



Introduction
Although adjuvant chemotherapy has improved osteosarcoma survival rate in recent years, development of multidrug resistance severely impacts prognosis and restricts success of curative attempts [1–3]. Therefore, new and effective drugs to treat osteosarcoma are clearly needed.
Naringin, a bioflavonoid abundant in grapefruit and other citrus, has multiple biological activities. It possesses sedative, antifungal, antispasmodic, and analgesic properties, and provides cardioprotective, neuroprotective, and anticancer effects [4]. In addition, naringin has been demonstrated to inhibit inflammatory responses, prevent bone degeneration, and exert anabolic effects on bone cells [5,6]. Naringin promotes the expression of β-catenin and increase Ser552 phosphorylation on β-catenin in UMR-106 osteosarcoma cells. This led to activation of lymphoid enhancer factor (LEF)/T-cell factor (TCF) transcription factors to stimulate bone development [7]. Naringin abrogates osteoclastogenesis and bone resorption by inhibiting RANKL-induced NF-κB and ERK activation [8], and demonstrated therapeutic potential to attenuate polymethylmethacrylate-induced osteoclastogenesis and osteolysis. There is substantial evidence supporting a role for naringin as an anticancer agent. Studies also indicated that naringin could reduce the release of inflammatory factors and inhibit the growth of W256 carcinosarcoma in rats [4,9,10]. Moreover, growth arrest and apoptosis were common effects of naringin in several in vitro and in vivo studies conducted on breast, cervical, ovarian, bladder, hepatic, skin, colorectal, and gastric cancer cells [11,12].
Zeb1 (zinc finger E-box binding homeobox 1) is a transcription factor that represses epithelial differentiation and promotes a mesenchymal phenotype [13]. Zeb1 is upregulated in several cancers, where it influences cell motility, cell cycle, and survival, and is an important contributor to tumor invasion and metastasis [14,15].
Studies have shown that Zeb1 can override the G1 checkpoint directly, by stimulating Cyclin D1 expression, and indirectly, by regulating the Wnt signaling pathway [16,17]. Zeb1 was shown to promote the progression of lung cancer by increasing the expression of MMP2, a member of the matrix metalloproteinases family that play an important role in cell migration and facilitate invasion and metastasis of tumor cells [18,19]. Zeb1 has also been shown to be upregulated in osteosarcoma, and to contribute to its development [20,21].
Using human osteosarcoma cell lines as experimental model, in the present study we provide in vitro and in vivo evidence that naringin suppresses proliferation and metastasis of osteosarcoma cells by inhibiting the expression of Zeb1. Our findings highlight the potential of naringin, a safe and natural flavonoid, for osteosarcoma therapy.
Results
Naringin inhibits the expression of Zeb1 in osteosarcoma cells
The expression of Zeb1 in human osteosarcoma samples was assessed by Western blot and real-time PCR (Figs. 1A, B). Both assays showed that Zeb1 was overexpressed in most samples, although heterogeneity was evident. In cultured cells, both Western blot and real-time PCR showed much stronger Zeb1 expression in osteosarcoma MG63 and U2OS cells than in control hFOB1.19 osteoblasts (Figs. 1C, D). Upon exposure to naringin (10 or 20 μmol/L) for 24 h, Zeb1 protein and mRNA levels were dramatically decreased, in dose-dependent manner, in both osteosarcoma cell lines (Figs. 1 E-H).
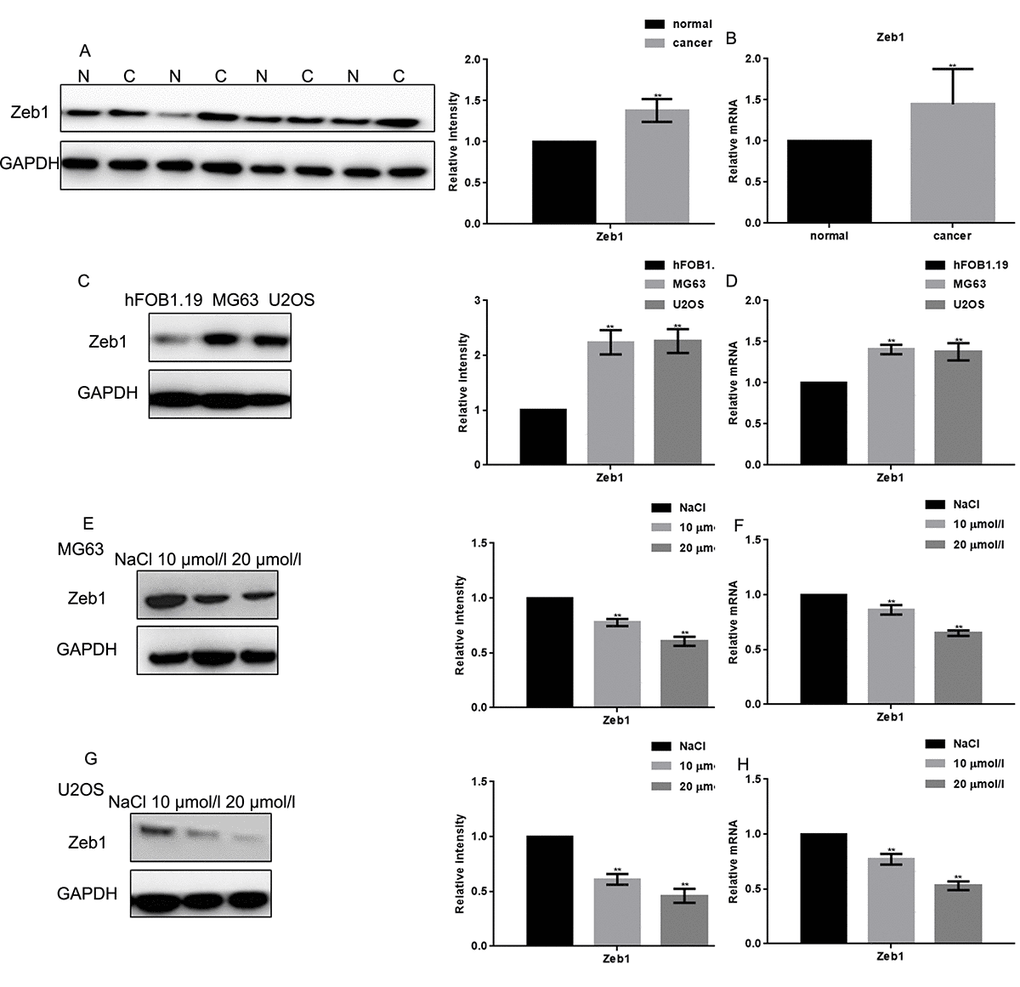
Figure 1. Naringin inhibits the expression of Zeb1 in osteosarcoma cells. (A, B) Zeb1 expression in 30 human osteosarcoma specimens and their adjacent normal tissue counterparts was detected by Western blot and real-time PCR. **P < 0.05, vs normal tissues. (C, D) Zeb1 expression in MG63, U2OS and hFOB1.19 cells, detected by Western blot and real-time PCR. **P < 0.05, vs hFOB1.19 cells. (E-H) Zeb1 expression detected by Western blot and real-time PCR in MG63 and U2OS cells treated with NaCl or indicated concentrations of naringin for 24 h. **P < 0.05, compared with NaCl.
Naringin inhibits proliferation and induces apoptosis in osteosarcoma cells
The MTT assay revealed that naringin treatment inhibited the proliferation of MG63 and U2OS cells in a concentration dependent manner (Fig. 2A). The inhibitory effect of naringin on the proliferation of hFOB1.19 was only obvious when the concentration of naringin was 20 μmol/L. The IC50 of naringin on MG63 and U2OS cells at 24 h was ~50 μmol/L and ~30 μmol/L, respectively (Fig. 2B). Next, we used flow cytometry to evaluate cell cycle staging in PI-stained MG63 and U2OS cells previously exposed to various concentrations of naringin for 24 h. Naringin induced a dose-dependent increase in the percentage of cells in G1 phase, and decreased the number of cells in S phase, compared to control (Figs. 2C, D). To assess whether naringin can promote apoptosis, flow cytometry was used in Annexin-V-FITC-stained osteosarcoma cells. Results showed a dose-dependent increase in apoptotic cells treated with naringin (Figs. 2E, F). In line with these antiproliferative and pro-apoptotic effects, both Western blot and real-time PCR assays showed that exposure to 10 or 20 μmol/L naringin for 24 h dramatically decreased the expression of Cyclin D1 and bcl-2 (Figs. 2G-J).
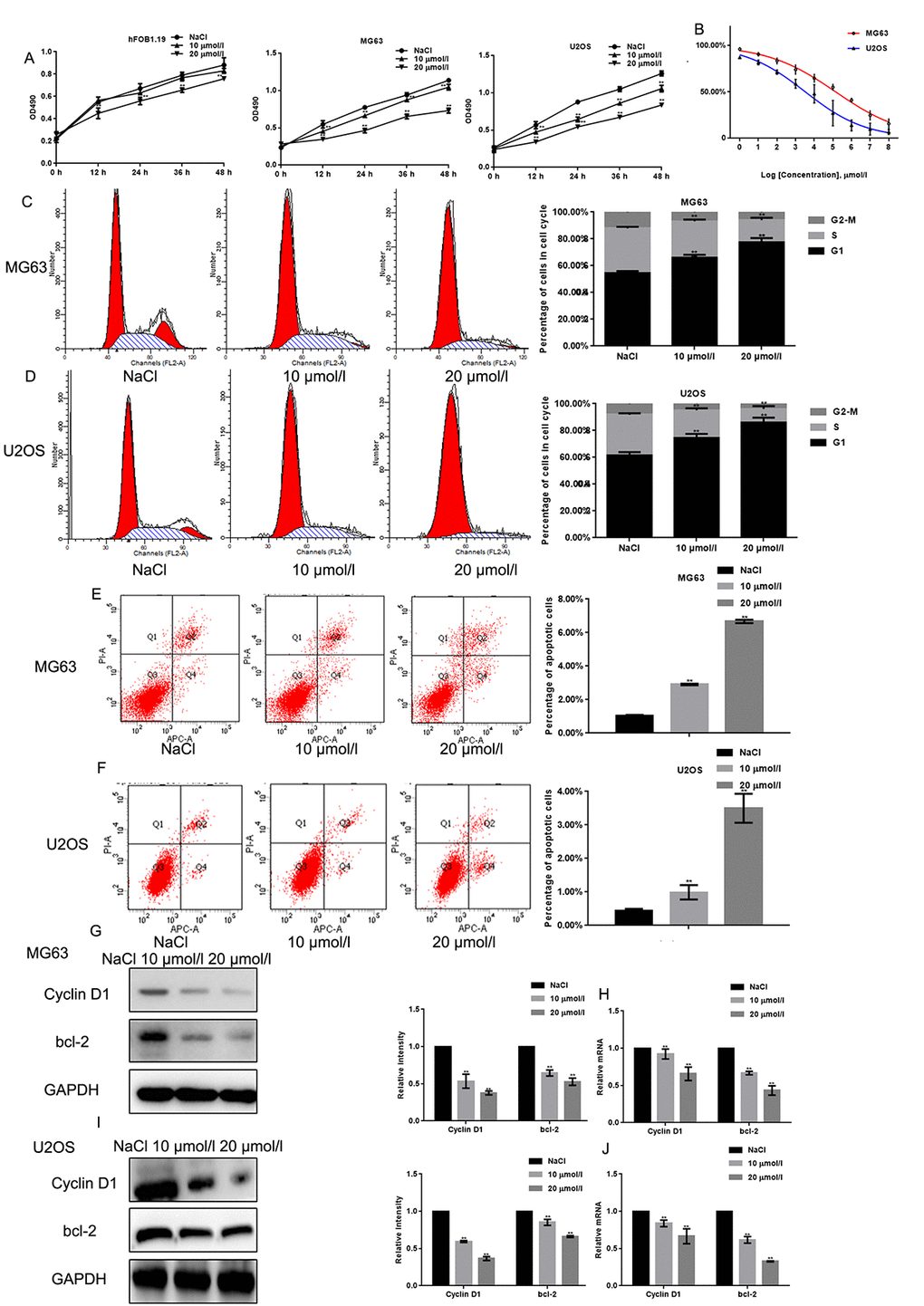
Figure 2. Naringin inhibits the proliferation of osteosarcoma cells. (A) Results of MTT proliferation assays in hFOB1.19, MG63, and U2OS cells cultured with various concentrations of naringin for different times. Results represent the mean ± SD of three experiments done in triplicate. **P < 0.05, compared with NaCl. (B) Proliferation inhibition rates induced by naringin on MG63 and U2OS cells. IC50 values were calculated through linear regression. (C, D) Flow cytometric analysis of cell cycle distribution in MG63 and U2OS cells pre-incubated with or without naringin for 24 h and stained with PI. The experiment was repeated three times. **P < 0.05, compared with NaCl. (E, F) Flow cytometric assay of apoptosis in MG63 and U2OS cells pre-incubated with or without naringin for 24 h and stained with Annexin V-FITC/PI. The experiment was repeated three times. **P < 0.05, compared with NaCl. (G-J) Expression of Cyclin D1 and bcl-2 detected by Western blot and real-time PCR in MG63 and U2OS cells treated with NaCl or naringin for 24 h. **P < 0.05, compared with NaCl.
Naringin inhibits migration of osteosarcoma cells
The effects of naringin on osteosarcoma cell migration and invasion was assessed using Transwell assays in the absence or presence, respectively, of Matrigel. Results showed that naringin exposure (10 or 20 μmol/L for 24 h) significantly decreased both migration and invasion of MG63 and U2OS cells in a dose-dependent manner (Figs. 3A-D). These effects were consistent with a decrease in MMP2 expression, detected in both cell lines in Western blot, real-time PCR, and zymography gel assays (Figs. 3E-H).
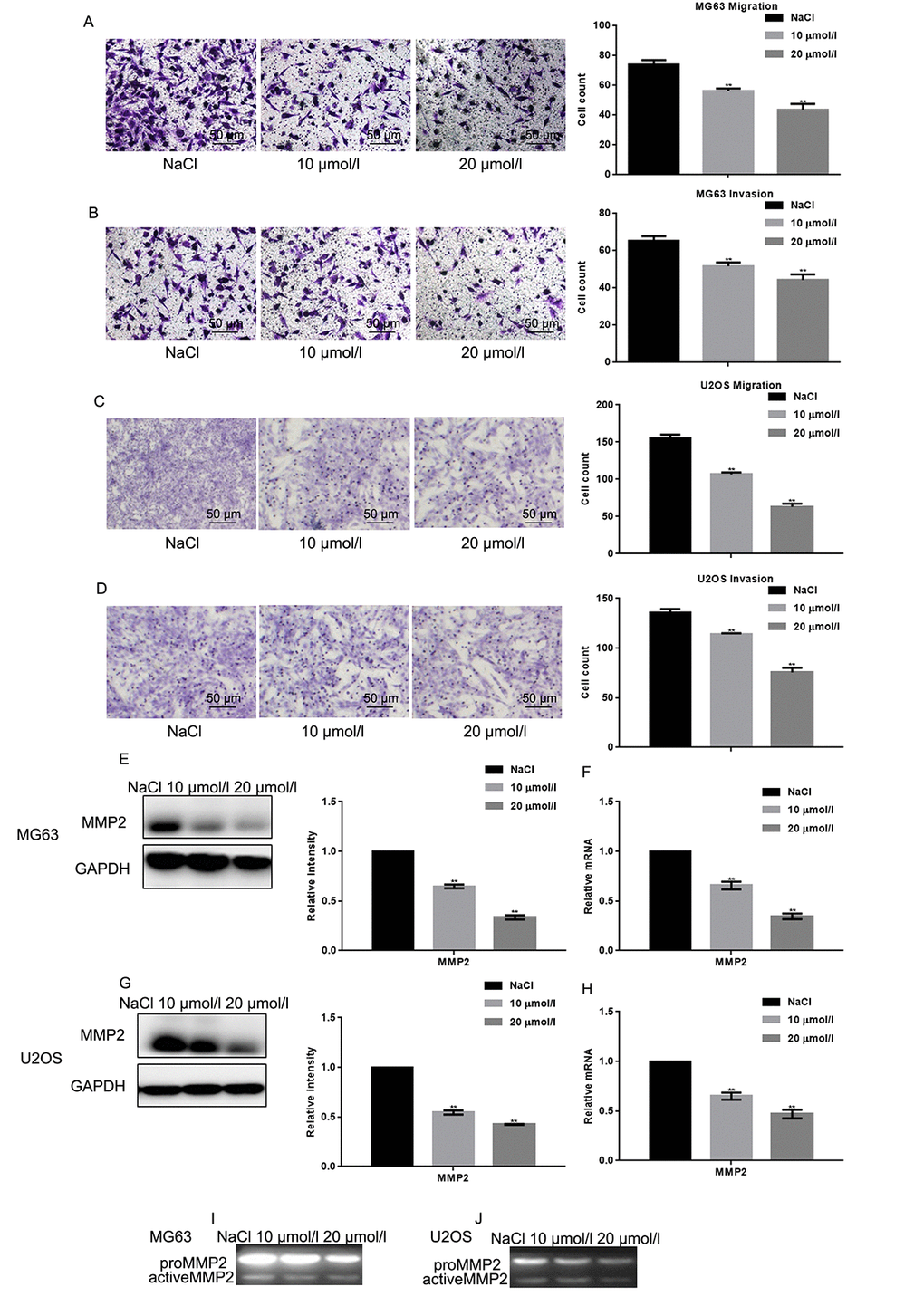
Figure 3. Naringin inhibits migration and invasion of osteosarcoma cells. (A-D) Migration and invasion were studied, respectively, using Transwell assays with or without Matrigel, in naringin-treated MG63 and U2OS cells. Cell count results represent the mean ± SD of three experiments. **P < 0.05, compared with NaCl. (E-H) MMP2 expression by Werstern blot and real-time PCR in MG63 and U2OS cells treated with NaCl or naringin for 24 h. **P < 0.05, compared with NaCl. (I, J) Zymography gel assay showing the inhibitory effect of naringin on MMP2 activity in MG63 and U2OS cells.
Naringin suppresses osteosarcoma cell proliferation and migration by inhibiting Zeb1
To test the hypothesis that naringin exerts antiproliferative and anti-invasive effects by inhibiting Zeb1, its effects were tested in MG63 cells transfected with a plasmid encoding Zeb1 (Zeb1 overexpression) or an empty vector backbone (control). Conversely, siRNAs were introduced to downregulate Zeb1 (si-Zeb1), and to serve as non-targeted, negative control (si-NC). MTT assays showed that proliferation was stimulated by Zeb1 overexpression, and decreased to control inhibition levels by naringin (20 μmol/L) (Fig. 4A). On the other hand, Zeb1 suppression, although partial, decreased cell proliferation, and naringin induced no further inhibition in these cells. (Fig. 4B). We next examined whether migration was affected in Zeb1-overexpressing and Zeb1-suppressed cells. Transwell assay results indicated that Zeb1 overexpression enhances the migration of MG63 cells, while Zeb1 silencing recapitulates the inhibitory effect of naringin on control cells (Figs. 4C, D). Finally, we analyzed the effects of Zeb1 overexpression and downregulation on Cyclin D1, MMP2, and bcl-2 expression. The results showed that expression of these proteins increased upon Zeb1 upregulation (Figs. 4 E-F). Meanwhile, Zeb1 silencing lowered protein expression to levels like those observed in naringin-treated control cells (Figs. 4 G-H).
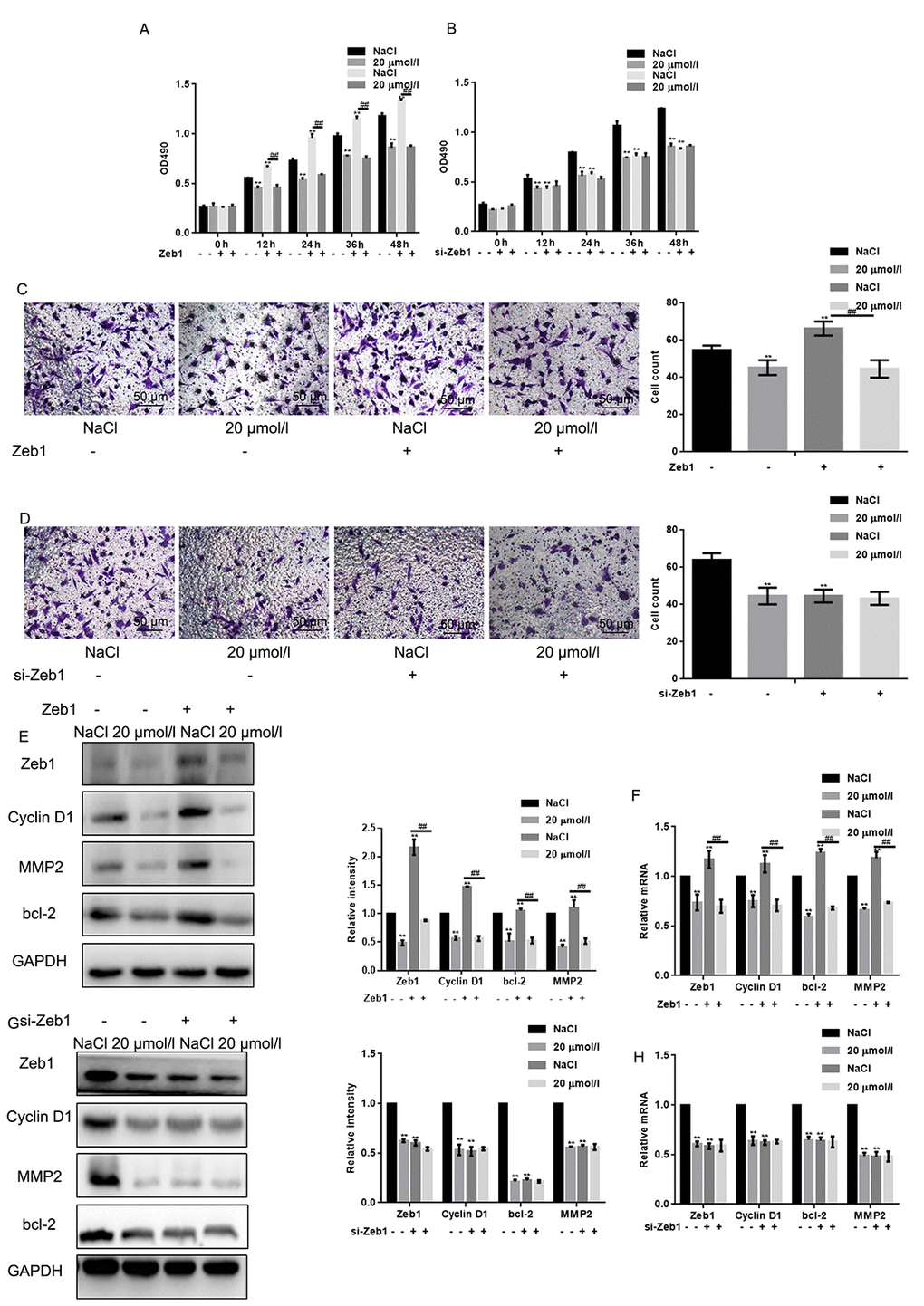
Figure 4. Naringin suppresses osteosarcoma cell proliferation and migration by inhibiting Zeb1. (A) MTT proliferation assay of MG63 cells expressing a control vector or Zeb1. Cells were incubated with 20 μmol/L of naringin or NaCl and assayed at the indicated times. Results represent the mean ± SD of three experiments done in triplicate. **P < 0.05, compared with NaCl. ## P < 0.05, compared with Zeb1 overexpressed group. (B) MTT proliferation assay of MG63 cells transfected with si-Zeb1 (Zeb1 silencing) or si-NC (negative control). Cells were treated with 20 μmol/L of naringin or NaCl and assayed at the indicated times. Results represent the mean ± SD of three experiments done in triplicate. **P < 0.05, compared with NaCl. (C) Results of Transwell migration assays (without Matrigel) performed in MG63 cells expressing control vectors or Zeb1. Cells were treated with 20 μmol/l of naringin or NaCl. **P < 0.05, compared with NaCl; ## P < 0.05, compared with Zeb1 overexpressed group. (D) Results of Transwell migration assays (without Matrigel) performed in MG63 cells transfected with si-Zeb1 (Zeb1 silencing) or si-NC (negative control). Cells were treated with 20 μmol/l of naringin or NaCl. **P < 0.05, compared with NaCl. (E, F) Western blot and real-time PCR assay results for Zeb1, Cyclin D1, bcl-2, and MMP2 expression in MG63 cells expressing Zeb1 or empty vector. Cells were incubated with 20 μmol/l of naringin or NaCl (control). **P < 0.05, compared with NaCl; ## P < 0.05, compared with Zeb1 overexpressed group. (G, H) Western blot and real-time PCR assay results for Zeb1, Cyclin D1, bcl-2, and MMP2 expression in MG63 cells transfected with si-Zeb1 or si-NC. Cells were treated with 20 μmol/l of naringin or NaCl. **P < 0.05, compared with NaCl.
Naringin inhibits the invasion of MG63 cells in vivo
To examine the effect of naringin on osteosarcoma cell tumorigenesis in vivo, MG63 cells were injected into nude mice via the tail vein. After daily administration of naringin (5 or 10 mg/kg) or 0.9% NaCl for 16 days, mice were sacrificed and lung tissues processed for microscopic histological analysis. Results showed that naringin significantly prevented lung degeneration and reduced the incidence of metastatic nodules (Fig. 5A). Moreover, Western blots (Fig. 5B) and real-time PCR assays (Fig. 5C) showed that the expression of Zeb1, Cyclin D1, MMP2, and bcl-2 was decreased in the livers of mice treated with naringin.
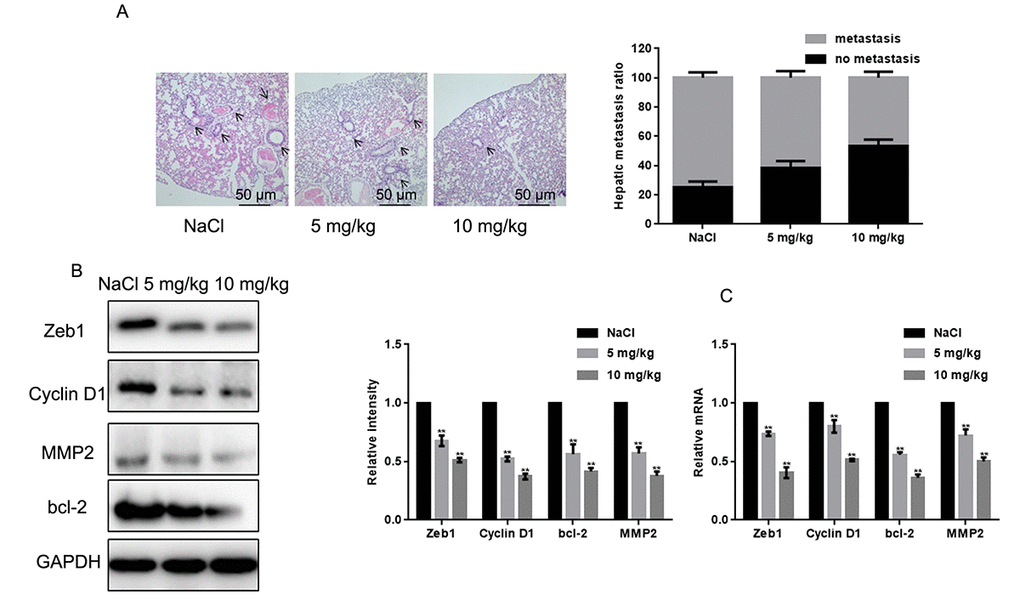
Figure 5. Naringin inhibits lung invasion by MG63 cells in vivo. (A) Representative images of lung histopathology (H&E staining; 400×) from mice injected with MG63 cells and treated daily with two different doses of naringin or NaCl (control) for 16 days. (B, C) Zeb1, Cyclin D1, bcl-2, and MMP2 expression in liver tumor samples, detected by Western blot (B) and real-time PCR (C). **P < 0.05, compared with NaCl-treated controls.
Discussion
Naringin, a flavonoid present in citrus fruits, has HMG-CoA reductase inhibitor activity, and at low (nM) doses increases osteogenic activities in an osteoblast cell model in vitro [22]. Moreover, naringin-induced osteogenic differentiation has been recently described in bone marrow stromal cells and stem cells [23].On the other hand, naringin has been shown to inhibit cell proliferation and promote cell apoptosis in breast cancer, cervical cancer, melanoma, and bladder cancer cells [24]. Thus, we speculated that naringin may have therapeutic effect on osteosarcoma as well.
In this study, we tested the hypothesis that naringin has anticancer actions through inhibition of Zeb1, a zinc finger homeodomain transcription factor implicated in invasiveness and metastasis development in several tumor types, including osteosarcoma and lung cancer [25–27]. We first confirmed high Zeb1 expression in human specimens and in human MG63 and U2OS osteosarcoma cell lines. In these cells, naringin dose-dependently inhibited the expression of Zeb1, reduced proliferation by arresting the cell cycle in the G1 phase, and promoted apoptosis. In contrast, naringin had weaker effects on normal osteoblasts. The induction of apoptosis by naringin correlated with a decrease in anti-apoptotic bcl-2 protein expression; the latter was also observed in a study evaluating naringin’s effects in an ovarian cancer mouse model [12]. Naringin-mediated apoptosis has been documented in cervical cancer cells, in two studies that implicated NF-κB/COX-2-caspase-1 pathway repression [9] and expression of caspases, p53, Bax, and Fas death receptor [28] respectively.
Our results also showed that both naringin exposure and Zeb1 silencing significantly suppressed osteosarcoma cell migration in Transwell assays. This is consistent with the down-regulation of MMP2, observed under both conditions, and the stimulation of MMP2 expression seen instead after ectopic expression of Zeb1 in these cells. These effects may be related to a well-known role of Zeb1 in promoting metastasis through epithelial-to-mesenchymal transition (EMT), although more research is needed to clarify the mechanisms at play in osteosarcoma [29].
Importantly, we demonstrated that naringin’s effects in vitro correlated with antimetastatic actions in vivo, as its administration to nude mice injected with osteosarcoma MG63 cells attenuated the formation of tumor nodules in the liver. In summary, our data showed that naringin inhibits the malignant phenotype of osteosarcoma cells by inhibiting the expression of Zeb1 and Zeb1-associated proteins such as Cyclin D1 and MMP2. The present findings support the potential of naringin as a novel therapeutic strategy for osteosarcoma.
Materials and Methods
Drugs and reagents
Naringin, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT), propidium iodide (PI), and Trypan Blue were purchased from Beyotime Biotechnology (Shanghai, China). Fetal bovine serum (FBS), Dulbecco’s modified Eagle’s medium (DMEM), Lipofectamine 2000, and TRIzol were obtained from Invitrogen (Carlsbad, CA, USA). RT-PCR kits were obtained from Promega (Beijing, China). SYBR Premix Ex Taq reagents were obtained from TaKaRa (Dalia, China). Anti-Zeb1, anti-cyclin D1, anti-bcl-2, anti-MMP2, and anti-GAPDH antibodies were purchased from Santa Cruz Biotechnology (Dallas, TX, USA). HRP-conjugated secondary antibody, BCA protein assay kit, and enhanced chemiluminescence (ECL) solution were purchased from Beyotime Biotechnology. All experiments were completed in the Central Laboratory of our hospital.
Human samples
Tissue samples were obtained from patients undergoing surgery at Shengjing Hospital of China Medical University (Table 1). The original histopathologic reports were obtained from each case, and the diagnosis of osteosarcoma was confirmed. Part of the excised tissue was embedded in paraffin, and part of the sample was snap-frozen at −80°C. Clinical samples were collected after written informed consent was obtained, and the study was approved by the Ethics Committee at the Academic Medical Center of Shengjing Hospital of China Medical University.
Table 1. Patient information.
| Group | N | Percent | |
| Sex | Male | 13 | 65% |
| Female | 7 | 35% | |
| Age | <20 | 12 | 60% |
| >20 | 8 | 40% | |
| History | Yes | 1 | 5% |
| No | 19 | 95% | |
| Site of primary disease | Tibia | 11 | 55% |
| Femur | 7 | 35% | |
| Humerus | 2 | 10% | |
| TNM | I | 9 | 45% |
| II | 7 | 35% | |
| III | 5 | 25% | |
Cell culture
Human osteosarcoma MG63 and U2OS cells, and human hFOB1.19 osteoblasts (SV40-transfected), used as control, were supplied by the Cell Pool Bank of China (Guangzhou, China). The cells were cultured in DMEM supplemented with 10% FBS at 37°C under an atmosphere of 5% CO2 and 95% air.
Cell viability assay
The MTT assay was employed to assess cell viability. Cells were cultured in 96-well plates at a concentration of 1×104 cells/ml and incubated with 10 or 20 µmol/L naringin for 12, 24, 36, or 48h. At those time intervals, 0.01 ml of MTT solution (5 mg/ml) was added to each well. After a 4 h incubation at 37 °C, medium was replaced by 0.15 ml DMSO. After 15 min incubation at 37 °C, optical densities (490 nm) were measured.
Cell cycle assay
Cells were incubated with 10 or 20 µmol/L naringin for 24 h and fixed in 75% ethanol at 4°C overnight. After resuspension in 10 μg/ml PI, cell cycle stages were determined using a FACS Vantage flow cytometer using CellQuest (Becton Dickinson and Co., San Jose, CA, USA).
Apoptosis assay
Cells were incubated with 10 or 20 µmol/L naringin for 24 h, washed twice with cold PBS, and stained with 5 μl ANNEXIN-V-FITC/10 μl PI for 15 min. After addition of 400 μl binding buffer to each tube, the apoptosis rate was measured by flow cytometry within 1 h.
Transwell migration assay
Transwell assays were performed using a modified Boyden chamber with Nuclepore polycarbonate membranes. After 24 h treatment with 10 or 20 µmol/L naringin, 1×105 cells in 100 μl FBS-free DMEM were placed in the upper part of the chamber with or without Matrigel, whereas the lower compartment was filled with 600 μl DMEM containing 10% FBS. After 8 h incubation at 37°C, the invading cells on the lower surface of the filter were fixed, stained with Trypan Blue, and counted under high-power magnification (400×).
Zymography
Cells were cultured in 12-well plates and treated with 10 or 20 µmol/L naringin. After 24 h, media was changed into DMEM containing 5% FBS (the source of proMMP2). After another 24 h, the media were harvested, cleared by centrifugation at 12,000 rpm for 10 min, and subjected to analysis by SDS-PAGE impregnated with 1 mg/ml gelatin. The gels were incubated at 37°C overnight, stained with Coomassie Blue, destained, and then scanned.
Transfection
To stably overexpress and silence Zeb1, cells were transfected with a pcDNA3.1 vector encoding Zeb1, and with a Zeb1-targeted siRNA, respectively (Shanghai GeneChem Company, Shanghai, China). An empty pcDNA3.1 vector and a non-targeted siRNA were transfected as respective controls, and cells were selected with puromycin (1.5 μg/mL). Lipofectamine 2000 was used for cell transfection according to the manufacturer's protocols. We extracted protein and total RNA at 24 h after transfection.
Real-time PCR
Total RNA was extracted after the indicated treatments (24 h) using TRIzol according to the manufacturer’s protocol. Cells or tissues were lysed by 0.2 mL chloroform and centrifuged (12,000 × g at 4°C for 15 min). The supernatant was then treated with 0.5mL isopropanol and centrifuged (12,000 × g at 4°C for 10 min). The RNA pellet was dissolved in 1 mL 75% ethanol, centrifuged (7,500 × g at 4°C for 5 min), and the supernatant discarded. After resuspension in DEPC water, 1 µg of RNA was reverse transcribed to cDNA using a RT-PCR kit. Real-time PCR was performed using an Mx 3000P real-time PCR system (Applied Biosystems, Shanghai, China) and SYBR Premix Ex Taq as a DNA-specific fluorescent dye. PCR was carried out for 40 cycles of 95°C for 10 s and 60°C for 30 s. Primer sequences for detection of mRNA expression were synthesized (Table 2). All the reactions were repeated at least three times. Gene expression levels were calculated relative to the housekeeping gene GAPDH using Stratagene Mx 3000P software.
Table 2. Primers for RT-PCR.
| Name | Forward primer (5'->3') | Reverse primer (5'->3') |
| Zeb1 (NM_001323643.1) | GCACAACCAAGTGCAGAAG | CATTTGCAGATTGAGGCTG |
| Cyclin D1 (NM_053056.2) | CCGAGGAGCTGCTGCAAATGGAGCT | TGAAATCGTGCGGGGTCATTGCGGC |
| MMP2 (NM_004530.5) | CGCATCTGGGGCTTTAAACAT | TCAGCACAAACAGGTTGCAG |
| GAPDH (NM_002046.6) | GAAGGCTGGGGCTCATTTG | AGGGGCCATCCACAGTCTTC |
Western blot
Tissues (homogenized by grinding) and treated cells were lysed with lysis solution at 4°C for 30 min, followed by centrifugation (12,000 × g at 4°C for 15 min). From each sample, 20 µg of protein was fractionated by 10% sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto polyvinylidene difluoride (PVDF) membranes (Amersham, Beijing, China). After blocking with 5% nonfat dry milk in TBST for 1 h at room temperature, proteins were probed with specific antibodies against Zeb1, Cyclin D1, or MMP2. To assure equal loading, gels were stripped and reprobed with an anti-GAPDH antibody. Following incubation with HRP-conjugated secondary antibodies, signals were detected by chemiluminescence. All the reactions were repeated at least three times.
In vivo experiments
Five- to six-week-old female, athymic nude BALB/c mice (Vital River Laboratory Animal Technology Co. Ltd., Shanghai, China) were split into three groups of six and received tail vein injections containing 2×106 MG63 cells in 0.1 ml saline. The following day, and once a day thereafter, the mice were given intravenous injections of naringin (5 or 10 mg/kg) or 0.9% NaCl (control). On day 16 following tumor cell injection, liver samples were collected for histological examination.
All experimental procedures involving animals were conducted in accordance with the Guide for the Care and Use of Laboratory Animals (NIH publication no. 80-23, revised 1996) and followed institutional ethical guidelines. The study was approved by the Ethics Committee at the Academic Medical Center of Shengjing Hospital of China Medical University.
Histopathology
Lung specimens were fixed with 4% paraformaldehyde. Serial sections were cut using a microtome and affixed onto positively charged slides. Tissues were deparaffinized and rehydrated through graded xylene and alcohol. The sections were lightly counterstained with hematoxylin–eosin, dehydrated through an ethanol series, cleared in xylene and mounted. Stained sections were viewed using a light microscope (400×).
Statistical analysis
All data are presented as the mean ± SD. Statistical significance between two groups of data was evaluated by Student's t test (two-tailed) using GraphPad Prism software (GraphPad, Inc., La Jolla, CA, USA). P < 0.05 was considered significant.
Ethics statement and consent to participate
Research involving human subjects, human material, or human data has been performed in accordance with the Declaration of Helsinki and was approved by the Research Ethics Committee of Shengjing Hospital (R20160965).
Compliance with ethical standards
For the use of clinical materials for research purposes, written consent and approval from patients were obtained from the Shengjing Hospital of China Medical University. Patient consent was obtained in writing according to institutional regulations.
Consent to publish
We have obtained consent to publish from the participants to report individual patient data.
Author Contributions
Ming He: conceived of the study and carried out molecular studies. Qiu Chuang: carried out molecular studies. Wang Jiashi: participated in the design of the study and performed statistical analysis. Li Bin: participated in the study design and coordination and helped to draft the manuscript. Wang Guangbin: conceived the study. Ji Xianglu: helped to draft the manuscript.
Acknowledgments
All personnel who have contributed to this article are in the list of authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- 1. Degnan AJ, Chung CY, Shah AJ. Quantitative diffusion-weighted magnetic resonance imaging assessment of chemotherapy treatment response of pediatric osteosarcoma and Ewing sarcoma malignant bone tumors. Clin Imaging. 2018; 47:9–13. https://doi.org/10.1016/j.clinimag.2017.08.003 [PubMed]
- 2. Shi J, Fu Q, Yang P, Liu H, Ji L, Wang K. Downregulation of microRNA-15a-3p is correlated with clinical outcome and negatively regulates cancer proliferation and migration in human osteosarcoma. J Cell Biochem. 2018; 119:1215–22. https://doi.org/10.1002/jcb.26294 [PubMed]
- 3. Davis JC, Daw NC, Navid F, Billups CA, Wu J, Bahrami A, Jenkins JJ, Snyder SE, Reddick WE, Santana VM, McCarville MB, Guo J, Shulkin BL. (18)F-FDG uptake during early adjuvant chemotherapy predicts histologic response in pediatric and young adult patients with osteosarcoma. J Nucl Med. 2018; 59:25–30. https://doi.org/10.2967/jnumed.117.190595 [PubMed]
- 4. Raha S, Yumnam S, Hong GE, Lee HJ, Saralamma VV, Park HS, Heo JD, Lee SJ, Kim EH, Kim JA, Kim GS. Naringin induces autophagy-mediated growth inhibition by downregulating the PI3K/Akt/mTOR cascade via activation of MAPK pathways in AGS cancer cells. Int J Oncol. 2015; 47:1061–69. https://doi.org/10.3892/ijo.2015.3095 [PubMed]
- 5. Yin FM, Xiao LB, Zhang Y. [Research progress on Drynaria fortunei naringin on inflammation and bone activity]. Zhongguo Gu Shang. 2015; 28:182–86. [PubMed]
- 6. Chen R, Qi QL, Wang MT, Li QY. Therapeutic potential of naringin: an overview. Pharm Biol. 2016; 54:3203–10. https://doi.org/10.1080/13880209.2016.1216131 [PubMed]
- 7. Wang D, Ma W, Wang F, Dong J, Wang D, Sun B, Wang B. Stimulation of Wnt/β-Catenin Signaling to Improve Bone Development by Naringin via Interacting with AMPK and Akt. Cell Physiol Biochem. 2015; 36:1563–76. https://doi.org/10.1159/000430319 [PubMed]
- 8. Ang ES, Yang X, Chen H, Liu Q, Zheng MH, Xu J. Naringin abrogates osteoclastogenesis and bone resorption via the inhibition of RANKL-induced NF-κB and ERK activation. FEBS Lett. 2011; 585:2755–62. https://doi.org/10.1016/j.febslet.2011.07.046 [PubMed]
- 9. Zeng L, Zhen Y, Chen Y, Zou L, Zhang Y, Hu F, Feng J, Shen J, Wei B. Naringin inhibits growth and induces apoptosis by a mechanism dependent on reduced activation of NF‑κB/COX‑2‑caspase-1 pathway in HeLa cervical cancer cells. Int J Oncol. 2014; 45:1929–36. https://doi.org/10.3892/ijo.2014.2617 [PubMed]
- 10. Kawaguchi K, Kikuchi S, Hasegawa H, Maruyama H, Morita H, Kumazawa Y. Suppression of lipopolysaccharide-induced tumor necrosis factor-release and liver injury in mice by naringin. Eur J Pharmacol. 1999; 368:245–50. https://doi.org/10.1016/S0014-2999(98)00867-X [PubMed]
- 11. Guo B, Zhang Y, Hui Q, Wang H, Tao K. Naringin suppresses the metabolism of A375 cells by inhibiting the phosphorylation of c-Src. Tumour Biol. 2016; 37:3841–50. https://doi.org/10.1007/s13277-015-4235-z [PubMed]
- 12. Cai L, Wu H, Tu C, Wen X, Zhou B. Naringin inhibits ovarian tumor growth by promoting apoptosis: An in vivo study. Oncol Lett. 2018; 16:59–64. [PubMed]
- 13. Aigner K, Dampier B, Descovich L, Mikula M, Sultan A, Schreiber M, Mikulits W, Brabletz T, Strand D, Obrist P, Sommergruber W, Schweifer N, Wernitznig A, et al. The transcription factor ZEB1 (deltaEF1) promotes tumour cell dedifferentiation by repressing master regulators of epithelial polarity. Oncogene. 2007; 26:6979–88. https://doi.org/10.1038/sj.onc.1210508 [PubMed]
- 14. Browne G, Sayan AE, Tulchinsky E. ZEB proteins link cell motility with cell cycle control and cell survival in cancer. Cell Cycle. 2010; 9:886–91. https://doi.org/10.4161/cc.9.5.10839 [PubMed]
- 15. Caramel J, Ligier M, Puisieux A. Pleiotropic Roles for ZEB1 in Cancer. Cancer Res. 2018; 78:30–35. https://doi.org/10.1158/0008-5472.CAN-17-2476 [PubMed]
- 16. Wang M, He SF, Liu LL, Sun XX, Yang F, Ge Q, Wong WK, Meng JY. Potential role of ZEB1 as a DNA repair regulator in colorectal cancer cells revealed by cancer-associated promoter profiling. Oncol Rep. 2017; 38:1941–48. https://doi.org/10.3892/or.2017.5888 [PubMed]
- 17. Hanrahan K, O’Neill A, Prencipe M, Bugler J, Murphy L, Fabre A, Puhr M, Culig Z, Murphy K, Watson RW. The role of epithelial-mesenchymal transition drivers ZEB1 and ZEB2 in mediating docetaxel-resistant prostate cancer. Mol Oncol. 2017; 11:251–65. https://doi.org/10.1002/1878-0261.12030 [PubMed]
- 18. Bae GY, Choi SJ, Lee JS, Jo J, Lee J, Kim J, Cha HJ. Loss of E-cadherin activates EGFR-MEK/ERK signaling, which promotes invasion via the ZEB1/MMP2 axis in non-small cell lung cancer. Oncotarget. 2013; 4:2512–22. https://doi.org/10.18632/oncotarget.1463 [PubMed]
- 19. Shan Y, Zhang L, Bao Y, Li B, He C, Gao M, Feng X, Xu W, Zhang X, Wang S. Epithelial-mesenchymal transition, a novel target of sulforaphane via COX-2/MMP2, 9/Snail, ZEB1 and miR-200c/ZEB1 pathways in human bladder cancer cells. J Nutr Biochem. 2013; 24:1062–69. https://doi.org/10.1016/j.jnutbio.2012.08.004 [PubMed]
- 20. Wang H, Xing D, Ren D, Feng W, Chen Y, Zhao Z, Xiao Z, Peng Z. MicroRNA‑643 regulates the expression of ZEB1 and inhibits tumorigenesis in osteosarcoma. Mol Med Rep. 2017; 16:5157–64. https://doi.org/10.3892/mmr.2017.7273 [PubMed]
- 21. Yi L, Liu M, Tang Z. MicroRNA‑130a inhibits growth and metastasis of osteosarcoma cells by directly targeting ZEB1. Mol Med Rep. 2017; 16:3606–12. https://doi.org/10.3892/mmr.2017.6968 [PubMed]
- 22. Li H, Yang B, Huang J, Xiang T, Yin X, Wan J, Luo F, Zhang L, Li H, Ren G. Naringin inhibits growth potential of human triple-negative breast cancer cells by targeting β-catenin signaling pathway. Toxicol Lett. 2013; 220:219–28. https://doi.org/10.1016/j.toxlet.2013.05.006 [PubMed]
- 23. Wang H, Li C, Li J, Zhu Y, Jia Y, Zhang Y, Zhang X, Li W, Cui L, Li W, Liu Y. Naringin enhances osteogenic differentiation through the activation of ERK signaling in human bone marrow mesenchymal stem cells. Iran J Basic Med Sci. 2017; 20:408–14. [PubMed]
- 24. Wong RW, Rabie AB. Effect of naringin on bone cells. J Orthop Res. 2006; 24:2045–50. https://doi.org/10.1002/jor.20279 [PubMed]
- 25. Zhang X, Zhang Z, Zhang Q, Zhang Q, Sun P, Xiang R, Ren G, Yang S. ZEB1 confers chemotherapeutic resistance to breast cancer by activating ATM. Cell Death Dis. 2018; 9:57. https://doi.org/10.1038/s41419-017-0087-3 [PubMed]
- 26. Yu P, Shen X, Yang W, Zhang Y, Liu C, Huang T. ZEB1 stimulates breast cancer growth by up-regulating hTERT expression. Biochem Biophys Res Commun. 2018; 495:2505–11. https://doi.org/10.1016/j.bbrc.2017.12.139 [PubMed]
- 27. Langer EM, Kendsersky ND, Daniel CJ, Kuziel GM, Pelz C, Murphy KM, Capecchi MR, Sears RC. ZEB1-repressed microRNAs inhibit autocrine signaling that promotes vascular mimicry of breast cancer cells. Oncogene. 2018; 37:1005–19. https://doi.org/10.1038/onc.2017.356 [PubMed]
- 28. Ramesh E, Alshatwi AA. Naringin induces death receptor and mitochondria-mediated apoptosis in human cervical cancer (SiHa) cells. Food Chem Toxicol. 2013; 51:97–105. https://doi.org/10.1016/j.fct.2012.07.033 [PubMed]
- 29. Liu Y, El-Naggar S, Darling DS, Higashi Y, Dean DC. Zeb1 links epithelial-mesenchymal transition and cellular senescence. Development. 2008; 135:579–88. https://doi.org/10.1242/dev.007047 [PubMed]