the Experimental Animal Care Committee of the Yokohama Safety Center of RIKEN.
Multiplex cytokine analysis
Plasma levels of cytokines were quantitated using the Luminex system (Bio-Rad).
Microarray and miRNA qPCR array
Total RNA was purified from primary MSCs or MSCs cultured for infection with lentiviruses. Gene and miRNA expression profiles were investigated using microarrays (Agilent, SurePrint G3 Mouse GE microarray) and miRNA qPCR arrays (Thermo Fisher Scientific Applied Biosystems, TaqMan Array Rodent MicroRNA A+B Cards Set v3.0).
Transplantation of MSCs
MSCs isolated from young and old mice were lentivirally transduced and intravenously transplanted into old mice as previously described [15]. The percentages of PB cell types (B220+ B cells, CD3ε + T cells, and CD11b+ myeloid cells) were analyzed by FACS using specific antibodies (BioLegend). After sacrifice, cells were prepared from BM, lungs, liver, and spleen using dispase/collagenase solution, and then analyzed by FACS to monitor MSC engraftment.
Identification of Gdf6
Of the 59,305 probes screened by the microarray analyses, the expression levels of 701 matched four criteria: (1) <0.7-fold lower in old than in young MSCs, (2) <0.7-fold lower in miR-LacZ-overexpressing old than in young MSCs, (3) >0.7-fold higher in miR-17-expressing old than in young MSCs, and (4) >1.5-fold higher in miR-17-expressing old than in miR-LacZ-overexpressing old MSCs. These candidate genes were then further restricted to those encoding secreted proteins using Gene Ontology annotation in the DAVID online software (http://david.abcc.ncifcrf.gov), decreasing the number of candidates to 17. Thirteen of these candidate genes could be cloned from a cDNA library prepared from young MSCs and validated as miR-17-dependent, young MSC-derived secretory factors. All 13 factors were functionally evaluated by MSC osteoblast differentiation assay with CM from 293T cells expressing each factor.
In vivo upregulation of hGDF6
Old mice were transduced with hGDF6 lentivirus by intraperitoneal injection (3×) every other day. Plasma levels of hGDF6 were investigated by western blotting with anti-GDF6 antibody (Sigma-Aldrich, PRS4691). To analyze the precursor form of hGDF6, blood plasma was purified using the Aurum Serum Protein Mini Kit (Bio-Rad, 7326701). Sustained upregulation of plasma levels of the active form of hGDF6 was confirmed for at least 16 weeks (Supplementary Fig. 3).
Muscle injury
Injury was induced by focal injection of CTX in the left tibialis anterior muscle, as previously described [55]. Five days after injury, samples were obtained from the lower leg, and paraffin-embedded tissue sections were prepared for immunohistochemistry.
Immunostaining
Immunohistochemistry was performed on 2 μm paraffin-embedded histological sections using antibodies against eMHC (MYH3; 1:50, Santa Cruz Biotechnology sc-53091) [56] and Sox2 (1:100, Abcam ab92494). Immunostained histological sections were quantitatively evaluated using the Hybrid Cell Count program of the BZ-X700 microscope (KEYENCE; Supplementary Fig. 5).
Statistical analyses
At least three independent experiments were included in each statistical analysis. Statistical significance was determined by t-test. P-values <0.05 were considered statistically significant.
Supplementary Materials
Author Contributions
H.N.-K. devised and planned the project. D.H., M.O.-O., and H.N.-K. performed experiments and analyzed and interpreted data. Y.M. supervised the MSC experiments. S.N. assisted with all experiments. H.N.-K. wrote the manuscript and supervised the entire project.
Acknowledgments
We thank Dr. H. Miyoshi (Keio University) for the lentiviral constructs and Dr. H. Kanki for technical advice and critical reading of the manuscript.
Funding
This study was supported by the Young Chief Investigator Program at RIKEN (H.N.-K.) and by MEXT KAKENHI Grant Numbers 26860200 and 24689017 (M.O.-O. and H.N.-K.).
Conflicts of Interest
The authors have no conflict of interests to declare.
References
- 1. Kuilman T, Michaloglou C, Vredeveld LC, Douma S, van Doorn R, Desmet CJ, Aarden LA, Mooi WJ, Peeper DS. Oncogene-induced senescence relayed by an interleukin-dependent inflammatory network. Cell. 2008; 133: 1019 -31. [PubMed] .
- 2. Coppe JP, Patil CK, Rodier F, Sun Y, Munoz DP, Goldstein J, Nelson PS, Desprez PY, Campisi J. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 2008; 6: 2853 -68. [PubMed] .
- 3. Acosta JC, O'Loghlen A, Banito A, Guijarro MV, Augert A, Raguz S, Fumagalli M, Da Costa M, Brown C, Popov N, Takatsu Y, Melamed J, d'Adda di Fagagna F, et al. Chemokine signaling via the CXCR2 receptor reinforces senescence. Cell. 2008; 133: 1006 -18. [PubMed] .
- 4. Villeda SA, Luo J, Mosher KI, Zou B, Britschgi M, Bieri G, Stan TM, Fainberg N, Ding Z, Eggel A, Lucin KM, Czirr E, Park JS, et al. The ageing systemic milieu negatively regulates neurogenesis and cognitive function. Nature. 2011; 477: 90 -94. [PubMed] .
- 5. Smith LK, He Y, Park JS, Bieri G, Snethlage CE, Lin K, Gontier G, Wabl R, Plambeck KE, Udeochu J, Wheatley EG, Bouchard J, Eggel A, et al. beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis. Nat Med. 2015; 21: 932 -37. [PubMed] .
- 6. Naito AT, Sumida T, Nomura S, Liu ML, Higo T, Nakagawa A, Okada K, Sakai T, Hashimoto A, Hara Y, Shimizu I, Zhu W, Toko H, et al. Complement C1q activates canonical Wnt signaling and promotes aging-related phenotypes. Cell. 2012; 149: 1298 -1313. [PubMed] .
- 7. Baker DJ, Wijshake T, Tchkonia T, LeBrasseur NK, Childs BG, van de Sluis B, Kirkland JL, van Deursen JM. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature. 2011; 479: 232 -36. [PubMed] .
- 8. Baker DJ, Childs BG, Durik M, Wijers ME, Sieben CJ, Zhong J, R AS, Jeganathan KB, Verzosa GC, Pezeshki A, Khazaie K, Miller JD, van Deursen JM. Naturally occurring p16-positive cells shorten healthy lifespan. Nature. 2016; 530: 184 -89. [PubMed] .
- 9. Kang TW, Yevsa T, Woller N, Hoenicke L, Wuestefeld T, Dauch D, Hohmeyer A, Gereke M, Rudalska R, Potapova A, Iken M, Vucur M, Weiss S, et al. Senescence surveillance of pre-malignant hepatocytes limits liver cancer development. Nature. 2011; 479: 547 -51. [PubMed] .
- 10. Rossi DJ, Bryder D, Zahn JM, Ahlenius H, Sonu R, Wagers AJ, Weissman IL. Cell intrinsic alterations underlie hematopoietic stem cell aging. Proc Natl Acad Sci U S A. 2005; 102: 9194 -99. [PubMed] .
- 11. Sudo K, Ema H, Morita Y, Nakauchi H. Age-associated characteristics of murine hematopoietic stem cells. J Exp Med. 2000; 192: 1273 -80. [PubMed] .
- 12. Renault V, Thornell LE, Eriksson PO, Butler-Browne G, Mouly V. Regenerative potential of human skeletal muscle during aging. Aging Cell. 2002; 1: 132 -39. [PubMed] .
- 13. Brack AS, Conboy MJ, Roy S, Lee M, Kuo CJ, Keller C, Ro TA. Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science. 2007; 317: 807 -10. [PubMed] .
- 14. Sousa-Victor P, Gutarra S, Garcia-Prat L, Rodriguez-Ubreva J, Ortet L, Ruiz-Bonilla V, Jardi M, Ballestar E, Gonzalez S, Serrano AL, Perdiguero E, Munoz-Canoves P. Geriatric muscle stem cells switch reversible quiescence into senescence. Nature. 2014; 506: 316 -21. [PubMed] .
- 15. Shen J, Tsai YT, Dimarco NM, Long MA, Sun X, Tang L. Transplantation of mesenchymal stem cells from young donors delays aging in mice. Sci Rep. 2011; 1: 67 [PubMed] .
- 16. Sun Y, Li W, Lu Z, Chen R, Ling J, Ran Q, Jilka RL, Chen XD. Rescuing replication and osteogenesis of aged mesenchymal stem cells by exposure to a young extracellular matrix. FASEB J. 2011; 25: 1474 -85. [PubMed] .
- 17. Wang YQ, Wang M, Zhang P, Song JJ, Li YP, Hou SH, Huang CX. Effect of transplanted mesenchymal stem cells from rats of different ages on the improvement of heart function after acute myocardial infarction. Chin Med J (Engl). 2008; 121: 2290 -98. [PubMed] .
- 18. D'Ippolito G, Schiller PC, Ricordi C, Roos BA, Howard GA. Age-related osteogenic potential of mesenchymal stromal stem cells from human vertebral bone marrow. J Bone Miner Res. 1999; 14: 1115 -22. [PubMed] .
- 19. Wilson A, Shehadeh LA, Yu H, Webster KA. Age-related molecular genetic changes of murine bone marrow mesenchymal stem cells. BMC Genomics. 2010; 11: 229 [PubMed] .
- 20. Moerman EJ, Teng K, Lipschitz DA, Lecka-Czernik B. Aging activates adipogenic and suppresses osteogenic programs in mesenchymal marrow stroma/stem cells: the role of PPAR-gamma2 transcription factor and TGF-beta/BMP signaling pathways. Aging Cell. 2004; 3: 379 -89. [PubMed] .
- 21. Chen L, Tredget EE, Wu PY, Wu Y. Paracrine factors of mesenchymal stem cells recruit macrophages and endothelial lineage cells and enhance wound healing. PLoS One. 2008; 3: 1886 .
- 22. Lee RH, Pulin AA, Seo MJ, Kota DJ, Ylostalo J, Larson BL, Semprun-Prieto L, Delafontaine P, Prockop DJ. Intravenous hMSCs improve myocardial infarction in mice because cells embolized in lung are activated to secrete the anti-inflammatory protein TSG-6. Cell Stem Cell. 2009; 5: 54 -63. [PubMed] .
- 23. Matsubara K, Matsushita Y, Sakai K, Kano F, Kondo M, Noda M, Hashimoto N, Imagama S, Ishiguro N, Suzumura A, Ueda M, Furukawa K, Yamamoto A. Secreted ectodomain of sialic acid-binding Ig-like lectin-9 and monocyte chemoattractant protein-1 promote recovery after rat spinal cord injury by altering macrophage polarity. J Neurosci. 2015; 35: 2452 -64. [PubMed] .
- 24. Morikawa S, Mabuchi Y, Kubota Y, Nagai Y, Niibe K, Hiratsu E, Suzuki S, Miyauchi-Hara C, Nagoshi N, Sunabori T, Shimmura S, Miyawaki A, Nakagawa T, et al. Prospective identification, isolation, and systemic transplantation of multipotent mesenchymal stem cells in murine bone marrow. J Exp Med. 2009; 206: 2483 -96. [PubMed] .
- 25. Houlihan DD, Mabuchi Y, Morikawa S, Niibe K, Araki D, Suzuki S, Okano H, Matsuzaki Y. Isolation of mouse mesenchymal stem cells on the basis of expression of Sca-1 and PDGFR-alpha. Nat Protoc. 2012; 7: 2103 -11. [PubMed] .
- 26. Naka-Kaneda H, Nakamura S, Igarashi M, Aoi H, Kanki H, Tsuyama J, Tsutsumi S, Aburatani H, Shimazaki T, Okano H. The miR-17/106-p38 axis is a key regulator of the neurogenic-to-gliogenic transition in developing neural stem/progenitor cells. Proc Natl Acad Sci U S A. 2014; 111: 1604 -09. [PubMed] .
- 27. Naka-Kaneda H, Shimazaki T, Okano H. MicroRNA-mediated regulation of the neurogenic-to-gliogenic competence transition of neural stem/progenitor cells. Neurogenesis. 2014; 1: 1 -4. .
- 28. Liu Y, Liu W, Hu C, Xue Z, Wang G, Ding B, Luo H, Tang L, Kong X, Chen X, Liu N, Ding Y, Jin Y. MiR-17 Modulates Osteogenic Differentiation Through a Coherent Feed-Forward Loop in Mesenchymal Stem Cells Isolated from Periodontal Ligaments of Patients with Periodontitis. Stem Cells. 2011; 29: 1804 -16. [PubMed] .
- 29. Jia J, Feng X, Xu W, Yang S, Zhang Q, Liu X, Feng Y, Dai Z. MiR-17-5p modulates osteoblastic differentiation and cell proliferation by targeting SMAD7 in non-traumatic osteonecrosis. Exp Mol Med. 2014; 46: 107 .
- 30. Liu W, Qi M, Konermann A, Zhang L, Jin F, Jin Y. The p53/miR-17/Smurf1 pathway mediates skeletal deformities in an age-related model via inhibiting the function of mesenchymal stem cells. Aging (Albany NY). 2015; 7: 205 -18. [PubMed] .
- 31. Mabuchi Y, Morikawa S, Harada S, Niibe K, Suzuki S, Renault-Mihara F, Houlihan DD, Akazawa C, Okano H, Matsuzaki Y. LNGFR(+)THY-1(+)VCAM-1(hi+) cells reveal functionally distinct subpopulations in mesenchymal stem cells. Stem Cell Reports. 2013; 1: 152 -65. [PubMed] .
- 32. Haraguchi T, Ozaki Y, Iba H. Vectors expressing efficient RNA decoys achieve the long-term suppression of specific microRNA activity in mammalian cells. Nucleic Acids Res. 2009; 37: 43 .
- 33. Asai-Coakwell M, French CR, Berry KM, Ye M, Koss R, Somerville M, Mueller R, van Heyningen V, Waskiewicz AJ, Lehmann OJ. GDF6, a novel locus for a spectrum of ocular developmental anomalies. Am J Hum Genet. 2007; 80: 306 -15. [PubMed] .
- 34. Clendenning DE and Mortlock DP. The BMP ligand Gdf6 prevents differentiation of coronal suture mesenchyme in early cranial development. PLoS One. 2012; 7: 36789 .
- 35. Settle SH Jr., Rountree RB, Sinha A, Thacker A, Higgins K, Kingsley DM. Multiple joint and skeletal patterning defects caused by single and double mutations in the mouse Gdf6 and Gdf5 genes. Dev Biol. 2003; 254: 116 -30. [PubMed] .
- 36. Mazerbourg S, Sangkuhl K, Luo CW, Sudo S, Klein C, Hsueh AJ. Identification of receptors and signaling pathways for orphan bone morphogenetic protein/growth differentiation factor ligands based on genomic analyses. J Biol Chem. 2005; 280: 32122 -32. [PubMed] .
- 37. Berasi SP, Varadarajan U, Archambault J, Cain M, Souza TA, Abouzeid A, Li J, Brown CT, Dorner AJ, Seeherman HJ, Jelinsky SA. Divergent activities of osteogenic BMP2, and tenogenic BMP12 and BMP13 independent of receptor binding affinities. Growth Factors. 2011; 29: 128 -39. [PubMed] .
- 38. Freund A, Patil CK, Campisi J. p38MAPK is a novel DNA damage response-independent regulator of the senescence-associated secretory phenotype. EMBO J. 2011; 30: 1536 -48. [PubMed] .
- 39. Kang C, Xu Q, Martin TD, Li MZ, Demaria M, Aron L, Lu T, Yankner BA, Campisi J, Elledge SJ. The DNA damage response induces inflammation and senescence by inhibiting autophagy of GATA4. Science. 2015; 349: 5612 .
- 40. Loffredo FS, Steinhauser ML, Jay SM, Gannon J, Pancoast JR, Yalamanchi P, Sinha M, Dall'Osso C, Khong D, Shadrach JL, Miller CM, Singer BS, Stewart A, et al. Growth differentiation factor 11 is a circulating factor that reverses age-related cardiac hypertrophy. Cell. 2013; 153: 828 -39. [PubMed] .
- 41. Katsimpardi L, Litterman NK, Schein PA, Miller CM, Loffredo FS, Wojtkiewicz GR, Chen JW, Lee RT, Wagers AJ, Rubin LL. Vascular and neurogenic rejuvenation of the aging mouse brain by young systemic factors. Science. 2014; 344: 630 -34. [PubMed] .
- 42. Sinha M, Jang YC, Oh J, Khong D, Wu EY, Manohar R, Miller C, Regalado SG, Loffredo FS, Pancoast JR, Hirshman MF, Lebowitz J, Shadrach JL, et al. Restoring systemic GDF11 levels reverses age-related dysfunction in mouse skeletal muscle. Science. 2014; 344: 649 -52. [PubMed] .
- 43. Mazerbourg S and Hsueh AJ. Genomic analyses facilitate identification of receptors and signalling pathways for growth differentiation factor 9 and related orphan bone morphogenetic protein/growth differentiation factor ligands. Hum Reprod Update. 2006; 12: 373 -383. [PubMed] .
- 44. Tassabehji M, Fang ZM, Hilton EN, McGaughran J, Zhao Z, de Bock CE, Howard E, Malass M, Donnai D, Diwan A, Manson FD, Murrell D, Clarke RA. Mutations in GDF6 are associated with vertebral segmentation defects in Klippel-Feil syndrome. Hum Mutat. 2008; 29: 1017 -27. [PubMed] .
- 45. Zhang L, Lim SL, Du H, Zhang M, Kozak I, Hannum G, Wang X, Ouyang H, Hughes G, Zhao L, Zhu X, Lee C, Su Z, et al. High temperature requirement factor A1 (HTRA1) gene regulates angiogenesis through transforming growth factor-beta family member growth differentiation factor 6. J Biol Chem. 2012; 287: 1520 -26. [PubMed] .
- 46. Hall CJ, Flores MV, Davidson AJ, Crosier KE, Crosier PS. Radar is required for the establishment of vascular integrity in the zebrafish. Dev Biol. 2002; 251: 105 -17. [PubMed] .
- 47. Goumans MJ, Valdimarsdottir G, Itoh S, Rosendahl A, Sideras P, ten Dijke P. Balancing the activation state of the endothelium via two distinct TGF-beta type I receptors. EMBO J. 2002; 21: 1743 -53. [PubMed] .
- 48. Egerman MA, Cadena SM, Gilbert JA, Meyer A, Nelson HN, Swalley SE, Mallozzi C, Jacobi C, Jennings LL, Clay I, Laurent G, Ma S, Brachat S, et al. GDF11 Increases with Age and Inhibits Skeletal Muscle Regeneration. Cell Metab. 2015; 22: 164 -74. [PubMed] .
- 49. David L, Mallet C, Mazerbourg S, Feige JJ, Bailly S. Identification of BMP9 and BMP10 as functional activators of the orphan activin receptor-like kinase 1 (ALK1) in endothelial cells. Blood. 2007; 109: 1953 -61. [PubMed] .
- 50. Scharpfenecker M, van Dinther M, Liu Z, van Bezooijen RL, Zhao Q, Pukac L, Lowik CW, ten Dijke P. BMP-9 signals via ALK1 and inhibits bFGF-induced endothelial cell proliferation and VEGF-stimulated angiogenesis. J Cell Sci. 2007; 120: 964 -72. [PubMed] .
- 51. Rudini N, Felici A, Giampietro C, Lampugnani M, Corada M, Swirsding K, Garre M, Liebner S, Letarte M, ten Dijke P, Dejana E. VE-cadherin is a critical endothelial regulator of TGF-beta signalling. EMBO J. 2008; 27: 993 -1004. [PubMed] .
- 52. Santibanez JF, Blanco FJ, Garrido-Martin EM, Sanz-Rodriguez F, del Pozo MA, Bernabeu C. Caveolin-1 interacts and cooperates with the transforming growth factor-beta type I receptor ALK1 in endothelial caveolae. Cardiovasc Res. 2008; 77: 791 -99. [PubMed] .
- 53. Lebrin F, Goumans MJ, Jonker L, Carvalho RL, Valdimarsdottir G, Thorikay M, Mummery C, Arthur HM, ten Dijke P. Endoglin promotes endothelial cell proliferation and TGF-beta/ALK1 signal transduction. EMBO J. 2004; 23: 4018 -28. [PubMed] .
- 54. Jurk D, Wilson C, Passos JF, Oakley F, Correia-Melo C, Greaves L, Saretzki G, Fox C, Lawless C, Anderson R, Hewitt G, Pender SL, Fullard N, et al. Chronic inflammation induces telomere dysfunction and accelerates ageing in mice. Nat Commun. 2014; 2: 4172 [PubMed] .
- 55. Elabd C, Cousin W, Upadhyayula P, Chen RY, Chooljian MS, Li J, Kung S, Jiang KP, Conboy IM. Oxytocin is an age-specific circulating hormone that is necessary for muscle maintenance and regeneration. Nat Commun. 2014; 5: 4082 [PubMed] .
- 56. Tran TH, Shi X, Zaia J, Ai X. Heparan sulfate 6-O-endosulfatases (Sulfs) coordinate the Wnt signaling pathways to regulate myoblast fusion during skeletal muscle regeneration. J Biol Chem. 2012; 287: 32651 -64. [PubMed] .