



Introduction
Stem cells, which are characterized by their ability to self-renew, present in most adult tissues produce daughter stem cells and differentiated cells. With these unique properties, stem cells replenish aged cells to maintain tissue homeostasis throughout lifespan. With age, most stem cells lose their self-renewing ability, leading to loss of stem cell number, which in turn results in tissue aging and deterioration [1–5]. Importantly, majority of stem cells reside in a specific microenvironment, the stem cell niche, which provides factors responsible for maintenance and differentiation of stem cells in a controlled and coordinated manner. These include local signals that ensure stem cell self-renewal and adhesion of stem cells to their niche compartment. Many evidences suggest that as niches age, their function deteriorates, causing age-related defects in the associated stem cell populations [2,6–8]. Molecular mechanisms underlying niche aging are still poorly understood.
The Drosophila testis has been used as an incisive genetic model to provide insights into the mechanisms underlying aging processes occurring in the male germline stem cells and their niches [9,10]. Hub cells, a rosette of 10-12 post-mitotic cells localized at the anterior end of the testis, are the main components of the niche and actively support germline stem cells (GSCs) self-renewal [11]. To this end, hub cells produce DE-cadherin, which mediates adhesion of stem cells to the niche, and secrete self-renewal signaling molecules that are required for stemness, such as unpaired (Upd) [11]. With age, Drosophila testis becomes slender, hub cell number decreases, and GSCs lose their ability to divide [1,6]. Both DE-cadherin and Upd are reduced in aging hub cells, which underlies stem cell loss from the niche [6]. In addition, the expression of IGF-II messenger RNA binding protein (IMP), which stabilizes upd mRNA, also declines with age in the Drosophila testis [12]. This age-related decline of IMP in the hub cells is caused by the gradual induction of the microRNA Let-7 in aging hub cells, which targets Imp RNA for degradation and results in upd reduction [12,13].
Lin28 is a conserved RNA-binding protein in higher eukaryotes with function in development, metabolism, differentiation and pluripotency. The best characterized function of Lin28 protein is to act as an inhibitor of the biogenesis of let-7 microRNAs to reduce mature Let-7 [14–16]. Alternatively, Lin28 acts as a regulator of mRNA stability and translation, by potentially binding thousands of mRNAs [17–26]. Using either one of these properties, Lin28 regulates diverse physiological processes [22,27]. Lin28, for example, functions as a heterochronic factor that regulates developmental timing in C. elegans [28]. Lin28 regulates early stage of development in C. elegans and Lin28 level is reduced as developmental process progresses [29,30]. In mammals, Lin28 plays a role in cell fate succession, specifying early cell fate, which is analogous to the heterochronic function originally revealed in C. elegans [20]. Although much progress has been made on the role of Lin28, particularly in developmental processes as an early cell fate regulator, little is known about the role of Lin28 in the aging process of tissues maintained by a resident stem cell population. Here we show that Drosophila Lin28 is specifically expressed in the testis stem cell niche and that its expression dramatically declines with age. Our results reveal a let-7 independent role of Lin28 in hub cells: Lin28 can directly bind and protect the upd mRNA. We finally show that maintaining Lin28 expression in old hub cells prevent the age-related decrease in Upd levels and decline in the niche function, strongly supporting the notion that decline in Lin28 protein in the old niche significantly contributes to the aging process of the Drosophila testis.
Results
Lin28 is expressed in hub cells
At the anterior tip of the Drosophila testis, hub cells assemble to constitute the niche that supports two stem cell populations, germline stem cells (GSCs) and somatic cyst stem cells (CySCs), with each GSC surrounded by two CySCs (Fig. 1A) [31,32]. To examine the expression of Lin28 in the testis, we generated an antibody directed against the Lin28 protein. We first confirmed the specificity of our antibody using two Lin28 mutant alleles: the insertion line lin28EP915 (lin28EP), with a P-element insertion in the 4th exon, and the deletion line lin28df30, which deletes several coding exons [27] (Fig. S1A). Western blot analysis showed that Lin28 protein, which is detectable in wild-type testis and embryo extracts, is not detectable in those of both lin28EP and lin28df30 homozygous animals (Fig. 1B), showing that Lin28 is expressed in the fly testis and these two lin28 mutant alleles are null with regard to protein level.
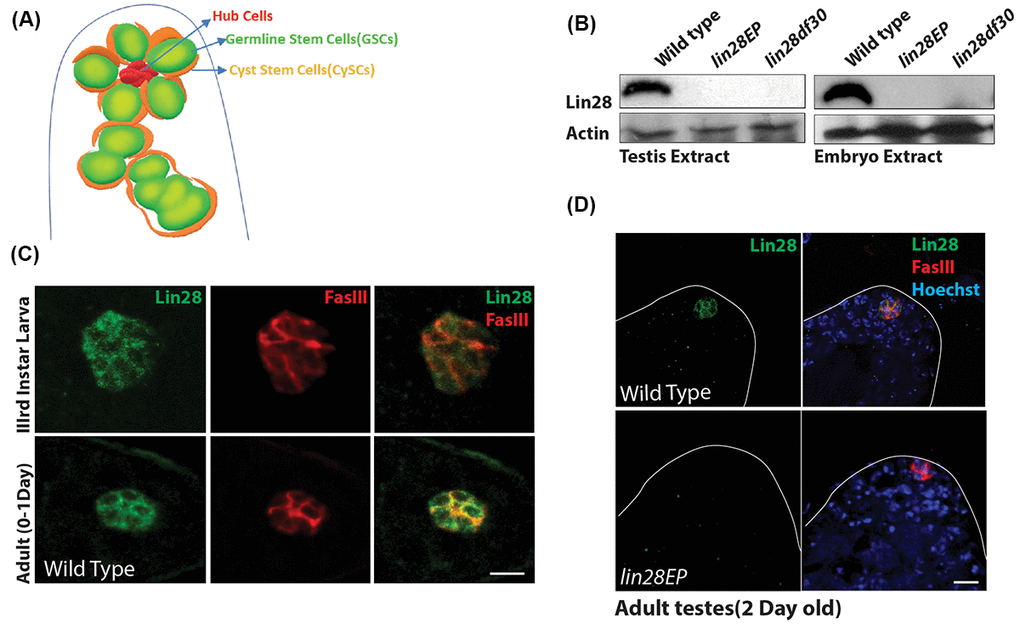
Figure 1. Expression of Lin28 in Drosophila testis. (A) A cartoon depicting the Drosophila testis tip, showing the location of hub cells, germline stem cells (GSCs) and cyst stem cells (CySCs). (B) Western blot showing levels of Lin28 in testis (left) and embryo extracts (right). Lin28 protein is not detectable in lin28 mutants. (C) Comparison of expression of Lin28 in 3rd larva (up) and young 0-1 Day old adult testis (down). Lin28 is specific to the hub cells. Testis were stained with antibodies specific to Lin28 (green) and hub cell marker FasIII (red). Scale 10µm. (D) Adult testis stained with antibodies specific to Lin28 (green), FasIII (red) and Hoechst (blue) in wild-type (top) and lin28 mutant (bottom), notice Lin28 expression is abolished in the lin28 mutant.
We next performed co-staining of the testis with an antibody against Fascillin-III protein (FasIII), a hub cell marker. We found that Lin28 is specifically expressed in the hub cells of the stem cell niche in the 3rd instar larva and young adult testis, while it is not expressed in other somatic and germline cells in this organ (Fig 1C, D; Fig. S1C). Importantly, no signal was detected in the hub cells in both lin28EP and lin28df30 homozygotes (Fig. 1D and Fig. S1B), confirming the specificity of the detected signal.
Hub cell defects in lin28 mutants
Since Lin28 is expressed in the hub cells we looked for any phenotype in lin28 mutants. To this end, we took advantage of the two lin28EP and lin28df30 alleles, which our western blot and immuno-staining confirmed as null or severely hypomorphic (Fig. 1B, D). Homozygotes for both alleles are viable, normal in size and show no observable morphological defects. While the fertility of the young mutant males is not affected (Fig. S1), we found that lin28 mutants present several defects in the hub. Young lin28 mutant have a significantly reduced number of hub cells with altered morphology (Fig. 2A-C). A typical young (0-1day old) Drosophila testis niche consists of approximately 10-12 somatic hub cells arranged in a dome like structure at the anterior tip of the testis [32]. Young lin28 mutant testis showed reduced hub cells number of 6.53 ± 0.2D8 (n=68) compared to 11.1 ± 0.20 (n=52) in young wild-type controls (Fig. 2B, C and Table S1). In addition to reduced number of hub cells, the morphology of the hub is largely aberrant in lin28 mutants, as shown by the irregular distribution of FasIII protein, which tends to accumulate at the periphery of mutant hubs, and reduced DE-cadherin levels (Fig. 2A, E, S2A-C), along with increased cell size in lin28 mutants (Fig. 2F). It is well established that hub cells are essential to maintain GSCs and CySCs population [1,6,9]. Compatible with reduced hub cell number and/or activity, lin28 mutants show a small but significant decrease in GSCs, identified as Vasa-positive cells immediately adjacent to hub cells, and Cyst Cells, identified by Traffic Jam (TJ) expression excluding the hub cell specific signal (Fig. 2B, D and Table S1).
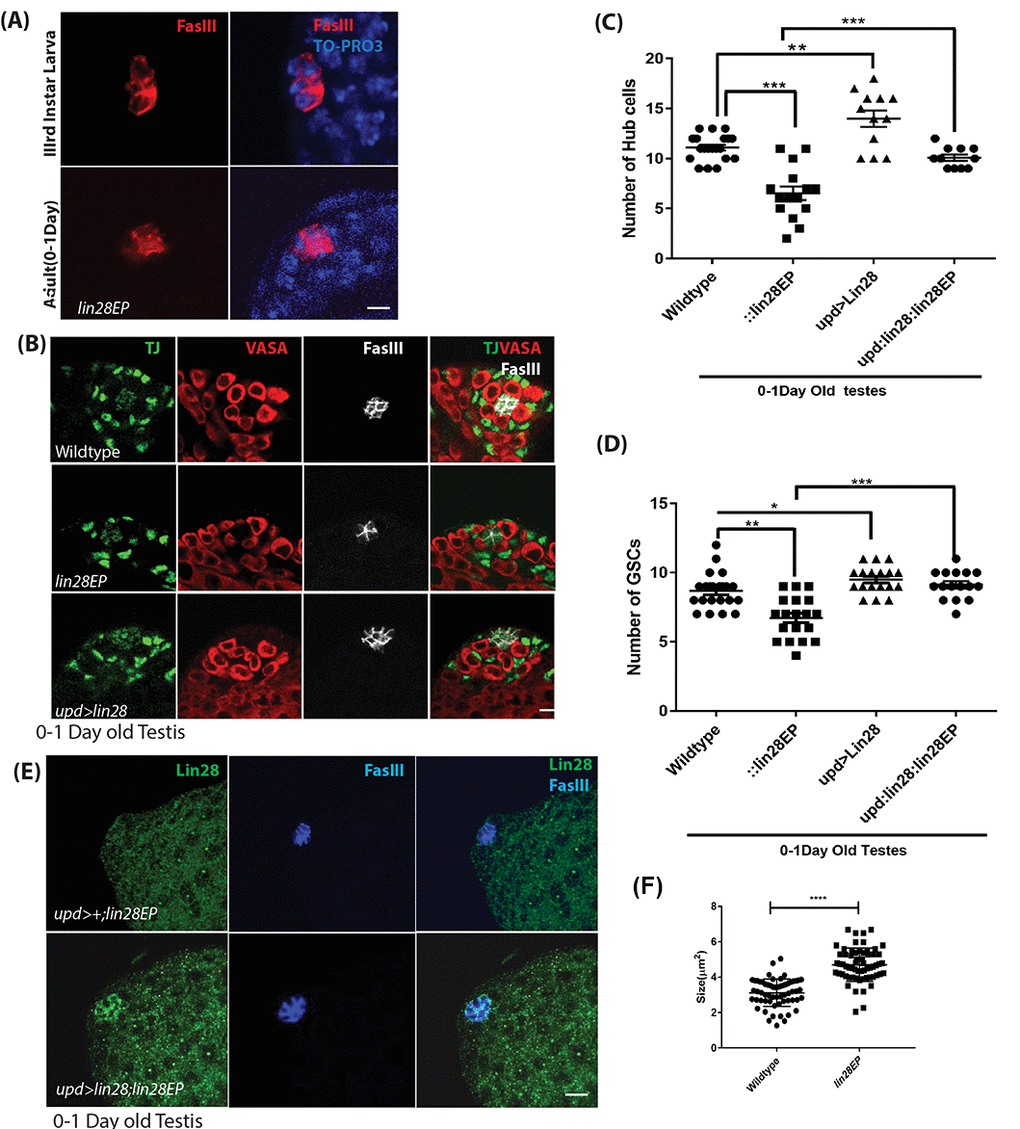
Figure 2. Phenotypes of lin28 mutants and overexpression of Lin28. (A) Comparison of the hub cells in 3rd larva (top) and young adult testis (bottom) stained with FasIII (red) and TO-PRO3 (Blue) showing defect in hub cell morphology in lin28 mutant. Scale 10µm. (B) Young adult testis stained with Traffic Jam (TJ), Vasa and FasIII. The number of hub cells is decreased in lin28 mutant testis. Hub cells are increased in Lin28 overexpressed testis. Scale 10µm. (C) Graph depicting the number of hub cells in 0-1Day old wildtype, lin28 mutant and overexpression of Lin28 along with the rescue of mutant testes by hub specific overexpression of Lin28. ** denotes P<0.01, *** denotes P <0.001 (from Table S1). (D) Number of GSCs in 0-1-day old wildtype, lin28 mutant and overexpressed Lin28 testis, along with rescue of mutant testes by hub specific expression of Lin28. *, **, *** denotes P<0.05, P <0.01, P<0.001 (from Table S1). (E) Adult rescued testis showing the expression of Lin28 and FasIII. Lin28 was over expressed in lin28 mutant background (upd-GAL4/+: UAS-lin28/+: lin28Ep915/lin28Ep915), which could rescue the mutant phenotype of lin28 mutant testes (see Table S1). (F) The significant increase in the cell size in lin28 mutant, compared to hub cells in wildtype testes. Scale bar 10µm.
Importantly, morphological defects and reduced number of hub cells and GSCs were rescued by directing Lin28 expression specifically in hub cells in lin28 mutants, using the Gal4-UAS system and the hub cell-specific driver upd-GAL4 (upd-GAL4: UAS-lin28: lin28EPEP915) (Fig. 2E, Table S1). Altogether, our data demonstrate that the function of Lin28 is required cell-autonomously in hub cells for their proper number and development.
Lin28 expression decreases with age and maintaining Lin28 expression prevents niche aging
Remarkably, most of the phenotypes observed in lin28 mutant testes, such as decreased cell number and DE-Cadherin levels, recapitulate changes observed during normal aging [1,6]. Thus, we asked whether Lin28 expression varies with ages and whether such expression changes could contribute to niche aging.
We found that Lin28 protein levels dramatically decreased in aging animals (Fig. 3A, B), becoming almost undetectable in 50-day-old testes. In addition, at a transcript level Lin28 expression is reduced significantly in the old testes (Fig. 3C). Next, we reasoned lin28 mutation may exacerbate testis aging phenotype and maintaining Lin28 expression in the hub cells of aging animals may counteract testis aging. To test this hypothesis, we quantified the number of hub cells in old (50-day-old) control males, lin28 mutants and animals with lin28 over-expressed specifically in Hub cells (upd-GAL4>UAS-Lin28) (Fig. 3D, E and Table S1). In wild-type males, 7.6 ± 0.33 (n=40) hub cells were observed while 3 ± 0.30 (n=60) hub cells were observed in lin28 mutant (Fig. 3 D, E). 5% of the testis showed complete loss of hub cells in the mutant old testis. This confirmed the previously reported decrease in hub cell number during normal aging [1,6] and suggested that aging is significantly accelerated in lin28 mutants. Conversely, hub cell specific over-expression of Lin28 significantly increased the number of hub cells from 11.1± 0.20 to 13.6 ± 0.26 (n=42) in young testis and from 7.6 ± 0.33 to 10.8 ± 0.36 (n=38) in 50-day old testis (Fig. 3D, E and Table S1), suggesting that ectopic Lin28 can enhance and maintain stem cell niche function in old testes. As for young testes, changes in hub cell numbers in old animals were mirrored by similar changes in the number of GSCs and apical somatic cyst cells excluding the hub cells (Fig. 3D, F and Table S1).
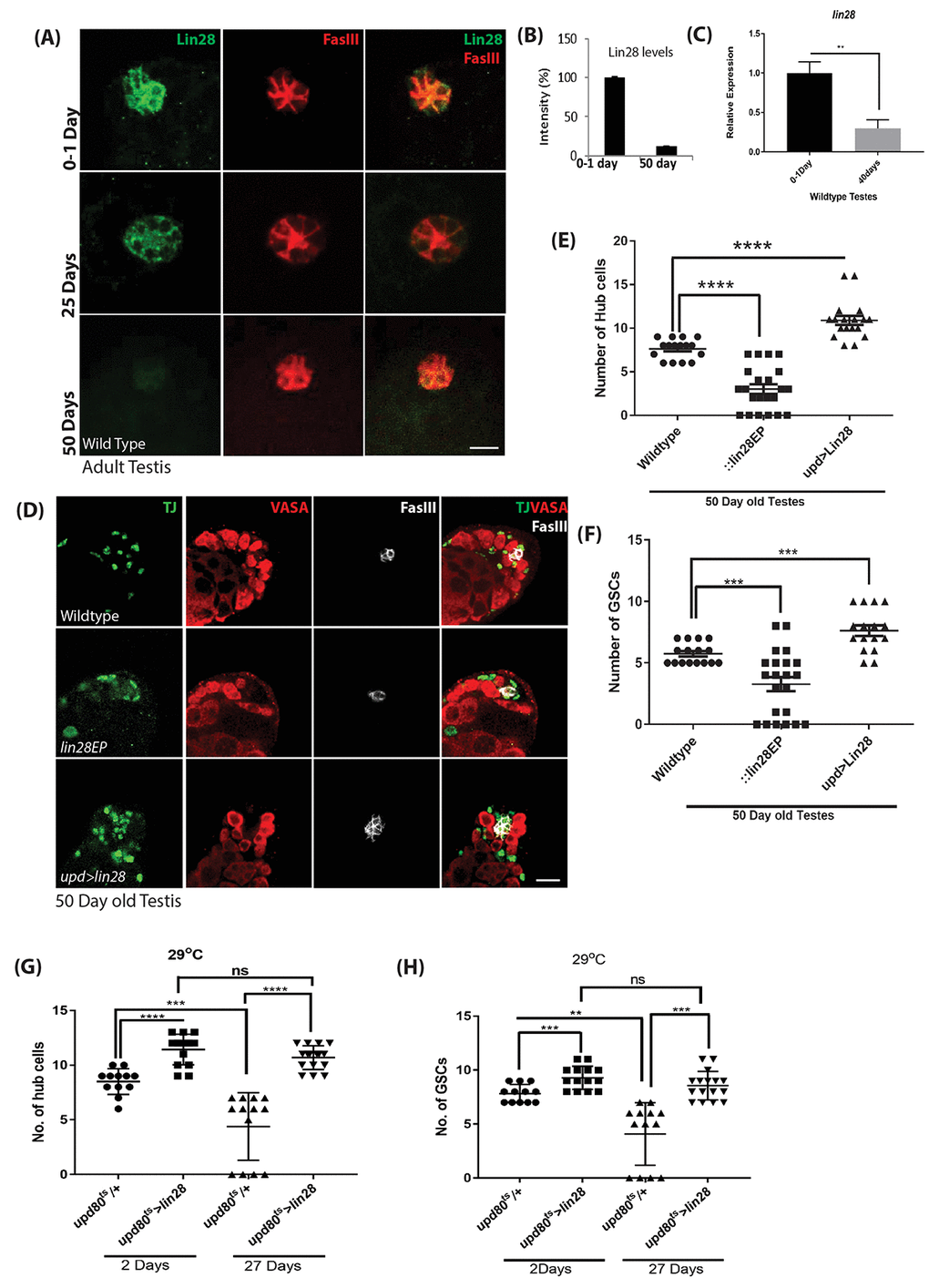
Figure 3. Age dependent loss of Lin28 in hub cells. (A) Adult testis stained with Lin28 (green) and FasIII (red) in an age dependent manner. Lin28 level, but not FasIII, is decreased in old testis. Scale 10µm. (B) Comparison of the level of intensity of green signal (Lin28 expression) in young (0-1 Day) vs old testis (50 Day). (C) Comparison for the transcript levels of lin28 in young (0-1 Day) and old (40 Days) wildtype testis, showing the reduction in Lin28 levels with age. (D) Hub cell number is not decreased in old testis when Lin28 is overexpressed (upd-Gal4>UAS-lin28) as compared to wild type testis. Testis were stained with Vasa, Traffic Jam (TJ) and FasIII antibodies. Scale 10µm. (E) The number of hub cells in old testis is comparatively reduced in lin28 mutant testis compared to wildtype testis. The number of hub cells in old upd>lin28 (upd-Gal4>UAS-lin28) testis is similar to a young wild type testis (data from Table S1). (F) The number of GSCs in 50-day old wildtype, lin28 mutant and upd>lin28 (upd-Gal4>UAS-lin28) testis (data from Table S1). (G) Comparison of the number of hub cells in wildtype vs adult specific overexpression of Lin28 in an age dependent manner (upd-GAl4; tubGAL80ts: UAS-lin28). Overexpression of Lin28 clearly increases the number of hub cells and with age the number of hub cells is maintained when lin28 is maintained compared to loss of hub cell in wildtype testes with age. (H) Comparison of the number GSCs in wildtype testes and adult specific over expression of Lin28 in hub cells. The number of GSCs reciprocates the number of cell number. * denotes, P<0.05. ** denotes P<0.01, *** denotes P<0.001, and **** denotes 0.0001.
We show that absence of lin28 or over-expression in hub cells throughout development results in small but significant changes in hub cells and GSCs number in young animals, raising the possibility that the effects observed in aging tissues results from developmental defects. Thus, to further characterize the adult specific role of Lin28 in the hub cells and its connection to the aging process, we carried out adult-specific Lin28 expression. To this end, we combined the upd-GAL4 driver with the tubulin-GAL80ts temperature sensitive GAL-80 inhibitor (updG80ts) [33] and analyzed animals grown at permissive temperature until adult and switch to 29oC for 2 days (young) or 27 days (old). Our data show that Lin28 over-expression increases the number of hub cells 2 days after transgene induction, (11.43 ± 0.37, n=42) compared to wild type control (8.5 ± 0.33, n=36) (Fig. 3G, S3A, Table S2). More importantly, we found that, while a strong age-related loss of hub cells is observed in control testes, no significant age-dependent change in hub cell number is observed when Lin28 expression is maintained throughout adulthood specifically in hub cells (young (11.43 ± 0.37, n=42) to old testes (10.69 ± 0.26, n=48), Table S2). These changes or absence of changes in hub cell numbers were mimicked by the number of GSCs in the niche (Fig. 3H, S3, Table S2).
Together these data strongly suggest that the aging niche becomes less functional and abnormal because of decreased expression of Lin28 in hub cells and that maintaining Lin28 expression in these cells is a viable approach to prevent testis aging.
Lin28 regulates Upd expression and STAT signaling in the hub
Lin28 expression pattern is identical to the expression pattern of the key self-renewal factors Upd and hub cell adhesion protein DE-cadherin, namely that they all are highly expressed in hub cells and show decreased expression with age [6]. This raised the possibility that upd and DE-cadherin reduction with age might be due to reduction of Lin28 with age. Therefore, we examined whether the expression of upd and DE-cadherin is dependent on Lin28. In-situ hybridization of upd transcripts confirmed that upd level is reduced in the mutant hub area (Fig. 4A), conversely hub specific expression of Lin28 leads to increased upd transcripts in the hub area. Quantitative RT-PCR showed that upd (Fig. 4B) and DE-Cadherin (Fig. S2C) transcript levels are reduced in lin28 mutant testes relative to wild-type. Conversely, over-expressing Lin28 in hub cells using upd-GAL4 significantly increases upd mRNA level (Fig. 4C). More importantly, confirming the role of Lin28 in hub cell aging, we observed an age-related reduction in the mRNA levels of upd in wild-type testes, while continued expression of Lin28 in hub cells of old testes maintains the expression of upd similar to young testes (Fig. 4C).
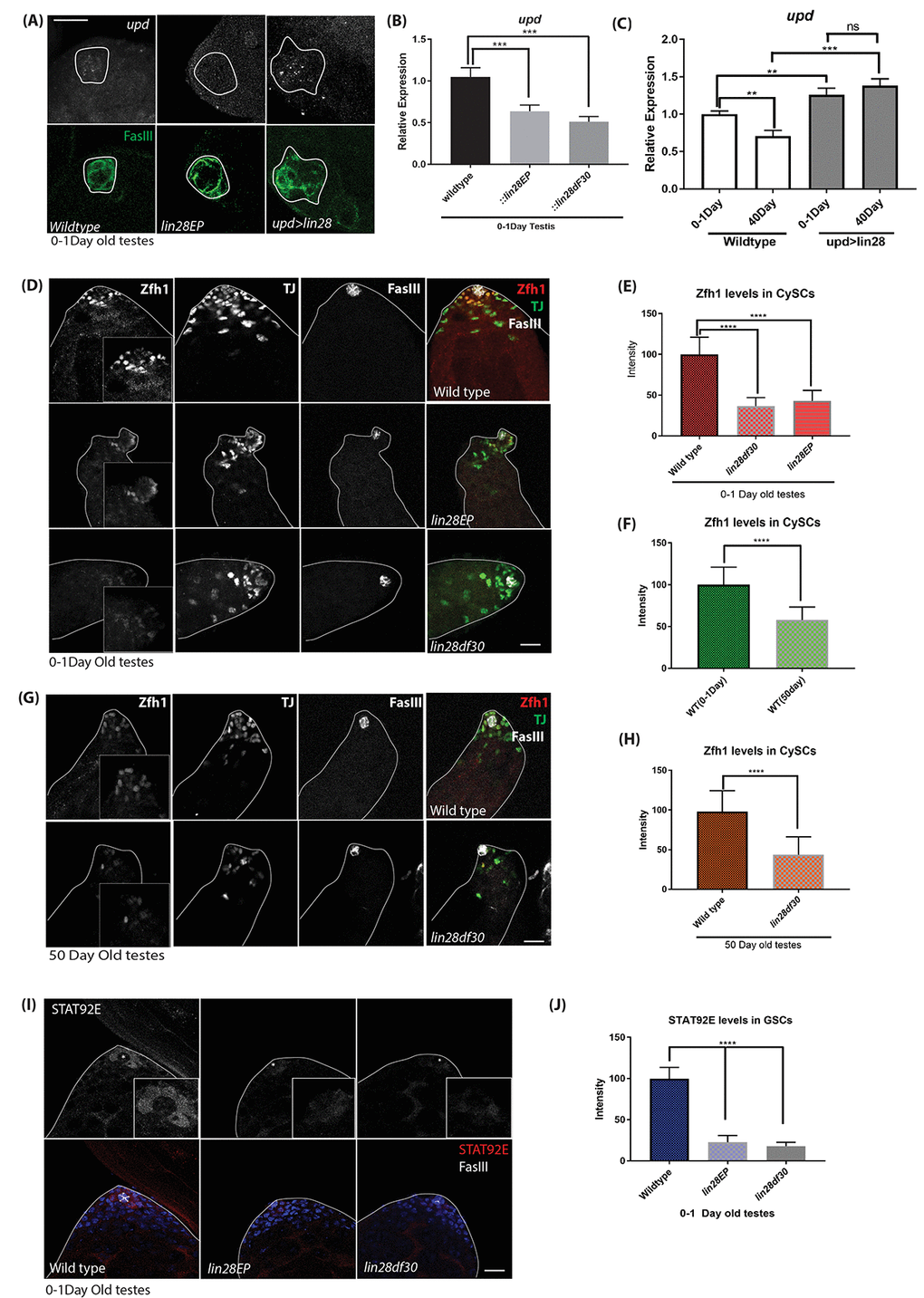
Figure 4. Upd-JAK-STAT signaling is affected with loss of Lin28. (A) In-situ hybridization of young (0-1 day old) testis. upd RNA level is decreased in lin28 mutant testis relative to wild-type and increase in upd levels in hub specific expression of Lin28. Scale 10µm. (B) quantitative RT-PCR of young (0-1day old) testis, upd mRNA level is decreased in lin28 mutants relative to wild-type. (C) RT-qPCR of young vs old testis showing reduced levels of upd mRNA in WT testis with age, while, maintained levels of upd in overexpression of Lin28 with age. (D) Young (0-1-day old) testis stained with Zfh1, TJ and FasIII antibodies. Zfh1 level, but not TJ and FasIII, is reduced in lin28 mutants. Scale 10µm. (E) Quantification of Zfh1 levels in CySCs shows reduced Zfh1 levels in mutants. [wildtype, n= 197; lin28EP, n=220; lin28df30, n= 185]. (F) Comparison of Zfh1 levels in young and old WT testes indicating that with age Zfh1 level goes down due to loss in STAT activity. [WT young, n= 197; WT old, n= 145]. (G) 50-day old testes stained with Zfh1, TJ and FasIII antibodies showing reduced levels of Zfh1 with age. Scale 10µm. (H) Comparison of Zfh1 levels in WT vs mutant [ wildtype, n= 145; lin28df30, n=125] (I) STAT92E protein level is reduced in the mutants. Scale 10µm. (J) Comparison of STAT92E levels in WT (n=115) vs mutants (lin28EP n=92; lin28df30, n=97). STAT92E protein level is reduced in lin28 mutants; * depicts the hub area. ** denotes P<0.01, *** denotes P<0.001, **** denotes P<0.0001, ns denotes not significant.
Secretion of upd in the hub cells activates the JAK-STAT signaling in the GSCs and CySCs, which is essential for their maintenance [11,34]. Zfh1, a transcriptional repressor is known as a somatic target of JAK-STAT signaling, expressed in CySCs [11,34]. Stabilized STAT92E protein can be used as a read-out for JAK-STAT signaling [35,36] Thus, we used these two markers to confirm that the Upd reduction in lin28 mutant hub cells affects STAT signaling in surrounding cells. Supporting our model, we found that Zfh1 protein level is reduced in CySCs of lin28 mutant testis (Fig. 4D, E) and that STAT92E protein is lost in the GSCs of lin28 mutants (Fig. 4I, J). In contrast, several factors that are expressed in hub cells such as FasIII and TJ protein levels were not altered in lin28 mutants (Fig. 4D). While with age the expression of Zfh1 goes down (Fig. 4F), we saw that the expression of Zfh1 further goes down in lin28 mutant testes (Fig. 4G, H).
Previously, it was shown that IMP stabilizes upd mRNA from siRNA mediated degradation in hub cells and with age, loss of IMP leads to loss of upd transcripts [12]. Age associated loss of IMP is related to post-transcriptional regulation of IMP transcripts by the microRNA let-7. Lin28 is a well-established inhibitor of biogenesis of let-7 microRNAs [37]. This raised the possibility that Lin28 controls upd expression through this let-7 / IMP axis. However, while we observed a slight reduction of IMP transcripts in young Lin28 mutant testes, we did not detect any reduction in expression of IMP protein in either young or old tissues (Fig. S4A, B, D). Similarly, over-expression of Lin28 in hub cells has limited effect on IMP expression in young or old testes, as measured by RT-qPCR or immunostaining (Fig S4A, C, D). Of note, an accumulation of IMP protein into granular structures, which co-localize with the stress granule marker Cup, in the hub area of the mutant testes can be observed in some samples (Fig. S4A, D, E), suggesting that IMP sub-cellular localization and function may be affected by the loss of Lin28. We next tested whether mature let-7 level is influenced by lin28 gene dose. We found that mature let-7 level is low in young animals and not significantly different between young wild-type and mutants (Fig. S4F). As reported previously [12], mature let-7 level dramatically increases with age in control tissues (Fig. S4F). However, this is not significantly different when Lin28 is mutated or over-expressed (Fig. S4F, G), suggesting that Lin28 does not regulate let-7 during the hub cell aging process.
Altogether, our results demonstrate that Lin28 controls upd expression in hub cells, in a largely let-7-independent manner.
Lin28 binds and stabilizes the upd transcript.
Lin28 is a translational regulator that was shown to bind several target mRNAs directly. Our in vivo experiments suggested to us that Lin28 may directly control the stability of the upd transcript. Thus, we examined the relationship between upd transcript level and Lin28 in S2 cells.
We confirmed that Lin28 over-expression in S2 cells increases the expression of a upd reporter which contain an intact 3’UTR, suggesting that the regulatory mechanisms present in hub cells are at least partially conserved in cultured cells (Fig. 5A). To first test whether Lin28 may bind the upd transcript directly, we mutated the putative binding site of Lin28 (GGAGA motif) present in upd3’UTR at position 228 and found that overexpression of Lin28 has no effect on the expression of this construct (Fig. 5A). We next carried out Lin28 immunoprecipitation followed by RT-PCR for upd mRNAs. Lin28 immunoprecipitates from S2 extracts are highly and specifically enriched in upd transcripts as compared to control reactions, demonstrating that Lin28 binds upd transcripts directly in these cells (Fig. 5B). The IMP protein has also been shown to bind upd mRNAs [12]. We confirmed that immunoprecipitating Flag-tagged IMP in S2 cells significantly enriches in upd mRNAs (Fig. 5C). We next asked whether these two binding events are dependent on each other. To this end we performed Lin28 immunoprecipitation while knocking-down IMP using siRNA and the converse experiment. Our results show that Lin28 and IMP proteins’ ability to bind to upd transcripts is not significantly affected in the absence of the other factor (Fig. 5B, C). Supporting the notion that both Lin28 and IMP can stabilize the upd transcript independent of each other, we found that overexpressing Lin28 while knocking down IMP is sufficient to increase upd levels and conversely IMP over-expression stabilize upd in the absence of Lin28 (Fig. 5D).
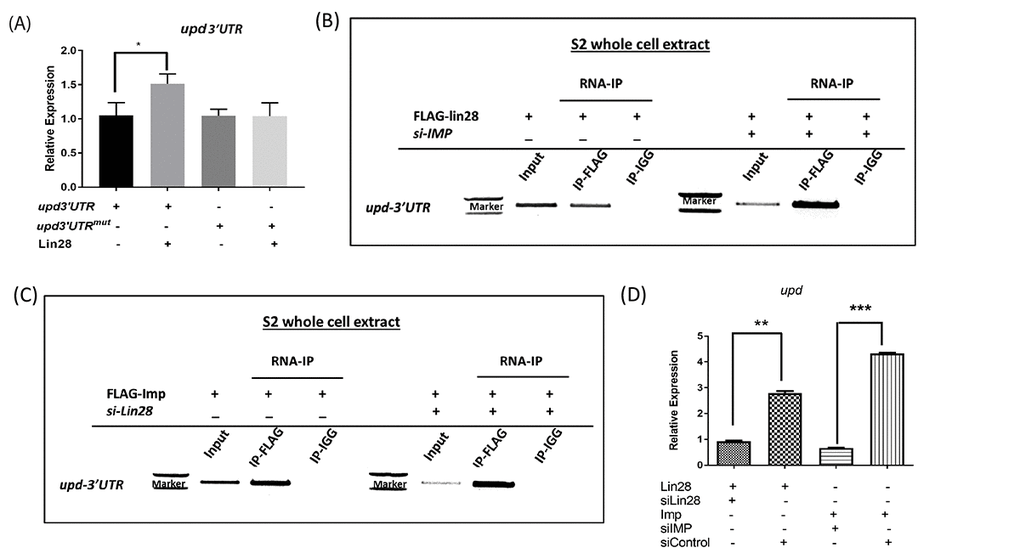
Figure 5. Lin28 and IMP associate with upd RNAs. (A) qRT-PCR showing the relative levels of Luc-Upd3’UTR reporters in cells co-transfected in the presence and absence of Lin28, showing increased Luc-Upd reporter expression in presence of lin28. Site directed mutagenesis of the Lin28 binding GGAGA motif abolishes the stabilization induced by Lin28 expression. (B and C) RNA-immunoprecipitation performed on S2 cells transfected with HA-tagged Lin28 and Flag-tagged IMP. Immunoprecipitation followed by RT-PCR detects that IMP and Lin28 immunoprecipitants contain upd RNAs. Depletion of either Lin28 or IMP does not affect RNA binding of the other protein. (D) Quantification of RNA-immunoprecipitation. siControl: scrambled siRNA. * denotes P<0.05, ** denotes P<0.01, *** denotes P<0.001.
Together, our biochemical studies demonstrate that Lin28 is capable of binding to and controlling the stability of the upd transcript, suggesting that it may control stem cell niche function in the testis at least in part through the regulation of Upd expression.
Discussion
Lin28 is required for niche development and long-term function
Lin28 has been known to function in diverse biological contexts like development, metabolism, pluripotency; along with a role in disease such as cancer [19,37]. In this report, our identification of Lin28 as one of the key factors regulating the male germline stem cell niche function extends our understanding of the diversity of roles Lin28 plays in stem cell biology. The study of the mechanism underlying the regulation of self-renewal factors in the hub cells provides us with new insights into the proper niche function. Our data establishes that Lin28, an RNA binding protein, plays a pivotal role in somatic post-mitotic hub cells of the Drosophila testis stem cell niche. We show that hub cell specific expression of Lin28 starts in early stages of testis development, plateaus in young adults and deteriorates with age. Similar to the niche deterioration observed in older testes, Lin28 mutations result in a loss in hub cell number, associated with impaired hub morphology due to aberrant increase in the cell size. Importantly, we show that hub cell specific expression of Lin28 is sufficient to rescue the mutant phenotype and that maintaining the expression of Lin28 in an adult specific manner is sufficient to preserve the number of hub cells in old testes, strongly suggesting a cell-autonomous function of Lin28 in regulation of long-term hub cell number, morphology and function.
While it is unclear how hub cells are lost and why their size increases in absence of Lin28, we attributed these phenotypes to a non-apoptotic mechanism: Tunnel assay and immunohistochemistry against cleaved caspase 3 (an apoptotic marker) did not show any significant change between control and lin28 mutant testes, suggesting the hub cell death or loss is not due to apoptosis (data not shown). Other mechanisms leading to hub cell loss are possible. For example, we show that E-cadherin accumulation around hub cells is strongly reduced in lin28 mutants, suggesting that the integrity of the niche is compromised, and hub cells may migrate away and lose their identity. Alternately, hub cells may trans-differentiate into CySCs when Lin28 is over-expressed similar to what was reported in [38], however, this possibility has been challenged by others [39]. Conversely, we show that over-expressing Lin28 throughout development or during adulthood results in a small but significant increase in hub cell number. Our studies have not allowed us to detect any dividing hub cells (positive for the phospho-Histone H3 mitotic marker) in these conditions (data not shown). However, previous studies have shown that germ cells and CySCs can repress hub cell development [39,40] the absence of these cues may trigger higher number of hub cells in Lin28 over-expressing conditions. Further experiments are required to understand whether Lin28 affects these mechanisms controlling specification and development of hub cells and CySCs and whether these signaling interactions between the cell types that compose the testis niche are maintained in the adult. We anticipate that the identification of Lin28 targets in hub cells may provide additional insights into the mechanism(s) by which Lin28 controls hub cell numbers.
Lin28 controls upd expression and regulate niche aging
Our data demonstrate that Lin28 controls the expression of self-renewal factor upd in hub cells and that its loss in the hub niche participates to the impaired Upd expression and niche function in older testes. Importantly, our data strongly suggest that Lin28 directly binds to the upd mRNA 3’UTR to control its stability in hub cells. In addition, we show that let-7 expression is largely not affected by Lin28 in testis, in particular in old animals where let-7 expression is highly induced and associated with the loss of Upd [12]. Recently, in Drosophila it was shown that Lin28 does not affect the biogenesis of let-7 [41] suggesting that the molecular mechanism for Lin28 function can vary. Our work thus identifies a novel mechanism that adds complexity to our previous view of the regulation of Upd expression during aging. In short, Upd is destabilized in old testis by two mechanisms acting in parallel: the increased let-7 expression which causes IMP loss; and the loss of Lin28 expression which directly affects the upd transcript. Why two parallel, although connected, pathways are necessary to maintain upd expression remains unclear at this point.
Our cell culture data suggest that both Lin28 and IMP can bind and stabilize the upd 3’UTR in the absence of the other factor. Our study indicates that Drosophila Lin28, like mammalian Lin28a [24], likely binds to mRNAs target including upd at the sequence specific motif (GGAGA). IMP binds to upd 3’UTR at a different sequence [12]. Whether single RNA molecule can be bound by both factors at the same time and whether this may further stabilize the target transcripts remains unexplored. It will also be interesting to investigate why both Lin28 and IMP can bind to upd mRNA independently and whether binding of Lin28 to upd induce any conformational changes which assists binding of IMP to upd mRNA or reciprocally. Interestingly in our hands, the absence of one of the 2 factors i.e. Lin28 or IMP, leads to an increase in upd mRNA level, suggesting that Lin28 and IMP can protect upd mRNA in absence of the other, and may have greater affinity for the upd 3’UTR in these conditions. This raises the possibility of Lin28 or IMP working in parallel pathways or at different stages of upd regulation. Whether this mechanism is mirrored in-vivo need to be further characterized. Functionally, it is clear from our studies and others that both factors are required to prevent accelerated aging in hub cells. It was observed that IMP protein accumulates in granular structures specific to the hub area of lin28 mutants. Preliminary investigation indicates that these foci are stress granules (Fig. S4E), whether this affects IMP function and upd stabilization, needs further characterization.
Other Lin28 targets in hub cells
Although the decreased number and morphological defects in Lin28 mutant hub cells may be attributed to loss of upd, it is likely that other factors are regulated by Lin28 and play a critical role in this phenotype and the overall decreased stem cell niche function. Other factors known to contribute to the niche function such as DE-cadherin, dpp, esg, Rbf are prime candidates that will have to be investigated. It is interesting to note that, in a different fly tissue Lin28 binds directly to the Insulin receptor InR mRNA in intestinal stem cells and regulate their symmetrical renewal [22]. InR function is required cell autonomously for GSC maintenance in Drosophila testis [42]; it will thus be interesting to test whether hub cell specific Lin28 has similar function. Further studies are required to understand how Lin28 might regulate other factors or signaling pathways present in the niche.
Age related changes in the stem cell niches have been known to directly influence stem cell function and self-renewal ability [6,8,9]. Age associated decline in stem cells function and niches is known to regulate aging. Understanding the mechanistic basis of stem cell behavior in the niche will lead us to the development of strategies to facilitate stem cell-based therapies. Future work involves broadening our understanding of the mechanism of stem cell renewal with Lin28 as a key regulator.
Materials and Methods
Fly stocks and husbandry
lin28 mutant line w[1118];P(w[+mC]=EP)lin-28[EP915] (#17298), were obtained from Bloomington Drosophila Stock Center. W1118 from Bloomington Drosophila Stock Center was used as a wildtype control. upd-GAL4 was a kind gift from S. Dinardo. lin28df30 was a kind gift from F. Michon. UAS-Lin28 was generated in the lab. upd-GAL4;tub-GAL80ts was a kind gift from Christian Bokel.
All updGal4; tubGAL80ts and updGal4;tub-GAL80ts>UAS-lin28 flies were raised at 18oC unless otherwise noted. Flies were raised at 18oC for 5 days post-eclosion to restrict GAL4 during final developmental stages. Flies were shifted to 29oC for 2 days to inhibit Gal80ts and activate GAL4 or aged for 27 days at 29oC.
Quantification of Hub cells and Germline Stem cells, and hub cell size
To quantify the number of hub cells, the testis was stained with antibody specific to hub cell, FasIII and TO-PRO3/Hoechst a nuclear marker. Densitometric analysis of Serial Z section was used to quantify the number of hub cells using LAS X software from Leica. The number of GSCs were quantified by the cells positively stained to VASA which were directly in contact to the FasIII positive hub cells. The size of the hub cells was measured as described [43].
Cloning
The lin28 cDNA (RE05342) was obtained from Drosophila Genetic Resource Centre. RE05342 consists of pFLC-1 vector which is a derivative of pBluescriptII-SK (+) with the cDNA insert (1165bp) of lin-28 (transcript 1152bp with 5’UTR and 3’UTR). The Not I-Xho I insert of the lin28 CDS (588bp) was sub-cloned into pAc5.1/V5- HisA-HA/FLAG (Invitrogen) vector for S2 cell expression, while Bgl II- Xho I insert and EcoR I-Xho I insert was sub-cloned into pUASt for fly expression studies. The KpnI-Not1 insert was sub-cloned into phmKGN/C-MC/MN vector for fragment complementation assay. The IGF-II mRNA binding protein (IMP) cDNA (RE72930) was obtained from Drosophila Genetic Resource Centre. RE72930 consists of derivative of pBluescriptII-SK (+) with cDNA insert (3955bp) of IMP-RJ (3942bp transcript with 5’UTR and 3’UTR). The Not I-Xho I insert of the IMP CDS (1743bp) was sub-cloned into pAc5.1/V5-HisA-FLAG for S2 cell related experiments.
The outstretched (os)/unpaired (upd) cDNA (BS-UPD) was obtained from Douglas A. Harrison. BS-UPD consists of the cDNA of upd (2520bp). The XhoI -Xba I insert of 3’UTR (740bp) of upd was sub-cloned into pAc5.1A(Invitrogen) following a luciferase reporter (pAc5.1A-Luc-upd3’UTR). Site directed mutagenesis was used to generate pAc5.1A-Luc- upd3’UTRmut (228GGAGA=AACAT).
Generation of Lin28 antibody
The N-Terminal peptide sequence Lin28- NGLERRTTSQSSTSSAN was used to generate the antibody against Lin28 in rat by Peptron (Korea).
Immunostaining
Immunostaining was done as described [44]. Briefly, for third instar larvae, the male larvae were hand dissected in Schneider’s media using micro scissors and micro forceps. Using the micro scissor, the larva was cut into half and the internal organs especially the gut was pulled out from the posterior region carefully, preventing the other organs from losing. This posterior region containing testis was fixed in 4% formaldehyde in PBS for 20minutes, followed by washing in PBX. It was then blocked in 5% Normal Goat serum for 1hour and incubated in appropriate antibody overnight at 4oC. Secondary antibodies were used at 1:500. After the secondary antibody treatment, the testis was carefully detached from the posterior region using tungsten needles and mounted in antifade. Adult testis of appropriately aged male flies were immunostained as previously described except they were hand dissected in Ringer’s solution and fixed in 4% formaldehyde for 40 minutes.
The following primary antibodies were used along with the following concentration: Rat anti-Lin28 1:100 (This study), mouse anti-FasIII 1:30, rat DE Cadherin 1:30, rat anti-VASA 1:400 (Developmental Studies Hybridoma Bank, DSHB), Rabbit anti-VASA 1:3000, Rabbit anti-Zfh1(1:1000) (Ruth. Lehmann), guinea pig anti-Traffic Jam 1:10000 (Dorothea Godt), rabbit anti-IMP 1:1000 (Paul MacDonald), Rat anti-Cup 1:1000 (A. Nakamura), Rabbit anti-GFP 1:500 (Invitrogen, USA), Rabbit anti- STAT92E 1:1000 (Denise Montell). The secondary antibodies were used in 1:500 (Alexa Series, Invitrogen, USA).
RNA-Immunoprecipitation
S2 cells transfected with HA tagged Lin28 and FLAG tagged IMP were washed twice in ice cold PBS and were subjected to RNA-IP. The washed cells were lysed using polysome lysis buffer containing protease inhibitor cocktail, RNase inhibitor and Vanadyl complexes along with DTT. The lysed cells were centrifuged at high speed for 15 minute and frozen for 30 minutes at -80oC. The thawed lysate was subjected to Immunoprecipitation using HA-Agarose or FLAG-Agarose beads for 4 hours at 4oC and washed 5 times in wash buffer followed by 3 times washing in wash buffer with 1M urea. The washed Agarose beads were subjected to DNAse1 treatment for 10 minutes at 37oC followed by proteinase K (10 mg/ml) treatment at 55oC for 30 minutes. The proteinase K treated beads were subjected to Phenol: Chloroform: Isoamyl alcohol extraction to extract RNA. The RNA was diluted and was subjected to reverse transcription using reverse transcriptase. The RT samples were used as template to amplify RNA bound to the flagged proteins by PCR.
When using the luciferase-based reporters, S2 cells were transfected with reporter constructs of Luc-upd3’UTR or Luc-upd3’UTRmut using DDAB transfection reagent (Fluka/Sigma). 72 hours after transfection cells were harvested, and total RNA was extracted with Trizol (Invitrogen). Following reverse transcription, qRT-PCR was used to measure Luc-upd3’UTR levels using upd3’UTR-specific primers with rp49 as control.
In-situ hybridization of upd RNA
RNA in situ hybridization was performed as previously described [11]. Full length cDNA of UPD cloned into plasmid was linearized to generate probes using the Roche RNA labeling kit (Roche, USA).
qRT-PCR
Total RNAs were extracted from young (0-1 day old) and old (30-32 days old or specified) testis using mirVana miRNA isolation kit (Invitrogen). Total RNA (200 ng) was used for cDNA synthesis with specific let-7 stem loop primer 5’-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACACTATA-3’ and let-7, forward, 5’-GCCGCTGAGGTAGTAGGTT GTA-3’ and reverse 5’-GTGCAGGGTCCGAGG-3’, as described previously [45]. Quantitative PCR was performed with SYBR green (Applied Biosystems, USA) on a Step One Real-Time PCR Systems from Thermo Scientific according to the manufacturer’s instructions. U6 snRNA (RT primer 5’-CGCTTCACGAATTTGCGTGTCAT-3’ and forward 5’-GCTTCGGCAGCACATATACTAAAAT-3’, reverse 5’-GTGCAGGGTCCGAGG-3’) was used as a reference RNA.
For qRT-PCR total RNA was extracted from the appropriately aged testes using RNeasy Mini Kit (Qiagen). Total RNA (500ng) was used for cDNA synthesis with oligo dT, Quantitative PCR was performed with SYBR green (Applied Biosystems, USA) on a Step One Real-Time PCR Systems from Bio-Rad according to the manufacturer’s instructions. Specific primers for qRT are the following: upd forward, 5’-GGGAGAGAGAGAGAAATAGAGAGA-3’ upd reverse, 5’-CGGGCGTGGCGAATAATA-3’; Imp forward, 5’-GCACCCACCACAATTTACAAC-3’, Imp reverse, 5’-CTCCAGGTTGCTTGCTTACT-3’; DE-Cadherin forward, 5’-CCGTAGACACTAAGACTCGATTAAG-3’, DE-cadherin, reverse, 5’-GTGTGGCTCTTCGTTTGTTG-3’; rp49, forward, 5’-CCAGTCGGATCGATATGCTAAG-3’, rp49, reverse, 5’-CCGATGTTGGGCATCAGATA-3’; lin28, forward 5’-CAGCCAGAGTTCCACTTCAT-3’, lin28, reverse 5’-GGCCACGTTGAACCATTTG-3’.
Intensity measurements.
The intensity measurements for the samples were created in ImageJ (NIH) using RGB Profiler Macro. A rectangular selection around the hub cells and or individual cells, which were normalized with the background to get the accurate measurement. Similar settings were used for the all the measurement in the same day for the same set of experiments.
Statistical analysis
All P values were calculated using a two-tailed unpaired t-test (GraphPad Prism 7) Average values are presented as mean ± s.e.m. Unless otherwise noted, *P<0.05, **P<0.01, ***P<0.001 and ****P<0.0001. n.s. not significant.
Supplementary Materials
Author Contributions
PS conducted most of the experiments. WJ, VT, PS performed RT-PCR analysis. YJ conducted confocal microscopy for Fig. 4 D, G. BB analyzed the results and wrote the paper. PS and CK conceived the idea, analyzed the results, and wrote the paper.
Acknowledgments
We thank the Bloomington Drosophila Stock Center, Vienna Drosophila RNAi Center, and Exelixis Drosophila Collection for flies; and the Drosophila Genetic Resource Center for cDNA and Developmental Studies Hybridoma Bank for antibodies.
Conflicts of Interest
None declared.
Funding
This research was supported by the National Research Foundation (NRF) of Korea, NRF-2018R1D1A1B07 049100 to CK and National Institutes of Health, United States, 5R01GM108712 to B.B.
References
- 1. Wallenfang MR, Nayak R, DiNardo S. Dynamics of the male germline stem cell population during aging of Drosophila melanogaster. Aging Cell. 2006; 5:297–304. https://doi.org/10.1111/j.1474-9726.2006.00221.x [PubMed]
- 2. Zhou S, Greenberger JS, Epperly MW, Goff JP, Adler C, Leboff MS, Glowacki J. Age-related intrinsic changes in human bone-marrow-derived mesenchymal stem cells and their differentiation to osteoblasts. Aging Cell. 2008; 7:335–43. https://doi.org/10.1111/j.1474-9726.2008.00377.x [PubMed]
- 3. Encinas JM, Michurina TV, Peunova N, Park JH, Tordo J, Peterson DA, Fishell G, Koulakov A, Enikolopov G. Division-coupled astrocytic differentiation and age-related depletion of neural stem cells in the adult hippocampus. Cell Stem Cell. 2011; 8:566–79. https://doi.org/10.1016/j.stem.2011.03.010 [PubMed]
- 4. Kao SH, Tseng CY, Wan CL, Su YH, Hsieh CC, Pi H, Hsu HJ. Aging and insulin signaling differentially control normal and tumorous germline stem cells. Aging Cell. 2015; 14:25–34. https://doi.org/10.1111/acel.12288 [PubMed]
- 5. Tseng CY, Kao SH, Wan CL, Cho Y, Tung SY, Hsu HJ. Notch signaling mediates the age-associated decrease in adhesion of germline stem cells to the niche. PLoS Genet. 2014; 10:e1004888. https://doi.org/10.1371/journal.pgen.1004888 [PubMed]
- 6. Boyle M, Wong C, Rocha M, Jones DL. Decline in self-renewal factors contributes to aging of the stem cell niche in the Drosophila testis. Cell Stem Cell. 2007; 1:470–78. https://doi.org/10.1016/j.stem.2007.08.002 [PubMed]
- 7. Pan KZ, Palter JE, Rogers AN, Olsen A, Chen D, Lithgow GJ, Kapahi P. Inhibition of mRNA translation extends lifespan in Caenorhabditis elegans. Aging Cell. 2007; 6:111–19. https://doi.org/10.1111/j.1474-9726.2006.00266.x [PubMed]
- 8. Chakkalakal JV, Jones KM, Basson MA, Brack AS. The aged niche disrupts muscle stem cell quiescence. Nature. 2012; 490:355–60. https://doi.org/10.1038/nature11438 [PubMed]
- 9. Wallenfang MR. Aging within the Stem Cell niche. Dev Cell. 2007; 13:603–04. https://doi.org/10.1016/j.devcel.2007.10.011 [PubMed]
- 10. Toledano H. The role of the heterochronic microRNA let-7 in the progression of aging. Exp Gerontol. 2013; 48:667–70. https://doi.org/10.1016/j.exger.2012.08.006 [PubMed]
- 11. Kiger AA, Jones DL, Schulz C, Rogers MB, Fuller MT. Stem cell self-renewal specified by JAK-STAT activation in response to a support cell cue. Science. 2001; 294:2542–45. https://doi.org/10.1126/science.1066707 [PubMed]
- 12. Toledano H, D’Alterio C, Czech B, Levine E, Jones DL. The let-7-Imp axis regulates ageing of the Drosophila testis stem-cell niche. Nature. 2012; 485:605–10. https://doi.org/10.1038/nature11061 [PubMed]
- 13. Geng C, Macdonald PM. Imp associates with squid and Hrp48 and contributes to localized expression of gurken in the oocyte. Mol Cell Biol. 2006; 26:9508–16. https://doi.org/10.1128/MCB.01136-06 [PubMed]
- 14. Viswanathan SR, Daley GQ, Gregory RI. Selective blockade of microRNA processing by Lin28. Science. 2008; 320:97–100. https://doi.org/10.1126/science.1154040 [PubMed]
- 15. Heo I, Joo C, Cho J, Ha M, Han J, Kim VN. Lin28 mediates the terminal uridylation of let-7 precursor MicroRNA. Mol Cell. 2008; 32:276–84. https://doi.org/10.1016/j.molcel.2008.09.014 [PubMed]
- 16. Piskounova E, Viswanathan SR, Janas M, LaPierre RJ, Daley GQ, Sliz P, Gregory RI. Determinants of microRNA processing inhibition by the developmentally regulated RNA-binding protein Lin28. J Biol Chem. 2008; 283:21310–14. https://doi.org/10.1074/jbc.C800108200 [PubMed]
- 17. Xu B, Zhang K, Huang Y. Lin28 modulates cell growth and associates with a subset of cell cycle regulator mRNAs in mouse embryonic stem cells. RNA. 2009; 15:357–61. https://doi.org/10.1261/rna.1368009 [PubMed]
- 18. Balzer E, Heine C, Jiang Q, Lee VM, Moss EG. LIN28 alters cell fate succession and acts independently of the let-7 microRNA during neurogliogenesis in vitro. Development. 2010; 137:891–900. https://doi.org/10.1242/dev.042895 [PubMed]
- 19. Shyh-Chang N, Daley GQ. Lin28: primal regulator of growth and metabolism in stem cells. Cell Stem Cell. 2013; 12:395–406. https://doi.org/10.1016/j.stem.2013.03.005 [PubMed]
- 20. Vadla B, Kemper K, Alaimo J, Heine C, Moss EG. lin-28 controls the succession of cell fate choices via two distinct activities. PLoS Genet. 2012; 8:e1002588. https://doi.org/10.1371/journal.pgen.1002588 [PubMed]
- 21. Yang M, Yang SL, Herrlinger S, Liang C, Dzieciatkowska M, Hansen KC, Desai R, Nagy A, Niswander L, Moss EG, Chen JF. Lin28 promotes the proliferative capacity of neural progenitor cells in brain development. Development. 2015; 142:1616–27. https://doi.org/10.1242/dev.120543 [PubMed]
- 22. Chen CH, Luhur A, Sokol N. Lin-28 promotes symmetric stem cell division and drives adaptive growth in the adult Drosophila intestine. Development. 2015; 142:3478–87. https://doi.org/10.1242/dev.127951 [PubMed]
- 23. Peng S, Chen LL, Lei XX, Yang L, Lin H, Carmichael GG, Huang Y. Genome-wide studies reveal that Lin28 enhances the translation of genes important for growth and survival of human embryonic stem cells. Stem Cells. 2011; 29:496–504. https://doi.org/10.1002/stem.591 [PubMed]
- 24. Wilbert ML, Huelga SC, Kapeli K, Stark TJ, Liang TY, Chen SX, Yan BY, Nathanson JL, Hutt KR, Lovci MT, Kazan H, Vu AQ, Massirer KB, et al. LIN28 binds messenger RNAs at GGAGA motifs and regulates splicing factor abundance. Mol Cell. 2012; 48:195–206. https://doi.org/10.1016/j.molcel.2012.08.004 [PubMed]
- 25. Cho J, Chang H, Kwon SC, Kim B, Kim Y, Choe J, Ha M, Kim YK, Kim VN. LIN28A is a suppressor of ER-associated translation in embryonic stem cells. Cell. 2012; 151:765–77. https://doi.org/10.1016/j.cell.2012.10.019 [PubMed]
- 26. Yang J, Bennett BD, Luo S, Inoue K, Grimm SA, Schroth GP, Bushel PR, Kinyamu HK, Archer TK. LIN28A modulates splicing and gene expression programs in breast cancer cells. Mol Cell Biol. 2015; 35:3225–43. https://doi.org/10.1128/MCB.00426-15 [PubMed]
- 27. Stratoulias V, Heino TI, Michon F. Lin-28 regulates oogenesis and muscle formation in Drosophila melanogaster. PLoS One. 2014; 9:e101141. https://doi.org/10.1371/journal.pone.0101141 [PubMed]
- 28. Rougvie AE, Moss EG. Developmental transitions in C. elegans larval stages. Curr Top Dev Biol. 2013; 105:153–80. https://doi.org/10.1016/B978-0-12-396968-2.00006-3 [PubMed]
- 29. Moss EG, Tang L. Conservation of the heterochronic regulator Lin-28, its developmental expression and microRNA complementary sites. Dev Biol. 2003; 258:432–42. https://doi.org/10.1016/S0012-1606(03)00126-X [PubMed]
- 30. Zheng K, Wu X, Kaestner KH, Wang PJ. The pluripotency factor LIN28 marks undifferentiated spermatogonia in mouse. BMC Dev Biol. 2009; 9:38. https://doi.org/10.1186/1471-213X-9-38 [PubMed]
- 31. Papagiannouli F, Lohmann I. Shaping the niche: lessons from the Drosophila testis and other model systems. Biotechnol J. 2012; 7:723–36. https://doi.org/10.1002/biot.201100352 [PubMed]
- 32. de Cuevas M, Matunis EL. The stem cell niche: lessons from the Drosophila testis. Development. 2011; 138:2861–69. https://doi.org/10.1242/dev.056242 [PubMed]
- 33. McGuire SE, Mao Z, Davis RL. Spatiotemporal gene expression targeting with the TARGET and gene-switch systems in Drosophila. Sci STKE. 2004; 2004:pl6. https://doi.org/10.1126/stke.2202004pl6 [PubMed]
- 34. Leatherman JL, Dinardo S. Zfh-1 controls somatic stem cell self-renewal in the Drosophila testis and nonautonomously influences germline stem cell self-renewal. Cell Stem Cell. 2008; 3:44–54. https://doi.org/10.1016/j.stem.2008.05.001 [PubMed]
- 35. Chen HW, Chen X, Oh SW, Marinissen MJ, Gutkind JS, Hou SX. mom identifies a receptor for the Drosophila JAK/STAT signal transduction pathway and encodes a protein distantly related to the mammalian cytokine receptor family. Genes Dev. 2002; 16:388–98. https://doi.org/10.1101/gad.955202 [PubMed]
- 36. Flaherty MS, Salis P, Evans CJ, Ekas LA, Marouf A, Zavadil J, Banerjee U, Bach EA. chinmo is a functional effector of the JAK/STAT pathway that regulates eye development, tumor formation, and stem cell self-renewal in Drosophila. Dev Cell. 2010; 18:556–68. https://doi.org/10.1016/j.devcel.2010.02.006 [PubMed]
- 37. Balzeau J, Menezes MR, Cao S, Hagan JP. The LIN28/let-7 Pathway in Cancer. Front Genet. 2017; 8:31. https://doi.org/10.3389/fgene.2017.00031 [PubMed]
- 38. Voog J, D’Alterio C, Jones DL. Multipotent somatic stem cells contribute to the stem cell niche in the Drosophila testis. Nature. 2008; 454:1132–36. https://doi.org/10.1038/nature07173 [PubMed]
- 39. Dinardo S, Okegbe T, Wingert L, Freilich S, Terry N. lines and bowl affect the specification of cyst stem cells and niche cells in the Drosophila testis. Development. 2011; 138:1687–96. https://doi.org/10.1242/dev.057364 [PubMed]
- 40. Kitadate Y, Shigenobu S, Arita K, Kobayashi S. Boss/Sev signaling from germline to soma restricts germline-stem-cell-niche formation in the anterior region of Drosophila male gonads. Dev Cell. 2007; 13:151–59. https://doi.org/10.1016/j.devcel.2007.05.001 [PubMed]
- 41. Lee M, Nguyen TM, Kim K. In-depth study of lin-28 suggests selectively conserved let-7 independent mechanism in Drosophila. Gene. 2019; 687:64–72. https://doi.org/10.1016/j.gene.2018.11.017 [PubMed]
- 42. McLeod CJ, Wang L, Wong C, Jones DL. Stem cell dynamics in response to nutrient availability. Curr Biol. 2010; 20:2100–05. https://doi.org/10.1016/j.cub.2010.10.038 [PubMed]
- 43. Resende LP, Boyle M, Tran D, Fellner T, Jones DL. Headcase promotes cell survival and niche maintenance in the Drosophila testis. PLoS One. 2013; 8:e68026. https://doi.org/10.1371/journal.pone.0068026 [PubMed]
- 44. Fidler A, Boulay L, Wawersik M. Sonication-facilitated immunofluorescence staining of late-stage embryonic and larval Drosophila tissues in situ. J Vis Exp. 2014; 90:e51528. https://doi.org/10.3791/51528 [PubMed]
- 45. Kramer MF. Stem-loop RT-qPCR for miRNAs. Curr Protoc Mol Biol. 2011; Chapter 15: Unit 15 0.