




Introduction
Paraquat is an herbicide used worldwide which has a strong toxic effect on humans [1]. Paraquat could be absorbed via skin and cause severe damage in multiple organs, including respiratory tract and the digestive tract. The high concentration of paraquat in lung tissue may lead to acute lung injury (ALI) such as progressive dyspnea, monocyte infiltration, alveolar stroma thickening and acute pulmonary edema [2]. Therefore, studying the mechanism of paraquat-induced ALI is of great significance for clinical treatment.
Adhesion G protein-coupled receptor F5 (ADGRF5, also known as Gpr116) belongs to the adhesion class of G protein-coupled receptors (GPCRs), but is unique from others because of comprising immunoglobulin (Ig)-like repeats and long extracellular domains [3]. ADGRF5 plays a critical role in lung surfactant homeostasis through regulating pulmonary surfactant-associated protein B (SFTPB) and SFPTD [4]. Alveolar surfactants have the effect of reducing the surface tension of the alveoli, which exhibit ability to maintain the relative stability of the alveolar volume and prevent the fluid in the alveolar capillaries from filtering out into the alveoli [5]. However, the potential molecular mechanisms by which paraquat lead to ALI and downregulate the expression of pulmonary surfactants still remain unexplored.
Histones, mainly comprising histone H2A, H2B, H3 and H4, are known as parts of the nucleosomes. Within a nucleosome, histones are surrounded by 147 bp of DNA, where the DNA fragments are fixed to the histones at 14 sites [6]. The protruding tails of histones consist of highly dynamic basic amino acids such as lysine and arginine, which could be modified by the addition of methyl, acetyl and phosphoryl groups, indicating that the transcription or expression of genes could be affected by the modified histones [7]. Among them, the trimethylation of lysine 20 at histone H4 (H4K20me3) has been widely studied. Several studies found that the H4K20me3 was correlated with the suppression of genes and loss of H4K20me3, which would result in preneoplasia of lung cancer [8, 9]. Retinoblastoma-like protein 1 (RBL1) could interact with histone-lysine N-methyltransferase KMT5B (also known as SUV420H1) and regulate the trimethylation of histone H4K20 [10]. The RBL1 could also regulate the expression of DNA-binding transcription factor E2F3 for further regulation of other genes involved in individual cellular processes [11]. However, whether paraquat could affect the RBL1/KMT5B/E2F3 pathway has not been reported.
In the present study, we constructed paraquat-induced ALI mice model to explore the change in the histological and inflammatory status of lung tissues. Whether the H4K20me3 participated in the protection of paraquat-induced ALI was investigated via transfected with mutated histone H4 plasmid. The expression of ADGRF5, RBL1, KMT5B, SFTPD and SFTPB was detected via western blotting, and whether E2F3 was binding to the promoter of ADGRF5 was determined via chromatin immunoprecipitation analysis. Briefly, this study focused on mining the role of signaling pathway RBL1/KMT5B/E2F3 in affecting the H4K20me3 and the expression of ADGRF5 in the protection of paraquat-induced ALI, which was significant for discovery of clinical treatments for ALI.
Results
Activation of KMT5B attenuated paraquat-induced ALI in mice
Pathological characteristics and morphological status in the lung of mice could be observed in H&E staining images. As shown in Figure 1, ALI group presented extensive morphological damage including inflammatory infiltration, interstitial edema, alveolar hemorrhage and alveolar thickening while no pathological changes were observed in NC group. After treatment of KMT5B activator, the paraquat-induced pathological damages was dramatically attenuated in the lung tissue of mice. However, the injection of KMT5B inhibitor or transfection with H4K20-mutated plasmid further promoted the pathological damages. These evidences indicated that the H4K20 trimethylation might play a positive role in the protection of lung damage induced by paraquat.
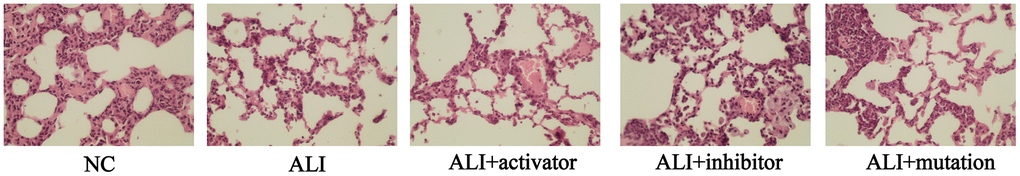
Figure 1. The histological status in lung tissue of mice in NC, ALI, ALI+activator, ALI+inhibitor and ALI+mutation group observed by H&E staining.
Paraquat downregulated the expression of ADGRF5 in mice
Immunohistochemical analysis was performed to determine the relative expression of ADGRF5 in lung tissue of mice. As Figure 2 illustrated, the level of ADGRF5 was the highest in NC group. The treatment of paraquat markedly downregulated the expression of ADGRF5, while activation of KMT5B could increase its level, which was still lower than the normal level in NC group. The level of ADGRF5 in ALI mice injected with KMT5B inhibitor or transfected with H4K20-mutated plasmid was reduced than ALI group, indicating that the unmethylated H4K20 would decrease the ADGRF5 expression.
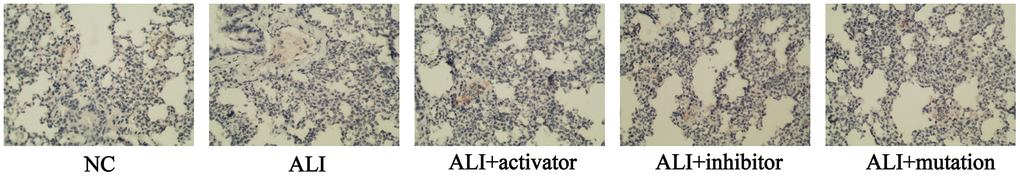
Figure 2. The expression of ADGRF5 in lung tissue of mice in NC, ALI, ALI+activator, ALI+inhibitor and ALI+mutation group detected by immunohistochemical analysis.
Activation of KMT5B decreased the level of inflammatory factors in ALI mice
The level of proinflammatory cytokines detected by ELISA analysis could reflect the inflammation status. Compared with NC group, the expression of IL-1β, IL-3, TNF-α, CCL4 and GM-CSF were all significantly decreased in mice those were orally administrated with paraquat (Figure 3). The upregulation of KMT5B using activator could downregulate the expression of these inflammatory factors in the lung tissue of ALI mice, which was unable to recover to the normal level. The inhibition of KMT5B or mutation of H4K20 further significantly enhanced the level of proinflammatory cytokines. The results suggested that the H4K20me3 could remarkably improve the inflammation status induced by paraquat in lung tissue of mice.
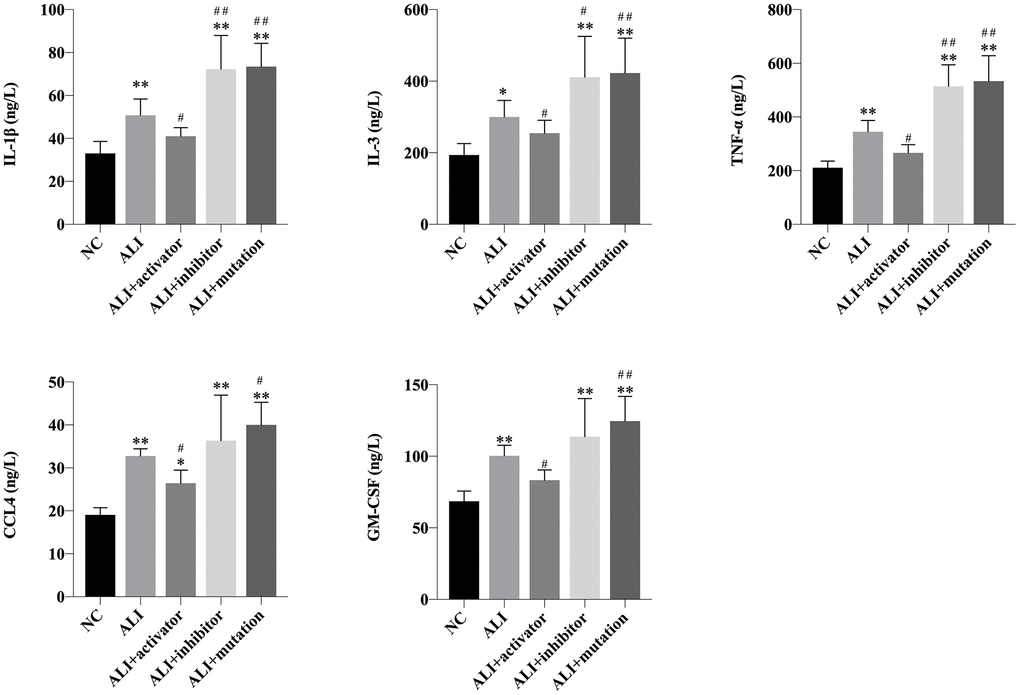
Figure 3. The level of IL-1β, IL-3, TNF-α, CCL4 and GM-CSF in lung tissue of mice in NC, ALI, ALI+activator, ALI+inhibitor and ALI+mutation group measured by ELISA analysis. *P<0.05 vs NC group, **P<0.01 vs NC group, #P<0.05 vs ALI group, ##P<0.01 vs ALI group.
Activation of KMT5B upregulated expression of ADGRF5, SFTPD and SFTPB protein
The RBL1 and KMT5B played important roles in regulating the methylation of H4K20, and SFTPD and SFTPB were important for maintaining the homeostasis of alveolar surfactants. The expressions of ADGRF5, RBL1, KMT5B, SFTPD and SFTPB proteins were determined by western blotting. As presented in Figure 4, all the protein expressions were downregulated in ALI mice. Activation of KMT5B could increase these protein levels, whereas unmethylated H4K20 constructed via inhibition of KMT5B or mutation of H4K20 could decrease the protein levels, compared with the levels of those in ALI mice.
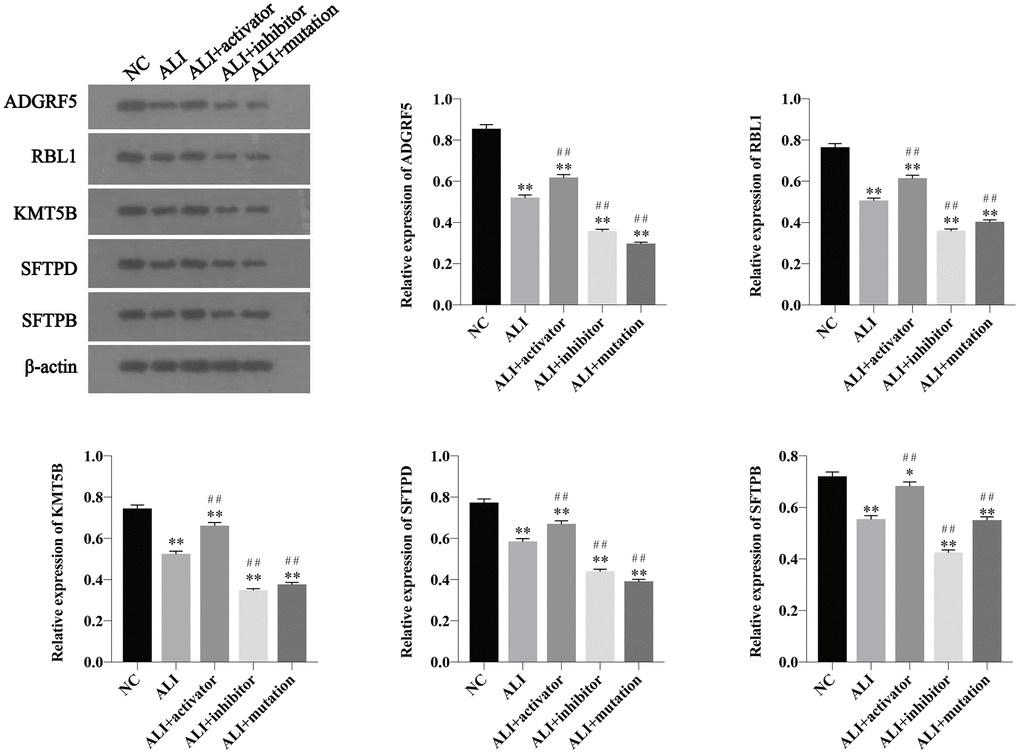
Figure 4. The expression of ADGRF5, RBL1, KMT5B, SFTPD and SFTPB in lung tissue of mice in NC, ALI, ALI+activator, ALI+inhibitor and ALI+mutation group determined by western blotting. The relative expression was calculated via normalized to the expression of β-actin. *P<0.05 vs NC group, **P<0.01 vs NC group, #P<0.05 vs ALI group, ##P<0.01 vs ALI group.
E2F3 regulated the expression of ADGRF5
Whether E2F3 could bind to the ADRGF5 gene promoter region and regulate the its expression was investigated using CHIP analysis. The ADRGF5 gene specific fragments were detected in the DNA fragments immunoprecipitated by E2F3 antibody. According to the agarose gel electrophoresis (Figure 5), we could conclude that content of E2F3-ADGRF5 complex was decreased in ALI mice. Higher level of the complex was detected after activation of KMT5B compared with ALI group, while fewer complexes could be obtained from ALI+inhibitor and ALI+mutation group.
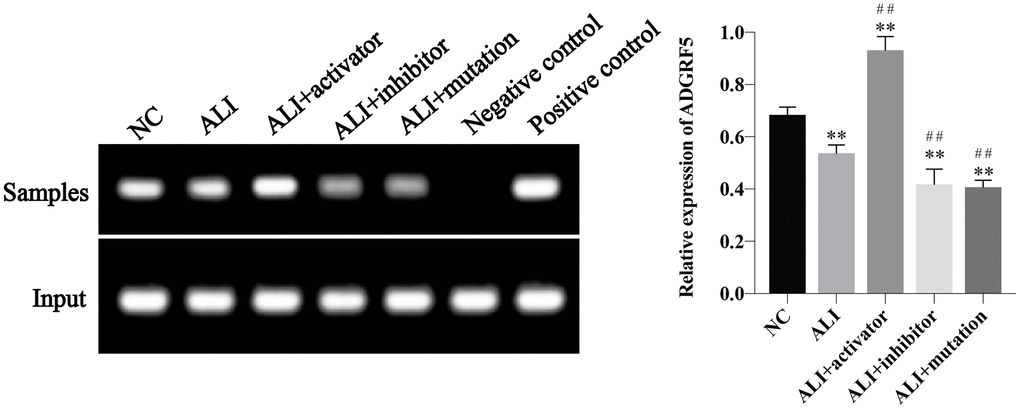
Figure 5. The binding of E2F3 and ADGRF5 was demonstrated using CHIP analysis and the relative expression of ADGRF5 was calculated via normalized to the input group. *P<0.05 vs NC group, **P<0.01 vs NC group, #P<0.05 vs ALI group, ##P<0.01 vs ALI group.
Discussion
Paraquat, a widely used herbicide in the developing countries, could primarily target lung tissues, leading to ALI and acute respiratory distress syndrome. ADGRF5, which belonged to the GPCRs, mainly participated in maintaining lung surfactant homeostasis through regulating pulmonary surfactant-associated proteins that affect the surface tension of the alveoli [4]. Gollamudi et al. found that pesticides including paraquat, pyridaben and maneb could affect the expression of GPCRs [12]. Pandey et al. also reported that GPCR could extend the life span of Drosophila melanogaster and increase resistance to paraquat-induced oxidative stress [13]. Consistently, our findings revealed that the expression of ADGRF5 was downregulated in mice treated with paraquat, while upregulation of ADGRF5 could remarkably improve the morphological status in lung tissue of ALI mice.
ADGRF5 was reported to regulate surfactant secretion and function as a molecular sensor of alveolar surfactant lipid pool sizes [14]. Gil et al. demonstrated that the level of SFTPB and SFTPD was decreased after paraquat treatment [15]. In the present study, the expression of SFTPB and SFTPD was downregulated when the level of ADGRF5 was decreased, indicating that ADGRF5 might exhibit its protection role via regulating SFTPB and SFTPD. Surfactant-associated proteins were reported to attenuate inflammatory status [16]. Luan et al. found the inflammatory factors including IL-6 and TNF-α were upregulated in paraquat-induced ALI mice [17]. The ELISA analysis implemented in this study indicated that overexpression of SFTPB and SFTPD through upregulating ADGRF5 could significantly improve the inflammatory status via decreasing levels of IL-1β, IL-3, TNF-α, CCL4 and GM-CSF.
Histone modifications were found to be involved in the regulation of various genes and were related to multiple diseases such as gastric cancer or acute kidney injury [18, 19]. A recent study also showed that histone deacetylase 7 played a protective role in ALI mice [20]. However, few studies focused on the relationship of H4K20me3 and paraquat-induced ALI [21, 22]. We investigated the effects of H4K20me3 via inhibition/activation of KMT5B and transfection of H4K20-mutated plasmid. Results revealed that the expression of RBL1 and KMT5B was downregulated in ALI mice, and activation of KMT5B could attenuate paraquat-induced damages in lung tissues. Moreover, inhibition of KMT5B or mutation of H4K20, which led to the unmethylation of H4K20, and resulted in the loss of protective ability. The expression of E2F3, a vital transcription factor which was regulated by RBL1, was also found binding to the promoter of ADGRF5 gene.
In summary, all these evidences suggested that paraquat-induced ALI could be inhibited through RBL1/KMT5B/E2F3 signaling, in which way the trimethylation of H4K20 as well as the expression of ADGRF5 were further regulated. This study provides valuable pathways and targets for the treatment of paraquat-induced ALI. However, investigations of other potential molecular pathways using transcriptome or proteome are also needed to understand related mechanisms comprehensively.
Materials and Methods
Tested animals grouping
The adult male BALB/c mice (20-25g, 6-8 weeks old) were obtained from Jiangsu ALF Biotechnology Co., LTD (Nanjing, China). A total of 100 mice were randomly divided into 5 groups (n=20 in each group) that included the NC group, ALI group, ALI+activator group (transfected with KMT5B overexpression plasmid), ALI+inhibitor group (transfected with plasmid that encoded anti-KMT5B antibody) and ALI+mutation group (transfected with plasmid of which histone H4K20 was mutated to tyrosine). The mice in NC group received saline at a dose of 25mg/kg body weight via intragastric administration. And for the ALI mice, paraquat at the equivalent dose of 25mg/kg was administered intragastrically. All the mice were housed under temperature of 22 ± 2° C with 12 h light/dark cycle, and free access to fresh water and food. The animal experiments were executed in accordance with institutional guidelines, following a protocol approved by the Ethics Committees of Frist Affiliated Hospital of Zhengzhou University.
Histological evaluation
The lung tissues of mice were fixed with 10 % neutral buffered formalin, after which the tissues were embedded in paraffin wax and cut into 2-μm sections. Hematoxylin and eosin (H&E) staining was performed to visualize the physiological status in lung tissue. Stained specimens were observed under an Axioplan 2 imaging analyzer.
Immunohistochemical staining analysis
The previously constructed 2-μm paraffin wax sections were dewaxed in xylene and rehydrated in grade alcohol. 3% hydrogen peroxide was added dropwise to the sectioned tissue to block endogenous peroxidase, and then incubated at room temperature for 15 min, and washed three times with PBS for 3 min each time. Then the mixture was added with 5% normal goat serum for blocking and maintained at the root temperature for 30 min. The sections were then incubated with primary monoclonal mouse anti-ADGRF5 antibody (1:100 (v/v)) at 4° C overnight, after which the specimens were washed for three times with PBS for 3 min each time and incubated horseradish peroxidase -conjugated secondary anti-mouse antibody for 30 min at 37° C. Diaminobiphenyl was added for staining, after which sections were rinsed with water to stop staining and were re-stained with hematoxylin for 1 min. The sections were rinsed in water, dehydrated with gradient alcohol and xylene, and air-dried to prepare sections using neutral gum. Stained sections were observed under an Axioplan 2 imaging analyzer.
Western blotting
After lung tissues were broke down with 2XSDS buffer supplemented with protease and phosphatase inhibitors and then homogenized. Supernatant was collected after centrifugations at 10000 r/min for 15 min and preserved for protein quantification using a BCA kit (Beyotime Biotechnology, Shanghai, China). Proteins (30 μg per lane) were separated through SDS-polyacrylamide gel electrophoresis (PAGE) and then electroblotted onto polyvinylidene fluoride membranes (PVDF, Thermo, CA, USA) at 25 V for 30 min. The PVDF membranes were blocked in 5% non-fat milk for 2 h, and were subsequently incubated overnight at 4° C with primary antibodies that included anti-ADGRF5 antibody (1:1000(v/v)), anti-RBL1 antibody (1:1000), anti-KMT5B antibody (1:1000), anti-SFTPD antibody (1:1000), anti-SFTPB antibody (1:1000) and anti-β-ACTIN antibody (1:1000) obtained from Abcam. Blots were washed with TBST for 10 min each in triplicates and then incubated for 1h min at 37° C with HRP-conjugated secondary anti-mouse antibodies (1:2000). Finally, the blot bands on PVDF membranes were detected using electrochemiluminescence reagent (Santa Cruz Biotechnology).
ELISA assay for inflammatory cytokines
A total of 400 mg of lung tissue was grinded thoroughly with a tissue homogenizer via adding 2 mL of normal saline, after which the mixture was centrifuged at 4000 r/min for 20 min and the supernatant was store in a -80° C refrigerator for later use. After sample pretreatment, ELISA kits (Nanjing Jiancheng Bioengineering Cooperation, Nanjing, China) were utilized to detect the expressions of inflammatory cytokines including IL-1β, IL-3, TNF-α, CCL4 and GM-CSF according to manufacturer’s instruction.
Chromatin immunoprecipitation
The chromatin of lung tissue of mice was fixed with a 1% formaldehyde solution for 10 min at room temperature, and glycine was added and maintained for 5 min to terminate fixation. After washing with PBS for twice, cells were collected through centrifugation at 5000rpm/min for 5 min at 4° C, and the precipitate was lysed with SDS lysis buffer. The isolated nuclear extract was sheared by sonication for 10 cycles (30 sec on and 30 sec off for each cycle), after which the DNA/protein complexes in precipitate were obtained via 12000 rpm/min for 10 min at 4° C and 20 μL of DNA fragments in supernatant was collected as Input group. DNA gels were performed to ensure that the isolated chromatin had been sheared into fragments of approximately 500-800 bp. The magnetic beads were prepared by incubation with the E2F3 antibody (IgG served as negative control and polymerase II served as positive control) at 4° C for 1 h, before which the beads were precleared with BSA and Salmon Sperm DNA. After being immunoprecipitated with the magnetic beads overnight at 4° C, the chromatin together with the Input group was washed and added with 5M NaCl to reverse the DNA/protein crosslinks at 65° C overnight. Finally, the DNA was purified with a MinElute column (catalog 28206, Qiagen), and the ADGRF5 fragments were detected using qPCR. The specific primers for detecting ADGRF5 fragments were F: 5’-CGAGCCGTTGCCACAAAAAG-3’, R: 5’-ATGTCGGTAATTTGGTCAGTGTT-3’, and the amplified fragment length is 155 bp. PCR parameters: 94° C denaturation for 5 min, 35 cycles of 94° C for 45 s, 60° C for 45s, 72° C for 60 s, and finally 72° C extension for 5 min. The PCR products were subjected to 2% agarose gel electrophoresis, stained with nucleic acid dyes, and the results were observed under ultraviolet light.
Statistical analysis
All experiments were performed in triplicates and all statistical data were expressed as ± standard deviation (SD). The comparisons among multiple groups were performed using one-way analysis of variance (ANOVA) followed by Turkey’s test. Student’s t-test was implemented for comparison between two groups using GraphPad Prism 7.0 software. A value of P < 0.05 or P<0.01 was considered statistically significant.
Ethics approval and consent to participate
The ethic approval was obtained from the Ethic Committee of Frist Affiliated Hospital of Zhengzhou University.
Consent to publish
All of the authors have Consented to publish this research.
Availability of data and materials
The data are free access to available upon request.
Author Contributions
YXY and LL conducted experiments, YXY and LL collected and analyzed data; YXY and LL for writing of the manuscript. YXY and LL for the discussion of and YXY and LL for discussion and comments on an earlier version of the manuscript. All authors read and approved the final manuscript.
Acknowledgments
We would like to acknowledge the everyone for their helpful contributions on this paper.
Conflicts of Interest
All authors declare no conflicts of interest.
References
- 1. Gawarammana IB, Buckley NA. Medical management of paraquat ingestion. Br J Clin Pharmacol. 2011; 72:745–57. https://doi.org/10.1111/j.1365-2125.2011.04026.x [PubMed]
- 2. Ding YW, Zhao GJ, Li XL, Hong GL, Li MF, Qiu QM, Wu B, Lu ZQ. SIRT1 exerts protective effects against paraquat-induced injury in mouse type II alveolar epithelial cells by deacetylating NRF2 in vitro. Int J Mol Med. 2016; 37:1049–58. https://doi.org/10.3892/ijmm.2016.2503 [PubMed]
- 3. Brown K, Filuta A, Ludwig MG, Seuwen K, Jaros J, Vidal S, Arora K, Naren AP, Kandasamy K, Parthasarathi K, Offermanns S, Mason RJ, Miller WE, et al. Epithelial Gpr116 regulates pulmonary alveolar homeostasis via Gq/11 signaling. JCI Insight. 2017; 2:e93700. https://doi.org/10.1172/jci.insight.93700 [PubMed]
- 4. Kubo F, Ariestanti DM, Oki S, Fukuzawa T, Demizu R, Sato T, Sabirin RM, Hirose S, Nakamura N. Loss of the adhesion G-protein coupled receptor ADGRF5 in mice induces airway inflammation and the expression of CCL2 in lung endothelial cells. Respir Res. 2019; 20:11. https://doi.org/10.1186/s12931-019-0973-6 [PubMed]
- 5. Kay S, Madan T. Fertility defects in surfactant associated protein D knockout female mice: altered ovarian hormone profile. Mol Immunol. 2016; 71:87–97. https://doi.org/10.1016/j.molimm.2016.01.002 [PubMed]
- 6. Luger K. Structure and dynamic behavior of nucleosomes. Curr Opin Genet Dev. 2003; 13:127–35. https://doi.org/10.1016/s0959-437x(03)00026-1 [PubMed]
- 7. Leszinski G, Gezer U, Siegele B, Stoetzer O, Holdenrieder S. Relevance of histone marks H3K9me3 and H4K20me3 in cancer. Anticancer Res. 2012; 32:2199–205. [PubMed]
- 8. Nelson DM, Jaber-Hijazi F, Cole JJ, Robertson NA, Pawlikowski JS, Norris KT, Criscione SW, Pchelintsev NA, Piscitello D, Stong N, Rai TS, McBryan T, Otte GL, et al. Mapping H4K20me3 onto the chromatin landscape of senescent cells indicates a function in control of cell senescence and tumor suppression through preservation of genetic and epigenetic stability. Genome Biol. 2016; 17:158. https://doi.org/10.1186/s13059-016-1017-x [PubMed]
- 9. Van Den Broeck A, Brambilla E, Moro-Sibilot D, Lantuejoul S, Brambilla C, Eymin B, Gazzeri S. Loss of histone H4K20 trimethylation occurs in preneoplasia and influences prognosis of non-small cell lung cancer. Clin Cancer Res. 2008; 14:7237–45. https://doi.org/10.1158/1078-0432.CCR-08-0869 [PubMed]
- 10. Isaac CE, Francis SM, Martens AL, Julian LM, Seifried LA, Erdmann N, Binné UK, Harrington L, Sicinski P, Bérubé NG, Dyson NJ, Dick FA. The retinoblastoma protein regulates pericentric heterochromatin. Mol Cell Biol. 2006; 26:3659–71. https://doi.org/10.1128/MCB.26.9.3659-3671.2006 [PubMed]
- 11. Rotgers E, Rivero-Müller A, Nurmio M, Parvinen M, Guillou F, Huhtaniemi I, Kotaja N, Bourguiba-Hachemi S, Toppari J. Retinoblastoma protein (RB) interacts with E2F3 to control terminal differentiation of sertoli cells. Cell Death Dis. 2014; 5:e1274. https://doi.org/10.1038/cddis.2014.232 [PubMed]
- 12. Gollamudi S, Johri A, Calingasan NY, Yang L, Elemento O, Beal MF. Concordant signaling pathways produced by pesticide exposure in mice correspond to pathways identified in human Parkinson’s disease. PLoS One. 2012; 7:e36191. https://doi.org/10.1371/journal.pone.0036191 [PubMed]
- 13. Pandey A, Khatoon R, Saini S, Vimal D, Patel DK, Narayan G, Chowdhuri DK. Efficacy of methuselah gene mutation toward tolerance of dichlorvos exposure in drosophila melanogaster. Free Radic Biol Med. 2015; 83:54–65. https://doi.org/10.1016/j.freeradbiomed.2015.02.025 [PubMed]
- 14. Bridges JP, Ludwig MG, Mueller M, Kinzel B, Sato A, Xu Y, Whitsett JA, Ikegami M. Orphan G protein-coupled receptor GPR116 regulates pulmonary surfactant pool size. Am J Respir Cell Mol Biol. 2013; 49:348–57. https://doi.org/10.1165/rcmb.2012-0439OC [PubMed]
- 15. Gil HW, Oh MH, Woo KM, Lee EY, Oh MH, Hong SY. Relationship between pulmonary surfactant protein and lipid peroxidation in lung injury due to paraquat intoxication in rats. Korean J Intern Med. 2007; 22:67–72. https://doi.org/10.3904/kjim.2007.22.2.67 [PubMed]
- 16. Schmiedl A, Grützner D, Hoffmann T, von Hörsten S, Stephan M. DPP4 inhibitors increase differentially the expression of surfactant proteins in fischer 344 rats. Acta Physiol (Oxf). 2014; 212:248–61. https://doi.org/10.1111/apha.12350 [PubMed]
- 17. Luan RL, Meng XX, Jiang W. Protective effects of apigenin against paraquat-induced acute lung injury in mice. Inflammation. 2016; 39:752–58. https://doi.org/10.1007/s10753-015-0302-2 [PubMed]
- 18. Liu X, Li Z, Song Y, Wang R, Han L, Wang Q, Jiang K, Kang C, Zhang Q. AURKA induces EMT by regulating histone modification through Wnt/β-catenin and PI3K/Akt signaling pathway in gastric cancer. Oncotarget. 2016; 7:33152–64. https://doi.org/10.18632/oncotarget.8888 [PubMed]
- 19. Fontecha-Barriuso M, Martin-Sanchez D, Ruiz-Andres O, Poveda J, Sanchez-Niño MD, Valiño-Rivas L, Ruiz-Ortega M, Ortiz A, Sanz AB. Targeting epigenetic DNA and histone modifications to treat kidney disease. Nephrol Dial Transplant. 2018; 33:1875–86. https://doi.org/10.1093/ndt/gfy009 [PubMed]
- 20. Kasotakis G, Purves JT, Agarwal S, Sullenger BA, Kintsurashvili E, Galvan MD, Graham C, Corcoran DL, Palmer SM, Remick DG. Histone deacetylase 7 in murine gram-negative acute lung injury. Shock. 2020; 53:375–77. https://doi.org/10.1097/SHK.0000000000001431 [PubMed]
- 21. Zhang F, Hu L, Wu YX, Fan L, Liu WT, Wang J, Sun H, Zhang JS. Doxycycline alleviates paraquat-induced acute lung injury by inhibiting neutrophil-derived matrix metalloproteinase 9. Int Immunopharmacol. 2019; 72:243–51. https://doi.org/10.1016/j.intimp.2019.04.015 [PubMed]
- 22. Hu X, Shen H, Wang Y, Zhang L, Zhao M. Aspirin-triggered resolvin D1 alleviates paraquat-induced acute lung injury in mice. Life Sci. 2019; 218:38–46. https://doi.org/10.1016/j.lfs.2018.12.028 [PubMed]