



Introduction
As the most prevalent malignant solid tumor in the world, breast cancer has a devastating impact on the lives of women [1, 2]. With the advancement of medical technology and the rising popularity of early breast cancer screening, the 5-year survival rate for early breast cancer patients after surgery, radiotherapy, and chemotherapy can reach 90 percent. However, the late breast tumor by direct infiltration or with the blood and lymph flow, transfer to other tissues or organs, and growth result in a poor prognosis for advanced breast cancer patients, with a 5-year survival rate of only about 26%, which has a significant impact on patients’ survival time and quality [3–5]. Moreover, due to the lack of early tumor-specific indicators or negligible initial symptoms of breast cancer, patients are diagnosed at stage III or IV and even miss the optimal treatment period, resulting in varied degrees of physical and mental burden [6, 7]. The molecular mechanism behind the onset and progression of breast cancer is still unclear. Consequently, research into the pathogenesis is critical for the development of effective therapies for breast cancer.
miRNA is an endogenous single-stranded non-coding RNA molecule composed of more than 20 nucleotides [8]. MiRNA mainly plays the role of gene regulation at the post-transcriptional level, and plays the role of tumor suppressor gene or oncogene by targeting mRNA degradation or inhibiting its translation process [9]. MiRNAs can affect the molecular expression of tumor cells through a variety of ways, thus affecting the ability of cell adhesion, changing the cytoskeleton structure, and participating in the regulation of cell-extracellular matrix interaction [10, 11]. Some scholars have demonstrated that miRNA can affect tumor bioactivity by regulating the signaling pathway of target genes. Cancer-related miRNAs are usually divided into two types. The first type of carcinogenic miRNA is often highly expressed in tumors, where it can promote the incidence and development of tumors and play a crucial role in tumor phenotypic maintenance. By controlling cell proliferation, apoptosis, and other processes, the second class of tumor suppressor microRNA decreases the occurrence, development, and medication resistance of cancers. In malignant tumors, miRNAs can therefore play either synergistic or antagonistic roles [12, 13]. It has been reported that miR-100, miR-122, miR-145, and miR-205 are down-regulated in breast cancer [14–17], while miR-1228, miR-150, miR-155, and miR-330-3p are up-regulated in breast cancer [18–21]. Recent study has shown that miR-874-3p expression is aberrant in liver cancer, osteosarcoma, colon cancer, and ovarian cancer [22–25], but our data shows that the expression of miR-874-3p is lowered in breast cancer.
Voltage-dependent anion channel 1 (VDAC1) is primarily positioned in the outer mitochondrial membrane of all eukaryotes and is the fundamental link between mitochondria and cellular metabolism [26]. VDAC1 has been demonstrated to be related with a number of disorders in the current investigation, and it is significantly expressed in a variety of tumors [27]. VDAC1 supports their metabolism via the transfer of diverse metabolites and mitochondrial ATP binding, resulting in mitochondrial regulation of glycolytic flow through the TCA cycle and the action of ATP synthase to meet tumor demand for metabolites or metabolite precursors [28]. By interacting with the anti-apoptotic proteins Bcl-XL, Bcl-2, and HK, VDAC1 regulates cancer cell apoptosis and shields tumor cells from cell death [29]. In addition, hyperglycemia increases VDAC1 expression in pancreatic -cells and kidneys, and VDAC1 levels are elevated in mouse coronary endothelial cells isolated from diabetic mice, because glucose-stimulated insulin secretion is dependent on the production of ATP and other metabolites in mitochondria, and VDAC1 regulates energy and metabolism [30]. Consequently, VDAC1 is necessary for insulin secretion.
We detected miR-874-3p and VDAC1 expression in breast cancer cells and tissues in our investigation. The targeted regulatory link between miR-874-3p and VDAC1 was confirmed, along with the associations between miR-874-3p expression, patient clinicopathology, and prognosis.
Results
VDAC1 expression is elevated in breast cancer
To determine the expression of VDAC1 in human breast cancer, western blot was utilized to identify the expression of VDAC1 in human breast cancer and normal breast tissues. Western blotting revealed that VDAC1 protein expression was considerably higher in breast cancer tissues than in surrounding tissues (Figure 1A, 1B). Simultaneously, we examined the expression of VDAC1 in human breast cancer cells and human mammary epithelial cells, and the findings revealed that, compared with normal mammary epithelial cells HMEC, the expression of VDAC1 was higher in human breast cancer cells BT549 and MCF-7 (Figure 1C). By studying the TCGA database, we discovered that the expression of VDAC1 is elevated in breast cancer patients, and that these patients had a bad prognosis (Figure 1D, 1E). All of these results indicate that VDAC1 may be associated with the incidence of breast cancer.
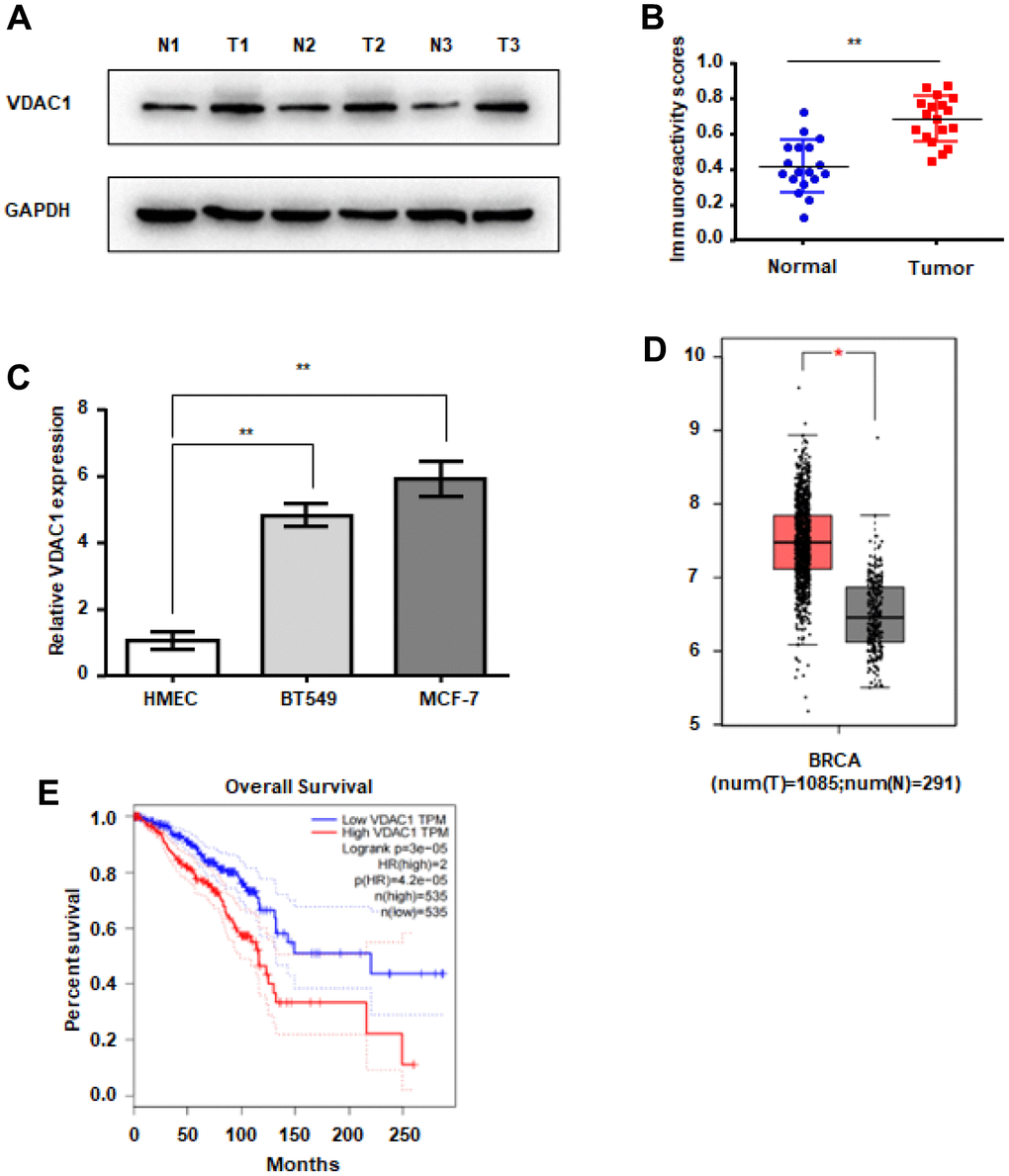
Figure 1. VDAC1 is highly expressed in breast cancer. (A, B) Analysis of VDAC1 expression in 18 breast cancer tissues and adjacent paired normal breast tissue samples. Representative western blotting images of VDAC1 levels in three breast cancer tissues and three normal breast tissues (A). VDAC1 and GAPDH protein levels were determined using ImageJ (B). (C) RT-qPCR was used to detect the mRNA expression of VDAC1 in normal breast cells and breast cancer cells. (D) Expression of VDAC1 in normal tissues and breast cancer tissues from the TCGA database. (E) Kaplan-Meier survival analysis of breast patients with positive or negative VDAC1 expression. The data are presented as the mean ± S.D. *P < 0.05, **P < 0.01.
VDAC1 promotes the proliferation, migration and invasion of breast cancer cells
Next, we evaluated the impact of VDAC1 on breast cancer in vitro by transfecting BT549 and McF-7 cells with VDAC1 overexpression plasmid plvx-VDAC1 and control plasmid plvx-Con, respectively. Overexpression of VDAC1 significantly boosted the proliferative potential of breast cancer cells compared to the control group, as determined by CCK8 (Figure 2A). In addition, EdU measurements revealed that in the VDAC1 overexpression group, the fraction of cells in the replication phase was considerably elevated (Figure 2B). The findings of the Transwell experiment indicated further that VDAC1 overexpression enhances the capacity of breast cancer cells to move and invade (Figure 2C, 2D). All of these data demonstrated that VDAC1 can enhance breast cancer malignancy in vitro.
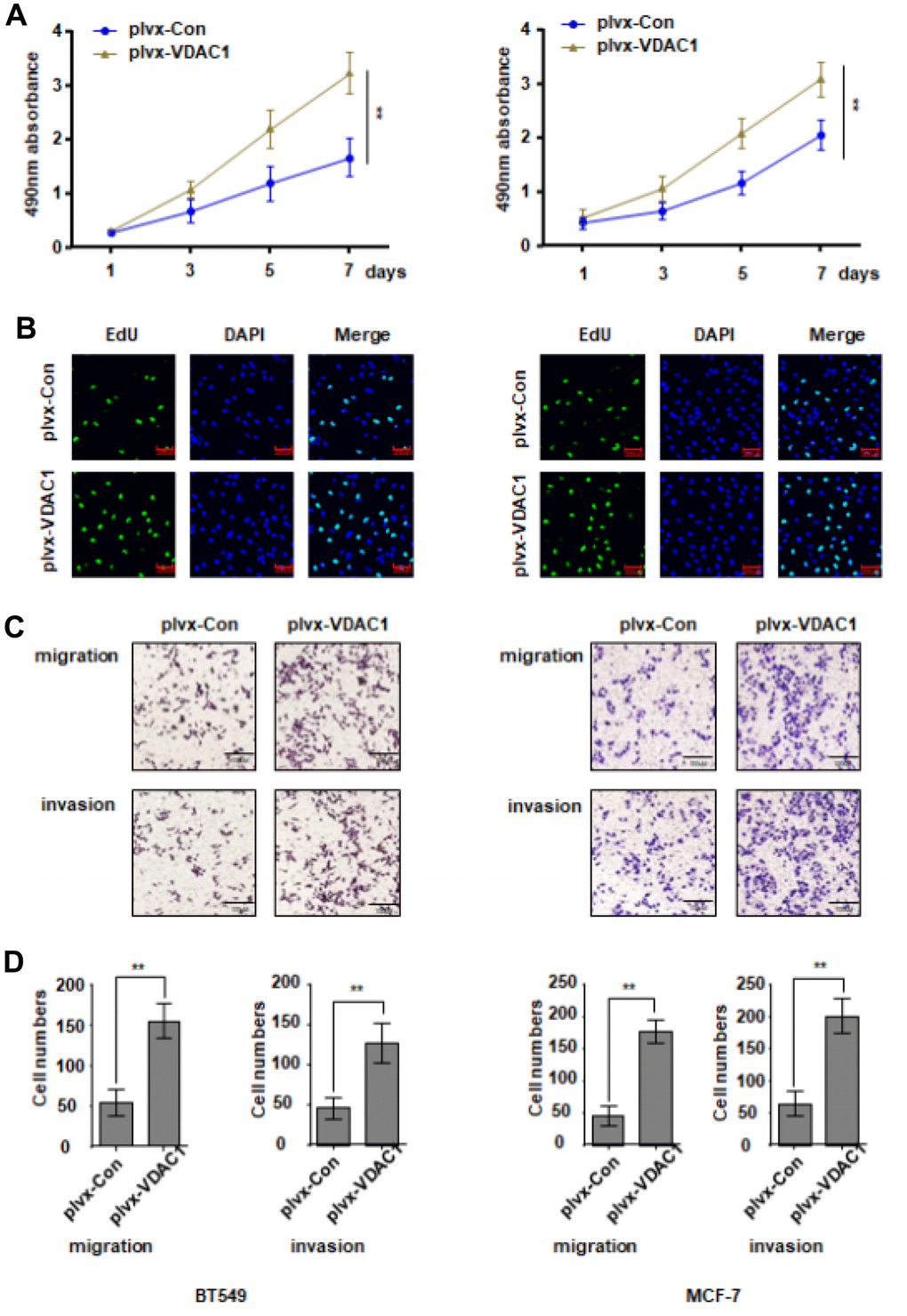
Figure 2. VDAC1 promotes the proliferation, migration and invasion of breast cancer. The VDAC1 overexpression plasmid plvx-VDAC1 or blank plasmid plvx-Con was transfected into BT549 and MCF-7 cells. (A, B) Cell proliferation was detected with CCK8 assay (A) and EDU assay (B). (C, D) Transwell assay was used to detect cell migration and invasion ability. The data are presented as the mean ± S.D. *P < 0.05, **P < 0.01.
VDAC1 promotes the occurrence and development of breast cancer in vivo
To investigate the influence of VDAC1 on breast cancer in vivo, MCF-7 cells stably overexpressed with VDAC1 and control cells were subcutaneously injected into immunodeficient mice to provide a subcutaneous tumor-bearing model of human breast cancer. The observation was started on the seventh day. Every four days, tumor growth and size were recorded. After 28 days, mice were killed and tumors were removed. The tumors generated in the VDAC1 overexpression group (plvx-VDAC1) were considerably larger than those in the control group (plvx-Con), as measured by tumor volume (Figure 3A–3C). In vitro and in vivo studies demonstrate that VDAC1 promotes breast cancer.
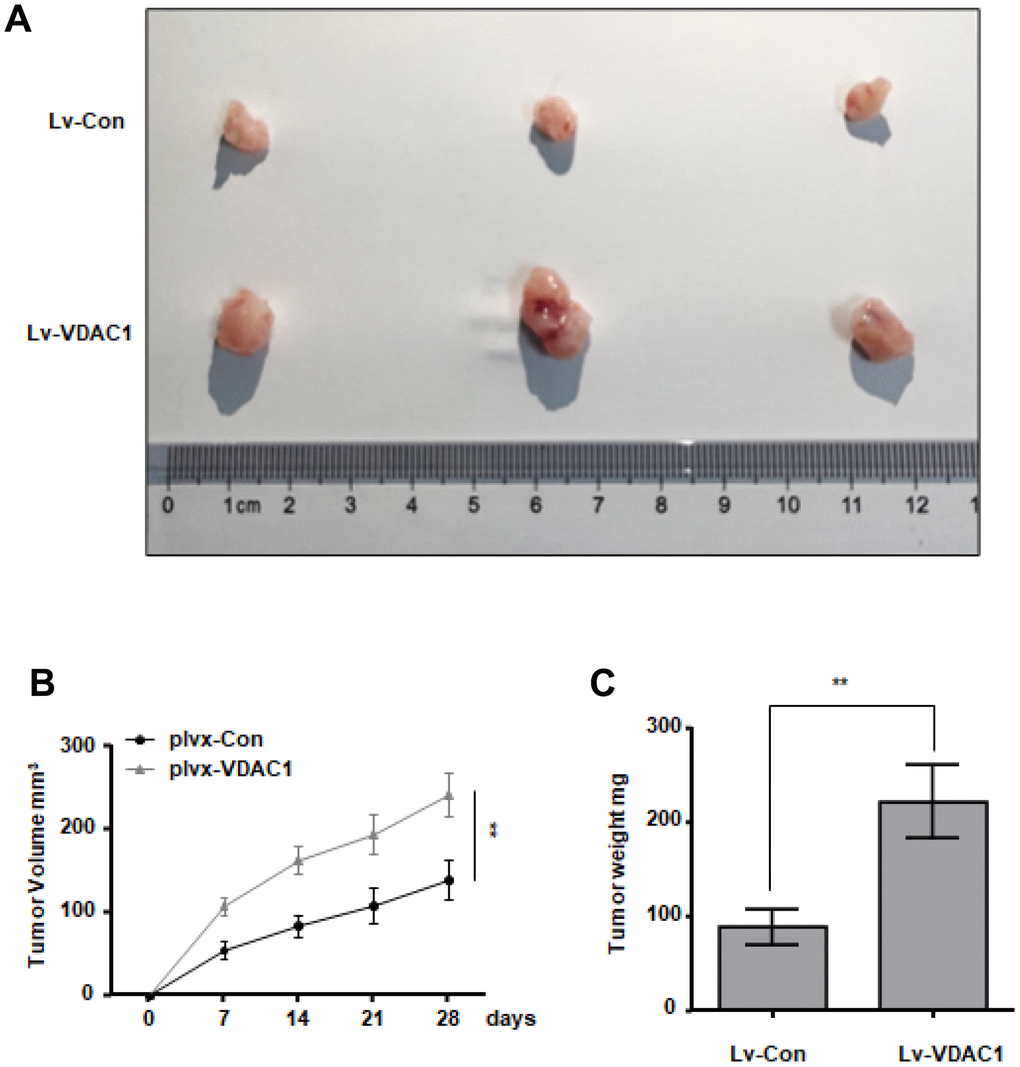
Figure 3. VDAC1 promotes breast cancer growth in vivo. Subcutaneous xenografts of MCF7 cells infected with VDAC1 overexpressing lentivirus or control lentivirus. (A) Images of the tumors at autopsy from nude mice are presented. (B, C) Tumor volumes (B) and average weight (C) of xenografted tumors were measured. Data represent the means ± S.D. **P < 0.01.
miR-874-3p directly binds to the VDAC1 3’-UTR
To explore the reasons for the abnormally high expression of VDAC1 in breast cancer, we predicted the miRNAs that bind to the 3’UTR region of VDAC1 mRNA through a bioinformatics website (Figure 4A). In accordance with our predictions, RT-qPCR detection revealed that only miR-874-3p was decreased in breast cancer (Figure 4B). Additionally, we discovered that miR-874-3p was downregulated in breast cancer tissues and was negatively linked with the expression of VDAC1 (Figure 4C, 4D). To establish the targeting interaction between miR-874-3p and VDAC1, luciferase reporter-tagged wild-type and mutant plasmids were generated (Figure 4E). The findings demonstrated that miR-874-3p mimics had no effect on the luciferase activity of mutant plasmids in breast cancer cells, but reduced the luciferase activity of wild-type plasmids (Figure 4F).
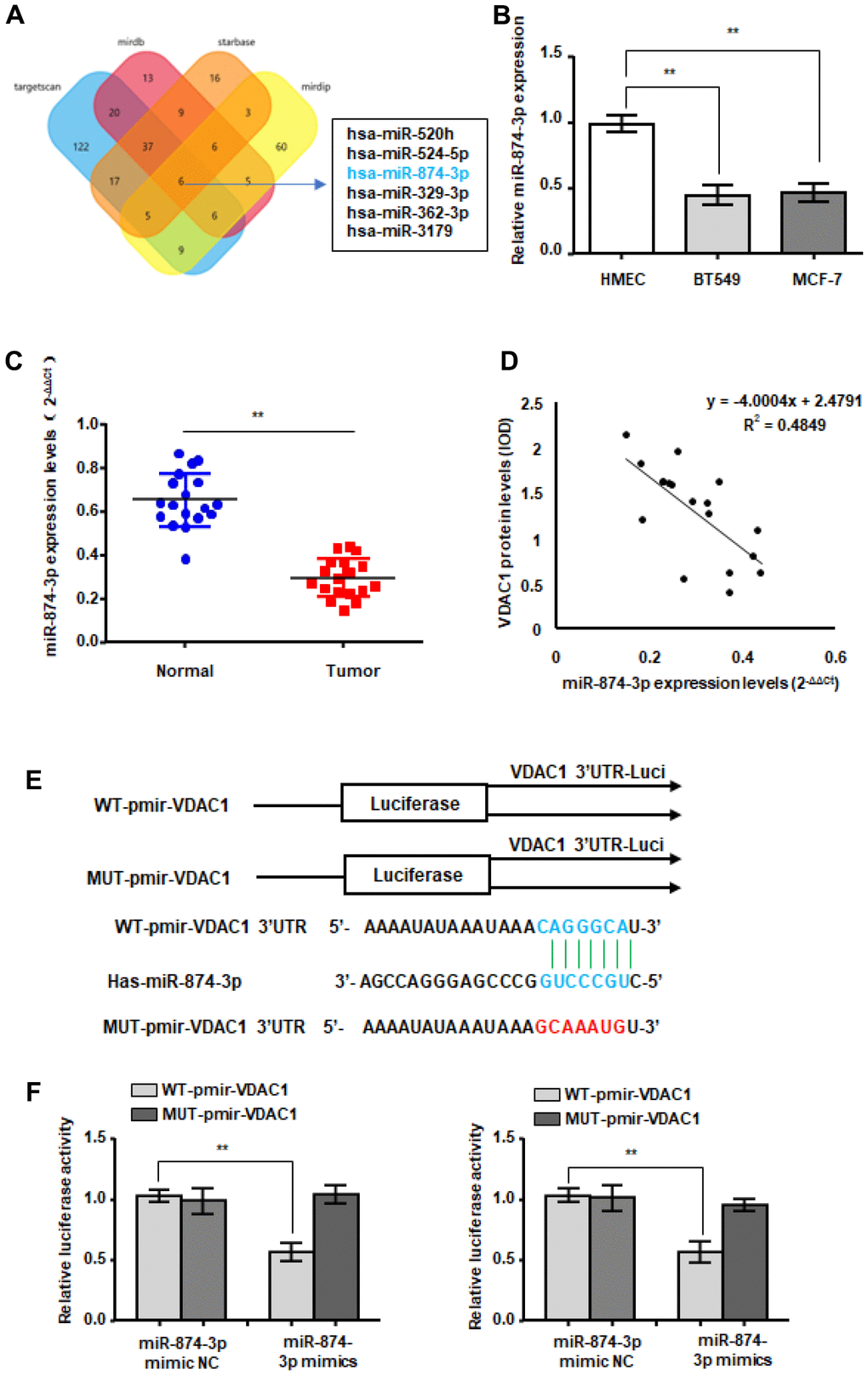
Figure 4. WTX is a direct target of miR-874-3p. (A) The four-way Venn diagram reveals the numbers of overlapping miRNAs obtained using four publicly available bioinformatics algorithms and the microarray-based VDAC1 signature. (B) RT-qPCR was used to detect the relative expression of miR-874-3p in normal breast cells and breast cancer cells. (C) RT-qPCR was used to detect the expression in 18 breast cancer tissues and adjacent paired normal breast tissue samples. (D) Correlation between miR-874-3p levels and VDAC1 levels in 18 breast cancer tissues. (E) Nucleotide predicted miR-874-3p-binding site in the VDAC1 mRNA 3′-UTR. (F) MCF-7 and BT549 cells were transfected with reporter plasmids containing WT-pmir-VDAC1 or MUT-pmir-VDAC1 and miR-874-3p mimic or miR-874-3p mimic NC, and luciferase activity was detected. Data represent the means ± S.D. **P < 0.01.
miR-874-3p inhibits the malignancy of breast cancer cells
During this period, breast cancer cells were transfected with miR-874-3p mimics or controls without mimics. Compared with the control group, the proliferation ability of MCF-7 and BT549 cells transfected with miR-874-3p mimic was significantly inhibited (Figure 5A, 5B). The Transwell experiment showed that transfection of miR-874-3p mimics made BT549 and MCF-7 cells less likely to move and invade compared to the blank control (Figure 5C, 5D).
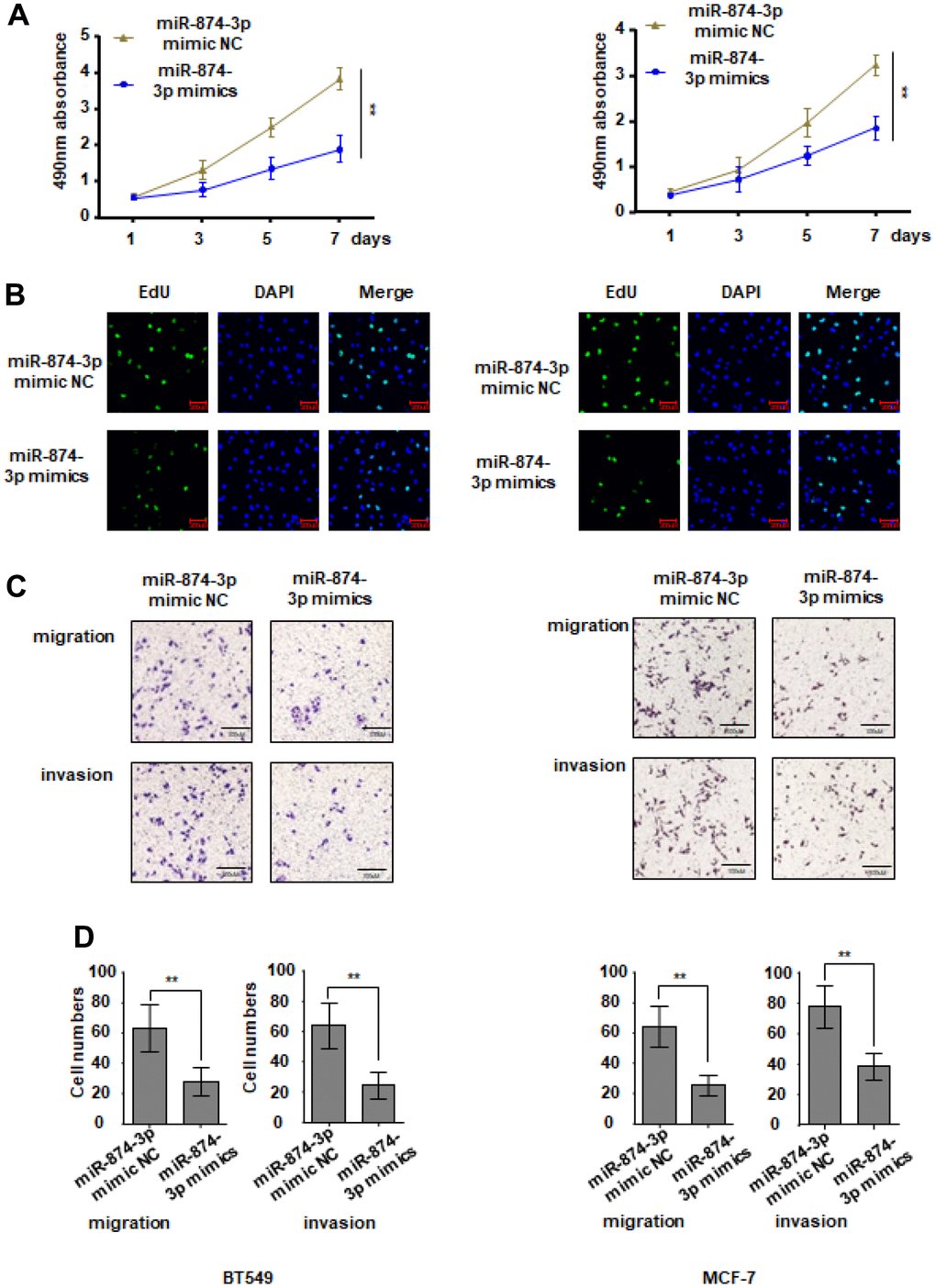
Figure 5. miR-874-3p inhibits proliferation, migration and invasion of breast cancer cells. MCF-7 and BT549 cells were transduced with miR-874-3p mimic NC or miR-874-3p mimics. (A, B) Cell proliferation was detected with CCK8 assay (A) and EDU assay (B). (C, D) Transwell assay was used to detect cell migration and invasion ability. The data are presented as the mean ± S.D. *P < 0.05, **P < 0.01.
miR-874-3p-VDAC1 axis regulates Wnt/β-catenin signaling in breast cells
The Wnt signaling pathway is a classic signaling pathway in tumors. Abnormal activation of wnt signaling often leads to tumorigenesis. Will VDAC1 activate wnt signaling? In MCF-7 and BT549 cells, VDAC1 can increase the activity of the β-catenin reporter gene (Figure 6A), although miR-874-3p has the opposite effect (Figure 6B). In addition, we discovered that the expression of β-catenin and downstream Cyclin D1 was reduced, while the expression of p-β-catenin was elevated, following transfection with miR-874-3p mimics (Figure 6C). These findings suggest that VDAC1 enhances the onset and progression of breast cancer via stimulating the wnt signaling pathway.
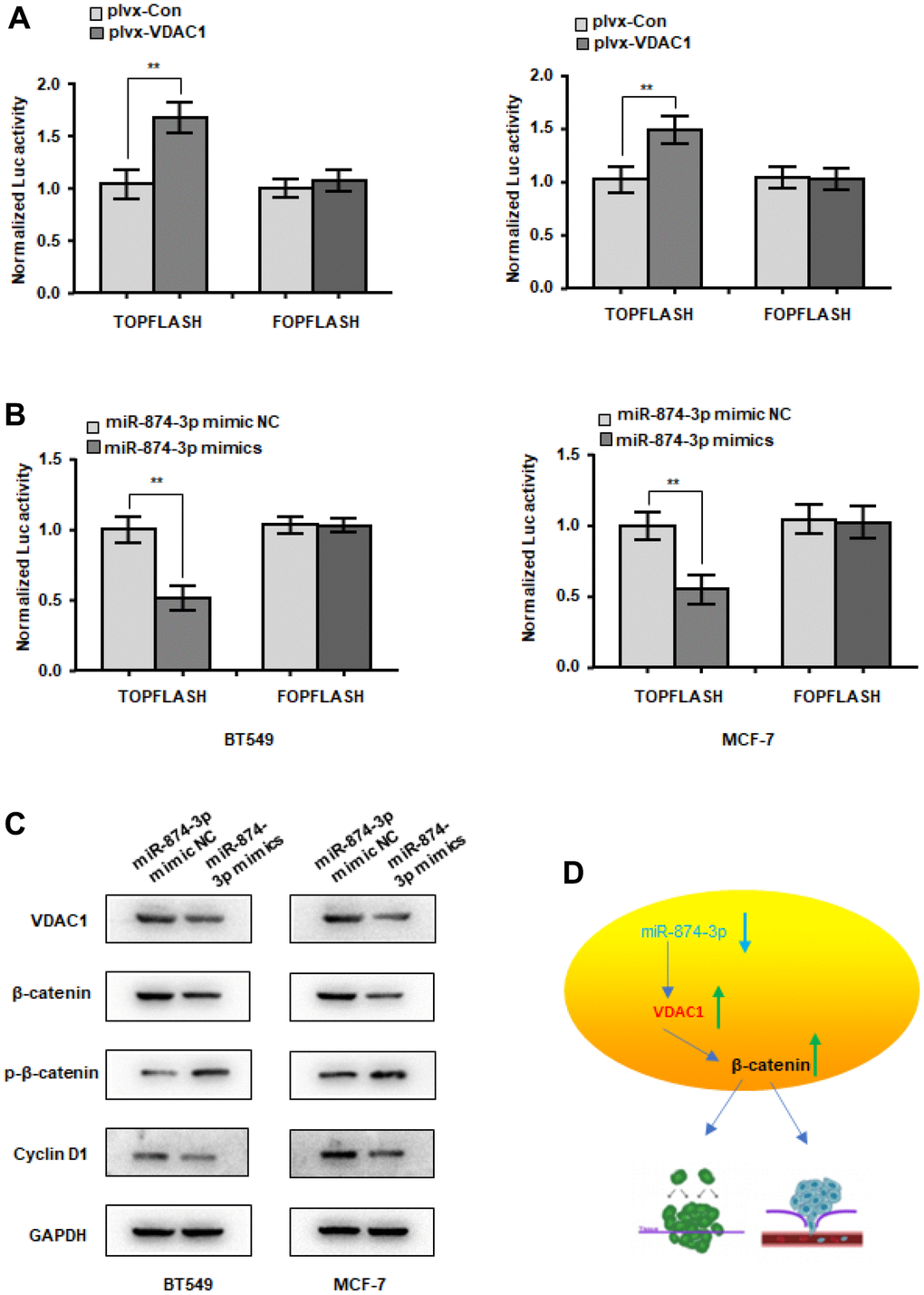
Figure 6. miR-874-3p-VDAC1 axis regulates Wnt/β-catenin signaling in breast cancer cells. (A) β-catenin reporter assay in MCF-7 and BT549 cells with VDAC1 overexpression (A) or miR-874-3p overexpression (B). (C) Effects of miR-874-3p on protein levels of total β-catenin, phosphorylated β-catenin (Ser33/37/Thr41) and cyclin D1. (D) Schematic of the mechanism. The data are presented as the mean ± S.D. *P < 0.05, **P < 0.01.
Discussion
Breast cancer is a common malignant tumor, accounting for 20% of female malignant tumors, the latest research reports show that breast cancer has become the highest incidence of cancer in the world, seriously affecting the survival of patients [31]. Our previous study discovered a link between VDAC1 and invasion and metastasis of breast cancer cell proliferation, but VDAC1 in the molecular regulation mechanism of breast cancer is unclear, so the VDAC1 in the incidence of breast cancer, development of molecular mechanism, and study the related regulation can help to find new breast cancer treatment targets, and promote the development of precision medical treatment of breast cancer. This gene has been discovered to be overexpressed in numerous malignancies, including lung cancer, cervical cancer, gastric cancer, pancreatic cancer, and laryngeal cancer [32–36]. In 2012, Brahimi-Horn et al., in their research on the drug resistance of lung cancer, found that cell lines with high expression of VDAC1 gene had higher resistance to apoptosis induced by staurosporine and etoposide, and silenced VDAC1 gene. Afterwards, the cell line regained its sensitivity to the above-mentioned drugs. In addition, high expression levels of VDAC1 gene can be detected in advanced lung cancer and tissues with larger lung cancer. It is concluded that VDAC1 may become a novel marker for early diagnosis and prognosis evaluation of lung cancer [37]. In 2010, Lan et al. found in their study on the mechanism of apoptosis in gastric cancer that the up-regulation of complexes and the down-regulation of proteins play a key role in the process of mediated apoptosis [34]. These results suggest that it is necessary to explore the role of VDAC1 in cancer.
Non-coding RNAs (ncRNAs) are RNA molecules that are produced by transcription but do not code for proteins; these molecules mostly consist of LncRNAs, CircRNAs, miRNAs, etc. At the post-transcriptional level, they play a significant role in gene expression regulation and epigenetic regulation [38]. This research investigated the function of microRNAs in the onset and progression of breast cancer. miRNAs are directly associated to the advancement of breast cancer, and they play varied roles at different phases of breast cancer development due to their involvement in numerous cancer processes. We believe miRNAs are involved in the regulation of malignant proliferation, escape from growth inhibition, cellular senescence, and genomic instability, despite the fact that their functions in numerous cancer processes have not been demonstrated. Several microRNAs have been discovered to be inappropriately expressed in breast tumors during the earliest stages of malignancy. VDAC1 is a miR-874-3p target gene, and miR-874-3p inhibits its expression in breast cancer cells. miR-874-3p has been shown to affect the expression of a variety of target genes, including GDPD5, FOXM1, FAM84A, and ADAM19 [22, 24, 39, 40]. However, no articles have reported the targeting link between miR-874-3p and VDAC1 and the role of miR-874-3p in breast cancer.
According to this study, miR-874-3p suppresses the proliferation, migration, and invasion of breast cancer cells as well as the expression of VDAC1, regulates the cell cycle, and promotes the death of breast cancer cells, as shown in Figure 6D. Therefore, miR-874-3p may be used as a biomarker for the early diagnosis of breast cancer. VDAC1 was validated as a target gene of miR-874-3p in the breast cancer molecular regulation mechanism, and its molecular regulation mechanism was investigated. We will improve the molecular regulatory mechanism of VDAC1 in the future. The findings imply that miR-874-3p may also serve as a novel therapeutic target for breast cancer, shedding fresh light on the investigation of miRNA expression profiles in breast cancer.
Materials and Methods
Tissue specimens
The breast cancer tissue specimens used in this study were obtained from patients who underwent breast cancer resection in Yueyang People’s Hospital.
Cell lines and cell culture
The BT549 and MCF-7 breast cancer cell lines were obtained from the Shanghai Cell Bank of the Chinese Academy of Sciences (Shanghai, China). MCF-7 was grown in DMEM medium (BI, Israel) with 10% fetal bovine serum, whereas BT549 was grown in RPMI 1640 medium (BI, Israel) with 10% fetal bovine serum. All cells were grown at 37° C in an incubator containing 5% CO2.
To construct cell lines stably overexpressing VDAC1, we infected MCF-7 cells with lentivirus containing VDAC1 and used 500 ng/mL puromycin for resistance selection. Cells were harvested a week later for verification.
qRT-PCR
Firstly, Trizol was added to the collected cells or tissues for mixing, then a quarter volume of chloroform was added, and then the supernatant was centrifuged, and the same volume of isopropanol was added for RNA deposition, all at 4° C. 1ugRNA was added to gDNA Wiper Mix solution to remove genomic DNA, and then HiScript qRT superMix solution was added for reverse transcription to obtain cDNA. Then take 2 μL of cDNA and add qPCR SuperMix solution for qRT-PCR. Primer sequences are as follows: VDAC1 forward, 5′-GGTGCTCTGGTGCTAGGTTA-3′ and reverse, 5′-CAGCGGTCTCCAACTTCTTG-3′ and GAPDH forward, 5′-ACCCACTCCTCCACCTTTGAC-3′ and reverse, 5′-TGTTGCTGTAGCCAAATTCGTT-3′.
Plasmids, miRNAs, and transfection
The plvx-VDAC1 plasmid and miR-874-3p mimics were obtained from Origene (Beijing, China). miR-874-3p mimics sequence: 5′-ACUGCGUUGAAACAUGGGUA-3′, and the sequence of the miR-mimic NC was 5′-UUGAGGCUUCAAUCGACGUTT-3′. All transfections were carried out utilizing Lipo2000 per the directions. First, cells were seeded in 6-well plates, then transfected 24 hours later, followed by additional functional testing 24 hours following transfection.
Transwell invasion and migration assays
36 hours after transfection, 5 × 104 cells were planted to the transwell chamber, the cells were incubated in the incubator for 36 hours, washed with PBS solution, cleaned with sterile cotton swabs, fixed with methanol, stained with crystal violet, and dried in a ventilation cabinet. Under an inverted microscope, photographs were taken and numbered. In preparation of the cell invasion assay, matrigel was spread in the Transwell chamber and dried overnight.
CCK8 assay
Transfected cells were inoculated into 96-well plates and absorbance was measured at 0, 1, 3, 5 and 7 days, respectively. Before each measurement, 10 μL CCK8 (Sigma) solution was added to each well and incubated for 2 h. At a wavelength of 450 nm, the absorbance of each well was measured.
EdU assay
The transfected cells were counted and then seeded on cell slides, EdU working solution was added at 24 hours and incubated for 3 hours, fixed with paraformaldehyde, and then EdU reaction solution was added, and photographed and analyzed under a fluorescence microscope.
Western blotting
The cells or tissues were treated with RIPA lysate, properly mixed, centrifuged, and the supernatant was added to electrophoresis loading buffer. The separated proteins in the gel were transferred to PVDF membranes, coated with primary antibody overnight, incubated with secondary antibody the next day, and then exposed and developed using a gel imaging system. Primary antibodies used were anti-VDAC1 (A19707; Abclonal), anti-β-Catenin (A0316; Abclonal), phosphorylated β-catenin (AP1076; Abclonal), anti-Cyclin D1 (A0310; Abclonal), anti-GAPDH (A19056; Abclonal).
Animal studies
In this study, 10 Balb/ C female nude mice (4 weeks old) were purchased from Beijing Spaifu Company and kept under SPF (temperature 25° C, humidity 55%). The Yueyang People’s Hospital Animal Ethics Committee authorized the trial. Mice were randomly assigned into two groups corresponding to the MCF-7-control group and MCF-7-overexpression group, and then the cells in different treatment groups were mixed with matrix glue at 1:1 and adjusted to a cell suspension of 5×107/ mL cells. Mice were subcutaneously sown with cells using a standard 1 ml syringe (100μl, 5×106 cells per mouse).
Statistical analysis
This study repeated each experiment at least three times. GraphPad Prism 8.0 was utilized to process and assess the experimental data. The data are given as the standard deviation of the mean. Student’s t-test, chi-square test, and Kaplan Meier analysis were utilized in the statistical study. P<0.05 signifies that the outcomes of the experiment are statistically significant.
Author Contributions
Z.Y.Y. conceived and devised the study. Z.Y.Y., W.L., H.S.Y. and L.C. designed the experiments and analysis. W.L., S.H. and Z.Y.Y. performed the experiments and analyzed the data. Z.W.W. and Z.Y.Y. contributed reagents and materials. H.S.Y. and L.C. supervised the research and wrote the manuscript. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Ethical Statement and Consent
Participants signed the informed consent form, and the studies were conducted with the approval of the Yueyang People’s Hospital Ethics Committee (YY369841). All animal experiments were approved by Institutional Ethics Committee of Yueyang People’s Hospital (AB173584).
Funding
This work was financially supported by Hunan Provincial Health Commission Scientific Research Fund Project (202204015494).
References
- 1. Liang Y, Zhang H, Song X, Yang Q. Metastatic heterogeneity of breast cancer: Molecular mechanism and potential therapeutic targets. Semin Cancer Biol. 2020; 60:14–27. https://doi.org/10.1016/j.semcancer.2019.08.012 [PubMed]
- 2. Chodosh LA. Breast cancer: current state and future promise. Breast Cancer Res. 2011; 13:113. https://doi.org/10.1186/bcr3045 [PubMed]
- 3. Fahad Ullah M. Breast Cancer: Current Perspectives on the Disease Status. Adv Exp Med Biol. 2019; 1152:51–64. https://doi.org/10.1007/978-3-030-20301-6_4 [PubMed]
- 4. Rossi L, Mazzara C, Pagani O. Diagnosis and Treatment of Breast Cancer in Young Women. Curr Treat Options Oncol. 2019; 20:86. https://doi.org/10.1007/s11864-019-0685-7 [PubMed]
- 5. McDonald ES, Clark AS, Tchou J, Zhang P, Freedman GM. Clinical Diagnosis and Management of Breast Cancer. J Nucl Med. 2016 (Suppl 1); 57:9S–16S. https://doi.org/10.2967/jnumed.115.157834 [PubMed]
- 6. Veronesi U, Boyle P, Goldhirsch A, Orecchia R, Viale G. Breast cancer. Lancet. 2005; 365:1727–41. https://doi.org/10.1016/S0140-6736(05)66546-4 [PubMed]
- 7. Low SK, Zembutsu H, Nakamura Y. Breast cancer: The translation of big genomic data to cancer precision medicine. Cancer Sci. 2018; 109:497–506. https://doi.org/10.1111/cas.13463 [PubMed]
- 8. Chen L, Heikkinen L, Wang C, Yang Y, Sun H, Wong G. Trends in the development of miRNA bioinformatics tools. Brief Bioinform. 2019; 20:1836–52. https://doi.org/10.1093/bib/bby054 [PubMed]
- 9. Lu TX, Rothenberg ME. MicroRNA. J Allergy Clin Immunol. 2018; 141:1202–7. https://doi.org/10.1016/j.jaci.2017.08.034 [PubMed]
- 10. Saliminejad K, Khorram Khorshid HR, Soleymani Fard S, Ghaffari SH. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J Cell Physiol. 2019; 234:5451–65. https://doi.org/10.1002/jcp.27486 [PubMed]
- 11. Liu B, Li J, Cairns MJ. Identifying miRNAs, targets and functions. Brief Bioinform. 2014; 15:1–19. https://doi.org/10.1093/bib/bbs075 [PubMed]
- 12. Hill M, Tran N. miRNA interplay: mechanisms and consequences in cancer. Dis Model Mech. 2021; 14:dmm047662. https://doi.org/10.1242/dmm.047662 [PubMed]
- 13. He B, Zhao Z, Cai Q, Zhang Y, Zhang P, Shi S, Xie H, Peng X, Yin W, Tao Y, Wang X. miRNA-based biomarkers, therapies, and resistance in Cancer. Int J Biol Sci. 2020; 16:2628–47. https://doi.org/10.7150/ijbs.47203 [PubMed]
- 14. Xie H, Xiao R, He Y, He L, Xie C, Chen J, Hong Y. MicroRNA-100 inhibits breast cancer cell proliferation, invasion and migration by targeting FOXA1. Oncol Lett. 2021; 22:816. https://doi.org/10.3892/ol.2021.13077 [PubMed]
- 15. Wang B, Wang H, Yang Z. MiR-122 inhibits cell proliferation and tumorigenesis of breast cancer by targeting IGF1R. PLoS One. 2012; 7:e47053. https://doi.org/10.1371/journal.pone.0047053 [PubMed]
- 16. Zou C, Xu Q, Mao F, Li D, Bian C, Liu LZ, Jiang Y, Chen X, Qi Y, Zhang X, Wang X, Sun Q, Kung HF, et al. MiR-145 inhibits tumor angiogenesis and growth by N-RAS and VEGF. Cell Cycle. 2012; 11:2137–45. https://doi.org/10.4161/cc.20598 [PubMed]
- 17. Zhang H, Fan Q. MicroRNA-205 inhibits the proliferation and invasion of breast cancer by regulating AMOT expression. Oncol Rep. 2015; 34:2163–70. https://doi.org/10.3892/or.2015.4148 [PubMed]
- 18. Lin L, Liu D, Liang H, Xue L, Su C, Liu M. MiR-1228 promotes breast cancer cell growth and metastasis through targeting SCAI protein. Int J Clin Exp Pathol. 2015; 8:6646–55. [PubMed]
- 19. Huang S, Chen Y, Wu W, Ouyang N, Chen J, Li H, Liu X, Su F, Lin L, Yao Y. miR-150 promotes human breast cancer growth and malignant behavior by targeting the pro-apoptotic purinergic P2X7 receptor. PLoS One. 2013; 8:e80707. https://doi.org/10.1371/journal.pone.0080707 [PubMed]
- 20. Shaker O, Maher M, Nassar Y, Morcos G, Gad Z. Role of microRNAs -29b-2, -155, -197 and -205 as diagnostic biomarkers in serum of breast cancer females. Gene. 2015; 560:77–82. https://doi.org/10.1016/j.gene.2015.01.062 [PubMed]
- 21. Mesci A, Huang X, Taeb S, Jahangiri S, Kim Y, Fokas E, Bruce J, Leong HS, Liu SK. Targeting of CCBE1 by miR-330-3p in human breast cancer promotes metastasis. Br J Cancer. 2017; 116:1350–7. https://doi.org/10.1038/bjc.2017.105 [PubMed]
- 22. Feng X, Xue H, Guo S, Chen Y, Zhang X, Tang X. MiR-874-3p suppresses cell proliferation and invasion by targeting ADAM19 in nasopharyngeal carcinoma. Panminerva Med. 2021; 63:238–9. https://doi.org/10.23736/S0031-0808.19.03682-6 [PubMed]
- 23. Liu WG, Zhuo L, Lu Y, Wang L, Ji YX, Guo Q. miR-874-3p inhibits cell migration through targeting RGS4 in osteosarcoma. J Gene Med. 2020; 22:e3213. https://doi.org/10.1002/jgm.3213 [PubMed]
- 24. Wang Y, Chen H, Wei X. Circ_0007142 downregulates miR-874-3p-mediated GDPD5 on colorectal cancer cells. Eur J Clin Invest. 2021; 51:e13541. https://doi.org/10.1111/eci.13541 [PubMed]
- 25. Xia B, Lin M, Dong W, Chen H, Li B, Zhang X, Hou Y, Lou G. Upregulation of miR-874-3p and miR-874-5p inhibits epithelial ovarian cancer malignancy via SIK2. J Biochem Mol Toxicol. 2018; 32:e22168. https://doi.org/10.1002/jbt.22168 [PubMed]
- 26. Shoshan-Barmatz V, Nahon-Crystal E, Shteinfer-Kuzmine A, Gupta R. VDAC1, mitochondrial dysfunction, and Alzheimer’s disease. Pharmacol Res. 2018; 131:87–101. https://doi.org/10.1016/j.phrs.2018.03.010 [PubMed]
- 27. Magrì A, Reina S, De Pinto V. VDAC1 as Pharmacological Target in Cancer and Neurodegeneration: Focus on Its Role in Apoptosis. Front Chem. 2018; 6:108. https://doi.org/10.3389/fchem.2018.00108 [PubMed]
- 28. Shoshan-Barmatz V, De S. Mitochondrial VDAC, the Na+/Ca2+ Exchanger, and the Ca2+ Uniporter in Ca2+ Dynamics and Signaling. Adv Exp Med Biol. 2017; 981:323–47. https://doi.org/10.1007/978-3-319-55858-5_13 [PubMed]
- 29. Shoshan-Barmatz V, Krelin Y, Chen Q. VDAC1 as a Player in Mitochondria-Mediated Apoptosis and Target for Modulating Apoptosis. Curr Med Chem. 2017; 24:4435–46. https://doi.org/10.2174/0929867324666170616105200 [PubMed]
- 30. Zhang E, Mohammed Al-Amily I, Mohammed S, Luan C, Asplund O, Ahmed M, Ye Y, Ben-Hail D, Soni A, Vishnu N, Bompada P, De Marinis Y, Groop L, et al. Preserving Insulin Secretion in Diabetes by Inhibiting VDAC1 Overexpression and Surface Translocation in β Cells. Cell Metab. 2019; 29:64–77.e6. https://doi.org/10.1016/j.cmet.2018.09.008 [PubMed]
- 31. Tray N, Taff J, Adams S. Therapeutic landscape of metaplastic breast cancer. Cancer Treat Rev. 2019; 79:101888. https://doi.org/10.1016/j.ctrv.2019.08.004 [PubMed]
- 32. Zerbib E, Arif T, Shteinfer-Kuzmine A, Chalifa-Caspi V, Shoshan-Barmatz V. VDAC1 Silencing in Cancer Cells Leads to Metabolic Reprogramming That Modulates Tumor Microenvironment. Cancers (Basel). 2021; 13:2850. https://doi.org/10.3390/cancers13112850 [PubMed]
- 33. Zhang C, Hua Y, Qiu H, Liu T, Long Q, Liao W, Qiu J, Wang N, Chen M, Shi D, Yan Y, Xie C, Deng W, et al. KMT2A regulates cervical cancer cell growth through targeting VDAC1. Aging (Albany NY). 2020; 12:9604–20. https://doi.org/10.18632/aging.103229 [PubMed]
- 34. Lan CH, Sheng JQ, Fang DC, Meng QZ, Fan LL, Huang ZR. Involvement of VDAC1 and Bcl-2 family of proteins in VacA-induced cytochrome c release and apoptosis of gastric epithelial carcinoma cells. J Dig Dis. 2010; 11:43–9. https://doi.org/10.1111/j.1751-2980.2009.00412.x [PubMed]
- 35. Wang W, Zhang T, Zhao W, Xu L, Yang Y, Liao Q, Zhao Y. A Single Talent Immunogenic Membrane Antigen and Novel Prognostic Predictor: voltage-dependent anion channel 1 (VDAC1) in Pancreatic Cancer. Sci Rep. 2016; 6:33648. https://doi.org/10.1038/srep33648 [PubMed]
- 36. Feng X, Ching CB, Chen WN. EBV up-regulates cytochrome c through VDAC1 regulations and decreases the release of cytoplasmic Ca2+ in the NPC cell line. Cell Biol Int. 2012; 36:733–8. https://doi.org/10.1042/CBI20110368 [PubMed]
- 37. Brahimi-Horn MC, Ben-Hail D, Ilie M, Gounon P, Rouleau M, Hofman V, Doyen J, Mari B, Shoshan-Barmatz V, Hofman P, Pouysségur J, Mazure NM. Expression of a truncated active form of VDAC1 in lung cancer associates with hypoxic cell survival and correlates with progression to chemotherapy resistance. Cancer Res. 2012; 72:2140–50. https://doi.org/10.1158/0008-5472.CAN-11-3940 [PubMed]
- 38. Anastasiadou E, Jacob LS, Slack FJ. Non-coding RNA networks in cancer. Nat Rev Cancer. 2018; 18:5–18. https://doi.org/10.1038/nrc.2017.99 [PubMed]
- 39. Zhang Z, Yang W, Li N, Chen X, Ma F, Yang J, Zhang Y, Chai X, Zhang B, Hou X, Luo S, Hua Y. LncRNA MCF2L-AS1 aggravates proliferation, invasion and glycolysis of colorectal cancer cells via the crosstalk with miR-874-3p/FOXM1 signaling axis. Carcinogenesis. 2021; 42:263–71. https://doi.org/10.1093/carcin/bgaa093 [PubMed]
- 40. Ding Y, Wu L, Zhuang X, Cai J, Tong H, Si Y, Zhang H, Wang X, Shen M. The direct miR-874-3p-target FAM84A promotes tumor development in papillary thyroid cancer. Mol Oncol. 2021; 15:1597–614. https://doi.org/10.1002/1878-0261.12941 [PubMed]