



Introduction
Frailty, as a manifestation of aging, is a state of increased vulnerability due to cumulative decline in multiple physiological systems [1]. The concept of frailty has been operationalized in two principal models: the frailty phenotype (FP) defines frailty as a clinical syndrome associated with unintentional weight loss, weak grip strength, slow walking speed, exhaustion, and low physical activity [2], whereas the Rockwood frailty index (FI) summarizes vulnerability quantitatively and defines frailty as the accumulation of deficits from a wide range of physical and psychosocial functioning [3, 4]. Regardless of the definition being used, frailty has been consistently linked to adverse outcomes such as falls, hospitalizations, loss of independence, and mortality [5], posing a significant public health concern. Tackling frailty is also recognized as a priority by the European Union [6], calling for better understanding of the syndrome. The prevalence of frailty increases with age, with an overall estimated prevalence of 10.7% among those aged ≥65 and 26.1% among those aged ≥85, varying depending on which frailty scale is used [7]. Meanwhile, there is sizable variability in the frailty status at all ages [8], highlighting the necessity to gain understanding on the underpinnings of the individual differences. The sex-paradox of frailty has been widely described, in which women experience higher levels of frailty but men are more vulnerable to death at any given level of frailty, yet the mechanisms behind remain elusive [9].
In accordance with its multifactorial nature, frailty is influenced by both genetic and environmental factors, with twin-based heritability estimates of the FI ranging from 30 to 45% [10, 11]. However, these studies included female twins only, leaving sex differences in the heritability of FI unaddressed. A vast body of literature has discerned the lifestyle and environmental determinants of frailty [12–14], and showed a persisting socioeconomic gradient in frailty, with increasing socioeconomic adversity associated with higher frailty and contributing to health inequalities even in old age [15]. Two of the most significant risk factors of frailty are unhealthy body mass index (BMI) ranges, including both underweight and obesity [16–18], and low educational attainment [12, 19–21]. A recent Mendelian randomisation analysis showed that the genetic risk of higher BMI and lower educational attainment had the strongest associations with frailty among all modifiable risk factors [22]. However, few studies have elucidated the mechanisms by which these risk factors influence frailty, and whether there may be overlap in the genetic and/or environmental underpinnings of FI with BMI and education. Moreover, it is not known whether the genetic and environmental influences on the FI are altered by BMI and education levels, that is, whether there is gene-environment interaction (G×E) such that genetic risk of frailty may be amplified or suppressed with different environmental circumstances. It is also unclear whether men and women may differ with respect to G×E.
Twin studies offer a natural experiment to partition variance of traits into genetic and environmental etiologies by comparing the genetic similarities between monozygotic (MZ) and dizygotic (DZ) twins, who share 100% and ~50% of their segregating alleles respectively. Large twin samples also allow robust assessment on whether the heritability of a trait differs quantitatively by sex, meaning whether it is the same genes but at different magnitudes that act upon the trait. Moreover, with the inclusion of opposite-sex twins, qualitative sex differences can be examined, informing whether the trait is influenced by different genetic factors in men and women. As the extension to classical twin models, bivariate models allow study of the sources of covariation between traits, while moderation models enable examination of G×E. To date, studies on the variance components of the FI are scarce and the potential sex differences remain unaddressed. The mechanisms by which high BMI and low education influence frailty are likewise poorly known. Not only is understanding the basis of the sex differences in frailty of importance, but identifying how the risk factors affect frailty will also aid in informing preventive strategies and public health interventions.
Towards this end, we performed structural equation modelling using data from the Screening Across the Lifespan Twin study (SALT), a large sample of Swedish twins (n = 42,994), with aims to (i) provide a comprehensive estimation of the contribution of genetic and environmental factors to the variance of the FI, and identify potential quantitative and qualitative sex differences therein; (ii) explore the common genetic and environmental influences on the covariation of FI with BMI and education, two of the most prominent risk factors of frailty; and (iii) test for G×E by levels of BMI and education.
Results
Sample characteristics
Table 1 shows the descriptive statistics of the sample, among the 42,994 twin individuals the mean age of which was 58.8 (standard deviation 10.7; range 41–99). The median FI, i.e., the proportion of endorsed frailty items out of 44 (items listed in Supplementary Table 1), was 0.108 (interquartile range 0.062–0.176). Women were on average older (mean age 59.2 vs 58.4 years) and had a higher median FI (0.119 vs 0.097) than men. Mean BMI and length of education were 25.0 kg/m2 and 10.5 years respectively; both were similar across sex.
Table 1. Characteristics of study population by sex.
| Characteristic | Total (n = 42,994) | Men (n = 19,940) | Women (n = 23,054) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Age at interview, mean (SD) | 58.8 (10.7) | 58.4 (10.4) | 59.2 (11.0) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FI, median (IQR) | 0.108 (0.062, 0.176) | 0.097 (0.057, 0.153) | 0.119 (0.068, 0.193) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BMI (kg/m2), mean (SD) | 25.0 (3.5) | 25.5 (3.1) | 24.5 (3.8) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Education (years), mean (SD) | 10.5 (3.2) | 10.5 (3.2) | 10.4 (3.3) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Zygosity, n (%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MZ | 10,785 (25.1) | 4,788 (24.0) | 5,997 (26.0) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Twins from complete pairs | 8,516 (79.0) | 3,640 (76.0) | 4,876 (81.3) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DZ same-sex | 16,448 (38.3) | 7,640 (38.3) | 8,808 (38.2) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Twins from complete pairs | 11,824 (71.9) | 5,266 (68.9) | 6,558 (74.5) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DZ opposite-sex | 15,761 (36.7) | 7,512 (37.7) | 8,249 (35.8) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Twins from complete pairs | 11,582 (73.5) | 5,791 (77.1) | 5,791 (70.2) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Note: BMI, body mass index; DZ, dizygotic twins; FI, frailty index; IQR, interquartile range; MZ, monozygotic twins; SD, standard deviation. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Phenotypic, intraclass and cross-twin cross-trait correlations
Age-adjusted correlations across zygosity are presented in Table 2 (unadjusted correlations in Supplementary Table 2). For FI, BMI and education, when correlating the same phenotype across twins in a pair (i.e., intraclass correlation), the MZ correlations were larger than DZ correlations and were less than 1, suggesting both genetic and individual-specific environmental influences on all three traits. We observed significantly greater intraclass correlations of FI in women than in men for same-sex twins, implying quantitative sex differences; nevertheless, intraclass correlation of FI in opposite-sex twins did not seem to be smaller than that in same-sex DZ twins, suggesting absence of qualitative sex differences.
Table 2. Adjusted phenotypic correlations, intraclass correlations and cross-twin cross-trait correlations for the frailty index (FI), body mass index (BMI) and education.
| Zygosity | Phenotypic correlations | Intraclass correlations | Cross-twin cross-trait correlations | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FI and BMI | FI and Education | FI | BMI | Education | FI and BMI | FI and Education | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Total | 0.13 (0.12, 0.14) | -0.09 (-0.10, -0.08) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MZ | 0.12 (0.10, 0.14) | -0.09 (-0.11, -0.07) | 0.51 (0.49, 0.53) | 0.68 (0.67, 0.70) | 0.65 (0.64, 0.67) | 0.10 (0.08, 0.12) | -0.07 (-0.10, -0.05) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DZ | 0.14 (0.13, 0.15) | -0.09 (-0.11, -0.07) | 0.18 (0.17, 0.20) | 0.23 (0.22, 0.25) | 0.42 (0.40, 0.43) | 0.06 (0.05, 0.07) | -0.08 (-0.09, -0.06) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MZ males | 0.14 (0.11, 0.17) | -0.10 (-0.13, -0.07) | 0.45 (0.41, 0.48) | 0.66 (0.63, 0.68) | 0.66 (0.63, 0.68) | 0.11 (0.08, 0.14) | -0.07 (-0.10, -0.04) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MZ females | 0.15 (0.12, 0.18) | -0.09 (-0.12, -0.06) | 0.53 (0.50, 0.56) | 0.69 (0.67, 0.71) | 0.65 (0.63, 0.67) | 0.13 (0.10, 0.16) | -0.08 (-0.11, -0.05) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DZ males | 0.12 (0.09, 0.14) | -0.09 (-0.11, -0.07) | 0.12 (0.09, 0.16) | 0.27 (0.24, 0.31) | 0.44 (0.42, 0.47) | 0.05 (0.02, 0.07) | -0.07 (-0.10, -0.05) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DZ females | 0.19 (0.17, 0.21) | -0.07 (-0.09, -0.05) | 0.24 (0.20, 0.27) | 0.31 (0.28, 0.34) | 0.46 (0.43, 0.48) | 0.10 (0.08, 0.13) | -0.06 (-0.09, -0.04) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DZ opposite-sex | 0.11 (0.09, 0.13) | -0.10 (-0.13, -0.08) | 0.17 (0.15, 0.20) | 0.18 (0.15, 0.20) | 0.38 (0.36, 0.41) | 0.03 (0.01, 0.06) | -0.09 (-0.11, -0.06) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Note: MZ, monozygotic twins; DZ, dizygotic twins. Phenotypic correlations are the within-individual correlations between FI and BMI, and between FI and education. Intraclass correlations indicate the extent to which each trait correlates within twin pairs. Cross-twin cross-trait correlations show the extent to which FI of the first twin correlate with the other trait (i.e., BMI or education) of the second twin. 95% confidence intervals are presented in parentheses. All correlations were adjusted for age. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
FI had a positive phenotypic correlation with BMI (r = 0.13) and a negative correlation with education (r = -0.09). The higher MZ than DZ cross-twin cross-trait correlation between FI and BMI indicated that part of their covariation could be explained by genetic influences in common. In contrast, cross-twin cross-trait correlations between FI and education were comparable for MZ and DZ twins, indicating environmental factors common to twins in a pair, rather than genetic factors, contributing to their covariance.
Univariate twin modelling
Univariate sex-limitation models of FI were fitted as shown in Table 3, providing estimates of the variance components in men and women: additive genetic factors (A), dominance genetic factors (D), common environmental factors (C), and unique environmental factors (E). Overall, the best-fitting model was an ADE model (i.e., a model including A, D and E variance components) with only quantitative sex differences. Compared to the saturated model (i.e., a model that fully describes the observed data), the full ADE model did not provide a worse model fit, indicating that the model fit the data well, but both the ACE and AE models provided a poor fit to the data. Removing quantitative sex differences (i.e., different magnitude of heritability) from the ADE quantitative sex-limitation model resulted in a reduced model fit. Meanwhile, the genetic correlation for opposite-sex twins (rfm), which is used for assessing qualitative sex difference (i.e., different sources of genetic variance), was close to, and statistically non-significantly different from unity. This suggests the presence of quantitative, but not qualitative sex differences in the genetic contributions to FI. We observed largely dissimilar proportions of additive and dominance genetic influences on FI for men and women. Alternatively, we estimated the broad-sense heritability, H, of FI (i.e., proportion of variance that can be explained by the total of additive and dominance genetic factors), which was estimated to be 45% [95% confidence interval (CI): 41–48%] in men, and at a statistically significantly higher proportion of 52% (95% CI: 50–55%) in women. The rest of the variation in FI was explained by unique environmental factors.
Table 3. Model fitting results and parameter estimates from univariate sex-limitation models of the frailty index (FI).
| Model | Model fit statistics | Parameter estimates for men and women (95% CI) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| AIC | ΔLL | Δdf | p | A | D/C | H | E | rfm | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Saturated | 19953 | - | - | - | - | - | - | - | - | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ADE full sex-limitation | 19940 | 19.1 | 16 | 0.264 | M: 7% (0, 23) | M: 38% (21, 55) | M: 44% (41, 48) | M: 56% (52, 59) | 0.69 (0.41, 0.96) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F: 41% (28, 55) | F: 11% (0, 25) | F: 52% (50, 55) | F: 48% (45, 50) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ADE quantitative sex-limitation | 19939 | 19.7 | 17 | 0.288 | M: 0% (0, 1) | M: 44% (41, 48) | M: 45% (41, 48) | M: 55% (52, 59) | 1.00 (NA) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F: 41% (28, 55) | F: 11% (0, 25) | F: 52% (50, 55) | F: 48% (45, 50) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ADE no sex difference | 19949 | 32.1 | 18 | 0.021 | M: 0% (0, 2) | M: 49% (45, 52) | M: 49% (47, 51) | M: 51% (49, 53) | 1.00 (NA) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F: 44% (31, 58) | F: 4% (0, 19) | F: 49% (47, 51) | F: 51% (49, 53) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ACE full sex-limitation | 19961 | 40.4 | 16 | 0.001 | M: 41% (37, 44) | M: 0% (0, 0) | M: 41% (37, 44) | M: 59% (56, 63) | 0.76 (0.64, 0.88) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F: 51% (49, 54) | F: 0% (0, 0) | F: 51% (49, 54) | F: 49% (46, 51) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| AE full sex-limitation | 19957 | 40.4 | 18 | 0.002 | M: 41% (37, 44) | M: 0% (NA) | M: 41% (37, 44) | M: 59% (56, 63) | 0.76 (0.64, 0.88) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F: 51% (49, 54) | F: 0% (NA) | F: 51% (49, 54) | F: 49% (46, 51) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Note: AIC, Akaike’s Information Criterion; LL, log-likelihood; df, degrees of freedom; p, p-values of likelihood ratio tests compared with the saturated model. CIs are Wald-type confidence intervals with lower and upper bounds of 0 and 1 respectively. A, additive genetic factors; D, dominance genetic factors; C, common environmental factors; H, total genetic factors/ broad-sense heritability; E, unique environmental factors; rfm, genetic correlation between men and women, estimated using opposite-sex twins. M and F represents parameter estimates for men and women respectively. Full sex-limitation models allowed both quantitative and qualitative sex differences. In ADE quantitative sex-limitation model, rfm was fixed to be 1. In ADE no sex difference model, broad-sense heritability of men and women were equated, but variance difference between sex was allowed. ACE and AE sub-models are not shown as the full models fit significantly worse than the saturated model. All models were adjusted for age. Best-fitting model is shown in bold. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Bivariate twin modelling
Using bivariate twin models, we assessed the overlap of genetic and environmental variance components of FI with BMI and education and allowed for sex differences. The best-fitting bivariate model for FI and BMI was an ADE model (Figure 1A; model fit statistics and parameter estimates in Supplementary Tables 3, 4). Variance of BMI was predominantly explained by genetic factors (men: H = 66%; women: H = 69%). We observed modest genetic correlations between FI and BMI, which were 0.19 (95% CI: 0.14–0.23) and 0.26 (95% CI: 0.22–0.29) in men and women, respectively. The bivariate heritability for FI and BMI, i.e., the proportion of covariance explained by genetic factors, was 81% (95% CI: 65–97%) in men and 87% (95% CI: 78–95%) in women, indicating that a substantial part of the correlation between FI and BMI could be attributable to genetic factors in common to both traits (Figure 1C). By contrast, the best-fitting bivariate model for FI and education according to the Akaike’s Information Criterion (AIC) was an ACE model (Figure 1B), although, all bivariate models between FI and education provided a worse fit than the saturated model. A moderate proportion of the variation in education was due to common environmental factors, which was 25% (95% CI: 19–31%) in men and 29% (95% CI: 23–34%) in women. The C component of FI was small and non-significant in both sexes, however, they had perfect negative correlations with that of education (rC = -1.00). Common environmental factors also accounted for considerable proportions of the phenotypic correlations between FI and education, which were 65% (95% CI: 23–107%) in men and 74% (95% CI: 22–126%) in women (Figure 1C). There was only weak overlap of genetic factors and unique environmental factors between FI and education.
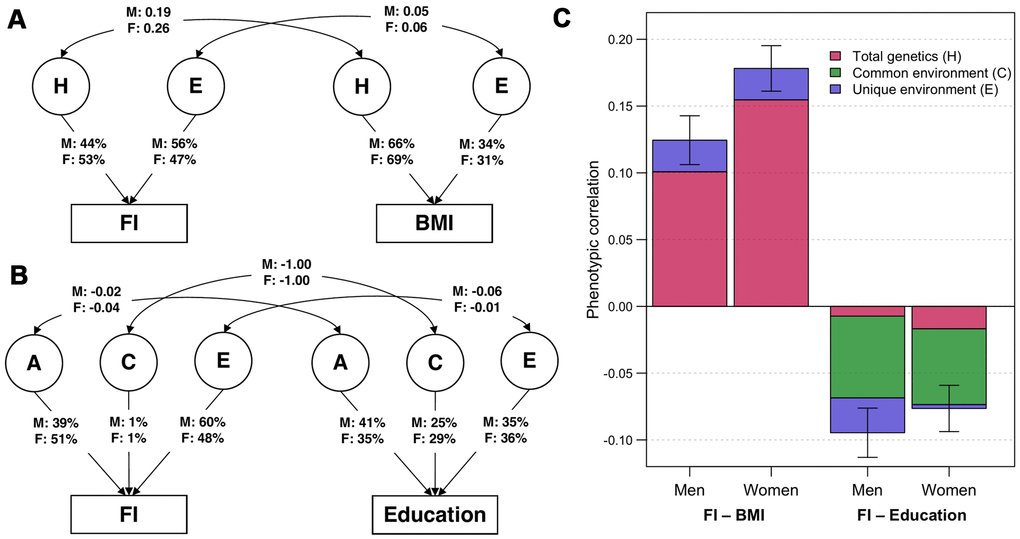
Figure 1. Parameter estimates from the best-fitting bivariate twin models. (A) ADE bivariate quantitative sex-limitation model between frailty index (FI) and body mass index (BMI), adjusted for age. Single headed-arrows represent the proportion of each traits explained by latent (circular) variance components; while double-headed arrows represent correlations between variance components. H indicates the sum of additive and dominance genetic factors; E indicates unique environmental factors; M and F are the estimates for men and women respectively. (B) ACE bivariate quantitative sex-limitation model between FI and education, adjusted for age. A indicates additive genetic factors; C indicates common environmental factors. (C) Phenotypic correlations of FI with BMI and education among men and women (with 95% confidence intervals), and the proportion of correlations explained by total genetic, common environmental and unique environmental factors. Note: Model-fitting results and parameter estimates can be found in Supplementary Tables 3, 4.
Moderation analysis
To determine whether genetic and environmental variance components of FI change with levels of BMI and education, we constructed a series of bivariate and univariate moderation models to examine whether there is moderation on the covariance between FI and the moderator (i.e., BMI and education), in addition to the variance unique to FI. For moderation by BMI, we observed significant moderating effects on the variance unique to FI as well as on the covariance between FI and BMI; in contrast, for moderation by education, we observed only significant moderating effects on the unique variance of FI (Supplementary Tables 5, 6). Therefore, a full bivariate moderation model and an extended univariate moderation model were fitted for moderation by BMI and education, respectively, as proposed by van der Sluis et al [23]. The H and E components of FI from the best-fitting moderation models are plotted in Figure 2 (see Supplementary Figure 1 for separate A and D components; and Supplementary Figure 2 for separate common and unique variance estimates of FI by BMI levels). Patterns of moderation were consistent in both men and women, although women had a higher absolute variance of FI than men. The total variance of FI was larger at low and high BMI levels and at fewer years of education. It appeared that genetic variance of FI increased at both ends of BMI levels, while unique environmental variance of FI was similar at all BMI values; hence, the heritability of FI was greater at low and high BMI. In contrast, both genetic and environmental variance of FI declined to a comparable extent with increased years of education; therefore, the relative proportions of genetic and environmental sources of FI variance did not seem to vary across education years.
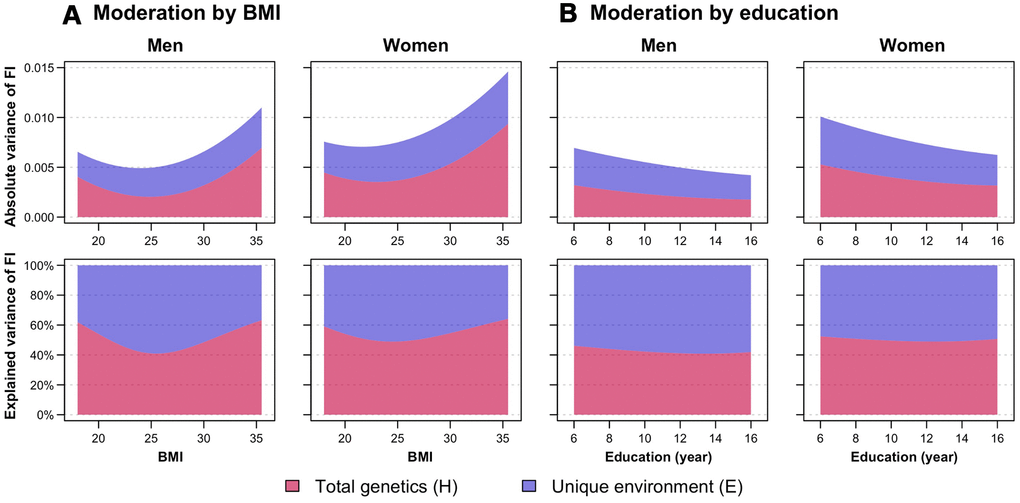
Figure 2. Moderation analysis of frailty index (FI) by (A) body mass index (BMI) and (B) education, stratified by sex. First row shows the absolute variance of FI, while the second row shows the proportion of FI variance explained by total genetic (H, indicating sum of additive and dominance genetic factors) and unique environmental (E) factors, with changes in BMI and education. Variance estimates of moderation by BMI were obtained from the full ADE bivariate moderation model between FI and BMI; while variance estimates of moderation by education were obtained from the ADE extended univariate moderation model between FI and education. Quantitative sex-differences were allowed in the models to obtain estimates separately for men and women. Models were adjusted for age. Note: Model-fitting results can be found in Supplementary Tables 5, 6.
Sensitivity analysis
We additionally performed sensitivity analysis to examine if the skewed distribution of FI affected its heritability estimates. Variance component estimates of the square-root transformed FI were largely consistent with that of the non-transformed FI variable (Supplementary Table 7). Moreover, similar patterns of moderating effects by BMI and education on the transformed FI were observed in men, but there was less evidence of moderation by BMI in women (Supplementary Figure 3). We also fitted the univariate and bivariate models using an alternative “direct symmetric approach” that allows negative variance components [24]; all the conclusions were essentially the same as in the main analysis (Supplementary Tables 8–10).
Discussion
Using a large sample of Swedish twins, we found that the variation in frailty was attributed rather equally to both genetic (with presence of additive and dominance effects) and individual-specific environmental influences. The twin-based heritability of FI was greater in women than in men, but there was no strong evidence of qualitative sex difference, indicating that the same genes, although at different magnitudes, are influencing FI in both sexes. There was small-to-moderate genetic overlap between FI and BMI, and a complete overlap of the environmental factors common to twins within pairs between FI and education, suggesting different mechanisms on how BMI and education influence the risk of frailty. In addition, moderation analysis showed that the total variance of FI varies with levels of BMI and years of education, so that genetic influence on frailty was greater at both low and high levels of BMI, but the heritability of FI did not seem to differ across education years.
Our finding that much of the FI variance is due to genetic factors is in line with two prior UK twin studies analyzing variance components of FI, which reported a heritability of 30–45% [10, 11]. By contrast, it is higher than the SNP-based heritability of 14% estimated in a recent genome-wide association study (GWAS) [22], which may be partly due to the overestimation of heritability in twin studies [25], or that GWAS does not include non-additive genetic effects, as observed in the current study, and it has limited power to detect rare genetic variants that normally exist in complex traits like frailty [25, 26]. Environmental factors shared by members within twin pairs, which are usually those from childhood such as family environment, have negligible influence on frailty. Instead, a substantial proportion of the variation of FI is shaped by environmental factors unique to individuals, although this component also includes measurement error. This may reflect the multidimensional nature of frailty, in which diverse physical, social, behavioral and psychological factors have been linked to the elevated risk of frailty [12–14].
To our knowledge, this is the first study that has formally examined sex differences in the heritability of frailty. We observed rather small, but significantly greater heritability in women (52%) than in men (45%); meanwhile, there was weak and statistically non-significant qualitative sex difference. A higher heritability of some psychological and neurological traits, such as depression [27], pain [28], and insomnia [29], has also been observed among women, which is attributed at least partly to the higher prevalence of these traits in women. Similarly, women may also be more genetically susceptible to frailty, given that the genetic underpinnings of frailty have been found to be associated with neurological pathways [22]. Another possible explanation to the difference in variance components across sex may be that men tend to report health problems less accurately [30], leading to an inflation of the E component. Notably, the total variance of FI was also higher in women than in men, reflecting the fact that women tend to have higher levels of frailty across the age range. The apparent sex difference may merely be owing to the overall greater variation of FI in women, instead of disparities in variance structure across sex, as shown in other traits such as BMI [31]. Furthermore, the observed sex difference is not immense, yet statistically significant, perhaps also due to our large sample that provided enough statistical power. More research is warranted to further investigate if sex differences in genetic and environmental factors contribute to the sex-specificity of frailty.
Our second aim was to assess the overlap in genetic and environmental influences of FI with BMI and education. There was a small-to-moderate genetic correlation between FI and BMI, suggesting that frailty and BMI may in part be influenced by the same genes. This may be attributable to their shared underlying mechanisms of energy metabolism and inflammation, known to be associated with both frailty [32, 33] and BMI [34, 35]. Also, genes related to synaptic pathways, were found to be enriched in the genetic architecture of frailty [22], and likewise may affect BMI [35]. On the other hand, we found a relatively low genetic correlation between FI and education; instead, despite small and non-significant common environmental variance component of FI, it explains a large proportion of the correlation between FI and education, suggesting the relative importance of family environment on the education-frailty association. Previous studies found that genes associated with educational attainment have an inverse relationship with frailty [22, 36]; our finding may therefore imply a possible mechanism that education-associated genes are not directly influencing frailty, but indirectly through affecting individuals’ socioeconomic circumstances [37]. The better family environment may then contribute to higher educational attainment and consequently prevention of frailty development, through the improved health literacy that higher education in the family brings [38, 39], as well as the health seeking behavior that is especially characteristic to women with high education [40]. It is however important to note that the common environmental factors of FI itself were small and not statistically significant and should be interpreted with caution.
Finally, we saw significant moderating effects on the genetic and unique environmental variance components of FI by both BMI and education, and the patterns were generally similar in men and women, although sensitivity analysis showed less evidence of moderation by BMI on the transformed FI in women. The overall variance of FI, as well as the relative proportion of genetic variance component were higher at both low and high BMI levels, which follows the U-shaped association between BMI and physical frailty reported in the literature [16–18]. This indicates that in individuals whose BMI falls outside a healthy range, especially those with obesity (BMI>30), the increased risk of frailty may be due to their more pronounced expression of genetic susceptibilities to frailty. On the other hand, the reduced FI variance with increased years of education is congruent to previous literature showing smaller variance of health status at higher education levels [41]. Meanwhile, the proportions of genetic and unique environmental variance components over the total variance did not seem to differ over education years, suggesting that genetic and individual-specific environmental factors may have a stable contribution to frailty, independent of education.
This study included a population-representative adult twin sample, which provided enough statistical power to examine sex differences in heritability of frailty. We also used a validated FI as the frailty measure, which was shown to predict higher risk of all-cause, cardiovascular disease, and respiratory-related mortality from midlife to old age [42]. Nevertheless, several limitations should be considered. First, given the cross-sectional nature of our data, we could not establish causal relationships among BMI, education and FI. Whether the variance components change with age should also be researched in future longitudinal studies with repeated measurements of frailty. Second, measures of frailty items, BMI, and education are all based on self-reported data, possibly causing misclassification and inaccurate reporting, especially in men [43]. Third, due to the different contribution of dominance genetic factors and common environmental factors to the variance of FI and education, all bivariate models for FI and education provided a poor fit, and we were unable to include both C and D parameters in the same statistical model with only reared-together twins in a classical twin design. Finally, there are some inherent limitations of twin modelling such as potential overestimation of heritability [25] and violation on the assumptions of equal environments (i.e., MZ and DZ twins are treated the same) and random mating in population (i.e., no assortment), although these should have minimal effects on the heritability estimates [44].
Overall, our findings demonstrate that individual differences in frailty are attributable to both genetic and individual-specific environmental factors. Sex differences are evident, in which women have a slightly higher heritability of frailty than men, yet there is limited evidence of different genetic factors influencing frailty in men and women. Furthermore, the two main risk factors of frailty, BMI outside a healthy range and low education, seem to operate through different mechanisms in frailty development, highlighting the relative importance of genes and family environment on the associations of frailty with BMI and education, respectively. These results would help in expanding our current understanding on the individual differences in frailty.
Materials and Methods
Study population
Twin participants were from SALT, which is part of the population-based Swedish Twin Registry [45]. During 1998–2002, all twins born in 1958 or before were invited to attend a telephone interview with questions on common diseases, symptoms, medication use, demographics, and lifestyle factors. With a response rate of 65% for those born in 1886–1925 and 74% for those born in 1926–1958, a total of 44,919 twin individuals participated in the survey [46]. Zygosity was determined based on DNA, or questions on intra-pair similarities during childhood; the latter method was over 95% accurate when validated against DNA testing [47]. This study has been approved by the Swedish Ethical Review Authority in Stockholm. Informed consent was obtained from all participants prior to data collection.
For the present analysis, we excluded those with uncertain zygosity and those who had over 20% missing data across the 44 frailty items, leaving n = 42,994 consisting of 31,922 paired and 11,072 single respondents, aged from 41 to 99 years. Single twin individuals were retained as they contribute to the mean, variance and within-individual covariance estimates. Twins were grouped by zygosity: 4,788 MZ males (1,820 complete pairs), 5,997 MZ females (2,438 complete pairs), 7,640 DZ same-sex males (2,633 complete pairs), 8,808 DZ same-sex females (3,279 complete pairs), and 15,761 DZ opposite-sex individuals (5,791 complete pairs).
Measures
We constructed an FI based on Rockwood’s deficit accumulation model [3], using a total of 44 self-reported frailty items selected from a wide range of health-status related symptoms, signs, disabilities, and diseases in various biological systems (items listed in Supplementary Table 1). Each respondent’s frailty items were summed up and divided by the total number of deficits measured (i.e., 44 in this study), yielding a continuous FI score ranging from 0 to 1. For instance, an individual with five deficits would have an FI of 5/44 = 0.11. The FI in SALT has previously been validated for its ability to predict mortality [42].
Information on body mass index (BMI) and education were obtained from self-reported data. BMI was calculated as weight (kg) divided by height-squared (m2). We defined education as a continuous variable of the number of years of education completed.
Statistical analysis
Twin design
The classical twin design allows decomposition of phenotypic variance into genetic and environmental sources, based on the fact that MZ twins are genetically identical while DZ twins share on average half of their segregating alleles. Genetic sources of variance include additive genetic variance (A, representing the sum of allelic effects at different loci that influence the trait) and dominance genetic variance (D, representing interactions between alleles at the same locus). In MZ twins, both A and D correlate 100%; while in DZ twins, A and D are assumed to correlate 50% and 25% respectively. The sum of A and D influences is referred to as the broad-sense heritability (H), which is the proportion of phenotypic variance that can be explained by genetic effects. On the other hand, environmental sources of variance include common environmental factors (C) such as family environment and environments that are in common or correlated in adulthood, which correlate 100% in both MZ and DZ twins; and unique environmental factors (E) which are uncorrelated between twins and contribute to differences between them. The E component also contains measurement error. In addition to variance, phenotypic covariance can also be partitioned into the same sources of influence, indicating the extent to which genetic and environmental variance components overlap across two or more traits.
Phenotypic, intraclass and cross-twin cross-trait correlations
For initial inspection of the familial similarity for MZ and DZ twin pairs, we estimated intraclass correlations for FI, BMI and education, which show how much each trait correlates within twin pairs. A higher MZ correlation than DZ correlation indicates contribution of genetics to variance in the trait. If DZ correlation is less than half of the MZ correlation, the D component is inferred; on the contrary, the C component is implied if DZ correlation is greater than half of the MZ correlation. A MZ correlation less than 1 indicates presence of E.
Within-individual between-trait correlations, which we refer to as “phenotypic correlations”, were calculated for FI with BMI and education. To explore the genetic and environmental sources of covariances, we computed cross-twin cross-trait correlations, which represent the strength of relationship between one trait of the first twin and another trait of the second twin. They can be interpreted in a similar way as intraclass correlations. For example, a larger cross-twin cross-trait correlation in MZ than DZ twins suggests that phenotypic correlation between the two traits is partially owing to common genetic influences; and if MZ cross-twin cross-trait correlation is less than the phenotypic correlation then E is present.
Phenotypic, intraclass and cross-twin cross-trait correlations across zygosity were estimated from constrained saturated models in which means, variances and phenotypic correlations were equated across twin order. Age was regressed out of the means (as linear effect for FI and linear+quadratic effect for BMI and education, after initial investigation of associations between variables), so that twin correlations are not spuriously inflated due to the same age of twins in pairs.
Univariate twin modelling
Before model fitting, we checked and observed no violations in the assumptions of equal means and variances across twin order and zygosity. Univariate twin models were fitted to estimate variance components of FI, after adjusting for age. Since C and D are confounded in classical twin modelling, either an ACE or ADE model can be fitted at a time. Sex differences were allowed in the models to examine whether genetic influence on FI differs across sex quantitatively (i.e., different magnitude of heritability in men and women) and qualitatively (i.e., different genetic sources in men and women). Quantitative sex differences were modelled by allowing variance components to differ among men and women. Qualitative sex differences were estimated by multiplying a genetic correlation term (rfm) to the expected genetic covariance of opposite-sex twin pairs, where a value of 1 would imply maximum genetic correlation (see Supplementary Methods for details). ACE or ADE sex-limitation models were then compared with their reduced models. We examined quantitative sex difference by constraining the broad-sense heritability to be equal in men and women (but allowing total variance to differ across sex) and tested if there was a significant reduction in model fit. Qualitative sex difference was assessed by testing whether rfm was significantly less than 1. AE models were fitted to determine if the C and D parameters can be excluded from ACE and ADE models.
Bivariate twin modelling
During assumption testing, all means and variances could be equated across twin order and zygosity, except that equating means of education across zygosity led to a significant decrease in model fit, so they were estimated separately in subsequent bivariate models. A “correlated factor model” [48] was applied to estimate the etiological correlations between genetic (rA and rD) and environmental (rC and rE) variance components of FI with BMI and education, apart from estimating variance components of each variables. We define the correlation between A+D in trait X and A+D in trait Y as the broad-sense genetic correlation, rH. Correlation of ±1 indicates complete overlap, while 0 indicates no overlap. Bivariate heritability was also calculated, which is referred to as the proportion of phenotypic correlation explained by genetic covariation. Since estimates of A and D components are highly dependent, as shown by non-significant A and D estimates, we estimated the broad-sense heritability and interpreted in ADE models to indicate the total genetic effects. Only same-sex twins were included in the bivariate quantitative sex-limitation models, and variance components and etiological correlations were estimated separately by sex, after adjusting for age.
Moderation analysis
In previous univariate and bivariate models, we assumed that variance components of FI were constant across the population (i.e., no gene-environment interaction). We relaxed this assumption and examined whether the genetic and environmental influences on FI are moderated by different levels of BMI and education. Moderation may occur on the variance that is unique to FI; for example, changes in genetic variance components of FI with years of education would suggest differential genetic sensitivity of FI to education levels. Furthermore, moderation may exist on the covariance between FI and the moderator, for example, the genetic correlation between FI and BMI may vary with different levels of BMI. To investigate these possible moderating effects, we adopted the full bivariate moderation models developed by Purcell [49], and the extended univariate moderation models proposed by van der Sluis et al [23] (Supplementary Figures 4, 5). By the bivariate moderation models, we first assessed if there were significant moderating effects on the covariance of FI with BMI and education on top of the variance unique to FI. In case of no statistically significant moderation on the covariance between FI and the moderator, the extended univariate moderation models were employed instead. All moderating effects were examined separately by sex in quantitative sex-limitation models using same-sex twins and controlled for age. Absolute genetic and environmental variance components of FI, as well as their proportions to the total variance, were plotted over BMI and education.
Sensitivity analysis
Since the FI variable is positively skewed (Supplementary Figure 6), we followed previous work and applied a square-root transformation to obtain an approximately normally-distributed FI [10, 11]. Univariate twin modelling and moderation analysis of the transformed FI were then performed to check for robustness of our results. We additionally compared our results of univariate and bivariate twin modelling with models fitted using the “direct symmetric approach” proposed by Verhulst et al. [24], which allows variance estimates to be negative and potentially reduces Type I error rates.
For all fitted models, we used the full information maximum-likelihood modelling in the R package OpenMx (version 2.17.3) for estimation of parameters that best fit the observed data. Changes in goodness of fit of models (distributed as χ2) were assessed by likelihood ratio tests, where significant values indicate worse fit of observed data. Akaike information criterion (AIC; a lower value is better) was used to select the most parsimonious and best-fitting models.
Author Contributions
C.A.R., S.H., J.J., and R.K.-H. contributed to the study design and statistical analysis plan. J.K.L.M., X.L., and R.K.-H. performed statistical analyses. N.L.P. is the leader of the SALT study and responsible for cohort recruitment, data collection, and funding. J.K.L.M., M.E., J.J., and R.K.-H. wrote the manuscript. All authors contributed to the interpretation of the results, and read and approved the final manuscript.
Acknowledgments
We acknowledge the Swedish Twin Registry for access to data. The Swedish Twin Registry is managed by Karolinska Institutet and receives funding through the Swedish Research Council under the grant number 2017-00641. This study was accomplished within the context of the Swedish National Graduate School for Competitive Science on Ageing and Health (SWEAH) funded by the Swedish Research Council.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
This work was supported by the Swedish Research Council (2018-02077, 2019-01272), the Loo and Hans Osterman Foundation, FORTE (the Swedish Research Council for Health, Working Life, and Welfare), the Strategic Research Program in Epidemiology at Karolinska Institutet and the Karolinska Institutet Foundations.
References
- 1. Clegg A, Young J, Iliffe S, Rikkert MO, Rockwood K. Frailty in elderly people. Lancet. 2013; 381:752–62. https://doi.org/10.1016/S0140-6736(12)62167-9 [PubMed]
- 2. Fried LP, Tangen CM, Walston J, Newman AB, Hirsch C, Gottdiener J, Seeman T, Tracy R, Kop WJ, Burke G, McBurnie MA, and Cardiovascular Health Study Collaborative Research Group. Frailty in older adults: evidence for a phenotype. J Gerontol A Biol Sci Med Sci. 2001; 56:M146–56. https://doi.org/10.1093/gerona/56.3.m146 [PubMed]
- 3. Searle SD, Mitnitski A, Gahbauer EA, Gill TM, Rockwood K. A standard procedure for creating a frailty index. BMC Geriatr. 2008; 8:24. https://doi.org/10.1186/1471-2318-8-24 [PubMed]
- 4. Rockwood K, Mitnitski A. Frailty in relation to the accumulation of deficits. J Gerontol A Biol Sci Med Sci. 2007; 62:722–27. https://doi.org/10.1093/gerona/62.7.722 [PubMed]
- 5. Vermeiren S, Vella-Azzopardi R, Beckwée D, Habbig AK, Scafoglieri A, Jansen B, Bautmans I, and Gerontopole Brussels Study group. Frailty and the Prediction of Negative Health Outcomes: A Meta-Analysis. J Am Med Dir Assoc. 2016; 17:1163.e1–17. https://doi.org/10.1016/j.jamda.2016.09.010 [PubMed]
- 6. Cano A, Dargent G, Carriazo A, López-Samaniego L, Apostolo J, Campos E, Holland C, Varela-Nieto I, Luz Sánchez-Sánchez M, Illario M, Iaccarino G, Roller RE, Goossens E, et al. Tackling frailty and functional decline: Background of the action group A3 of the European innovation partnership for active and healthy ageing. Maturitas. 2018; 115:69–73. https://doi.org/10.1016/j.maturitas.2018.06.009 [PubMed]
- 7. Collard RM, Boter H, Schoevers RA, Oude Voshaar RC. Prevalence of frailty in community-dwelling older persons: a systematic review. J Am Geriatr Soc. 2012; 60:1487–92. https://doi.org/10.1111/j.1532-5415.2012.04054.x [PubMed]
- 8. Rockwood K, Song X, Mitnitski A. Changes in relative fitness and frailty across the adult lifespan: evidence from the Canadian National Population Health Survey. CMAJ. 2011; 183:E487–94. https://doi.org/10.1503/cmaj.101271 [PubMed]
- 9. Gordon EH, Peel NM, Samanta M, Theou O, Howlett SE, Hubbard RE. Sex differences in frailty: A systematic review and meta-analysis. Exp Gerontol. 2017; 89:30–40. https://doi.org/10.1016/j.exger.2016.12.021 [PubMed]
- 10. Young AC, Glaser K, Spector TD, Steves CJ. The Identification of Hereditary and Environmental Determinants of Frailty in a Cohort of UK Twins. Twin Res Hum Genet. 2016; 19:600–09. https://doi.org/10.1017/thg.2016.72 [PubMed]
- 11. Livshits G, Ni Lochlainn M, Malkin I, Bowyer R, Verdi S, Steves CJ, Williams FM. Shared genetic influence on frailty and chronic widespread pain: a study from TwinsUK. Age Ageing. 2018; 47:119–25. https://doi.org/10.1093/ageing/afx122 [PubMed]
- 12. Chamberlain AM, St Sauver JL, Jacobson DJ, Manemann SM, Fan C, Roger VL, Yawn BP, Finney Rutten LJ. Social and behavioral factors associated with frailty trajectories in a population-based cohort of older adults. BMJ Open. 2016; 6:e011410. https://doi.org/10.1136/bmjopen-2016-011410 [PubMed]
- 13. Feng Z, Lugtenberg M, Franse C, Fang X, Hu S, Jin C, Raat H. Risk factors and protective factors associated with incident or increase of frailty among community-dwelling older adults: A systematic review of longitudinal studies. PLoS One. 2017; 12:e0178383. https://doi.org/10.1371/journal.pone.0178383 [PubMed]
- 14. Raymond E, Reynolds CA, Dahl Aslan AK, Finkel D, Ericsson M, Hägg S, Pedersen NL, Jylhävä J. Drivers of Frailty from Adulthood into Old Age: Results from a 27-Year Longitudinal Population-Based Study in Sweden. J Gerontol A Biol Sci Med Sci. 2020; 75:1943–50. https://doi.org/10.1093/gerona/glaa106 [PubMed]
- 15. Brunner EJ, Shipley MJ, Ahmadi-Abhari S, Valencia Hernandez C, Abell JG, Singh-Manoux A, Kawachi I, Kivimaki M. Midlife contributors to socioeconomic differences in frailty during later life: a prospective cohort study. Lancet Public Health. 2018; 3:e313–22. https://doi.org/10.1016/S2468-2667(18)30079-3 [PubMed]
- 16. Hubbard RE, Lang IA, Llewellyn DJ, Rockwood K. Frailty, body mass index, and abdominal obesity in older people. J Gerontol A Biol Sci Med Sci. 2010; 65:377–81. https://doi.org/10.1093/gerona/glp186 [PubMed]
- 17. Rietman ML, van der A DL, van Oostrom SH, Picavet HS, Dollé ME, van Steeg H, Verschuren WM, Spijkerman AM. The Association between BMI and Different Frailty Domains: A U-Shaped Curve? J Nutr Health Aging. 2018; 22:8–15. https://doi.org/10.1007/s12603-016-0854-3 [PubMed]
- 18. Watanabe D, Yoshida T, Watanabe Y, Yamada Y, Kimura M, Kyoto-Kameoka Study Gr. A U-Shaped Relationship Between the Prevalence of Frailty and Body Mass Index in Community-Dwelling Japanese Older Adults: The Kyoto-Kameoka Study. J Clin Med. 2020; 9:1367. https://doi.org/10.3390/jcm9051367 [PubMed]
- 19. Szanton SL, Seplaki CL, Thorpe RJ
Jr , Allen JK, Fried LP. Socioeconomic status is associated with frailty: the Women’s Health and Aging Studies. J Epidemiol Community Health. 2010; 64:63–67. https://doi.org/10.1136/jech.2008.078428 [PubMed] - 20. Etman A, Burdorf A, Van der Cammen TJ, Mackenbach JP, Van Lenthe FJ. Socio-demographic determinants of worsening in frailty among community-dwelling older people in 11 European countries. J Epidemiol Community Health. 2012; 66:1116–21. https://doi.org/10.1136/jech-2011-200027 [PubMed]
- 21. Hoogendijk EO, van Hout HP, Heymans MW, van der Horst HE, Frijters DH, Broese van Groenou MI, Deeg DJ, Huisman M. Explaining the association between educational level and frailty in older adults: results from a 13-year longitudinal study in the Netherlands. Ann Epidemiol. 2014; 24:538–44.e2. https://doi.org/10.1016/j.annepidem.2014.05.002 [PubMed]
- 22. Atkins JL, Jylhava J, Pedersen N, Magnusson P, Lu Y, Wang Y, Hagg S, Melzer D, Williams D, Pilling LC. A Genome-Wide Association Study of the Frailty Index Highlights Synaptic Pathways in Aging. medRxiv. 2019. https://doi.org/10.1101/19007559
- 23. van der Sluis S, Posthuma D, Dolan CV. A note on false positives and power in G × E modelling of twin data. Behav Genet. 2012; 42:170–86. https://doi.org/10.1007/s10519-011-9480-3 [PubMed]
- 24. Verhulst B, Prom-Wormley E, Keller M, Medland S, Neale MC. Type I Error Rates and Parameter Bias in Multivariate Behavioral Genetic Models. Behav Genet. 2019; 49:99–111. https://doi.org/10.1007/s10519-018-9942-y [PubMed]
- 25. Young AI. Solving the missing heritability problem. PLoS Genet. 2019; 15:e1008222. https://doi.org/10.1371/journal.pgen.1008222 [PubMed]
- 26. Wainschtein P, Jain D, Zheng Z, Cupples LA, Shadyab AH, McKnight B, Shoemaker BM, Mitchell BD, Psaty BM, Kooperberg C, Liu CT, et al. Recovery of trait heritability from whole genome sequence data. bioRxiv. 2021. https://doi.org/10.1101/588020
- 27. Chen J, Yu J. Sex Differences in Genetic and Environmental Influences on Adolescent Depressive Symptoms: A Meta-Analytic Review. Depress Res Treat. 2015; 2015:476238. https://doi.org/10.1155/2015/476238 [PubMed]
- 28. Hartvigsen J, Nielsen J, Kyvik KO, Fejer R, Vach W, Iachine I, Leboeuf-Yde C. Heritability of spinal pain and consequences of spinal pain: a comprehensive genetic epidemiologic analysis using a population-based sample of 15,328 twins ages 20-71 years. Arthritis Rheum. 2009; 61:1343–51. https://doi.org/10.1002/art.24607 [PubMed]
- 29. Lind MJ, Aggen SH, Kirkpatrick RM, Kendler KS, Amstadter AB. A Longitudinal Twin Study of Insomnia Symptoms in Adults. Sleep. 2015; 38:1423–30. https://doi.org/10.5665/sleep.4982 [PubMed]
- 30. Gordon EH, Hubbard RE. Do sex differences in chronic disease underpin the sex-frailty paradox? Mech Ageing Dev. 2019; 179:44–50. https://doi.org/10.1016/j.mad.2019.02.004 [PubMed]
- 31. Schousboe K, Willemsen G, Kyvik KO, Mortensen J, Boomsma DI, Cornes BK, Davis CJ, Fagnani C, Hjelmborg J, Kaprio J, De Lange M, Luciano M, Martin NG, et al. Sex differences in heritability of BMI: a comparative study of results from twin studies in eight countries. Twin Res. 2003; 6:409–21. https://doi.org/10.1375/136905203770326411 [PubMed]
- 32. Mekli K, Marshall A, Nazroo J, Vanhoutte B, Pendleton N. Genetic variant of Interleukin-18 gene is associated with the Frailty Index in the English Longitudinal Study of Ageing. Age Ageing. 2015; 44:938–42. https://doi.org/10.1093/ageing/afv122 [PubMed]
- 33. Inglés M, Mas-Bargues C, Gimeno-Mallench L, Cruz-Guerrero R, García-García FJ, Gambini J, Borrás C, Rodríguez-Mañas L, Viña J. Relation Between Genetic Factors and Frailty in Older Adults. J Am Med Dir Assoc. 2019; 20:1451–57. https://doi.org/10.1016/j.jamda.2019.03.011 [PubMed]
- 34. Akiyama M, Okada Y, Kanai M, Takahashi A, Momozawa Y, Ikeda M, Iwata N, Ikegawa S, Hirata M, Matsuda K, Iwasaki M, Yamaji T, Sawada N, et al. Genome-wide association study identifies 112 new loci for body mass index in the Japanese population. Nat Genet. 2017; 49:1458–67. https://doi.org/10.1038/ng.3951 [PubMed]
- 35. Locke AE, Kahali B, Berndt SI, Justice AE, Pers TH, Day FR, Powell C, Vedantam S, Buchkovich ML, Yang J, Croteau-Chonka DC, Esko T, Fall T, et al. Genetic studies of body mass index yield new insights for obesity biology. Nature. 2015; 518:197–206. https://doi.org/10.1038/nature14177 [PubMed]
- 36. Huibregtse BM, Newell-Stamper BL, Domingue BW, Boardman JD. Genes Related to Education Predict Frailty Among Older Adults in the United States. J Gerontol B Psychol Sci Soc Sci. 2021; 76:173–83. https://doi.org/10.1093/geronb/gbz092 [PubMed]
- 37. Belsky DW, Domingue BW, Wedow R, Arseneault L, Boardman JD, Caspi A, Conley D, Fletcher JM, Freese J, Herd P, Moffitt TE, Poulton R, Sicinski K, et al. Genetic analysis of social-class mobility in five longitudinal studies. Proc Natl Acad Sci USA. 2018; 115:E7275–84. https://doi.org/10.1073/pnas.1801238115 [PubMed]
- 38. van der Heide I, Wang J, Droomers M, Spreeuwenberg P, Rademakers J, Uiters E. The relationship between health, education, and health literacy: results from the Dutch Adult Literacy and Life Skills Survey. J Health Commun. 2013 (Suppl 1); 18:172–84. https://doi.org/10.1080/10810730.2013.825668 [PubMed]
- 39. Lastrucci V, Lorini C, Caini S, Bonaccorsi G, and Florence Health Literacy Research Group. Health literacy as a mediator of the relationship between socioeconomic status and health: A cross-sectional study in a population-based sample in Florence. PLoS One. 2019; 14:e0227007. https://doi.org/10.1371/journal.pone.0227007 [PubMed]
- 40. Mackian S. A Review of Health Seeking Behavior: Problems and Prospects. Heal Syst Dev Program. Manchester; 2003. http://r4d.dfid.gov.uk/pdf/outputs/healthsysdev_kp/05-03_health_seeking_behaviour.pdf.
- 41. Johnson W, Kyvik KO, Mortensen EL, Skytthe A, Batty GD, Deary IJ. Education reduces the effects of genetic susceptibilities to poor physical health. Int J Epidemiol. 2010; 39:406–14. https://doi.org/10.1093/ije/dyp314 [PubMed]
- 42. Li X, Ploner A, Karlsson IK, Liu X, Magnusson PK, Pedersen NL, Hägg S, Jylhävä J. The frailty index is a predictor of cause-specific mortality independent of familial effects from midlife onwards: a large cohort study. BMC Med. 2019; 17:94. https://doi.org/10.1186/s12916-019-1331-8 [PubMed]
- 43. Theou O, O’Connell MD, King-Kallimanis BL, O’Halloran AM, Rockwood K, Kenny RA. Measuring frailty using self-report and test-based health measures. Age Ageing. 2015; 44:471–77. https://doi.org/10.1093/ageing/afv010 [PubMed]
- 44. Barnes JC, Wright JP, Boutwell BB, Schwartz JA, Connolly EJ, Nedelec JL, Beaver KM. Demonstrating the validity of twin research in criminology. Criminology. 2014; 52:588–626. https://doi.org/10.1111/1745-9125.12049
- 45. Zagai U, Lichtenstein P, Pedersen NL, Magnusson PK. The Swedish Twin Registry: Content and Management as a Research Infrastructure. Twin Res Hum Genet. 2019; 22:672–80. https://doi.org/10.1017/thg.2019.99 [PubMed]
- 46. Lichtenstein P, Sullivan PF, Cnattingius S, Gatz M, Johansson S, Carlström E, Björk C, Svartengren M, Wolk A, Klareskog L, de Faire U, Schalling M, Palmgren J, Pedersen NL. The Swedish Twin Registry in the third millennium: an update. Twin Res Hum Genet. 2006; 9:875–82. https://doi.org/10.1375/183242706779462444 [PubMed]
- 47. Lichtenstein P, De Faire U, Floderus B, Svartengren M, Svedberg P, Pedersen NL. The Swedish Twin Registry: a unique resource for clinical, epidemiological and genetic studies. J Intern Med. 2002; 252:184–205. https://doi.org/10.1046/j.1365-2796.2002.01032.x [PubMed]
- 48. Neale MC, Maes HHM. Methodology for Genetic Studies of Twins and Families. Dordrecht: Kluwer Academic Publishers; 2004. http://ibgwww.colorado.edu/workshop2004/cdrom/HTML/book2004a.pdf.
- 49. Purcell S. Variance components models for gene-environment interaction in twin analysis. Twin Res. 2002; 5:554–71. https://doi.org/10.1375/136905202762342026 [PubMed]