



Introduction
Chronic wounds in patients with diabetes mellitus present a significant clinical challenge. They often result in long-term suffering, adversely affecting patients’ quality of life and imposing a substantial burden on healthcare systems, as the absence of normal stages of healing characterizes these types of wounds. Numerous factors contribute to their preservation, including impaired blood supply, infection, biofilm formation, malnutrition, and underlying medical conditions. In some cases, if these types of wounds are not treated properly, they can lead to severe illness and potential amputation [1].
Antibacterial therapy (ABT) is one of the most critical components of conservative treatment of chronic wounds in diabetic foot syndrome (DFS). In patients with diabetes, an uncontrolled or poorly controlled infection significantly worsens the prognosis of limb preservation and even life. The depth of the lesion, the severity of the disease, and the use of antibiotics in the previous stages of treatment affect the nature of the infection in DFS [2]. Aerobic Gram-positive cocci that colonize the skin are known to be the first to contaminate a wound or skin defect. S. aureus employs various virulence factors, including cytolytic proteins, super-antigenic factors, cellular proteins, and molecules to evade immunity, to cause infectious inflammation in DFS.
Staphylococcus aureus (S. aureus) and beta-hemolytic streptococci of groups A, B, and C are frequently identified in patients with infectious complications of DFS [3]. S. aureus employs various virulence factors, including cytolytic proteins, super-antigenic factors, cellular proteins, and molecules to evade immunity, to cause infectious inflammation in DFS. Antimicrobial resistance (AMR) poses a significant threat as microorganisms evolve defense mechanisms, reducing treatment efficacy and increasing the risk of treatment failure, disease progression, severe illness or death. Misuse and overuse of antimicrobial agents, combined with ineffective infection prevention and control (IPC) practices, are recognized as significant drivers of the increasing prevalence of AMR [4]. Nevertheless, at least five main types of resistance to antibiotics have been described for isolated (planktonic) microbial cells: antibiotic inactivation, target modification, active removal of the antibiotic from the microbial cell (efflux), violation of the permeability of the outer membrane of the microbial cell, and the formation of a metabolic shunt [5–7]. Current scientific knowledge indicates that that almost all types of known ‘planktonic’ resistance are also present in biofilms. The effectiveness of penetration of antibiotics is primarily linked to their ability to overcome the surface membrane and intercellular matrix of biofilms. The latter’s composition contains many different lipids, qualitatively similar to membrane lipids [8]. Despite the relatively simple organization, bacteria exhibit a well-developed cellular structure responsible for biological features and properties. The cellular structure of bacteria is a fundamental focus of classical microbiology, encompassing the study of microorganisms morphology. Bacteria vary in cell wall structure, dividing into two main groups: gram-negative and gram-positive. Gram-positive bacteria are characterized by a thick layer of peptidoglycan and an additional outer membrane containing phospholipids and lipopolysaccharides. The chemical structure of lipopolysaccharides in the outer membrane can be specific and contribute to the antigenic properties of strains [9].
The surface components of the bacterial cell wall serve multiple functions, including acting as a barrier, possessing adhesive properties, accommodating several enzymes, as well as signaling proteins that respond to the factors such as temperature, osmosis, light, oxygen, chemical compounds, and others. The cell wall facilitates all bacterial interactions with the environment. Thus, the primary organizational structure perceives the influence of any external factors. The process of capturing outer material by the cell or protecting against immune factors of the macroorganism is accomplished by forming membrane vesicles. Typically, bacterial vesicles are formed due to protrusion and separation of a fragment of the outer membrane [10, 11].
S. aureus generates and releases extracellular vesicles (EV), which are packed by cytosol. These vesicles are associated with the cell wall and membrane proteins, as well as glycopolymers and exoproteins, including alpha hemolysine, leukocids, phenol-modules, superstigens and enzymes. EV represents a novel secretory system in S. aureus, impacting various stress reactions and facilitating the delivery of biologically active transformative toxins and other determinants of virulence for host cells. Numerous studies have shown that EVs secreted by Methicillin-resistant Staphylococcus aureus (MRSA) are linked to drug resistance in S. aureus. These EVs contain multiple cytokines such as bio-enzyme and drug resistance genes, which are crucial for the bacteria’s survival. Research indicated that stimulation by β-lactam antibiotics creates holes in the peptidoglycan layer of cell wall of S. aureus, which results in the protrusion of cytoplasmic membrane material into the extracellular region, leading to increased production of EVs.
Fulsundar et al. highlighted the potential lytic effect of vesicles on gram-positive bacteria, suggesting a possible mechanism for the development of antibiotic resistance in microorganisms [12]. It is assumed that the lipid component of vesicles can bind to polypeptides of the membrane of gram-positive bacteria. In the case of gram-negative microorganisms, β-lactamases, known enzymes that resist β-lactam antibiotics, are lipoproteins that can also be found in vesicles [13–15].
This study aims to investigate resistance to antibacterial drugs and the role of lytic vesicles as factors of antibiotic resistance.
Materials and Methods
Cultures of S. aureus isolates, and reference strains of S. aureus and B. subtilis were used to determine the lytic effect of vesicles. Clinical isolates of S. aureus were obtained from the wounds of 182 patients with ischemic and mixed forms of DFS, who were treated from 2016 to 2022. All of them suffered from Type II Diabetes Mellitus (DM). According to the classification indicating the severity of peripheral arterial disease, they had Fontaine’s IV degree of chronic ischemia of the lower limb with localization of ulcerative-necrotic lesions of soft tissues in the area of the foot. The average age of the patients was 68 ± 4.6 years; the duration of diabetes was 13 ± 4.4 years. All patients received signed informed consent, and the Ethics Committee of Ukrainian Association of Biobank Austria approved the study (No. 1123-AU). The contents of patients’ wounds were analyzed using MALDI-ToF MS, and the samples were processed on a Bruker Daltonik Biotyper sirius GP or RUO System, Bruker Taxonomy (Bruker Scientific LLC., Billerica, MA, USA).
Vesicles were obtained from inoculums of S. aureus and B. subtilis. Cells from 0.6 l of culture were removed by centrifugation at 4500 g for 20 min and repeated centrifugation of the supernatant at 22 000 g for 25 min. to remove cell debris. Vesicles were isolated from the resulting suspension by centrifugation at 140,000 g for 2 hours at 4° Celcius. Then, the precipitate of vesicles was resuspended in 3 ml of 10 mM Tris-HCl (Sigma-Aldrich offers Roche-10812846001), pH 8.0, and stored at minus 20° Celcius.
Subsequently, vesicle suspensions were isolated from both clinical isolates of S. aureus and reference strains of S. aureus ATCC 25923 and B. subtilis ATCC 6633. These isolates were cultivated on a medium promoting the secretion of bacteriolytic enzymes. The purification of EVs was carried out from S. aureus isolates and reference strains of S. aureus ATCC 25923 and B. subtilis ATCC 6633 cultures harvested during the post-exponential growth phase. To remove non-membranous proteins, protein aggregates, and other contaminants, the EV sample was further purified by OptiPrep™ density gradient (OptiPrep™ is a sterile endotoxin tested solution of 60% iodixanol in water with a density of 1.32 g/ml, OptiPrep™ Density Gradient Medium, Sigma-Aldrich) ultracentrifugation. OptiPrep™ density gradient ultracentrifugation was employed to further purify the EV samples, effectively removing non-membranous proteins, protein aggregates, and other contaminants.
Lowry’s method, based on protein absorption of ultraviolet light with wavelengths of 263 and 280 nm, was used to quantify low protein concentrations in vesicles and membranes of isolates and reference strains [16]. To determine the lytic action of vesicles, 25 μl of vesicle suspension was applied to the lawn of target cells, corresponding to 24 μg of total protein. The plates continued to be incubated at 29° Celcius until the appearance of lysis zones. The adhesive properties of isolates and reference strains of S. aureus were also studied. A quantitative assessment of adhesive activity was carried out according to the method described by V. I. Brilis and co-authors [17].
Determination of genetic markers in clinical isolates of S. aureus was conducted using multiplex polymerase chain reaction (PCR). Accelerated sample preparation was used for DNA isolation. Gel electrophoresis in 1.6% agarose gel stained with ethidium bromide was used to detect cloned DNA samples. An ultraviolet transilluminator TSR-25 M (Vilber Lourmat, France) with a wavelength of 312 nm of view was accustomed to photographing the gels. In the positive control sample, five distinct luminous pink-colored bands were identified, measuring 1000 bp, 745 bp, 512 bp, 360 bp, and 197 bp. Conversely, these bands were absent in the negative control sample. In the studied samples showing a positive PCR result, between one to five luminous bands were observed, positioned according to the genome size at the same level as the bands in the positive control. DNA detection was conducted through electrophoresis, and separation of amplification products was carried out by horizontal electrophoresis in 2% agarose gel. A TSR-25 M transilluminator (Vilber Lourmat, France) with a radiation wavelength of 312 nm was used for viewing and photographing gels. In the examined samples, genes at the chromosomal loci responsible for the formation of resistance to antibacterial drugs were identified, including mes (resistance to cephalosporins), vanA (resistance to glycopeptides), vanB (resistance to vancomycin), OXA-48, blaNDM and VIM (resistance to carbapenems). Plasmid-encoded genes qnrA and qnrB were also identified. The gels were photographed in transmitted ultraviolet light to document the obtained results with a digital video camera connected to a computer.
All the studies were carried out in the biotechnological laboratory of the Ukrainian Association of Biobanks in Austria (Graz, Austria), Institute of Bio-Stem Cell Rehabilitation (Kharkiv, Ukraine).
The study materials underwent statistical processing using parametric analysis methods. Accumulation, correction, systematization of initial information, and visualization of the obtained results were performed in Microsoft Office Excel 2016 spreadsheets. Statistical analysis was conducted using STATISTICA 13.3 software (developer - StatSoft, Inc.). For quantitative indicators with a normal distribution, the obtained data were organized into variation series, in which arithmetic mean (M) standard deviations (S.D.) and 95% confidence interval limits (95% CI) were calculated. When comparing mean values in normally distributed populations, quantitative Student’s t-criterion values were evaluated by comparing them with critical values. Statistically significant differences were considered at a significance level of p < 0.05.
Availability of data and materials
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Results
Table 1 lists the identified resistance genes of clinical S. aureus isolates, and their distribution is shown. The blaCTX-M-2 gene, responsible for cephalosporin-1 resistance, was most frequently detected in clinical strains of S. aureus as the causative agent of wound infection in 60 out of 182 strains of clinical isolates of S. aureus (33%). The differences were statistically significant and reliable (p = 0.026). The second gene controlling resistance to cephalosporins, Mec-1, was less frequently detected in 20 out of 182 strains (frequency 10,9%), and the differences were statistically significant (p = 0.031). Resistance to glycopeptide antibiotics (vancomycin, teicoplanin), coded by vanA and vanB genes, was observed in two studied strains of S. aureus (frequency 10,9%). The erm gene, encoding resistance to macrolides, was detected in 14 strains of S. aureus (7,6%). The differences were statistically significant (p = 0.05). The tet gene, encoding resistance to tetracyclines, was identified in 25 (13,7%) of the studied strains (p = 0.025). The genes tet-38 and 23S rRNA gene, important for macrolides and tetracycline resistance, and the multigene regulators arlR/arlS, mgrA, crucial for multidrug resistance and virulence gene expression, were ubiquitously expressed among the isolates and are not indicated on the table. Plasmid resistance to fluoroquinolones mediated by the qnrB gene was revealed in 12 clinical strains of S. aureus (6,5%) (p = 0.027). Among the analyzed cultures, 11 (6,0%) strains of S. aureus were identified with multiple antibiotic resistance to 4 groups of drugs.
Table 1. Core genome of clinical Staphylococcus aureus isolates identified.
| Screen gene-positive isolates | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| qnrB | blaCTX-M-2 | mec-1 | erm | Van A | Van B | blaOXA-48 | tet | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S. aureus clinical isolates | 12 | 60 | 20 | 14 | 20 | 20 | 11 | 25 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| P-value | 0.027 | 0.026 | 0.031 | 0.05 | 0.031 | 0.032 | 0.005 | 0.025 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Total (n = 182) | 12 | 60 | 20 | 14 | 20 | 20 | 11 | 25 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| *Ubiquitously expressed genes are not indicated (23S, arlR, arlS, mgrA, and tet38). | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Suggestion: Genetic screening of resistance-associated genes in clinical isolates of Staphylococcus aureus. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The study revealed a significant difference in the number of vesicles among all groups, with isolates and reference strains exhibiting a notable disparity (p ≤ 0.05).
Table 2 provides a comprehensive summary of our results, and includes data from genome groups of clinical S. aureus, along with reference strains of S. aureus ATCC 25923 and B. subtilis ATCC 6633, analyzing both the protein content and EVs. Interestingly, the number of vesicles in the reference strains of S. aureus ATCC 25923 was found to be two times smaller than that of the isolates, despite a minimal difference in the protein content within the cell membranes. The results of the mean values for the three samples are presented in Table 2.
Table 2. Protein content in outer membranes and extracellular vesicles of S. aureus ATCC 25923 reference strain, S. aureus clinical strains carrying antibiotic-resistance genes, and B. subtilis reference strain.
| Sample | S. aureus ATCC 25923 | S. aureus blaCTX-M-2 | S. aureus mec-1 | S. aureus erm | S. aureus vanA and vanB | B. subtilis ATCC 6633 |
| Protein, mg/ml | Protein, mg/ml | Protein, mg/ml | Protein, mg/ml | Protein, mg/ml | Protein, mg/ml | |
| Vesicles | 0,48 ± 0.009 | 0,91 ± 0.004 | 1,02 ± 0.005 | 0,96 ± 0.007 | 1,12 ± 0.006 | 0,64 ± 0.006 |
| Outer membrane protein | 42,5 ± 0.06 | 43,02 ± 0.02 | 43,25 ± 0.05 | 43,80 ± 0.05 | 44,09 ± 0.03 | 44,75 ± 0.009 |
As a result of the study, it was established that vesicles exhibited a lytic effect, forming lysis zones around both reference strains of S. aureus ATCC 25923 and clinical isolates of S. aureus. Additionally, lysis zones were observed around vesicles of reference strains of B. subtilis ATCC 6633.
S. aureus EVs were found to mediate the survival of ampicillin-susceptible gram-positive bacteria in the presence of ampicillin. Interestingly, B. subtilis ATCC 6633 shoved susceptibility to both vancomycin hydrochloride and ampicillin. To investigate the role of S. aureus ATCC 25923 EVs in antibiotic resistance, we incubated ampicillin-susceptible B. subtilis ATCC 6633 with the EVs in the presence of ampicillin. The results showed that S. aureus ATCC 25923 EVs mediated the survival of B. subtilis ATCC 6633 in LB-ampicillin (100 μg/ml) medium (A5354, Sigma-Aldrich) in a dose-dependent manner (P < 0.001) (Figure 1B). When S. aureus ATCC 25923 EVs were directly applied to a lawn of B. subtilis ATCC 6633 in LB-ampicillin (100 μg/ml) agar plate (A5354, Sigma-Aldrich), a zone of survived bacteria was revealed (Figure 1C). Notably, no colonies grew on LB and LB-ampicillin (100 μg/ml) agar plate with isolated S. aureus ATCC 25923 EVs (data not shown), indicating that the purified EVs were not contaminated with any bacterial strain.
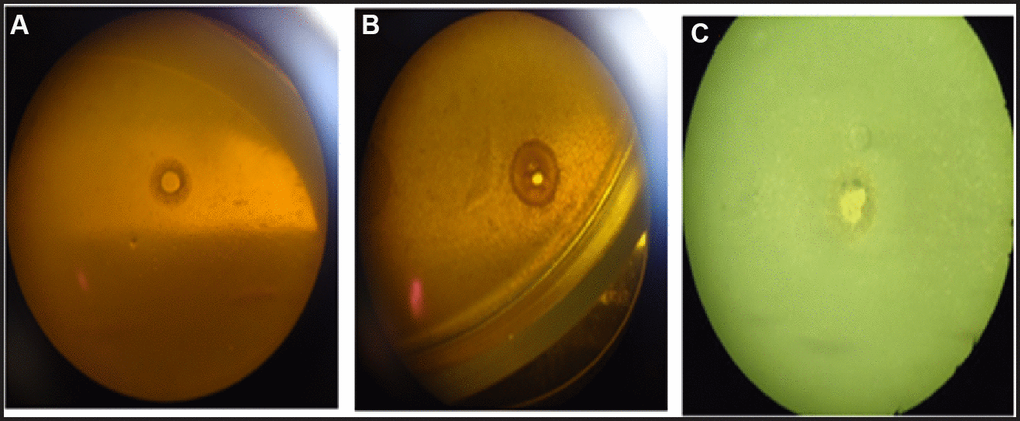
Figure 1. A lytic effect of vesicles: reference strains of S. aureus on B. subtilis (A); isolates of S. aureus on B. subtilis (B); reference strains of B. subtilis on reference strains of S. aureus (C).
Discussion
The study revealed that clinical strains of S. aureus, responsible for wound infection in patients with DFS, harbored resistance genes against almost all groups of antibacterial drugs prescribed for treating these patients. Specifically, blaCTX-M-2 genes were identified in 16.7% of clinical strains, mec-1 in 5.5%, vanA and vanB genes encoding resistance to glycopeptides in 5.5%, while erm genes in 11.1%. Gene qnrB associated with plasmid-mediated resistance to fluoroquinolones was detected in 5.5% of strains. In addition, multiple resistance genes were found in 11.1% of clinical strains of S. aureus.
Although the autolytic systems of both groups of S. aureus were known to be identical, the significantly higher vesicle activity observed in S. aureus clinical isolates suggested the presence of more active extracellular bacteriolytic enzymes. Although the autolytic systems of both groups of S. aureus were known to be identical, the adhesive properties of S. aureus clinical isolates indirectly supported the existence of more active extracellular enzymes. Overall, our findings suggest that vesicles in bacteria act as aggression factors, which in turn allows them to gain an advantage over other bacteria in environmental conditions. This aligns with the findings of Manning & Kuehn, who investigated similar effects in Gram-negative bacteria [18]. These authors demonstrated that outer membrane vesicles produced by Gram-negative bacteria throughout growth played an essential role in virulence, inflammation, and defense responses against external membrane stressors. They found that a hypervascular mutant of Escherichia coli survived treatment with antimicrobial peptides, polymyxin B, and colistin. The authors’ findings demonstrated the role of outer membrane vesicles in innate bacterial defense, and their production might be an essential factor in neutralizing environmental agents targeting the outer membrane of Gram-negative bacteria. Simultaneously, Schaar et al. showed that vesicles secreted by bacteria hydrolyze antibiotics and act as “decoys” for antimicrobial drugs, thereby protecting bacteria from peptides and ABT [19].
At the same time, we observed that vesicles exhibit a more pronounced lytic effect in clinical isolates of S. aureus. We hypothesize that the site of vesicle attachment is likely the concentrated action site of lytic enzymes. Furthermore, it is conceivable that vesicles are associated with peptidoglycans, potentially shielding the contents of the bacterial cell from external influences. Vesicles significantly broaden the range of microorganisms with which a given bacterium can compete in nature and resist 12 microorganism immune factors, as both pathogenic and non-pathogenic bacteria produce outer membrane vesicles as a canonical end product of secretion [20–23].
Staphylococcus aureus extracellular vesicles in interbacterial communication to cope with antibiotic stress. The Lee J and Lee EY (2013) found that S. aureus liberated BlaZ, a β-lactamase protein, via extracellular vesicles. These extracellular vesicles enabled other ampicillin-susceptible Gram-negative bacteria to survive in the presence of ampicillin. However, S. aureus extracellular vesicles did not mediate the survival of tetracycline-, chloramphenicol-, or kanamycin-susceptible bacteria. Moreover, S. aureus extracellular vesicles did not contain the blaZ gene. In addition, the heat-treated S. aureus extracellular vesicles did not mediate the survival of ampicillin-susceptible bacteria. The β-lactamase activities of S. aureus soluble and extracellular vesicle-associated BlaZ were similar, but only the extracellular vesicle-associated BlaZ was resistant to protease digestion, which suggests that the enzymatic activity of BlaZ in extracellular vesicles is largely protected by the vesicle structure. Our observations provide evidence of the important role of S. aureus extracellular vesicles in antibiotic resistance, which allows the polymicrobial community to continue to evolve and prosper against antibiotics.
In the literature analysis of theses in PubMed, no prior studies had reported on the role of S. aureus bacterial EVs in interbacterial communication and genotype variability. In the present study, we showed that S. aureus EVs are genotype-dependent and carry biologically active β-lactamase protein, and these EVs allow other ampicillin-sensitive Gram-positive bacteria (B. subtilis) to survive in the presence of ampicillin. This is the first report demonstrating that S. aureus EVs are genotype dependent and play a functional role in interbacterial communication, similar to Gram-negative bacteria.
Our study confirmed this by revealing differences in the adhesive properties of reference strains of S. aureus and clinical isolates of S. aureus. This expands our understanding of the mechanisms behind antibiotic resistance formation in clinical strains of surgical infection pathogens and may contribute to the development of effective measures against the spread of multidrug-resistant pathogens.
Limitations of the study
Most of our investigations into the ability of S. aureus to form membrane vesicles were conducted on a small clinic group with limited number of patients. We believe that future studies should delve into the contribution of the relevant genes as described in the context of adhesion and membrane vesicle formation, and their importance in the pathogenesis of wound healing. This applies not only to patients with DFS but also to those with wounds and infections of other origins, as recommended by the worldwide cross-sectional survey in 2022 [24].
Conclusions
In this study, we investigated the ability of membrane vesicle formation with lytic action in S. aureus isolates from patients with DFS. While S. aureus isolates are known for their resistance to most β-lactam antibiotics, the existence of different virulence factors, which could potentially complicate infections, remains a topic of debate. We explored antibiotic resistance and the role of lytic vesicles as AMR factors in cultures of S. aureus clinical isolates and reference strains. Utilizing B. subtilis reference strain, we aimed to determine the lytic effect of vesicles in cases of ischaemic and mixed forms of DFS. The results revealed that vesicles have a lytic effect, forming lysis zones around both vesicles of S. aureus reference strains and S. aureus clinical isolates and around B. subtilis reference strains. Furthermore, we identified genes blaCTX-M-2, Mec-1, vanA, vanB, erm encoding resistance to different types of antibiotics, along with qnrB gene associated with plasmid-mediated resistance to fluoroquinolones. Notably, multiple resistance to this category of antimicrobial agents has been identified in 11.1% of clinical strains of S. aureus.
Author Contributions
IAK, SG, and YVI constructed the study design. IAK, AMF and YVI contributed to data interpretation and manuscript drafting. YVI, MS and SG, contributed to the statistical analysis. IAK, AMF, SG, and MNB prepared figures. AMF, SG, and YVI participated in the clinical investigation and contributed to the epidemiological data collection. SG, AMF, and MNB revised the manuscript. All experiments mentioned in the article were conducted in the Biotechnology and Microbiology Laboratories of the Ukrainian Association of Biobanks, with the involvement of the article’s authors. All laboratories hold the necessary licenses and accreditations required for this type of research. All authors read and approved the final manuscript.
Acknowledgments
The authors would like to thank the patients and clinical staff who gave their time to take part in this clinical study.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Ethical Statement and Consent
This all-institution retrospective cohort study was handled in accordance with the Declaration of Helsinki. The use of registered data follows the General Data Protection Regulation of the European Union. This research was part of the research work of the Bioethics Committee of the Ukrainian Association of Biobanks which reviewed and approved the research protocol pilot project on quality control of stem cells and cell lines developed and implemented in partnership with the Ukrainian Association of Biobanks (UAB), Institute of Bio-Stem Cell Rehabilitation, International Biobanking, Department of Surgery No. 1, Kharkiv National Medical University (protocol no. 1123-AU, dated 11.11.2021).
Funding
This research was part of the research work of the Ukraine Association of Biobank “Mechanisms of Antibiotic Resistance in Clinical Isolates: The Influence of Extracellular Vesicles and Genotypic Profiles in Staphylococcus aureus from Diabetic Foot Syndrome Patients”. The registration number is UAB_IKB832904y. This research received no external funding.
References
- 1. Spanos K, Saleptsis V, Athanasoulas A, Karathanos C, Bargiota A, Chan P, Giannoukas AD. Factors Associated With Ulcer Healing and Quality of Life in Patients With Diabetic Foot Ulcer. Angiology. 2017; 68:242–50. https://doi.org/10.1177/0003319716651166 [PubMed]
- 2. Ivanova Y, Gramatiuk S, Kryvoruchko I, Tymchenko M, Goltsev K, Sargsyan K. Investigating the joint application of negative pressure wound treatment and tissue therapy for chronic wounds in patients with diabetes. J Med Life. 2023; 16:1098–104. https://doi.org/10.25122/jml-2023-0020 [PubMed]
- 3. Selva Olid A, Solà I, Barajas-Nava LA, Gianneo OD, Bonfill Cosp X, Lipsky BA. Systemic antibiotics for treating diabetic foot infections. Cochrane Database Syst Rev. 2015; 2015:CD009061. https://doi.org/10.1002/14651858.CD009061.pub2 [PubMed]
- 4. Worldwide Antimicrobial Resistance National/International Network Group (WARNING) Collaborators. Ten golden rules for optimal antibiotic use in hospital settings: the WARNING call to action. World J Emerg Surg. 2023; 18:50. https://doi.org/10.1186/s13017-023-00518-3 [PubMed]
- 5. Chopra I, Roberts M. Tetracycline antibiotics: mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol Mol Biol Rev. 2001; 65:232–60. https://doi.org/10.1128/MMBR.65.2.232-260.2001 [PubMed]
- 6. Antibiotic resistance threats in the United States, 2019. Corporate Authors(s): Centers for Disease Control and Prevention (U.S.); National Center for Emerging Zoonotic and Infectious Diseases (U.S.). Division of Healthcare Quality Promotion. Antibiotic Resistance Coordination and Strategy Unit.Published Date: 2019. stacks.cdc.gov/view/cdc/82532.
- 7. Sharma S, Mohler J, Mahajan SD, Schwartz SA, Bruggemann L, Aalinkeel R. Microbial Biofilm: A Review on Formation, Infection, Antibiotic Resistance, Control Measures, and Innovative Treatment. Microorganisms. 2023; 11:1614. https://doi.org/10.3390/microorganisms11061614 [PubMed]
- 8. Lewis K. Persister cells. Annu Rev Microbiol. 2010; 64:357–72. https://doi.org/10.1146/annurev.micro.112408.134306 [PubMed]
- 9. Hancock RE. The bacterial outer membrane as a drug barrier. Trends Microbiol. 1997; 5:37–42. https://doi.org/10.1016/S0966-842X(97)81773-8 [PubMed]
- 10. Wagner T, Joshi B, Janice J, Askarian F, Škalko-Basnet N, Hagestad OC, Mekhlif A, Wai SN, Hegstad K, Johannessen M. Enterococcus faecium produces membrane vesicles containing virulence factors and antimicrobial resistance related proteins. J Proteomics. 2018; 187:28–38. https://doi.org/10.1016/j.jprot.2018.05.017 [PubMed]
- 11. Liu Y, Defourny KAY, Smid EJ, Abee T. Gram-Positive Bacterial Extracellular Vesicles and Their Impact on Health and Disease. Front Microbiol. 2018; 9:1502. https://doi.org/10.3389/fmicb.2018.01502 [PubMed]
- 12. Fulsundar S, Harms K, Flaten GE, Johnsen PJ, Chopade BA, Nielsen KM. Gene transfer potential of outer membrane vesicles of Acinetobacter baylyi and effects of stress on vesiculation. Appl Environ Microbiol. 2014; 80:3469–83. https://doi.org/10.1128/AEM.04248-13 [PubMed]
- 13. Vasilyeva NV, Tsfasman IM, Suzina NE, Stepnaya OA, Kulaev IS. Secretion of bacteriolytic endopeptidase L5 of Lysobacter sp. XL1 into the medium by means of outer membrane vesicles. FEBS J. 2008; 275:3827–35. https://doi.org/10.1111/j.1742-4658.2008.06530.x [PubMed]
- 14. Kim JH, Lee J, Park J, Gho YS. Gram-negative and Gram-positive bacterial extracellular vesicles. Semin Cell Dev Biol. 2015; 40:97–104. https://doi.org/10.1016/j.semcdb.2015.02.006 [PubMed]
- 15. Kim SW, Lee JS, Park SB, Lee AR, Jung JWS, Chun JH, Lazarte JM, Kim J, Seo JS, Kim JH, Song JW, Ha MW, Thompson KD, et al. The Importance of Porins and β-Lactamase in Outer Membrane Vesicles on the Hydrolysis of β-Lactam Antibiotics. Int J Mol Sci. 2020; 21:2822. https://doi.org/10.3390/ijms21082822 [PubMed]
- 16. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Biol Chem. 1951; 193:265–75. [PubMed]
- 17. Brilis VI, Brilene TA, Lentsner Kh, Lentsner AA. [Method of studying the adhesive process of microorganisms]. Lab Delo. 1986; 210–2. [PubMed]
- 18. Manning AJ, Kuehn MJ. Contribution of bacterial outer membrane vesicles to innate bacterial defense. BMC Microbiol. 2011; 11:258. https://doi.org/10.1186/1471-2180-11-258 [PubMed]
- 19. Schaar V, Nordström T, Mörgelin M, Riesbeck K. Moraxella catarrhalis outer membrane vesicles carry β-lactamase and promote survival of Streptococcus pneumoniae and Haemophilus influenzae by inactivating amoxicillin. Antimicrob Agents Chemother. 2011; 55:3845–53. https://doi.org/10.1128/AAC.01772-10 [PubMed]
- 20. Kim SW, Park SB, Im SP, Lee JS, Jung JW, Gong TW, Lazarte JMS, Kim J, Seo JS, Kim JH, Song JW, Jung HS, Kim GJ, et al. Outer membrane vesicles from β-lactam-resistant Escherichia coli enable the survival of β-lactam-susceptible E. coli in the presence of β-lactam antibiotics. Sci Rep. 2018; 8:5402. https://doi.org/10.1038/s41598-018-23656-0 [PubMed]
- 21. Subsomwong P, Teng W, Ishiai T, Narita K, Sukchawalit R, Nakane A, Asano K. Extracellular vesicles from Staphylococcus aureus promote the pathogenicity of Pseudomonas aeruginosa. Microbiol Res. 2024; 281:127612. https://doi.org/10.1016/j.micres.2024.127612 [PubMed]
- 22. Anand D, Chaudhuri A. Bacterial outer membrane vesicles: New insights and applications. Mol Membr Biol. 2016; 33:125–37. https://doi.org/10.1080/09687688.2017.1400602 [PubMed]
- 23. Kulkarni HM, Nagaraj R, Jagannadham MV. Protective role of E. coli outer membrane vesicles against antibiotics. Microbiol Res. 2015; 181:1–7. https://doi.org/10.1016/j.micres.2015.07.008 [PubMed]
- 24. Sartelli M, Labricciosa FM, Coccolini F, Coimbra R, Abu-Zidan FM, Ansaloni L, Al-Hasan MN, Ansari S, Barie PS, Caínzos MA, Ceresoli M, Chiarugi M, Claridge JA, et al. It is time to define an organizational model for the prevention and management of infections along the surgical pathway: a worldwide cross-sectional survey. World J Emerg Surg. 2022; 17:17. https://doi.org/10.1186/s13017-022-00420-4 [PubMed]