



Introduction
Most proteins undergo several modifications after synthesis, encompassing acetylation, methylation, phosphorylation, glycosylation, and ubiquitination [1–4]. Numerous signaling pathways are influenced by post-translational modification via the ubiquitination mechanism, which regulates target proteins’ location and/or activity. The ubiquitination process relies on three specific enzymes known as Ubiquitin enzymes (UBEs) that facilitate the covalent attachment of ubiquitin molecules to target proteins. Three key enzymes are ubiquitin-activating enzyme (E1), ubiquitin-conjugating enzyme (E2), and ubiquitin ligase or E3 ubiquitin ligase (E3). The target proteins first interact with E1 and E2, then selectively interact with E3 to regulate the specificity of substrates. Different forms of ubiquitination [5, 6], including monoubiquitination, polyubiquitination, and branching ubiquitination, are essential for intracellular activities and are engaged in various cellular functions [7], such as controlling the cell cycle and responding to DNA damage. Additionally, ubiquitination significantly affects many human diseases by regulating cell division, proliferation, and apoptosis [8, 9].
Ubiquitination is an essential process for both genesis and apoptosis; therefore, any disturbance in this process could potentially lead to cancer development [10–12]. Deubiquitinating enzymes (DUBs) may reverse the ubiquitination process of targeted proteins [13–18]. DUBs work by removing ubiquitin chains from their target proteins, which are vital for the stability or activity of the proteins [19, 20]. So far, there are about 100 kinds of DUBs in human genome [21], which can be divided into five families based on the organization of their catalytic domain [22]: (1) Ubiquitin-specific proteases (USPs), (2) ovarian tumor proteases (OTUs), (3) Ubiquitin C-terminal hydrolases (UCHs), (4) Machado-Joseph domain proteases (MJDs), and (5) JAB1/MNP/MOV34 metalloproteases (JAMMs).
The USPs family is the most recognized group of DUBs, characterized by a wide range of structural and functional variability [23]. The distinguishing features of USPs include the unique catalytic core, known as the histidine and cysteine boxes, as well as the presence of zinc finger domains, ubiquitin-binding domains, or ubiquitin-like domains at the N and/or C-termini of the catalytic domain [21]. Protein degradation or oversynthesis is associated with malignancy’s development, spread, or growth. In this regard, a growing body of research has shown that USPs exert an influence on cancer progression through their ability to regulate and promote the growth of cancerous cells [12, 23, 24] (Figure 1). Previous studies have connected USPs, such as USP7 [25], USP21, USP22, USP33 [26], USP39, USP54 [27], and others, to carcinogenesis in several cancer types. While USP7 affects the cell cycle of breast cancer, which regulates its growth [28], USP9X influences breast cancer growth by influencing nuclear replication [29]. Additionally, USP9X controls mitosis, cell death, and treatment resistance in aggressive B-cell lymphomas [30]. Studies have indicated that the pathophysiology of Von Hippel-Lindau Syndrome (VHL) is linked to USP20 and USP33 [31–33]. The association between USPs and cell signaling pathways in malignancies have been demonstrated in previous studies.
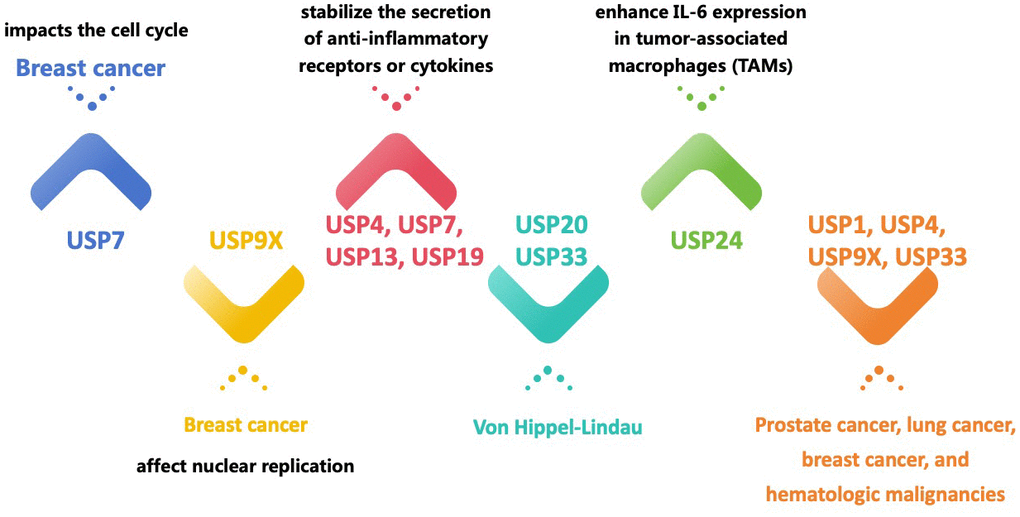
Figure 1. Different USPs regulate and promote the growth of cancerous cells and other diseases.
Prominent signaling pathways implicated in the initiation and advancement of cancer, namely p53, NF-B, Wnt, and TGF-β, are impacted by the activities of USPs, including USP2, USP4, USP5, USP10, USP11, USP15, USP29, and USP34 [34]. Furthermore, USPs can control cytokine release patterns, with USP24 increasing IL-6 expression in tumor-associated macrophages (TAMs), whereas USP4, USP7, USP13, and USP19 can stabilize the release of anti-inflammatory receptors or cytokines [32–35]. It is worth mentioning that different types of cancer display divergent reactions to the identical USP protein. For instance, although USP24 inhibits the proliferation of tumor cells, it promotes the spread of metastases in other types of malignancies [4, 35]. In terms of treatment, USPs can be used as cancer therapeutic targets, such as inhibitors of USP1, USP4, USP7, USP9X, and USP33 for prostate cancer, lung cancer, breast cancer, and hematological malignancies, among others [26–30].
Nevertheless, the biological mechanisms of various proteins in the USPs family, such as USP43, remain unclear as limited research has been conducted on its role in malignancies. Published studies have explored the connection between USP43 and various types of cancer, including osteosarcoma, non-small cell lung cancer, lung squamous cell carcinoma, breast cancer, and colorectal cancer. These studies have shed light on the specific molecular processes and signaling pathways that are characteristic of each type of cancer.
USP43 affects the proliferation and invasion of breast cancer
The involvement of USP43 in breast cancer has been documented in two previous studies suggesting that Cav2.2 upregulates USP43 to promote tumorigenesis in breast cancer [36]. Cav2.2 is an essential component of the voltage-dependent calcium channel family, which is responsible for regulating Ca2+ levels within physiological thresholds [37]. In support of this, the FDA has approved Ziconotide as a specific inhibitor of Cav2.2 for the treatment of chronic pain [38–41]. While Cav2.2 expression is primarily restricted to brain tissue and absent in epithelial tissue, it has also been shown to be downregulated in normal breast epithelial cells and tissues, as opposed to an upregulation observed in breast cancer tissues and cell lines, particularly in triple-negative breast cancer (TNBC).
The extracellular matrix (ECM) plays a pivotal role in impeding the dissemination and metastasis of primary cancers, while certain cancer cells possess the capability to invade adjacent tissues through ECM degradation [42]. A specialized membrane structure with proteolytic activity is referred to as an invadopodium by which the invasive cells accomplish the invasion and metastasis. For instance, breast cancer cells utilize invadopodia to invade neighboring tissues and degrade the ECM [43]. The actin-binding protein cortactin plays a crucial role in the development of invadopodia [44]. Cortical actin also facilitates the synthesis and secretion of matrix metalloproteinases, which regulate ECM degradation [45, 46]. Similarly, Cav2.2 enhances the production of invadopodia and the degradation of ECM, playing a crucial role in the dissemination and invasion of breast cancer. Further investigation into the underlying mechanisms of this procedure revealed that Cav2.2 regulates USP43 synthesis through NFAT2 dephosphorylation, thereby facilitating the promotion of cortactin and subsequent growth of invadopodia, ultimately leading to metastasis in breast cancer. Addressing F-actin and actin regulatory proteins, which constitute the majority of invadopodia, poses a formidable challenge due to their intricate structural characteristics, rendering them an arduous therapeutic target. According to the aforementioned findings, USP43 exerts regulatory control over invadopodia development, thereby emerging as a promising therapeutic target for impeding invadopodia formation and attenuating the dissemination of breast cancer.
A subsequent investigation revealed a contrasting finding, suggesting that USP43 may function as a suppressive gene for breast cancer. The study proposed an imbalance in the inhibitory relationship between USP43 and EGFR/PI3K/AKT as a potential cause of cancer growth [47]. Numerous investigations have unequivocally established the pivotal role of the nucleosome remodeling and deacetylation (NuRD) complex in breast cancer development and metastasis [48–54]. Multiple subunits of the NuRD complex have been identified, and their expression varies depending on the type of cell and tissue. Histone H2BK120 can be deubiquitinated by the complex of USP43 and NuRD complex and be reactivated. It has been discovered that the USP43/NuRD complex inhibits the expression of numerous cancer-related genes, including the epidermal growth factor receptor (EGFR). EGFR mutations or amplifications are frequently associated with the progression of cancer and an unfavorable prognosis [55–59]. The PI3K-AKT pathway, strongly associated with cancer growth and prognosis, represents one of the signaling cascades that undergo hyperactivation upon EGFR activation [60–62]. The USP43/NuRD complex inhibits the activity of EGFR and AKT, while the EGFR/PI3K/AKT signaling pathway regulates the behavior of USP43. Phosphorylation by AKT enables USP43 to interact with the 14-3-3β/ε heterodimer and remain localized in the cytoplasm. The response inhibits the transcriptional function of USP43, while concurrently attenuating its negative regulation on the EGFR/PI3K/AKT signaling pathway. In line with this argument, a mutually inhibitory loop exists, the imbalance of which promotes breast cancer. These findings suggest that USP43 may serve as an essential component in the regulatory network of the EGFR/PI3K/AKT signaling pathway [47]. Therefore, it has been established that the USP43/NuRD complex inhibits the growth, invasion, and metastasis of breast cancer cells; thus, USP43 may serve as a suppressor of breast cancer.
USP43 affects the proliferation and invasion of colorectal cancer
As a member of the zinc finger E-box (ZEB) family of transcription factors, ZEB1 plays a crucial role in regulating cell differentiation [63], especially in controlling epithelial-mesenchymal transformation (EMT), which is vital for cancer progression. Aberrant expression of ZEB1 has been detected in a wide range of cancer types, such as cervical cancer, pancreatic cancer, osteosarcoma, lung cancer, liver cancer, stomach cancer, colorectal cancer, and breast cancer [64]. According to this study [65], The expression of USP43 is significantly upregulated in colorectal cancer and correlates with an unfavorable prognosis. During the investigation into the role of USP43 in the pathogenesis of colorectal cancer, it has been revealed that USP43 exhibits a capacity to enhance in vitro proliferation, migration, and invasion of colorectal cancer cells. Further investigation revealed a direct interaction between USP43 and ZEB1 in colorectal cell lines, wherein USP43 downregulates ZEB1 protein expression independent of its transcriptional regulation, thereby modulating the abundance of ZEB1 protein. Consequently, USP43 exerts an influence on the process of ubiquitination, thereby affecting the degradation of ZEB1 protein. Moreover, modulation of USP43 expression or knockdown in combination with ZEB1 knockdown or overexpression can exert an impact on various EMT-related biomarkers including E-cadherin, N-cadherin, and vimentin, as well as cell invasion and migration. Therefore, it demonstrates that USP43 and ZEB1 govern the regulation of EMT, which plays a pivotal role in initiating and advancing colorectal cancer.
ZEB1 exerts a significant impact on not only the formation and progression of malignancies but also the development of resistance to chemotherapy. For instance, the knockdown of ZEB1 reduces both the invasiveness of glioblastoma cells and their susceptibility to the chemotherapeutic agent temozolomide [66]. The expression of ZEB1 is strongly associated with the resistance of cancer cells to chemotherapeutic agents including gemcitabine, 5-fluorouracil, and cisplatin in pancreatic cancer cells. Furthermore, the impact of USP43 on the susceptibility of colorectal cancer cells to chemotherapy, reveals a potential role for USP43 in upregulating ZEB1 expression and downregulating chemotherapy resistance in colorectal cancer.
Therefore, USP43 emerges as a promising therapeutic target for the management of colorectal cancer, implying that the aberrant functioning of USP43 may contribute to the progression of this malignancy.
USP43 affects the proliferation and prognosis of pancreatic ductal adenocarcinoma (PDAC)
The majority of pancreatic cancer cases, approximately 90%, are attributed to PDAC, which unfortunately has a low 5-year survival rate of only 8% to 10% [67, 68]. Moreover, a significant proportion of patients with PDAC who are diagnosed with Stage IV generally pass away within a year. As a result of the considerable biological diversity and aggressiveness demonstrated by PDAC, in addition to the broad spectrum of patient prognoses and therapeutic responses, the current treatment options are quite limited in scope [69]. Immunotherapy has recently made significant advancements in the treatment of various malignancies, such as melanoma and breast cancer [70–73], which have been demonstrated to significantly extend cancer survival. Nevertheless, the potential of PDAC for immunotherapy is constrained by three key factors. Firstly, the unique tumor immune microenvironment (TIME) associated with PDAC poses a physical barrier that hampers immune system functionality, impeding the identification and elimination of PDAC by immune cells [74, 75]. Secondly, the majority of immune cells that have infiltrated the peritumoral microenvironment in PDAC are immunosuppressive cells [76, 77]. Thirdly, the TIME of PDAC may be impacted by the intricate network between cytokines and cytokine receptors [78], thereby promoting cancer progression. Consequently, PDAC is considered an “immune desert” or an immunologically “cold tumor”.
A recent study reports that USP43 is significantly upregulated in PDAC, suggesting its potential role in influencing tumor formation by promoting the proliferation of the expression of USP43 in PDAC [79]. Additionally, a positive correlation has been observed between elevated levels of USP43 expression and an unfavorable prognosis in patients with PDAC. These findings suggest that USP43 may serve as a pivotal regulator of PDAC proliferation and hold potential as an independent prognostic indicator. Further analysis of USP43 expression in PDAC revealed a negative association with the chemokine signaling pathway and cytokine-cytokine receptor interactions. The complex interaction of cytokines and receptors can have a substantial influence on PDAC TIME, potentially resulting in immune evasion and faster malignant development. As a result, identifying targets that modify TIME in PDAC, moving it from a “cold” to a “hot” state, has the potential to improve PDAC’s responsiveness to immune checkpoint inhibitors. An additional study of the TIME surrounding PDAC has shown an inverse relationship between USP43 expression and CD8+ T cell activation, indicating that USP43-mediated suppression of CD8+ T cell infiltration leads to a poor prognosis [80–83].
USP43 affects the proliferation and invasion of lung cancer
Lung cancer, which is the most common cancer type among other organs, has a poor prognosis, and scientists are interested in finding new modes of treatment, one of which is ubiquitination. A recent study established a correlation between USP43 and lung squamous cell carcinoma (LUSC), indicating that USP43 facilitated LUSC proliferation and invasion, indicating an unfavorable prognosis [84]. In patients with LUSC, increased USP43 expression is substantially correlated with tumor invasiveness, including larger tumors and advanced TNM stages, and with a reduced overall survival rate. Notably, variability in the expression of USP43 within LUSC tissues is observed among patients. The results of their study provide support for the claim that USP43 has the potential to function as an innovative and autonomous prognostic indicator that can forecast survival outcomes. The in vitro investigations have shown that USP43 stimulates the growth and spread of LUSC, which is consistent with its reported influence on breast and colorectal cancers [65, 85]. The expressions of cyclin-dependent kinase 1 (CDK1), vimentin, and snail are shown to be downregulated after USP43 knockdown, giving more evidence for USP43’s carcinogenic function in LUSC. As a result, the downstream ramifications of USP43 were studied; nevertheless, the specific underlying mechanism remains unknown to the researchers, emphasizing the need for more study. The data presented above suggest that USP43 is a unique prognostic biomarker for patients with LUSC. Furthermore, its deletion has a significant anti-cancer impact by efficiently reducing the proliferation and invasion of LUSC [84].
A further investigation [86] was undertaken to construct a radiation prediction model for early-stage or locally progressed non-small cell lung cancer (NSCLC) using six genes, including APOBEC3B, GOLM1, FAM117A, KCNQ1OT1, PCDHB2, and USP43. The high-risk group for those genes had considerably lower overall survival (OS) and progression-free survival (PFS) than the low-risk group; however, this study did not include a comprehensive examination of individual genes. In conclusion, USP43 is a predictive gene in NSCLC patients, particularly LUSC patients, and further research with large cohorts is needed to explain its specific involvement.
USP43 affects the proliferation and invasion of osteosarcoma
In investigating the role of USP43 in osteosarcoma [87], it was observed that patients with osteosarcoma displayed high levels of USP43, which contributes to the regulation of EMT, a critical cellular mechanism implicated in the progression of osteosarcoma. Furthermore, the upregulation of EMT transcription factors such as Snail, ZEB, or Twist promotes the production of EMT. Notably, elevated expression levels of ZEB1 were observed in osteosarcoma tissue compared to healthy bone tissue. In patients with osteosarcoma and lung metastasis, USP43 deubiquitinates ZEB1 and maintains its transcription [65], resulting in increased invasion of osteosarcoma cells via inducing EMT [88, 89].
USP43 promotes glycolysis and metastasis in bladder cancer (BLCA)
The primary factors contributing to the unfavorable prognosis of BLCA are invasion and recurrence [90]. Under aerobic conditions, cancer cells obtain energy via glycolytic metabolism rather than oxidative phosphorylation; this process is known as the Warburg effect [91]. Warburg effect is closely related to the pathogenesis and aggressiveness of BLCA [92]. c-Myc plays a crucial role in regulating aerobic glycolysis [93]. It has been observed that BLCA induces amplification of the MYC oncogene, and its products may contribute to the tumorigenesis of BLCA [94–96]. Nevertheless, c-Myc is prone to ubiquitase degradation, thereby impeding tumor progression [97–100].
A recent study [101] demonstrates that USP43 is substantially upregulated in BLCA and that its expression increases with tumor grade. Moreover, USP43 may stimulate the metastasis of BLCA. The examination of differentially expressed genes revealed a positive correlation between USP43 and both the glycolysis pathway and the MYC target pathway. As a result, the researchers conducted additional investigations into the correlation between USP43 and c-Myc and discovered that USP43 deubiquitinated c-Myc at K148 and K289, thereby stabilizing its expression. c-Myc oncoprotein promotes the transcription of USP43 and regulates the transcription of at least 15% of the entire genome as a transcription factor. Consequently, USP43 and c-Myc form a feedback loop in which their activity is reciprocal; an asymmetry in this loop results in atypical glycolysis and c-Myc accumulation, both of which initiate the malignant behavior of BLCA. Furthermore, therapeutic targeting of c-Myc is challenging due to its localization and reaction affinity within the nucleus. In summary, USP43 emerges as a prospective therapeutic target for BLCA.
USP43 promotes the proliferation of epithelial ovarian cancer (EOC) and impairs its cisplatin sensitivity
Epithelial ovarian cancer (EOC) is the predominant form, representing over 90% of all ovarian cancer cases, with the majority being diagnosed in advanced stages [102–104]. Primary treatments for EOC include debulking surgery and a combination of cisplatin and taxane chemotherapy [105]. Cisplatin administration frequently leads to drug resistance, impacting the effectiveness of chemotherapy, so it is crucial to identify new effective treatment targets [106, 107].
Previous research discovered that USP43 acts as a promoter in various types of cancer, including EOC. According to a study [108], the cancer’s malignant characteristics are more pronounced in patients with high levels of USP43 in EOC. High levels of USP43 in EOC are linked to a poor prognosis. The study delves into the tumor-promoting impact in EOC, showing that USP43 enhances the proliferation, invasion, and migration of EOC, and facilitates EOC cells to enter the cell cycle’s proliferation phase.
Cisplatin is a primary chemotherapeutic treatment for EOC. Researchers investigate the impact of USP43 on the efficacy of cisplatin in EOC cells that are resistant to the drug. USP43 suppressed DNA damage and apoptosis, thereby reducing the sensitivity of EOC cells to cisplatin. Genes associated with USP43 in EOC primarily focus on controlling cancer advancement and histone deacetylation (HDAC). HDAC2, a member of the HDAC family, is shown to be significantly upregulated in EOC, and its high expression is linked to a negative prognosis. The same study also explores the correlation between USP43 and HDAC2. USP43 is discovered to remove ubiquitin from HDAC2, leading to the stabilization of the HDAC2 protein. USP43 hindered the responsiveness of EOC cells to cisplatin by targeting HDAC2. Upon further investigation, it was discovered that HDAC2 triggers the Wnt/β-catenin pathway, leading to decreased sensitivity of EOC cells to cisplatin.
Overall, USP43 affects the susceptibility of EOC cells to cisplatin and is identified as a therapeutic target to overcome cisplatin resistance.
Conclusion and future perspectives
TP53 is the most important cancer-related gene. This tumor suppressor gene, found on chromosome 17p13.1, is linked to both hereditary and sporadic malignancies. Mutations in the TP53 gene, which affects 17p13.1 of the human genome, are among humans’ most common genetic alterations and are thought to represent hereditary malignant transformation [109–111]. Interestingly, the presence of the USP43 gene in this chromosomal region implies that mutations in 17p13.1, particularly deletions, may have a negative impact on USP43 functioning. Notably, online database studies revealed frequent alterations and/or lack of USP43 in numerous types of cancer, thereby compelling further investigation into its function in the progression of cancer and potential targets as a novel therapeutic approach.
Studies in databases and clinicopathological specimens have confirmed that USP43 expression is significantly elevated in breast, pancreatic, lung, bladder, and epithelial ovarian cancers. Furthermore, the USP43 level is associated with poor prognosis and its involvement in different cancer pathways. USP43 potentially regulates cancer invasion by modulating EMT-associated transcription factors, such as ZEB1 or SNAIL, in breast cancer, colorectal cancer, and osteosarcoma. In breast cancer, USP43 promotes invasion and metastasis through invadopodia formation. In BLCA, USP43 promotes glycolysis and metastasis. In EOC, USP43 promotes the proliferation and impairs cisplatin sensitivity. Moreover, it may govern the duration of PDAC progression while facilitating CD8+ T cell infiltration. Additionally, USP43 can be employed in conjunction with other genes to predict lung cancer outcomes. Therefore, USP43 not only affects the proliferation and metastasis of tumors directly but also influences the immune microenvironment of some kinds of cancers, which affects their biological behavior (Figure 2 and Table 1).
Table 1. The prognostic significance of ubiquitin-specific protease 43 (USP43) across diverse cancer entities.
| Cancer | Pathway | Year |
| Breast cancer | Cav2.2-NFAT2-USP43 axis facilitates invadopodia formation and metastasis [36]. | 2022 |
| Imbalance of the reciprocally inhibitory loop between the USP43 and EGFR/PI3K/AKT drives breast carcinogenesis [47]. | 2018 | |
| Pancreas | Modulating the proliferation and infiltration of surrounding immune cells [79]. | 2023 |
| Lung cancer | Promoting growth and invasion of LUSC may relate to CDK1, vimentin, and snail [84]. | 2022 |
| Predicting the risk of NSCLC with the other five genes [86]. | 2022 | |
| Colorectal cancer | Regulating ZEB1 protein and mediating proliferation and metastasis [65]. | 2021 |
| Osteosarcoma | Regulating EMT and progression [87]. | 2021 |
| Bladder cancer | USP43 stabilizes c-Myc, and c-Myc promotes the translation of USP43. This loop imbalance between USP43 and c-Myc drives glycolysis and bladder carcinogenesis [101]. | 2024 |
| Epithelial ovarian cancer | The stabilization of HDAC2 by USP43 and the modulation of the Wnt/β catenin signaling pathway result in a reduction in cisplatin sensitivity [108]. | 2024 |
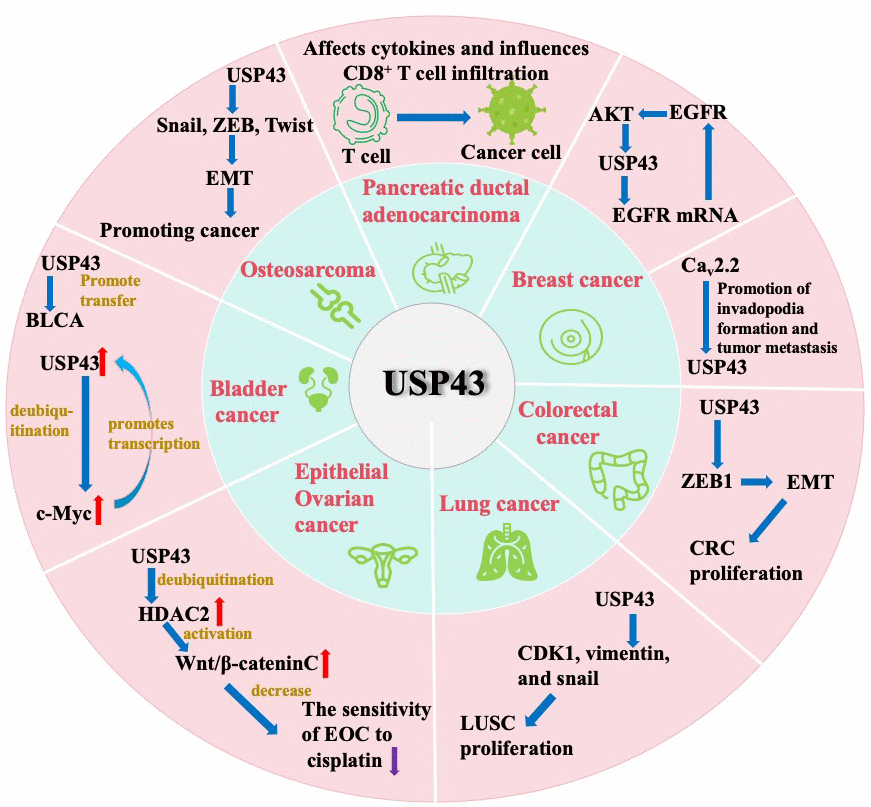
Figure 2. The potential significance of USP43 across diverse cancer entities.
In conclusion, our findings demonstrate a high frequency of USP43 mutations across various human cancers, implicating its pivotal role in tumorigenesis and prognosis prediction for chemo- and radiotherapy efficacy. USP43 may be a potential indicator for predicting the prognosis of cancer and may also be a conducive target for monitoring cancer therapy in the future. These results highlight the potential of targeting USP43 as a promising therapeutic strategy for future cancer treatment.
Author Contributions
ZZ and ML designed the study and wrote the main manuscript. ZL, MZ, RF, and AA explored most of the research and analyzed the data. ZZ, LL, and XZ revised the manuscript. GT and HH supervised the study. All authors reviewed and approved the final manuscript.
Conflicts of Interest
There are no conflicts of interest in this review.
Funding
This research was supported by the 2023 Liaoning Science and Technology Joint Plan (2023-BSBA-100).
References
- 1. Millar AH, Heazlewood JL, Giglione C, Holdsworth MJ, Bachmair A, Schulze WX. The Scope, Functions, and Dynamics of Posttranslational Protein Modifications. Annu Rev Plant Biol. 2019; 70:119–51. https://doi.org/10.1146/annurev-arplant-050718-100211 [PubMed]
- 2. Tang W, Wan S, Yang Z, Teschendorff AE, Zou Q. Tumor origin detection with tissue-specific miRNA and DNA methylation markers. Bioinformatics. 2018; 34:398–406. https://doi.org/10.1093/bioinformatics/btx622 [PubMed]
- 3. Liu H, Wang D, Zhang Q, Zhao Y, Mamonova T, Wang L, Zhang C, Li S, Friedman PA, Xiao K. Parallel Post-Translational Modification Scanning Enhancing Hydrogen-Deuterium Exchange-Mass Spectrometry Coverage of Key Structural Regions. Anal Chem. 2019; 91:6976–80. https://doi.org/10.1021/acs.analchem.9b01410 [PubMed]
- 4. Wang Y, Wang F. Post-Translational Modifications of Deubiquitinating Enzymes: Expanding the Ubiquitin Code. Front Pharmacol. 2021; 12:685011. https://doi.org/10.3389/fphar.2021.685011 [PubMed]
- 5. Davis NM, Sokolosky M, Stadelman K, Abrams SL, Libra M, Candido S, Nicoletti F, Polesel J, Maestro R, D’Assoro A, Drobot L, Rakus D, Gizak A, et al. Deregulation of the EGFR/PI3K/PTEN/Akt/mTORC1 pathway in breast cancer: possibilities for therapeutic intervention. Oncotarget. 2014; 5:4603–50. https://doi.org/10.18632/oncotarget.2209 [PubMed]
- 6. Lander GC, Estrin E, Matyskiela ME, Bashore C, Nogales E, Martin A. Complete subunit architecture of the proteasome regulatory particle. Nature. 2012; 482:186–91. https://doi.org/10.1038/nature10774 [PubMed]
- 7. Fuchs G, Shema E, Vesterman R, Kotler E, Wolchinsky Z, Wilder S, Golomb L, Pribluda A, Zhang F, Haj-Yahya M, Feldmesser E, Brik A, Yu X, et al. RNF20 and USP44 regulate stem cell differentiation by modulating H2B monoubiquitylation. Mol Cell. 2012; 46:662–73. https://doi.org/10.1016/j.molcel.2012.05.023 [PubMed]
- 8. Han ZJ, Feng YH, Gu BH, Li YM, Chen H. The post-translational modification, SUMOylation, and cancer (Review). Int J Oncol. 2018; 52:1081–94. https://doi.org/10.3892/ijo.2018.4280 [PubMed]
- 9. Grabbe C, Husnjak K, Dikic I. The spatial and temporal organization of ubiquitin networks. Nat Rev Mol Cell Biol. 2011; 12:295–307. https://doi.org/10.1038/nrm3099 [PubMed]
- 10. Liu H, Zhang Q, Li K, Gong Z, Liu Z, Xu Y, Swaney MH, Xiao K, Chen Y. Prognostic significance of USP33 in advanced colorectal cancer patients: new insights into β-arrestin-dependent ERK signaling. Oncotarget. 2016; 7:81223–40. https://doi.org/10.18632/oncotarget.13219 [PubMed]
- 11. Mennerich D, Kubaichuk K, Kietzmann T. DUBs, Hypoxia, and Cancer. Trends Cancer. 2019; 5:632–53. https://doi.org/10.1016/j.trecan.2019.08.005 [PubMed]
- 12. Young MJ, Hsu KC, Lin TE, Chang WC, Hung JJ. The role of ubiquitin-specific peptidases in cancer progression. J Biomed Sci. 2019; 26:42. https://doi.org/10.1186/s12929-019-0522-0 [PubMed]
- 13. Ciechanover A, Schwartz AL. The ubiquitin system: pathogenesis of human diseases and drug targeting. Biochim Biophys Acta. 2004; 1695:3–17. https://doi.org/10.1016/j.bbamcr.2004.09.018 [PubMed]
- 14. Petroski MD. The ubiquitin system, disease, and drug discovery. BMC Biochem. 2008 (Suppl 1); 9:S7. https://doi.org/10.1186/1471-2091-9-S1-S7 [PubMed]
- 15. Kirkin V, Dikic I. Ubiquitin networks in cancer. Curr Opin Genet Dev. 2011; 21:21–8. https://doi.org/10.1016/j.gde.2010.10.004 [PubMed]
- 16. Pfoh R, Lacdao IK, Saridakis V. Deubiquitinases and the new therapeutic opportunities offered to cancer. Endocr Relat Cancer. 2015; 22:T35–54. https://doi.org/10.1530/ERC-14-0516 [PubMed]
- 17. Amerik AY, Hochstrasser M. Mechanism and function of deubiquitinating enzymes. Biochim Biophys Acta. 2004; 1695:189–207. https://doi.org/10.1016/j.bbamcr.2004.10.003 [PubMed]
- 18. Tyagi N, Tyagi M, Pachauri M, Ghosh PC. Potential therapeutic applications of plant toxin-ricin in cancer: challenges and advances. Tumour Biol. 2015; 36:8239–46. https://doi.org/10.1007/s13277-015-4028-4 [PubMed]
- 19. Komander D. The emerging complexity of protein ubiquitination. Biochem Soc Trans. 2009; 37:937–53. https://doi.org/10.1042/BST0370937 [PubMed]
- 20. Avvakumov GV, Walker JR, Xue S, Finerty PJ
Jr , Mackenzie F, Newman EM, Dhe-Paganon S. Amino-terminal dimerization, NRDP1-rhodanese interaction, and inhibited catalytic domain conformation of the ubiquitin-specific protease 8 (USP8). J Biol Chem. 2006; 281:38061–70. https://doi.org/10.1074/jbc.M606704200 [PubMed] - 21. Nijman SM, Luna-Vargas MP, Velds A, Brummelkamp TR, Dirac AM, Sixma TK, Bernards R. A genomic and functional inventory of deubiquitinating enzymes. Cell. 2005; 123:773–86. https://doi.org/10.1016/j.cell.2005.11.007 [PubMed]
- 22. Reyes-Turcu FE, Ventii KH, Wilkinson KD. Regulation and cellular roles of ubiquitin-specific deubiquitinating enzymes. Annu Rev Biochem. 2009; 78:363–97. https://doi.org/10.1146/annurev.biochem.78.082307.091526 [PubMed]
- 23. Pfoh R, Lacdao IK, Georges AA, Capar A, Zheng H, Frappier L, Saridakis V. Crystal Structure of USP7 Ubiquitin-like Domains with an ICP0 Peptide Reveals a Novel Mechanism Used by Viral and Cellular Proteins to Target USP7. PLoS Pathog. 2015; 11:e1004950. https://doi.org/10.1371/journal.ppat.1004950 [PubMed]
- 24. Pal A, Young MA, Donato NJ. Emerging potential of therapeutic targeting of ubiquitin-specific proteases in the treatment of cancer. Cancer Res. 2014; 74:4955–66. https://doi.org/10.1158/0008-5472.CAN-14-1211 [PubMed]
- 25. Tavana O, Li D, Dai C, Lopez G, Banerjee D, Kon N, Chen C, Califano A, Yamashiro DJ, Sun H, Gu W. HAUSP deubiquitinates and stabilizes N-Myc in neuroblastoma. Nat Med. 2016; 22:1180–6. https://doi.org/10.1038/nm.4180 [PubMed]
- 26. Yuasa-Kawada J, Kinoshita-Kawada M, Rao Y, Wu JY. Deubiquitinating enzyme USP33/VDU1 is required for Slit signaling in inhibiting breast cancer cell migration. Proc Natl Acad Sci USA. 2009; 106:14530–5. https://doi.org/10.1073/pnas.0801262106 [PubMed]
- 27. Fraile JM, Quesada V, Rodríguez D, Freije JM, López-Otín C. Deubiquitinases in cancer: new functions and therapeutic options. Oncogene. 2012; 31:2373–88. https://doi.org/10.1038/onc.2011.443 [PubMed]
- 28. Wang Q, Ma S, Song N, Li X, Liu L, Yang S, Ding X, Shan L, Zhou X, Su D, Wang Y, Zhang Q, Liu X, et al. Stabilization of histone demethylase PHF8 by USP7 promotes breast carcinogenesis. J Clin Invest. 2016; 126:2205–20. https://doi.org/10.1172/JCI85747 [PubMed]
- 29. Li X, Song N, Liu L, Liu X, Ding X, Song X, Yang S, Shan L, Zhou X, Su D, Wang Y, Zhang Q, Cao C, et al. USP9X regulates centrosome duplication and promotes breast carcinogenesis. Nat Commun. 2017; 8:14866. https://doi.org/10.1038/ncomms14866 [PubMed]
- 30. Engel K, Rudelius M, Slawska J, Jacobs L, Ahangarian Abhari B, Altmann B, Kurutz J, Rathakrishnan A, Fernández-Sáiz V, Brunner A, Targosz BS, Loewecke F, Gloeckner CJ, et al. USP9X stabilizes XIAP to regulate mitotic cell death and chemoresistance in aggressive B-cell lymphoma. EMBO Mol Med. 2016; 8:851–62. https://doi.org/10.15252/emmm.201506047 [PubMed]
- 31. Li Z, Na X, Wang D, Schoen SR, Messing EM, Wu G. Ubiquitination of a novel deubiquitinating enzyme requires direct binding to von Hippel-Lindau tumor suppressor protein. J Biol Chem. 2002; 277:4656–62. https://doi.org/10.1074/jbc.M108269200 [PubMed]
- 32. Li Z, Wang D, Na X, Schoen SR, Messing EM, Wu G. Identification of a deubiquitinating enzyme subfamily as substrates of the von Hippel-Lindau tumor suppressor. Biochem Biophys Res Commun. 2002; 294:700–9. https://doi.org/10.1016/S0006-291X(02)00534-X [PubMed]
- 33. Allen MD, Bycroft M. The solution structure of the ZnF UBP domain of USP33/VDU1. Protein Sci. 2007; 16:2072–5. https://doi.org/10.1110/ps.072967807 [PubMed]
- 34. Singh N, Singh AB. Deubiquitinases and cancer: A snapshot. Crit Rev Oncol Hematol. 2016; 103:22–6. https://doi.org/10.1016/j.critrevonc.2016.04.018 [PubMed]
- 35. Wang SA, Wang YC, Chuang YP, Huang YH, Su WC, Chang WC, Hung JJ. EGF-mediated inhibition of ubiquitin-specific peptidase 24 expression has a crucial role in tumorigenesis. Oncogene. 2017; 36:2930–45. https://doi.org/10.1038/onc.2016.445 [PubMed]
- 36. Xue Y, Li M, Hu J, Song Y, Guo W, Miao C, Ge D, Hou Y, Wang X, Huang X, Liu T, Zhang X, Huang Q. Cav2.2-NFAT2-USP43 axis promotes invadopodia formation and breast cancer metastasis through cortactin stabilization. Cell Death Dis. 2022; 13:812. https://doi.org/10.1038/s41419-022-05174-0 [PubMed]
- 37. Nanou E, Catterall WA. Calcium Channels, Synaptic Plasticity, and Neuropsychiatric Disease. Neuron. 2018; 98:466–81. https://doi.org/10.1016/j.neuron.2018.03.017 [PubMed]
- 38. Heinke B, Balzer E, Sandkühler J. Pre- and postsynaptic contributions of voltage-dependent Ca2+ channels to nociceptive transmission in rat spinal lamina I neurons. Eur J Neurosci. 2004; 19:103–11. https://doi.org/10.1046/j.1460-9568.2003.03083.x [PubMed]
- 39. Matthews EA, Dickenson AH. Effects of spinally delivered N- and P-type voltage-dependent calcium channel antagonists on dorsal horn neuronal responses in a rat model of neuropathy. Pain. 2001; 92:235–46. https://doi.org/10.1016/s0304-3959(01)00255-x [PubMed]
- 40. Winquist RJ, Pan JQ, Gribkoff VK. Use-dependent blockade of Cav2.2 voltage-gated calcium channels for neuropathic pain. Biochem Pharmacol. 2005; 70:489–99. https://doi.org/10.1016/j.bcp.2005.04.035 [PubMed]
- 41. Wall MJ, Subasinghe NL, Winters MP, Lubin ML, Finley MFA, Qin N, Brandt MR, Neeper MP, Schneider CR, Colburn RW, Flores CM, Sui Z. Discovery and optimization of a novel series of pyrazolyltetrahydropyran N-type calcium channel (Cav 2.2) blockers for the treatment of pain. Bioorg Med Chem Lett. 2018; 28:3780–3. https://doi.org/10.1016/j.bmcl.2018.10.007 [PubMed]
- 42. Chiang AC, Massagué J. Molecular basis of metastasis. N Engl J Med. 2008; 359:2814–23. https://doi.org/10.1056/NEJMra0805239 [PubMed]
- 43. Meirson T, Gil-Henn H. Targeting invadopodia for blocking breast cancer metastasis. Drug Resist Updat. 2018; 39:1–17. https://doi.org/10.1016/j.drup.2018.05.002 [PubMed]
- 44. Hill A, McFarlane S, Mulligan K, Gillespie H, Draffin JE, Trimble A, Ouhtit A, Johnston PG, Harkin DP, McCormick D, Waugh DJ. Cortactin underpins CD44-promoted invasion and adhesion of breast cancer cells to bone marrow endothelial cells. Oncogene. 2006; 25:6079–91. https://doi.org/10.1038/sj.onc.1209628 [PubMed]
- 45. Artym VV, Zhang Y, Seillier-Moiseiwitsch F, Yamada KM, Mueller SC. Dynamic interactions of cortactin and membrane type 1 matrix metalloproteinase at invadopodia: defining the stages of invadopodia formation and function. Cancer Res. 2006; 66:3034–43. https://doi.org/10.1158/0008-5472.CAN-05-2177 [PubMed]
- 46. Zhao G, Zhang H, Huang Z, Lv L, Yan F. Cortactin and Exo70 mediated invasion of hepatoma carcinoma cells by MMP-9 secretion. Mol Biol Rep. 2016; 43:407–14. https://doi.org/10.1007/s11033-016-3972-4 [PubMed]
- 47. He L, Liu X, Yang J, Li W, Liu S, Liu X, Yang Z, Ren J, Wang Y, Shan L, Guan C, Pei F, Lei L, et al. Imbalance of the reciprocally inhibitory loop between the ubiquitin-specific protease USP43 and EGFR/PI3K/AKT drives breast carcinogenesis. Cell Res. 2018; 28:934–51. https://doi.org/10.1038/s41422-018-0079-6 [PubMed]
- 48. Si W, Huang W, Zheng Y, Yang Y, Liu X, Shan L, Zhou X, Wang Y, Su D, Gao J, Yan R, Han X, Li W, et al. Dysfunction of the Reciprocal Feedback Loop between GATA3- and ZEB2-Nucleated Repression Programs Contributes to Breast Cancer Metastasis. Cancer Cell. 2015; 27:822–36. https://doi.org/10.1016/j.ccell.2015.04.011 [PubMed]
- 49. Wang Y, Zhang H, Chen Y, Sun Y, Yang F, Yu W, Liang J, Sun L, Yang X, Shi L, Li R, Li Y, Zhang Y, et al. LSD1 is a subunit of the NuRD complex and targets the metastasis programs in breast cancer. Cell. 2009; 138:660–72. https://doi.org/10.1016/j.cell.2009.05.050 [PubMed]
- 50. Li Q, Shi L, Gui B, Yu W, Wang J, Zhang D, Han X, Yao Z, Shang Y. Binding of the JmjC demethylase JARID1B to LSD1/NuRD suppresses angiogenesis and metastasis in breast cancer cells by repressing chemokine CCL14. Cancer Res. 2011; 71:6899–908. https://doi.org/10.1158/0008-5472.CAN-11-1523 [PubMed]
- 51. Fujita N, Jaye DL, Kajita M, Geigerman C, Moreno CS, Wade PA. MTA3, a Mi-2/NuRD complex subunit, regulates an invasive growth pathway in breast cancer. Cell. 2003; 113:207–19. https://doi.org/10.1016/s0092-8674(03)00234-4 [PubMed]
- 52. Li DQ, Pakala SB, Nair SS, Eswaran J, Kumar R. Metastasis-associated protein 1/nucleosome remodeling and histone deacetylase complex in cancer. Cancer Res. 2012; 72:387–94. https://doi.org/10.1158/0008-5472.CAN-11-2345 [PubMed]
- 53. Lai AY, Wade PA. Cancer biology and NuRD: a multifaceted chromatin remodelling complex. Nat Rev Cancer. 2011; 11:588–96. https://doi.org/10.1038/nrc3091 [PubMed]
- 54. Shan L, Zhou X, Liu X, Wang Y, Su D, Hou Y, Yu N, Yang C, Liu B, Gao J, Duan Y, Yang J, Li W, et al. FOXK2 Elicits Massive Transcription Repression and Suppresses the Hypoxic Response and Breast Cancer Carcinogenesis. Cancer Cell. 2016; 30:708–22. https://doi.org/10.1016/j.ccell.2016.09.010 [PubMed]
- 55. Appert-Collin A, Hubert P, Crémel G, Bennasroune A. Role of ErbB Receptors in Cancer Cell Migration and Invasion. Front Pharmacol. 2015; 6:283. https://doi.org/10.3389/fphar.2015.00283 [PubMed]
- 56. Chan SK, Hill ME, Gullick WJ. The role of the epidermal growth factor receptor in breast cancer. J Mammary Gland Biol Neoplasia. 2006; 11:3–11. https://doi.org/10.1007/s10911-006-9008-2 [PubMed]
- 57. Holbro T, Civenni G, Hynes NE. The ErbB receptors and their role in cancer progression. Exp Cell Res. 2003; 284:99–110. https://doi.org/10.1016/s0014-4827(02)00099-x [PubMed]
- 58. Dheeraj A, Rigby CM, O’Bryant CL, Agarwal C, Singh RP, Deep G, Agarwal R. Silibinin Treatment Inhibits the Growth of Hedgehog Inhibitor-Resistant Basal Cell Carcinoma Cells via Targeting EGFR-MAPK-Akt and Hedgehog Signaling. Photochem Photobiol. 2017; 93:999–1007. https://doi.org/10.1111/php.12727 [PubMed]
- 59. Vergara D, Valente CM, Tinelli A, Siciliano C, Lorusso V, Acierno R, Giovinazzo G, Santino A, Storelli C, Maffia M. Resveratrol inhibits the epidermal growth factor-induced epithelial mesenchymal transition in MCF-7 cells. Cancer Lett. 2011; 310:1–8. https://doi.org/10.1016/j.canlet.2011.04.009 [PubMed]
- 60. Alanazi IO, Khan Z. Understanding EGFR Signaling in Breast Cancer and Breast Cancer Stem Cells: Overexpression and Therapeutic Implications. Asian Pac J Cancer Prev. 2016; 17:445–53. https://doi.org/10.7314/apjcp.2016.17.2.445 [PubMed]
- 61. Engelman JA, Luo J, Cantley LC. The evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nat Rev Genet. 2006; 7:606–19. https://doi.org/10.1038/nrg1879 [PubMed]
- 62. Engelman JA. Targeting PI3K signalling in cancer: opportunities, challenges and limitations. Nat Rev Cancer. 2009; 9:550–62. https://doi.org/10.1038/nrc2664 [PubMed]
- 63. Zhang P, Sun Y, Ma L. ZEB1: at the crossroads of epithelial-mesenchymal transition, metastasis and therapy resistance. Cell Cycle. 2015; 14:481–7. https://doi.org/10.1080/15384101.2015.1006048 [PubMed]
- 64. Yang Y, Ahn YH, Chen Y, Tan X, Guo L, Gibbons DL, Ungewiss C, Peng DH, Liu X, Lin SH, Thilaganathan N, Wistuba II, Rodriguez-Canales J, et al. ZEB1 sensitizes lung adenocarcinoma to metastasis suppression by PI3K antagonism. J Clin Invest. 2014; 124:2696–708. https://doi.org/10.1172/JCI72171 [PubMed]
- 65. Ye DX, Wang SS, Huang Y, Wang XJ, Chi P. USP43 directly regulates ZEB1 protein, mediating proliferation and metastasis of colorectal cancer. J Cancer. 2021; 12:404–16. https://doi.org/10.7150/jca.48056 [PubMed]
- 66. Siebzehnrubl FA, Silver DJ, Tugertimur B, Deleyrolle LP, Siebzehnrubl D, Sarkisian MR, Devers KG, Yachnis AT, Kupper MD, Neal D, Nabilsi NH, Kladde MP, Suslov O, et al. The ZEB1 pathway links glioblastoma initiation, invasion and chemoresistance. EMBO Mol Med. 2013; 5:1196–212. https://doi.org/10.1002/emmm.201302827 [PubMed]
- 67. Fesinmeyer MD, Austin MA, Li CI, De Roos AJ, Bowen DJ. Differences in survival by histologic type of pancreatic cancer. Cancer Epidemiol Biomarkers Prev. 2005; 14:1766–73. https://doi.org/10.1158/1055-9965.EPI-05-0120 [PubMed]
- 68. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2020. CA Cancer J Clin. 2020; 70:7–30. https://doi.org/10.3322/caac.21590 [PubMed]
- 69. Chiaravalli M, Reni M, O’Reilly EM. Pancreatic ductal adenocarcinoma: State-of-the-art 2017 and new therapeutic strategies. Cancer Treat Rev. 2017; 60:32–43. https://doi.org/10.1016/j.ctrv.2017.08.007 [PubMed]
- 70. Deng Y, Xia X, Zhao Y, Zhao Z, Martinez C, Yin W, Yao J, Hang Q, Wu W, Zhang J, Yu Y, Xia W, Yao F, et al. Glucocorticoid receptor regulates PD-L1 and MHC-I in pancreatic cancer cells to promote immune evasion and immunotherapy resistance. Nat Commun. 2021; 12:7041. https://doi.org/10.1038/s41467-021-27349-7 [PubMed]
- 71. Hodi FS, O’Day SJ, McDermott DF, Weber RW, Sosman JA, Haanen JB, Gonzalez R, Robert C, Schadendorf D, Hassel JC, Akerley W, van den Eertwegh AJ, Lutzky J, et al. Improved survival with ipilimumab in patients with metastatic melanoma. N Engl J Med. 2010; 363:711–23. https://doi.org/10.1056/NEJMoa1003466 [PubMed]
- 72. Topalian SL, Hodi FS, Brahmer JR, Gettinger SN, Smith DC, McDermott DF, Powderly JD, Carvajal RD, Sosman JA, Atkins MB, Leming PD, Spigel DR, Antonia SJ, et al. Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N Engl J Med. 2012; 366:2443–54. https://doi.org/10.1056/NEJMoa1200690 [PubMed]
- 73. Brahmer JR, Tykodi SS, Chow LQ, Hwu WJ, Topalian SL, Hwu P, Drake CG, Camacho LH, Kauh J, Odunsi K, Pitot HC, Hamid O, Bhatia S, et al. Safety and activity of anti-PD-L1 antibody in patients with advanced cancer. N Engl J Med. 2012; 366:2455–65. https://doi.org/10.1056/NEJMoa1200694 [PubMed]
- 74. Neesse A, Algül H, Tuveson DA, Gress TM. Stromal biology and therapy in pancreatic cancer: a changing paradigm. Gut. 2015; 64:1476–84. https://doi.org/10.1136/gutjnl-2015-309304 [PubMed]
- 75. Murakami T, Hiroshima Y, Matsuyama R, Homma Y, Hoffman RM, Endo I. Role of the tumor microenvironment in pancreatic cancer. Ann Gastroenterol Surg. 2019; 3:130–7. https://doi.org/10.1002/ags3.12225 [PubMed]
- 76. Gentles AJ, Newman AM, Liu CL, Bratman SV, Feng W, Kim D, Nair VS, Xu Y, Khuong A, Hoang CD, Diehn M, West RB, Plevritis SK, Alizadeh AA. The prognostic landscape of genes and infiltrating immune cells across human cancers. Nat Med. 2015; 21:938–45. https://doi.org/10.1038/nm.3909 [PubMed]
- 77. Ali HR, Chlon L, Pharoah PD, Markowetz F, Caldas C. Patterns of Immune Infiltration in Breast Cancer and Their Clinical Implications: A Gene-Expression-Based Retrospective Study. PLoS Med. 2016; 13:e1002194. https://doi.org/10.1371/journal.pmed.1002194 [PubMed]
- 78. Balkwill F. Cancer and the chemokine network. Nat Rev Cancer. 2004; 4:540–50. https://doi.org/10.1038/nrc1388 [PubMed]
- 79. Zhao Z, Lin Z, Guo X, Al-Danakh A, He H, Qin H, Ma C, Zhang N, Tan G. Ubiquitin-Specific Protease 43 Impacts Pancreatic Ductal Adenocarcinoma Prognosis by Altering Its Proliferation and Infiltration of Surrounding Immune Cells. J Immunol Res. 2023; 2023:4311388. https://doi.org/10.1155/2023/4311388 [PubMed]
- 80. Liu R, Liao YZ, Zhang W, Zhou HH. Relevance of Immune Infiltration and Clinical Outcomes in Pancreatic Ductal Adenocarcinoma Subtypes. Front Oncol. 2021; 10:575264. https://doi.org/10.3389/fonc.2020.575264 [PubMed]
- 81. Fukunaga A, Miyamoto M, Cho Y, Murakami S, Kawarada Y, Oshikiri T, Kato K, Kurokawa T, Suzuoki M, Nakakubo Y, Hiraoka K, Itoh T, Morikawa T, et al. CD8+ tumor-infiltrating lymphocytes together with CD4+ tumor-infiltrating lymphocytes and dendritic cells improve the prognosis of patients with pancreatic adenocarcinoma. Pancreas. 2004; 28:e26–31. https://doi.org/10.1097/00006676-200401000-00023 [PubMed]
- 82. Zhang J, Wang YF, Wu B, Zhong ZX, Wang KX, Yang LQ, Wang YQ, Li YQ, Gao J, Li ZS. Intraepithelial Attack Rather than Intratumorally Infiltration of CD8+T Lymphocytes is a Favorable Prognostic Indicator in Pancreatic Ductal Adenocarcinoma. Curr Mol Med. 2017; 17:689–98. https://doi.org/10.2174/1566524018666180308115705 [PubMed]
- 83. Balachandran VP, Łuksza M, Zhao JN, Makarov V, Moral JA, Remark R, Herbst B, Askan G, Bhanot U, Senbabaoglu Y, Wells DK, Cary CIO, Grbovic-Huezo O, et al, Australian Pancreatic Cancer Genome Initiative, and Garvan Institute of Medical Research, and Prince of Wales Hospital, and Royal North Shore Hospital, and University of Glasgow, and St Vincent’s Hospital, and QIMR Berghofer Medical Research Institute, and University of Melbourne, Centre for Cancer Research, and University of Queensland, Institute for Molecular Bioscience, and Bankstown Hospital, and Liverpool Hospital, and Royal Prince Alfred Hospital, Chris O’Brien Lifehouse, and Westmead Hospital, and Fremantle Hospital, and St John of God Healthcare, and Royal Adelaide Hospital, and Flinders Medical Centre, and Envoi Pathology, and Princess Alexandria Hospital, and Austin Hospital, and Johns Hopkins Medical Institutes, and ARC-Net Centre for Applied Research on Cancer. Identification of unique neoantigen qualities in long-term survivors of pancreatic cancer. Nature. 2017; 551:512–6. https://doi.org/10.1038/nature24462 [PubMed]
- 84. Sun Q, Zhang H, Zong L, Julaiti A, Jing X, Zhang L. Prognostic Value and Oncogenic Effects of Ubiquitin-Specific Protease 43 in Lung Squamous Cell Carcinoma. Tohoku J Exp Med. 2022; 257:135–45. https://doi.org/10.1620/tjem.2022.J008 [PubMed]
- 85. Lin F, Xie Z, Chang L, Li W, Wang L, Hou Y, Li L, Zhu J, Xia Y, He W, Li W. USP43 promotes tumorigenesis through regulating cell cycle and EMT in breast cancer. Int J Clin Exp Pathol. 2017; 10:11014–21. [PubMed]
- 86. Peinado-Serrano J, Quintanal-Villalonga Á, Muñoz-Galvan S, Verdugo-Sivianes EM, Mateos JC, Ortiz-Gordillo MJ, Carnero A. A Six-Gene Prognostic and Predictive Radiotherapy-Based Signature for Early and Locally Advanced Stages in Non-Small-Cell Lung Cancer. Cancers (Basel). 2022; 14:2054. https://doi.org/10.3390/cancers14092054 [PubMed]
- 87. Lavaud M, Mullard M, Tesfaye R, Amiaud J, Legrand M, Danieau G, Brion R, Morice S, Regnier L, Dupuy M, Brounais-Le Royer B, Lamoureux F, Ory B, et al. Overexpression of the Ubiquitin Specific Proteases USP43, USP41, USP27x and USP6 in Osteosarcoma Cell Lines: Inhibition of Osteosarcoma Tumor Growth and Lung Metastasis Development by the USP Antagonist PR619. Cells. 2021; 10:2268. https://doi.org/10.3390/cells10092268 [PubMed]
- 88. Yang G, Yuan J, Li K. EMT transcription factors: implication in osteosarcoma. Med Oncol. 2013; 30:697. https://doi.org/10.1007/s12032-013-0697-2 [PubMed]
- 89. Verrecchia F, Rédini F. Transforming Growth Factor-β Signaling Plays a Pivotal Role in the Interplay Between Osteosarcoma Cells and Their Microenvironment. Front Oncol. 2018; 8:133. https://doi.org/10.3389/fonc.2018.00133 [PubMed]
- 90. Patel VG, Oh WK, Galsky MD. Treatment of muscle-invasive and advanced bladder cancer in 2020. CA Cancer J Clin. 2020; 70:404–23. https://doi.org/10.3322/caac.21631 [PubMed]
- 91. Vander Heiden MG, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science. 2009; 324:1029–33. https://doi.org/10.1126/science.1160809 [PubMed]
- 92. Massari F, Ciccarese C, Santoni M, Iacovelli R, Mazzucchelli R, Piva F, Scarpelli M, Berardi R, Tortora G, Lopez-Beltran A, Cheng L, Montironi R. Metabolic phenotype of bladder cancer. Cancer Treat Rev. 2016; 45:46–57. https://doi.org/10.1016/j.ctrv.2016.03.005 [PubMed]
- 93. Dang CV, Le A, Gao P. MYC-induced cancer cell energy metabolism and therapeutic opportunities. Clin Cancer Res. 2009; 15:6479–83. https://doi.org/10.1158/1078-0432.CCR-09-0889 [PubMed]
- 94. Sardi I, Dal Canto M, Bartoletti R, Guazzelli R, Travaglini F, Montali E. Molecular genetic alterations of c-myc oncogene in superficial and locally advanced bladder cancer. Eur Urol. 1998; 33:424–30. https://doi.org/10.1159/000019629 [PubMed]
- 95. Watters AD, Latif Z, Forsyth A, Dunn I, Underwood MA, Grigor KM, Bartlett JM. Genetic aberrations of c-myc and CCND1 in the development of invasive bladder cancer. Br J Cancer. 2002; 87:654–8. https://doi.org/10.1038/sj.bjc.6600531 [PubMed]
- 96. Jeong KC, Kim KT, Seo HH, Shin SP, Ahn KO, Ji MJ, Park WS, Kim IH, Lee SJ, Seo HK. Intravesical instillation of c-MYC inhibitor KSI-3716 suppresses orthotopic bladder tumor growth. J Urol. 2014; 191:510–8. https://doi.org/10.1016/j.juro.2013.07.019 [PubMed]
- 97. von der Lehr N, Johansson S, Wu S, Bahram F, Castell A, Cetinkaya C, Hydbring P, Weidung I, Nakayama K, Nakayama KI, Söderberg O, Kerppola TK, Larsson LG. The F-box protein Skp2 participates in c-Myc proteosomal degradation and acts as a cofactor for c-Myc-regulated transcription. Mol Cell. 2003; 11:1189–200. https://doi.org/10.1016/s1097-2765(03)00193-x [PubMed]
- 98. Welcker M, Orian A, Jin J, Grim JE, Harper JW, Eisenman RN, Clurman BE. The Fbw7 tumor suppressor regulates glycogen synthase kinase 3 phosphorylation-dependent c-Myc protein degradation. Proc Natl Acad Sci USA. 2004; 101:9085–90. https://doi.org/10.1073/pnas.0402770101 [PubMed]
- 99. Paul I, Ahmed SF, Bhowmik A, Deb S, Ghosh MK. The ubiquitin ligase CHIP regulates c-Myc stability and transcriptional activity. Oncogene. 2013; 32:1284–95. https://doi.org/10.1038/onc.2012.144 [PubMed]
- 100. Mei Z, Zhang D, Hu B, Wang J, Shen X, Xiao W. FBXO32 Targets c-Myc for Proteasomal Degradation and Inhibits c-Myc Activity. J Biol Chem. 2015; 290:16202–14. https://doi.org/10.1074/jbc.M115.645978 [PubMed]
- 101. Li M, Yu J, Ju L, Wang Y, Jin W, Zhang R, Xiang W, Ji M, Du W, Wang G, Qian K, Zhang Y, Xiao Y, Wang X. USP43 stabilizes c-Myc to promote glycolysis and metastasis in bladder cancer. Cell Death Dis. 2024; 15:44. https://doi.org/10.1038/s41419-024-06446-7 [PubMed]
- 102. Torre LA, Trabert B, DeSantis CE, Miller KD, Samimi G, Runowicz CD, Gaudet MM, Jemal A, Siegel RL. Ovarian cancer statistics, 2018. CA Cancer J Clin. 2018; 68:284–96. https://doi.org/10.3322/caac.21456 [PubMed]
- 103. Orr B, Edwards RP. Diagnosis and Treatment of Ovarian Cancer. Hematol Oncol Clin North Am. 2018; 32:943–64. https://doi.org/10.1016/j.hoc.2018.07.010 [PubMed]
- 104. Kuroki L, Guntupalli SR. Treatment of epithelial ovarian cancer. BMJ. 2020; 371:m3773. https://doi.org/10.1136/bmj.m3773 [PubMed]
- 105. Lheureux S, Braunstein M, Oza AM. Epithelial ovarian cancer: Evolution of management in the era of precision medicine. CA Cancer J Clin. 2019; 69:280–304. https://doi.org/10.3322/caac.21559 [PubMed]
- 106. Kurnit KC, Fleming GF, Lengyel E. Updates and New Options in Advanced Epithelial Ovarian Cancer Treatment. Obstet Gynecol. 2021; 137:108–21. https://doi.org/10.1097/AOG.0000000000004173 [PubMed]
- 107. Lheureux S, Gourley C, Vergote I, Oza AM. Epithelial ovarian cancer. Lancet. 2019; 393:1240–53. https://doi.org/10.1016/S0140-6736(18)32552-2 [PubMed]
- 108. Pei L, Zhao F, Zhang Y. USP43 impairs cisplatin sensitivity in epithelial ovarian cancer through HDAC2-dependent regulation of Wnt/β-catenin signaling pathway. Apoptosis. 2024; 29:210–28. https://doi.org/10.1007/s10495-023-01873-x [PubMed]
- 109. Lee JH, Kavanagh JJ, Wildrick DM, Wharton JT, Blick M. Frequent loss of heterozygosity on chromosomes 6q, 11, and 17 in human ovarian carcinomas. Cancer Res. 1990; 50:2724–8. [PubMed]
- 110. Russell SE, Hickey GI, Lowry WS, White P, Atkinson RJ. Allele loss from chromosome 17 in ovarian cancer. Oncogene. 1990; 5:1581–3. [PubMed]
- 111. Liu Y, Chen C, Xu Z, Scuoppo C, Rillahan CD, Gao J, Spitzer B, Bosbach B, Kastenhuber ER, Baslan T, Ackermann S, Cheng L, Wang Q, et al. Deletions linked to TP53 loss drive cancer through p53-independent mechanisms. Nature. 2016; 531:471–5. https://doi.org/10.1038/nature17157 [PubMed]