



Introduction
Cellular senescence is a stress response mechanism induced by different types of insults such as telomere attrition, DNA damage, and oncogenic mutations, among others [1]. First described in cultured human diploid fibroblasts after successive rounds of division [2], its main hallmarks are irreversible growth arrest, alterations of cell size and morphology, increased lysosomal activity, expression of anti-proliferative proteins, resistance to apoptosis, activation of damage-sensing signaling routes. Another important characteristic is the regulated secretion of interleukins (ILs), inflammatory factors, chemokines, proteases and growth factors, termed the senescence-associated secretory phenotype (SASP) [3].
As there is ample evidence placing senescent cells as one of the causes of age-related dysfunctions, it has been considered to be one of the hallmarks of aging [4]. It was recently demonstrated that elimination of senescent cells by genetic or pharmacological approaches delays the onset of aging-related diseases, such as cancer, neurodegenerative disorders or cardiovascular diseases, among others, showing that the chronic presence of these cells is not essential [5–7]. Conversely, local injections of senescent cells drive aging-related diseases [8, 9]. This data, together with that obtained from tissues of patients with different diseases and ages, has established causality of senescent cells in some aging-related pathologies [10, 11].
Current therapies targeting senescent cells are focused on: i) specific killing of these cells by senolytics; ii) specific inhibition of the secretory phenotype (anti-SASP strategy); and iii) improving clearance of senescent cells by the immune system [12]. In addition, currently available senescence-inducing therapies for cancer stop tumor growth while causing accumulation of senescent cells [13, 14], which subsequently become a problem for the organism [15].
This review will summarize the hypothetical scenarios that each anti-cell senescence approach (described above) could face, either alone or in combination, with a discussion of open questions that should be kept in mind when targeting senescent cells.
Triggers of cell senescence
The onset of senescence in healthy tissue occurs in response to different internal and external stimuli, such as telomere attrition, DNA damage (alkylating agents, radiation), oncogene activation, mitochondrial dysfunction, and spindle, epigenetic, endoplasmic reticulum (ER) and proteotoxic stress [16–19]. The type and duration of the stimulus dictates the final effect on the senescent cells [20]. These cells display a characteristic phenotype comprising specific cell/nuclear morphology (increased size, abnormal shape and nuclear envelope changes), apoptosis resistance, chromatin redistribution (senescence-associated heterochromatin foci and senescence-associated distension of satellites), epigenetic markers (e.g. H3K9Me3), lipofuscin accumulation, SASP, and overexpression of proteins such as p53, p16Ink4a, p21WAF1, Differentiated Embryo Chondrocyte-expressed gene 1 (DEC1) and senescence-associated β-Gal (SA-β-Gal) [13, 21–24]. To date there is no universal marker for senescence, and identification of senescent cells is based on the combined detection of two or more phenotypic aspects mentioned above, such as SA-β-Gal, p16Ink4a or p21WAF1 [10].
One of the characteristic phenotypic hallmarks of cell senescence is the secretion of a plethora of factors that affect their environment (SASP), which also serves as a call for the immune system to recognize and eliminate the senescent cells [3, 25]. Among the SASP factors that seem responsible for attraction of immune cells are CSF (colony stimulating factor 1), CXCL-1 (chemokine C-X-C motif ligand 1), MCP-1 (monocyte chemoattractant protein 1) and ICAM-1 (intercellular adhesion molecule 1) [25]. In this scenario of acute or short-term senescence, the tissue returns to normal after a regeneration process [17] (Figure 1, steps 1-4). The regeneration is a fundamental process to avoid tissue atrophy and dysfunction. In this scenario of replacement of senescent cells, we should keep in mind the different capacity of renewal of some tissues with respect to others, and the exhausted or damaged state of stem cells that can lead to functionally compromised differentiated cells or carcinogenesis [26].
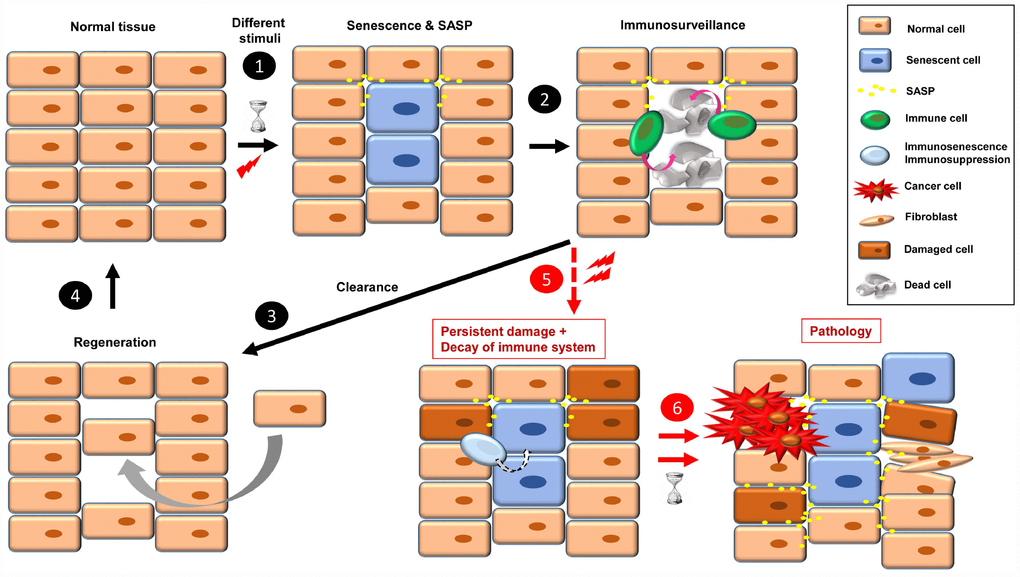
Figure 1. The onset of cellular senescence in normal tissue takes place in response to different stimuli (1). Some SASP factors are involved in immune cell recruitment, which act in the clearance of the senescent cells (2). Then, to restore the normal tissue, a regeneration process is necessary (3, 4). When a combination of persistent damage and immune system decay occurs, senescent cells accumulate, creating a pro-inflammatory and pro-tumorigenic environment and fibrotic tissue. Over time, this leads to disease, such as cancer progression, insulin resistance, osteoarthritis, atherosclerosis, and brain pathologies, among others (5, 6).
Implication of cell senescence in disease
Acute senescent cells play a direct role in tumor suppression, efficient wound healing, embryogenesis, placental formation, and tissue regeneration, among other processes [17]. At this point, both their onset and primary effect are positive for the organism [17, 20].
When senescence-inducing stimuli persist and decrease the ability of the immune system to recognize and eliminate senescent cells (by either immunosenescence or immunosuppression), these cells accumulate. The continual presence of senescent cells negatively affects their environment, inducing damage, instability or senescence in other cells through SASP [1, 27]. Over time, these “secondary” damaged cells can become either pro-tumorigenic or senescent, which increases the cellular instability of the tissue, leading to dysfunction and disease [27] (Figure 1, steps 5 and 6). In this sense, some SASP factors play a direct role in fibroblast activation and uncontrolled fibrotic scarring [28].
Chronic senescent cells (also termed “zombie” cells) have been associated with the onset of several diseases [1, 10, 13, 17]. In the last few years there have been extensive studies to elucidate the causative role of senescence in the onset of different pathologies [17]. These studies were mainly based on: i) detection of senescent cells in tissues/organs from patients or animal models; or ii) improvement in tissue/organ functions upon removal of senescent cells in mice, by either genetic or pharmacological interventions. This is a list of some age-related diseases where cellular senescence seems to play an important role:
Cancer
Aging is the main cause of cancer [29], and the presence of senescent cells in aged tissues or xenograft models correlates with the incidence of cancer [30, 31]. Their specific removal led to a delay in tumor formation and reduced metastasis [6]. It is also important to note that both senolytics and senomorphics are currently being used in clinical trials for the treatment of numerous types of cancer, such as leukemia, lung cancer, melanoma and glioblastoma, among others [16].
Neurodegenerative disorders
Senescent cell accumulation has been detected at sites of brain pathology [7, 32, 33]. The presence of senescent astrocytes correlates with the onset of pathologies such as Parkinson’s and Alzheimer’s disease [34]. Interestingly, Tau protein induces cellular senescence in neurons, and specific clearance of senescent astrocytes and microglia, reduced Tau-containing neurofibrillary tangle, neuron loss and ventricular enlargement [7, 8]. Moreover, it has been proposed a role of senescent cells in multiple sclerosis [33].
Cardiovascular disease
Senescent cells play a key role in atherosclerosis, and their specific removal reduced progression of the disease [35]. Moreover, senescent macrophages seem to contribute to coronary heart disease, and cell senescence in the aorta increases vascular stiffness [13].
Osteoarthritis
This degenerative disease causes the joints to become painful and stiff, and accumulation of senescent cells correlates with its progression [36]. In mouse models, local injections of these cells induce an osteoarthritis-like condition [9], whereas their clearance improves health by attenuating development of post-traumatic osteoarthritis [37].
Type 2 diabetes
Aging is the main cause of type 2 diabetes, and there is association between disease progression and detection of senescent markers. Senescent β-cells affect glucose homeostasis, although further work is needed to elucidate the exact role of senescence [20, 38, 39].
Idiopathic pulmonary fibrosis (IPF)
This chronic lung disease results in scarring, affecting primarily older adults. Tissues from IPF patients display some phenotypical characteristics of senescent cells, and when these cells were removed by senolytics, pulmonary functions improved [104].
Cachexia
In this disease adipocyte differentiation is disrupted by senescent cells, causing weight loss, muscle wasting and loss of body fat, leading to metabolic dysfunction and loss of adaptive thermogenic capacity [10]. When senescent cells were removed, tissue homeostasis recovered [6, 75].
Cataracts
Characterized by opacity of the lens of the eye [109], the lens capsules from patients suffering cataracts show accumulation of senescent human lens epithelial cells [105]. Removal of these cells by genetic approaches decreased the incidence of cataracts in old mice [6].
Liver diseases
The presence of senescent cells correlates with the onset of liver fibrosis, cirrhosis and non-alcoholic fatty liver disease. Elimination of these cells reduced liver fat accumulation [10, 106].
Metabolic syndrome
A collection of metabolic disorders such as increased blood pressure, high blood sugar, excess body fat (around the waist) and abnormal cholesterol levels. Endothelial cell senescence is involved in systemic metabolic dysfunction and glucose intolerance [13, 107].
Erectile dysfunction
The presence of senescent cells is directly related to endothelial dysfunction. SASP factors seem mediate this effect, and importantly, removal of senescent cells led to improvement of erectile function in mice [40].
Altogether, this data highlights the importance of targeting these cells in order to delay or cure different diseases.
Strategies to suppress senesent cells
Senolytics
An option to eliminate the negative effects of chronic senescent cells is to kill them specifically, using compounds called senolytics (Figure 2), which target pathways activated in senescent cells [16]. The list of these senolytic tool compounds is extensive and continuously growing. In Table 1 are shown the noteworthy ones. Chronic/periodic administration of senolytics kills senescent cells that are generated in the tissues, and the immune system is responsible for clearing apoptotic bodies for subsequent regeneration with new cells (Figure 2, steps 1-3). Senolytics target key proteins mainly involved in apoptosis, such as Bcl-2, Bcl-XL, p53, p21, PI3K, AKT, FOXO4 and p53. See Table 1 for references.
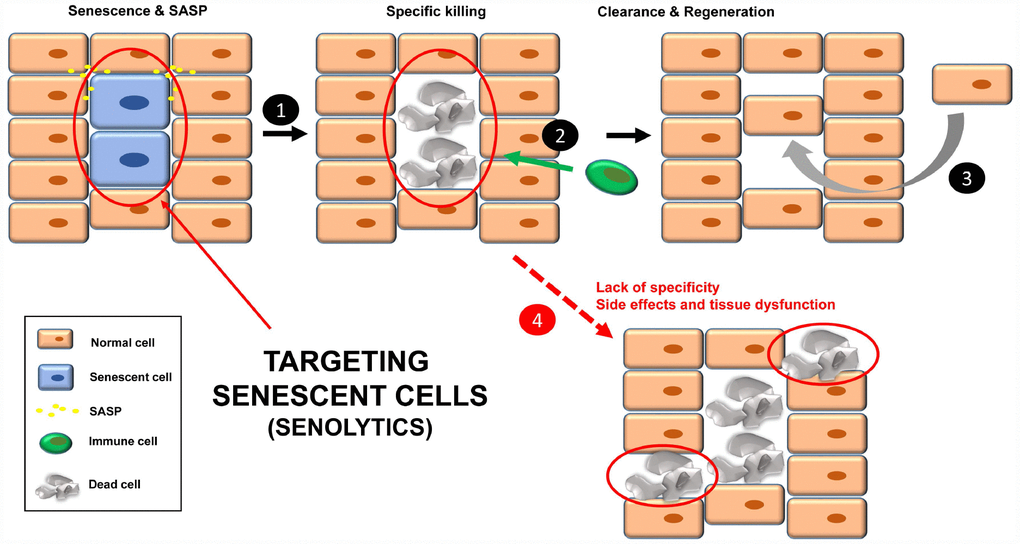
Figure 2. Treatment with senolytics to specifically kill senescent cells (1). Over time, these apoptotic bodies will be cleared by the immune system (2). Finally, a regenerative process will lead to normal tissue functions (3). Normal cells could be affected by either the lack of specificity of the senolytics or chronic treatment, leading to tissue dysfunction (4).
Table 1. List of senolytics and their targets.
| Senolytic | Target/function | References | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Apoptosis | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Dasatinib (D) | Inhibitor EFNB*-dependent suppression of apoptosis | [51] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Quercetin (Q) | PI3K/AKT, BCL-2, p53, p21, Serpine | [51] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ABT 737 | BCL-W and BCL-XL inhibitor | [52] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ABT 263 (Navitoclax; UBX0101) | BCL-2, BCL-XL and BCL-W inhibitors | [37, 53, 54] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1331852, A1155463 | BCL-XL | [55] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Fisetin | PI3K/AKT and ROS | [55] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FOXO4-related peptide (DRI) | Inhibitor of FOXO4-p53 interaction | [44] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Delivery options** | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gal-encapsulated cytotoxics | SA-β-Gal | [42] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| *AKT; protein kinase B. BCL; B-cell lymphoma. EFNB; ephrin ligand B. FOXO; forkhead box proteins O. PI3K; phosphatidylinositol 3-kinase. ROS; reactive oxygen species. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| **It helps improve senolysis by directed targeting. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Although senolytics are supposed to be specific for senescent cells, there are always unwanted damage/side effects since the administration is not directed [41] (Figure 2, step 4). In this regard, a new strategy has been recently described to specifically target senescent cells in mice, using nanocapsules containing toxins (or senolytics) [42]. The outer layer of these nanocapsules are composed of substrates for enzymes that are overexpressed in senescent cells. In this way, the toxin (senolytic) will only be released inside senescent cells, killing them [42]. Thus, these nanocapsules are a vehicle to specifically deliver any type of senolytic into senescent cells in mice. The specificity of the delivery is important in non-targeted senolytics (natural product derivatives with less defined biological activities), such as quercetin and fisetin.
Though there have been numerous reports showing the benefits of senolytics, it is important to highlight the recently described effects of dasatinib + quercetin (D + Q) treatment on lifespan in old animals [43]. Transplant of senescent cells into healthy mice caused physical dysfunction, which was reversed by oral administration of D + Q [43]. Also, clearance of senescent neurons improved neurological functions in transgenic mice mimicking Tau aggregation-dependent neurodegenerative disease [8]. It is also important to note that the treatment with the peptide FOXO4-DRI restored renal functions in both old (normal) mice and mice with accelerated aging [44]. As indicated above, some senolytics are currently being used in clinical trials for treating different diseases [16]. In this sense it is important to mention that MDM2 inhibitors, targeting p53, are also in clinical phases as anti-cancer therapies [45].
Remaining questions
There is reasonable doubt about the fate of the dead senescent cells, especially when the immune system of the patient is depressed (by either immunosenescence or immunosuppression). The accumulation of these apoptotic bodies may have undesired side effects (i.e. further release pro-inflammatory factors in an already-damaged tissue) [10]. Also, as indicated before, the possible side effects of periodic/chronic treatments should not be ignored. In fact, toxic effects after systemic administration of BCL family inhibitors have been described in patients, such as thrombocytopenia and neutropenia [41]. It would be desirable that treatments with senolytics are as sporadic as possible, without affecting efficacy. Lastly, and as indicated above, the regeneration process is an important issue to be analyzed in the tissues where senescence clearance has taken place.
SASP inhibitors (or senomorphics)
Another strategy to inhibit the functions of senescent cells is through the specific silencing of SASP [16, 46], the complex mixture of soluble factors such as cytokines, chemokines, growth factors, proteases and angiogenic factors that mediates the paracrine and autocrine functions of senescent cells [3, 25] (Figure 3). The qualitative and quantitative composition of this secretome is different depending on the cell type and the senescence-inducing stimulus, and becomes fully active a few days after the persistent stimulus [3, 47, 48]. Senomorphics inhibit SASP functions by targeting pathways such as p38 mitogen-activated protein kinase (MAPK), NF-κB, IL-1α, mTOR and PI3K/AKT (Table 2), which act at the level of transcription, translation or mRNA stabilization [21]. Alternatively, inhibition may be achieved by specific neutralizing antibodies against individual SASP factors (protein function inhibition), as is the case for IL-1α, IL-8 and IL-6.
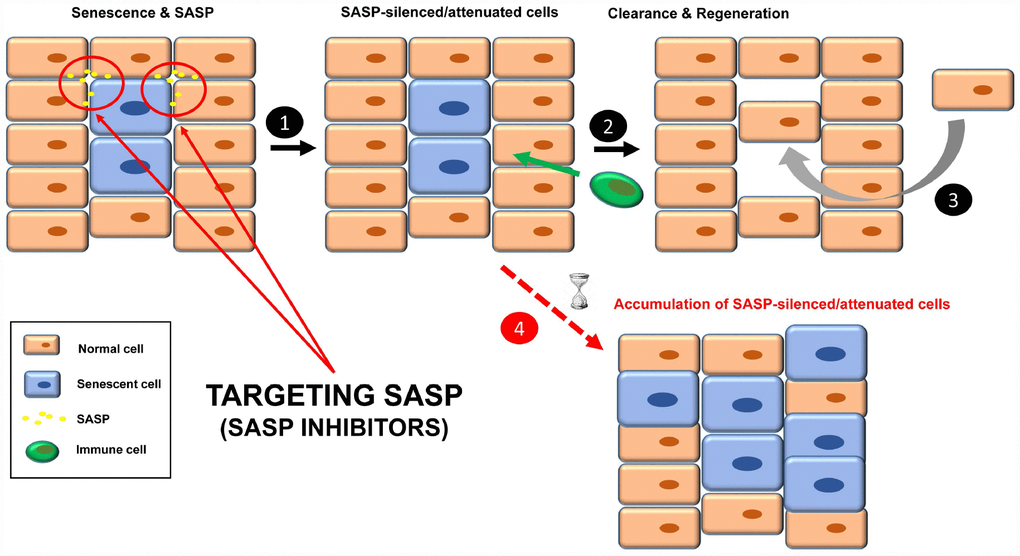
Figure 3. Treatment with senomorphics to inhibit SASP factors in senescent cells (1). Over time, these cells will be removed by immune cells (2). Finally, a regenerative process will lead to normal tissue functions (3). In aged or immunosuppressed individuals, this strategy would lead to an accumulation of SASP-silenced/attenuated senescent cells (4).
Table 2. List of senomorphics and their targets.
| SASP inhibitor | Target/function* | References | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SB 203580 | p38 MAPK** inhibitor | ([60] Reviewed by [12]) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| UR-135756, BIRB 796 | p38 MAPK inhibitor | [61] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Resveratrol | NF-ƙB inhibitor (IĸB-kinase inhibitor), AMPK and SIRT1 activator, others | [62–64] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Apigenin, Wogonin, Kaempferol | NF-ƙB inhibitors (IĸB-zeta) | [65] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Metformin | Inhibition of IKK/NF-ƙB, mitochondrial electron tranport, mitochondrial GPDH, and KDM6A/UTX, AMPK activator, others | [66–70] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Cortisol/corticosterone | IL-1α/NF-ƙB pathway inhibitors | [71] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NDGA | ROS (free radical scavenger) | [72] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Rapamycin | mTOR inhibitor, membrane-bound IL-1A translation inhibition, prelamin A, 53BP1 | [73] [74] [110] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ruxolitinib | Inhibition of JAK1/2 and ROCK | [75, 76] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| *For many of the SASP inhibitors listed there have been described several targets. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| **53BP1; p53 binding protein 1. AMPK; AMP-activated protein kinase. IKK; IĸB kinase. JAK; Janus kinase. KDM6A/UTX; lysine demethylase 6A. MAPK; mitogen-activated protein kinase. mTOR; mammalian target of rapamycin. NDGA; nordihydroguaiaretic acid. NF-ƙB; nuclear factor kappa light chain enhancer of activated B cells. ROS; reactive oxygen species. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
As IL-1α plays a direct role in SASP regulation, targeting either the receptor (IL-1αR) or the ligand (IL-1α) leads to decreased global SASP expression, with special emphasis on oncogene-induced senescence (OIS) [49, 50].
Importantly, the MABp1 antibody (neutralizing anti-human IL-1α monoclonal) has proven efficient in clinical trials against type 2 diabetes, sarcopenia and inflammation [56–58], diseases in which senescent cells play an important causative role [10].
IL-8 is a member of the CXC motif chemokine upregulated in SASP, and is associated with some types of cancer [50]. ABX-IL-8 is a humanized monoclonal antibody against IL-8 that acts as an antagonist, impairing IL-8 signaling. Treatment with ABX-IL-8 attenuates the growth of some cancer xenografts models [59].
IL-6 is a pleiotropic cytokine also upregulated in SASP that is involved in tumor proliferation, invasion and immunosuppression. Specific inhibition of IL-6 by a neutralizing monoclonal antibody (Mab-IL-6.8) completely abolished JAK/STAT signaling [50, 77] and relieved symptoms of arthritis in a primate model (Olokizumab) [78]. Arthritis has also been causally associated with the presence of senescent cells [37].
Finally, SASP-silenced/attenuated senescent cells should be recognized by the immune system for subsequent clearance and regeneration (Figure 3, steps 2 and 3).
Remaining questions
One doubt about this strategy is how SASP-silenced/attenuated senescent cells would be cleared. Given that some SASP factors are involved in the recruitment of immune cells, SASP inhibition could make senescent cells effectively “invisible” to the immune system, therefore remaining chronically within the tissue. In fact, two senomorphics (apigenin and kaempferol) showed inhibition in cultured cells of SASP components involved in immune cell recruitment, such as CXCL-1 and CSF [65]. What would the influence of SASP-silenced senescent cells be in the tissue? Perhaps instead of being dysfunctional, the tissue would be non-functional.
Likewise, as senomorphics require chronic/continuous treatment, a major problem of these types of SASP inhibitors is the lack of specificity for senescent cells. Perhaps inhibition of individual SASP components by neutralizing antibodies (as described above) would minimize the potential side effects. As indicated for senolytics, it would be desirable if over time, the treatments with senomorphics were as sporadic as possible without affecting efficacy.
Improving immune system function
A third strategy to target senescent cells is to strengthen the immune system for efficient recognition and elimination of these cells, a process termed immunosurveillance (Figure 4, steps 1-3). The role of the immune system in the elimination of senescent cells is fundamental, and a decline in immune function is associated with an increase in the number of senescent cells and finally, disease (Figure 4, step 4) [12, 20, 79, 80].
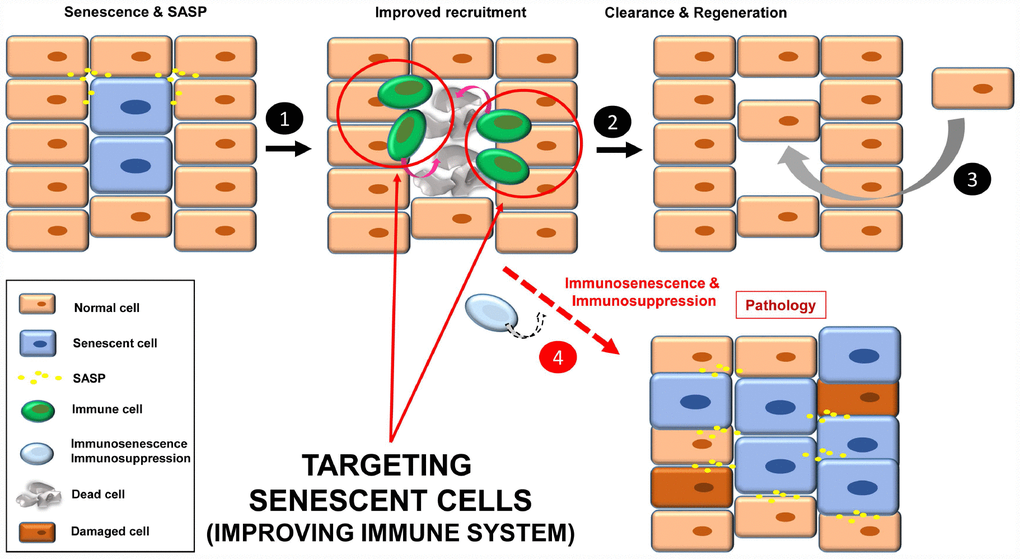
Figure 4. Improving immune system functions to efficiently remove senescent cells (1). A robust immune system targets senescent cells, leading to their removal (2). Then a regenerative process will maintain normal tissue functions (3). In situations where the immune system decays (e.g. immunosenescence or immunodepression), there will be an accumulation of senescent cells, increasing instability in the tissue/organ (4).
In this regard, there are two strategies: i) improving the specific anti-senescent cell functions; and ii) general enhancement of immune functions (to avoid senescence of immune cells involved in recognition of senescent cells).
Anti-senescent cell functions have been described in NK cells, macrophages and CD4+ T cells [20, 81]. Since these functions take place through membrane receptors, one option is to increase the binding affinity of the involved receptors. In this sense, the use of chimeric antigen receptor (CAR) T cells to target specific senescent-related molecules would be an attractive approach. This strategy is currently showing extraordinary results as anti-cancer therapy [82]. Alternatively, specifically increasing the surface expression of these receptors in senescent cells could be attempted. NK cells recognize the CD58/ICAM1 receptor present in senescent cells [83]. In the case of macrophages this recognition is not clear, and may occur through modified membrane receptors in senescent cells (glycans, lipids or vimentin), recognized by receptors present in macrophages such as CD36, IgM, SIRPα, and leptins. For T cells this process would be mainly mediated by TCRs [84].
Another possibility is to reduce the number of senescent immune cells, perhaps by depletion using specific antibodies recognizing surface markers of senescence, and in this way “rejuvenate” the immune system [84]. In this sense the recent identification of a targetable senescent cell surface marker supports this strategy [85].
NK and T cell functions decrease in older individuals. The constitutive activation of the nutrient-sensing component adenosine 5´-monophosphate-activated protein kinase (AMPK) seems to play a central role in this process [86]. Thus, an alternative approach to increase functions of these immune cells is to target AMPK functions, as the p38 MAPK inhibitor does [87]. Another approach would be to inhibit the killer cell lectin-like receptor G1 (KLRG1, or CD57 in humans), which increases on NK and T cells of older individuals. Activation of KLRG1 in NK cells is associated with activation of AMPK (via protein stabilization), which in turn would inhibit cell functions. In the case of CD8+ T cells, this mechanism may involve other inhibitory receptors, such as programmed death 1 (PD-1) and cytotoxic T lymphocyte antigen 4 (CTLA-4) [86].
The down-regulation of the CD28 receptor is a hallmark of human CD8+ T cell senescence. Interestingly these senescent T cells have been found not only in old individuals (aging process), but also associated to diseases such as cancer and arthrosis [83], which are aging-related diseases where senescent cells seem to play a causative role, as discussed above.
This fact reinforces the idea of a pivotal role of immune cells by delaying the onset of diseases related to the accumulation of chronic senescent cells. In this regard, a recent article shows that mice lacking the main cytotoxic functions of NK and T cells (perforin pathway), accelerates both senescent cell burden and aging [80].
Some current anti-cancer therapies are based on immunotherapy, that stimulates the immune system to recognize and kill disease-associated cells based on differences in the expression of antigens between pathogenic and normal cells [88]. Immunotherapy is currently used not only for different types of cancer, but also for infectious diseases, Alzheimer’s disease, and even some types of addictions [89, 90]. Senescent cells display a characteristic phenotype, which make them suitable targets for this strategy. Cell and antibody mediated responses are possible approaches, however, the specificity of senescent antigens would be the bottleneck to avoid undesirable side effects [108].
Remaining questions
Improving immune system functions to target senescent cells could be difficult in scenarios such as immunosenescence (in older individuals or patients suffering from premature aging of the immune system [91]) or immunosuppression (i.e. patients treated with corticosteroids or radiation, in cases of organ transplant, autoimmune disease or cancer). CAR-based strategies and immune system “rejuvenation” would be personalized treatments, and thus very time consuming and expensive. These strategies would rely on specific (universal) senescence receptors, and a limiting factor when detecting cell senescence is the lack of universal markers [13]. Although novel technologies are making detection of senescent cells in tissues more reliable [92, 93], the use of a combination of different biomarkers is still necessary for confirmation. Thus, personalized treatment targeting at least 2 senescence markers would increase the challenge and difficulty of the process.
Moreover, the described connection between NK and T cell activation and nutrient-sensing machinery suggests that dietary interventions could be a promising approach to maintain a healthy immune system in older individuals, and thus the ability to efficiently clear senescent cells. The up-regulation of CD28 (by forced expression of either the receptor itself or other receptor related to T cell activation) could be another attractive approach to delay the senescence process in CD8+ T cells. Last, but not least, it is important to keep in mind that a general stimulation of the components of the immune system might also induce autoimmune diseases or may also promote some hematopoietic malignancies [94, 95].
Targeting senescent cancer cells
A way to stop cancer progression is to induce senescence in tumor cells (TIS; therapy-induced senescence), through treatments targeting key pathways activated in highly proliferative cells. These treatments include DNA damage inducers (e.g. mitoxantrone, doxorubicin, γ-radiation), and inhibitors of Aurora kinase A (i.e. MLN8054, alisertib) and CDK4/6 (abemaciclib, palbociclib, ribociclib), among others [14, 96–98]. While stopping tumor growth, TIS becomes a problem for the organism in the long-term, as cancer survivors have a higher incidence of age-related diseases linked to senescence, including cardiovascular disease, neurodegeneration, sarcopenia and secondary neoplasia [19]. Cancer cells that escape from TIS (or “senescence-like” cancer cells) display some features, such as polyploidy, stemness and aggressiveness. It has been calculated that only 1 in 106 of senescent cancer cells escape from TIS. Although it seems to be a rare event, it occurs [99, 100].
At this point, it is conceivable to imagine a tissue that is already damaged, not only by tumor cells but also a mix of pre-tumorigenic and senescent cells, together with fibrosis and SASP (Figure 5). The newly senescent cells (from the tumor; TIS) would increase the level of SASP in the tissue, leading to: i) growth of new tumors (or sprouts of the former); ii) senescence induction in neighboring cells; as well as iii) an increase in fibrotic tissue. This scenario would lead to an exacerbation of the pathology that was described in the starting point (step 3).
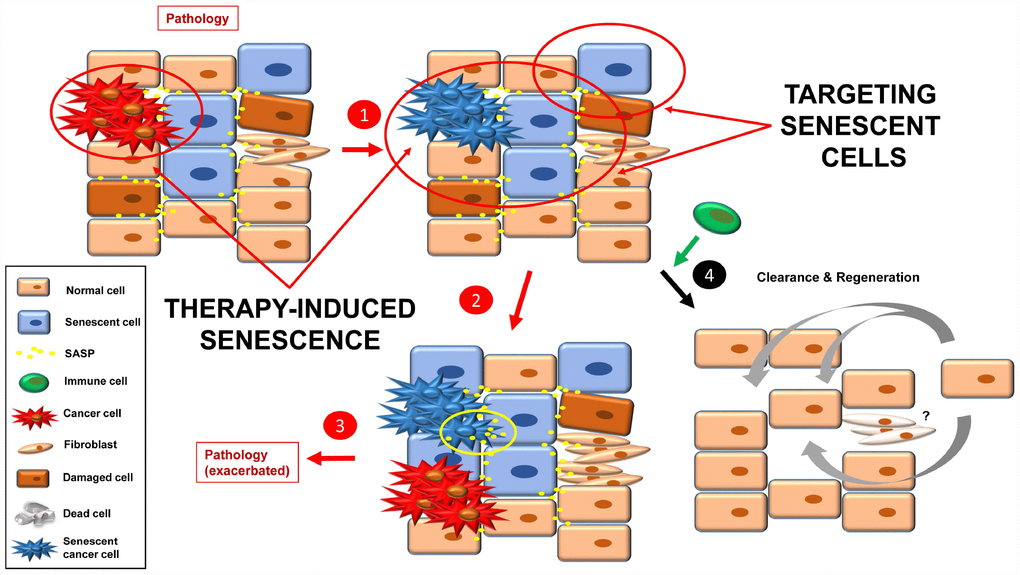
Figure 5. Inducing senescence in tumor cells will lead to an accumulation of senescence burden (1). The pro-inflammatory and pro-tumorigenic environment (more SASP factors) leads to exacerbation of the pathology (e.g. cancer relapse, fibrosis, inflammation) (2, 3). By targeting senescent cells with a combination of the approaches currently used, a better final scenario is possible (4). Fibrotic scarring may be treated by other means, or cured over time.
One solution to this situation would be to combine TIS (effective therapy to stop the growth of the tumor that is already present) with one or more of the three anti-senescent strategies presented above (senolytics, senomorphics and improved immune function) (Figure 5). Then clearance and tissue renewal processes will be necessary to restore tissue functions (Figure 5, step 4).
Remaining questions
Importantly these therapies would rely on the state of the patient´s immune system, and many patients have been affected by treatments they have received previously (immunosuppression), or by age (immunosenescence). In this sense, it is likely that in some cases it would only be necessary to inhibit SASP and not specifically induce death of the senescent cells, to avoid depending on the immune system for removal of apoptotic bodies.
And what about fibrosis? Fibrotic scarring can resolve over time, being replaced by new tissue. However, if this process is not completed (e.g. older people), the normal function of key organs can be compromised. Thus, alternative therapies should be kept in mind to treat senescence-associated fibrosis [101].
Concluding remarks
Targeting senescent cells has become an alternative therapy for treating different aging-related diseases. This therapy can be approached on three levels: i) specific killing of these cells; ii) inhibition of their secretory phenotype, therefore making them less efficient; and iii) improving our immune system for elimination of senescent cells.
The use of senolytics and senomorphics are showing promising results, although is still too early to draw conclusions. It is necessary to improve the specificity of these compounds, as well as optimize the treatment (i.e. dosage) to avoid unwanted effects. In this sense, progress has been made on the specific delivery of drugs into senescent cells by using nanocapsules. This elegant approach may overcome the problem of specificity of senolytic tool compounds when administrated in a chronic manner [42]. Importantly, senolytics and senomorphics are found in natural compounds, showing new (nutraceutical) approaches to treat aging-related diseases, although in a non-targeted way [102].
The “transformation” of normal cells into senescent ones is accomplished by a multitude of internal and external stressors in different physiological situations. Cancer cells can become senescent as well after different therapies, though the new tumor-induced senescent cells (TIS) generated are harmful in the long term. In this scenario, the three options presented here to either eliminate or “silence” the senescent cells are important to combat TIS. The combination of these pro- and anti-senescence approaches (TIS + senolytics and/or senomorphics and/or improved immune system), will play an important role in the cure of some types of cancer [98].
In future clinical trials focused on eliminating senescent cells, it will be important to determine when to initiate the treatments (age of the patients), the schedule (continuous, periodic and/or sporadic), as well as the specific markers to determine the efficacy of the therapy (see Table 3 for comparison of the therapies presented in this review). Clinical trials should be supported by robust preclinical results obtained in proper animal models.
Table 3. Comparison of the therapies presented in this review.
| Therapy | Pros | Cons |
| Senolytics | High specificity - Targeted drugs Sporadic treatments - Depending on compound efficacy | Low specificity - Non-targeted compounds Side effects - BCL family inhibitors Increase in apoptotic bodies Chronic treatments - Depending on compound efficacy |
| Senomorphics | High specificity - Targeting individual SASP components Sporadic treatments - Depending on compound efficacy | Low specificity - Targeting central pathways Chronic treatments - Depending on compound efficacy Lack of senescent cell clearance? Side effects |
| Improving immune system | High specificity - Personalized treatments - Immunotherapy Sporadic treatments Dietary interventions | Time consuming and expensive - Personalized treatments Low specificity - General activation Side effects - Autoimmunity? - Hematopoietic malignancies? Chronic treatments Patients affected by immunosuppression and/or immunosenescence |
| TIS | High specificity - Specific targets Sporadic treatments Stops tumor growth Possibility to combine with other therapies | Low specificity - General damage (chemo-radiotherapies) Side effects - New tumors - Fibrosis Chronic treatments Increasing senescence burden |
Senescent cells are the cause of several age-related diseases, which account for a high percentage of all causes of death worldwide and an expansion of morbidity. Likewise, it is estimated that by the year 2045, the number of people older than 60 will surpass, for the first time in history, the number of people under the age of 15 [103]. Thus, the approaches presented in this review highlight the urgent need for new therapies to delay or cure age/senescence-related diseases.
Acknowledgments
I am grateful to Adrián V. and Victoria Colombo for their productive discussions and support. The professional editing service NB Revisions was used for technical editing of the manuscript prior to submission.
Conflicts of Interest
The author declares that he is co-founder of SenCell Therapeutics.
Funding
Consejo Superior de Investigaciones Científicas (CSIC).
References
- 1. He S, Sharpless NE. Senescence in Health and Disease. Cell. 2017; 169:1000–11. https://doi.org/10.1016/j.cell.2017.05.015 [PubMed]
- 2. Hayflick L, Moorhead PS. The serial cultivation of human diploid cell strains. Exp Cell Res. 1961; 25:585–621. https://doi.org/10.1016/0014-4827(61)90192-6 [PubMed]
- 3. Coppé JP, Patil CK, Rodier F, Sun Y, Muñoz DP, Goldstein J, Nelson PS, Desprez PY, Campisi J. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 2008; 6:2853–68. https://doi.org/10.1371/journal.pbio.0060301 [PubMed]
- 4. López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013; 153:1194–217. https://doi.org/10.1016/j.cell.2013.05.039 [PubMed]
- 5. Baker DJ, Wijshake T, Tchkonia T, LeBrasseur NK, Childs BG, van de Sluis B, Kirkland JL, van Deursen JM. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature. 2011; 479:232–36. https://doi.org/10.1038/nature10600 [PubMed]
- 6. Baker DJ, Childs BG, Durik M, Wijers ME, Sieben CJ, Zhong J, Saltness RA, Jeganathan KB, Verzosa GC, Pezeshki A, Khazaie K, Miller JD, van Deursen JM. Naturally occurring p16(Ink4a)-positive cells shorten healthy lifespan. Nature. 2016; 530:184–89. https://doi.org/10.1038/nature16932 [PubMed]
- 7. Bussian TJ, Aziz A, Meyer CF, Swenson BL, van Deursen JM, Baker DJ. Clearance of senescent glial cells prevents tau-dependent pathology and cognitive decline. Nature. 2018; 562:578–82. https://doi.org/10.1038/s41586-018-0543-y [PubMed]
- 8. Musi N, Valentine JM, Sickora KR, Baeuerle E, Thompson CS, Shen Q, Orr ME. Tau protein aggregation is associated with cellular senescence in the brain. Aging Cell. 2018; 17:e12840. https://doi.org/10.1111/acel.12840 [PubMed]
- 9. Xu M, Bradley EW, Weivoda MM, Hwang SM, Pirtskhalava T, Decklever T, Curran GL, Ogrodnik M, Jurk D, Johnson KO, Lowe V, Tchkonia T, Westendorf JJ, Kirkland JL. Transplanted Senescent Cells Induce an Osteoarthritis-Like Condition in Mice. J Gerontol A Biol Sci Med Sci. 2017; 72:780–85. https://doi.org/10.1093/gerona/glw154 [PubMed]
- 10. McHugh D, Gil J. Senescence and aging: Causes, consequences, and therapeutic avenues. J Cell Biol. 2018; 217:65–77. https://doi.org/10.1083/jcb.201708092 [PubMed]
- 11. Waaijer ME, Parish WE, Strongitharm BH, van Heemst D, Slagboom PE, de Craen AJ, Sedivy JM, Westendorp RG, Gunn DA, Maier AB. The number of p16INK4a positive cells in human skin reflects biological age. Aging Cell. 2012; 11:722–25. https://doi.org/10.1111/j.1474-9726.2012.00837.x [PubMed]
- 12. Velarde MC, Demaria M. Targeting Senescent Cells: Possible Implications for Delaying Skin Aging: A Mini-Review. Gerontology. 2016; 62:513–18. https://doi.org/10.1159/000444877 [PubMed]
- 13. Myrianthopoulos V, Evangelou K, Vasileiou PVS, Cooks T, Vassilakopoulos TP, Pangalis GA, Kouloukoussa M, Kittas C, Georgakilas AG, Gorgoulis VG. Senescence and senotherapeutics: a new field in cancer therapy. Pharmacol Ther. 2019; 193:31–49. https://doi.org/10.1016/j.pharmthera.2018.08.006 [PubMed]
- 14. Ewald JA, Desotelle JA, Wilding G, Jarrard DF. Therapy-induced senescence in cancer. J Natl Cancer Inst. 2010; 102:1536–46. https://doi.org/10.1093/jnci/djq364 [PubMed]
- 15. Childs BG, Durik M, Baker DJ, van Deursen JM. Cellular senescence in aging and age-related disease: from mechanisms to therapy. Nat Med. 2015; 21:1424–35. https://doi.org/10.1038/nm.4000 [PubMed]
- 16. Sun Y, Coppé JP, Lam EW. Cellular Senescence: The Sought or the Unwanted? Trends Mol Med. 2018; 24:871–85. https://doi.org/10.1016/j.molmed.2018.08.002 [PubMed]
- 17. Muñoz-Espín D, Serrano M. Cellular senescence: from physiology to pathology. Nat Rev Mol Cell Biol. 2014; 15:482–96. https://doi.org/10.1038/nrm3823 [PubMed]
- 18. Kirkland JL, Tchkonia T. Cellular Senescence: A Translational Perspective. EBioMedicine. 2017; 21:21–28. https://doi.org/10.1016/j.ebiom.2017.04.013 [PubMed]
- 19. Childs BG, Gluscevic M, Baker DJ, Laberge RM, Marquess D, Dananberg J, van Deursen JM. Senescent cells: an emerging target for diseases of ageing. Nat Rev Drug Discov. 2017; 16:718–35. https://doi.org/10.1038/nrd.2017.116 [PubMed]
- 20. von Kobbe C. Cellular senescence: a view throughout organismal life. Cell Mol Life Sci. 2018; 75:3553–67. https://doi.org/10.1007/s00018-018-2879-8 [PubMed]
- 21. Hernandez-Segura A, Nehme J, Demaria M. Hallmarks of Cellular Senescence. Trends Cell Biol. 2018; 28:436–53. https://doi.org/10.1016/j.tcb.2018.02.001 [PubMed]
- 22. Salama R, Sadaie M, Hoare M, Narita M. Cellular senescence and its effector programs. Genes Dev. 2014; 28:99–114. https://doi.org/10.1101/gad.235184.113 [PubMed]
- 23. Swanson EC, Manning B, Zhang H, Lawrence JB. Higher-order unfolding of satellite heterochromatin is a consistent and early event in cell senescence. J Cell Biol. 2013; 203:929–42. https://doi.org/10.1083/jcb.201306073 [PubMed]
- 24. Qian Y, Zhang J, Yan B, Chen X. DEC1, a basic helix-loop-helix transcription factor and a novel target gene of the p53 family, mediates p53-dependent premature senescence. J Biol Chem. 2008; 283:2896–905. https://doi.org/10.1074/jbc.m708624200 [PubMed]
- 25. Coppé JP, Desprez PY, Krtolica A, Campisi J. The senescence-associated secretory phenotype: the dark side of tumor suppression. Annu Rev Pathol. 2010; 5:99–118. https://doi.org/10.1146/annurev-pathol-121808-102144 [PubMed]
- 26. He S, Nakada D, Morrison SJ. Mechanisms of stem cell self-renewal. Annu Rev Cell Dev Biol. 2009; 25:377–406. https://doi.org/10.1146/annurev.cellbio.042308.113248 [PubMed]
- 27. da Silva PFL, Ogrodnik M, Kucheryavenko O, Glibert J, Miwa S, Cameron K, Ishaq A, Saretzki G, Nagaraja-Grellscheid S, Nelson G, von Zglinicki T. The bystander effect contributes to the accumulation of senescent cells in vivo. Aging Cell. 2019; 18:e12848. https://doi.org/10.1111/acel.12848 [PubMed]
- 28. Schafer MJ, Haak AJ, Tschumperlin DJ, LeBrasseur NK. Targeting Senescent Cells in Fibrosis: Pathology, Paradox, and Practical Considerations. Curr Rheumatol Rep. 2018; 20:3. https://doi.org/10.1007/s11926-018-0712-x [PubMed]
- 29. DePinho RA. The age of cancer. Nature. 2000; 408:248–54. https://doi.org/10.1038/35041694 [PubMed]
- 30. de Magalhães JP. How ageing processes influence cancer. Nat Rev Cancer. 2013; 13:357–65. https://doi.org/10.1038/nrc3497 [PubMed]
- 31. Liu D, Hornsby PJ. Senescent human fibroblasts increase the early growth of xenograft tumors via matrix metalloproteinase secretion. Cancer Res. 2007; 67:3117–26. https://doi.org/10.1158/0008-5472.CAN-06-3452 [PubMed]
- 32. Golde TE, Miller VM. Proteinopathy-induced neuronal senescence: a hypothesis for brain failure in Alzheimer’s and other neurodegenerative diseases. Alzheimers Res Ther. 2009; 1:5. https://doi.org/10.1186/alzrt5 [PubMed]
- 33. Kritsilis M, V Rizou S, Koutsoudaki PN, Evangelou K, Gorgoulis VG, Papadopoulos D. Ageing, Cellular Senescence and Neurodegenerative Disease. Int J Mol Sci. 2018; 19:E2937. https://doi.org/10.3390/ijms19102937 [PubMed]
- 34. Chinta SJ, Woods G, Rane A, Demaria M, Campisi J, Andersen JK. Cellular senescence and the aging brain. Exp Gerontol. 2015; 68:3–7. https://doi.org/10.1016/j.exger.2014.09.018 [PubMed]
- 35. Childs BG, Baker DJ, Wijshake T, Conover CA, Campisi J, van Deursen JM. Senescent intimal foam cells are deleterious at all stages of atherosclerosis. Science. 2016; 354:472–77. https://doi.org/10.1126/science.aaf6659 [PubMed]
- 36. Price JS, Waters JG, Darrah C, Pennington C, Edwards DR, Donell ST, Clark IM. The role of chondrocyte senescence in osteoarthritis. Aging Cell. 2002; 1:57–65. https://doi.org/10.1046/j.1474-9728.2002.00008.x [PubMed]
- 37. Jeon OH, Kim C, Laberge RM, Demaria M, Rathod S, Vasserot AP, Chung JW, Kim DH, Poon Y, David N, Baker DJ, van Deursen JM, Campisi J, Elisseeff JH. Local clearance of senescent cells attenuates the development of post-traumatic osteoarthritis and creates a pro-regenerative environment. Nat Med. 2017; 23:775–81. https://doi.org/10.1038/nm.4324 [PubMed]
- 38. Helman A, Klochendler A, Azazmeh N, Gabai Y, Horwitz E, Anzi S, Swisa A, Condiotti R, Granit RZ, Nevo Y, Fixler Y, Shreibman D, Zamir A, et al. p16(Ink4a)-induced senescence of pancreatic beta cells enhances insulin secretion. Nat Med. 2016; 22:412–20. https://doi.org/10.1038/nm.4054 [PubMed]
- 39. Aguayo-Mazzucato C, van Haaren M, Mruk M, Lee TB
Jr , Crawford C, Hollister-Lock J, Sullivan BA, Johnson JW, Ebrahimi A, Dreyfuss JM, Van Deursen J, Weir GC, Bonner-Weir S. β Cell Aging Markers Have Heterogeneous Distribution and Are Induced by Insulin Resistance. Cell Metab. 2017; 25:898–910.e5. https://doi.org/10.1016/j.cmet.2017.03.015 [PubMed] - 40. Nishimatsu H, Suzuki E, Saito Y, Niimi A, Nomiya A, Fukuhara H, Kume H, Homma Y. Senescent Cells Impair Erectile Function through Induction of Endothelial Dysfunction and Nerve Injury in Mice. PLoS One. 2015; 10:e0124129. https://doi.org/10.1371/journal.pone.0124129 [PubMed]
- 41. Rudin CM, Hann CL, Garon EB, Ribeiro de Oliveira M, Bonomi PD, Camidge DR, Chu Q, Giaccone G, Khaira D, Ramalingam SS, Ranson MR, Dive C, McKeegan EM, et al. Phase II study of single-agent navitoclax (ABT-263) and biomarker correlates in patients with relapsed small cell lung cancer. Clin Cancer Res. 2012; 18:3163–69. https://doi.org/10.1158/1078-0432.CCR-11-3090 [PubMed]
- 42. Muñoz-Espín D, Rovira M, Galiana I, Giménez C, Lozano-Torres B, Paez-Ribes M, Llanos S, Chaib S, Muñoz-Martín M, Ucero AC, Garaulet G, Mulero F, Dann SG, et al. A versatile drug delivery system targeting senescent cells. EMBO Mol Med. 2018; 10:e9355. https://doi.org/10.15252/emmm.201809355 [PubMed]
- 43. Xu M, Pirtskhalava T, Farr JN, Weigand BM, Palmer AK, Weivoda MM, Inman CL, Ogrodnik MB, Hachfeld CM, Fraser DG, Onken JL, Johnson KO, Verzosa GC, et al. Senolytics improve physical function and increase lifespan in old age. Nat Med. 2018; 24:1246–56. https://doi.org/10.1038/s41591-018-0092-9 [PubMed]
- 44. Baar MP, Brandt RM, Putavet DA, Klein JD, Derks KW, Bourgeois BR, Stryeck S, Rijksen Y, van Willigenburg H, Feijtel DA, van der Pluijm I, Essers J, van Cappellen WA, et al. Targeted Apoptosis of Senescent Cells Restores Tissue Homeostasis in Response to Chemotoxicity and Aging. Cell. 2017; 169:132–147.e16. https://doi.org/10.1016/j.cell.2017.02.031 [PubMed]
- 45. Wiley CD, Schaum N, Alimirah F, Lopez-Dominguez JA, Orjalo AV, Scott G, Desprez PY, Benz C, Davalos AR, Campisi J. Small-molecule MDM2 antagonists attenuate the senescence-associated secretory phenotype. Sci Rep. 2018; 8:2410. https://doi.org/10.1038/s41598-018-20000-4 [PubMed]
- 46. Tchkonia T, Zhu Y, van Deursen J, Campisi J, Kirkland JL. Cellular senescence and the senescent secretory phenotype: therapeutic opportunities. J Clin Invest. 2013; 123:966–72. https://doi.org/10.1172/JCI64098 [PubMed]
- 47. Ito Y, Hoare M, Narita M. Spatial and Temporal Control of Senescence. Trends Cell Biol. 2017; 27:820–32. https://doi.org/10.1016/j.tcb.2017.07.004 [PubMed]
- 48. Ovadya Y, Krizhanovsky V. Senescent cells: SASPected drivers of age-related pathologies. Biogerontology. 2014; 15:627–42. https://doi.org/10.1007/s10522-014-9529-9 [PubMed]
- 49. Orjalo AV, Bhaumik D, Gengler BK, Scott GK, Campisi J. Cell surface-bound IL-1alpha is an upstream regulator of the senescence-associated IL-6/IL-8 cytokine network. Proc Natl Acad Sci USA. 2009; 106:17031–36. https://doi.org/10.1073/pnas.0905299106 [PubMed]
- 50. Di Mitri D, Alimonti A. Non-Cell-Autonomous Regulation of Cellular Senescence in Cancer. Trends Cell Biol. 2016; 26:215–26. https://doi.org/10.1016/j.tcb.2015.10.005 [PubMed]
- 51. Zhu Y, Tchkonia T, Pirtskhalava T, Gower AC, Ding H, Giorgadze N, Palmer AK, Ikeno Y, Hubbard GB, Lenburg M, O’Hara SP, LaRusso NF, Miller JD, et al. The Achilles’ heel of senescent cells: from transcriptome to senolytic drugs. Aging Cell. 2015; 14:644–58. https://doi.org/10.1111/acel.12344 [PubMed]
- 52. Yosef R, Pilpel N, Tokarsky-Amiel R, Biran A, Ovadya Y, Cohen S, Vadai E, Dassa L, Shahar E, Condiotti R, Ben-Porath I, Krizhanovsky V. Directed elimination of senescent cells by inhibition of BCL-W and BCL-XL. Nat Commun. 2016; 7:11190. https://doi.org/10.1038/ncomms11190 [PubMed]
- 53. Chang J, Wang Y, Shao L, Laberge RM, Demaria M, Campisi J, Janakiraman K, Sharpless NE, Ding S, Feng W, Luo Y, Wang X, Aykin-Burns N, et al. Clearance of senescent cells by ABT263 rejuvenates aged hematopoietic stem cells in mice. Nat Med. 2016; 22:78–83. https://doi.org/10.1038/nm.4010 [PubMed]
- 54. Zhu Y, Tchkonia T, Fuhrmann-Stroissnigg H, Dai HM, Ling YY, Stout MB, Pirtskhalava T, Giorgadze N, Johnson KO, Giles CB, Wren JD, Niedernhofer LJ, Robbins PD, Kirkland JL. Identification of a novel senolytic agent, navitoclax, targeting the Bcl-2 family of anti-apoptotic factors. Aging Cell. 2016; 15:428–35. https://doi.org/10.1111/acel.12445 [PubMed]
- 55. Zhu Y, Doornebal EJ, Pirtskhalava T, Giorgadze N, Wentworth M, Fuhrmann-Stroissnigg H, Niedernhofer LJ, Robbins PD, Tchkonia T, Kirkland JL. New agents that target senescent cells: the flavone, fisetin, and the BCL-XL inhibitors, A1331852 and A1155463. Aging (Albany NY). 2017; 9:955–63. https://doi.org/10.18632/aging.101202 [PubMed]
- 56. Dinarello CA, Simon A, van der Meer JW. Treating inflammation by blocking interleukin-1 in a broad spectrum of diseases. Nat Rev Drug Discov. 2012; 11:633–52. https://doi.org/10.1038/nrd3800 [PubMed]
- 57. Timper K, Seelig E, Tsakiris DA, Donath MY. Safety, pharmacokinetics, and preliminary efficacy of a specific anti-IL-1alpha therapeutic antibody (MABp1) in patients with type 2 diabetes mellitus. J Diabetes Complications. 2015; 29:955–60. https://doi.org/10.1016/j.jdiacomp.2015.05.019 [PubMed]
- 58. O’Sullivan Coyne G, Burotto M. MABp1 for the treatment of colorectal cancer. Expert Opin Biol Ther. 2017; 17:1155–61. https://doi.org/10.1080/14712598.2017.1347631 [PubMed]
- 59. Waugh DJ, Wilson C. The interleukin-8 pathway in cancer. Clin Cancer Res. 2008; 14:6735–41. https://doi.org/10.1158/1078-0432.CCR-07-4843 [PubMed]
- 60. Goldstein DM, Kuglstatter A, Lou Y, Soth MJ. Selective p38alpha inhibitors clinically evaluated for the treatment of chronic inflammatory disorders. J Med Chem. 2010; 53:2345–53. https://doi.org/10.1021/jm9012906 [PubMed]
- 61. Alimbetov D, Davis T, Brook AJ, Cox LS, Faragher RG, Nurgozhin T, Zhumadilov Z, Kipling D. Suppression of the senescence-associated secretory phenotype (SASP) in human fibroblasts using small molecule inhibitors of p38 MAP kinase and MK2. Biogerontology. 2016; 17:305–15. https://doi.org/10.1007/s10522-015-9610-z [PubMed]
- 62. Pitozzi V, Mocali A, Laurenzana A, Giannoni E, Cifola I, Battaglia C, Chiarugi P, Dolara P, Giovannelli L. Chronic resveratrol treatment ameliorates cell adhesion and mitigates the inflammatory phenotype in senescent human fibroblasts. J Gerontol A Biol Sci Med Sci. 2013; 68:371–81. https://doi.org/10.1093/gerona/gls183 [PubMed]
- 63. Holmes-McNary M, Baldwin AS
Jr . Chemopreventive properties of trans-resveratrol are associated with inhibition of activation of the IkappaB kinase. Cancer Res. 2000; 60:3477–83. [PubMed] - 64. Kulkarni SS, Cantó C. The molecular targets of resveratrol. Biochim Biophys Acta. 2015; 1852:1114–23. https://doi.org/10.1016/j.bbadis.2014.10.005 [PubMed]
- 65. Lim H, Park H, Kim HP. Effects of flavonoids on senescence-associated secretory phenotype formation from bleomycin-induced senescence in BJ fibroblasts. Biochem Pharmacol. 2015; 96:337–48. https://doi.org/10.1016/j.bcp.2015.06.013 [PubMed]
- 66. Moiseeva O, Deschênes-Simard X, St-Germain E, Igelmann S, Huot G, Cadar AE, Bourdeau V, Pollak MN, Ferbeyre G. Metformin inhibits the senescence-associated secretory phenotype by interfering with IKK/NF-κB activation. Aging Cell. 2013; 12:489–98. https://doi.org/10.1111/acel.12075 [PubMed]
- 67. Soberanes S, Misharin AV, Jairaman A, Morales-Nebreda L, McQuattie-Pimentel AC, Cho T, Hamanaka RB, Meliton AY, Reyfman PA, Walter JM, Chen CI, Chi M, Chiu S, et al. Metformin Targets Mitochondrial Electron Transport to Reduce Air-Pollution-Induced Thrombosis. Cell Metab. 2019; 29:335–347.e5. https://doi.org/10.1016/j.cmet.2018.09.019 [PubMed]
- 68. Cuyàs E, Verdura S, Llorach-Pares L, Fernández-Arroyo S, Luciano-Mateo F, Cabré N, Stursa J, Werner L, Martin-Castillo B, Viollet B, Neuzil J, Joven J, Nonell-Canals A, et al. Metformin directly targets the H3K27me3 demethylase KDM6A/UTX. Aging Cell. 2018; 17:e12772. https://doi.org/10.1111/acel.12772 [PubMed]
- 69. Thakur S, Daley B, Gaskins K, Vasko VV, Boufraqech M, Patel D, Sourbier C, Reece J, Cheng SY, Kebebew E, Agarwal S, Klubo-Gwiezdzinska J. Metformin Targets Mitochondrial Glycerophosphate Dehydrogenase to Control Rate of Oxidative Phosphorylation and Growth of Thyroid Cancer In Vitro and In Vivo.. Clin Cancer Res. 2018; 24:4030–43. https://doi.org/10.1158/1078-0432.CCR-17-3167 [PubMed]
- 70. Fryer LG, Parbu-Patel A, Carling D. The Anti-diabetic drugs rosiglitazone and metformin stimulate AMP-activated protein kinase through distinct signaling pathways. J Biol Chem. 2002; 277:25226–32. https://doi.org/10.1074/jbc.M202489200 [PubMed]
- 71. Laberge RM, Zhou L, Sarantos MR, Rodier F, Freund A, de Keizer PL, Liu S, Demaria M, Cong YS, Kapahi P, Desprez PY, Hughes RE, Campisi J. Glucocorticoids suppress selected components of the senescence-associated secretory phenotype. Aging Cell. 2012; 11:569–78. https://doi.org/10.1111/j.1474-9726.2012.00818.x [PubMed]
- 72. Fuhrmann-Stroissnigg H, Ling YY, Zhao J, McGowan SJ, Zhu Y, Brooks RW, Grassi D, Gregg SQ, Stripay JL, Dorronsoro A, Corbo L, Tang P, Bukata C, et al. Identification of HSP90 inhibitors as a novel class of senolytics. Nat Commun. 2017; 8:422. https://doi.org/10.1038/s41467-017-00314-z [PubMed]
- 73. Herranz N, Gallage S, Mellone M, Wuestefeld T, Klotz S, Hanley CJ, Raguz S, Acosta JC, Innes AJ, Banito A, Georgilis A, Montoya A, Wolter K, et al. mTOR regulates MAPKAPK2 translation to control the senescence-associated secretory phenotype. Nat Cell Biol. 2015; 17:1205–17. Erratum in: Nat. Cell. Biol. 2015. 17, 1370 https://doi.org/10.1038/ncb3225 [PubMed]
- 74. Lattanzi G, Ortolani M, Columbaro M, Prencipe S, Mattioli E, Lanzarini C, Maraldi NM, Cenni V, Garagnani P, Salvioli S, Storci G, Bonafè M, Capanni C, Franceschi C. Lamins are rapamycin targets that impact human longevity: a study in centenarians. J Cell Sci. 2014; 127:147–57. https://doi.org/10.1242/jcs.133983 [PubMed]
- 75. Xu M, Palmer AK, Ding H, Weivoda MM, Pirtskhalava T, White TA, Sepe A, Johnson KO, Stout MB, Giorgadze N, Jensen MD, LeBrasseur NK, Tchkonia T, Kirkland JL. Targeting senescent cells enhances adipogenesis and metabolic function in old age. eLife. 2015; 4:e12997. https://doi.org/10.7554/eLife.12997 [PubMed]
- 76. Rudolph J, Heine A, Quast T, Kolanus W, Trebicka J, Brossart P, Wolf D. The JAK inhibitor ruxolitinib impairs dendritic cell migration via off-target inhibition of ROCK. Leukemia. 2016; 30:2119–23. https://doi.org/10.1038/leu.2016.155 [PubMed]
- 77. Kuilman T, Michaloglou C, Vredeveld LC, Douma S, van Doorn R, Desmet CJ, Aarden LA, Mooi WJ, Peeper DS. Oncogene-induced senescence relayed by an interleukin-dependent inflammatory network. Cell. 2008; 133:1019–31. https://doi.org/10.1016/j.cell.2008.03.039 [PubMed]
- 78. Shaw S, Bourne T, Meier C, Carrington B, Gelinas R, Henry A, Popplewell A, Adams R, Baker T, Rapecki S, Marshall D, Moore A, Neale H, Lawson A. Discovery and characterization of olokizumab: a humanized antibody targeting interleukin-6 and neutralizing gp130-signaling. MAbs. 2014; 6:774–82. https://doi.org/10.4161/mabs.28612 [PubMed]
- 79. van Deursen JM. The role of senescent cells in ageing. Nature. 2014; 509:439–46. https://doi.org/10.1038/nature13193 [PubMed]
- 80. Ovadya Y, Landsberger T, Leins H, Vadai E, Gal H, Biran A, Yosef R, Sagiv A, Agrawal A, Shapira A, Windheim J, Tsoory M, Schirmbeck R, et al. Impaired immune surveillance accelerates accumulation of senescent cells and aging. Nat Commun. 2018; 9:5435. https://doi.org/10.1038/s41467-018-07825-3 [PubMed]
- 81. Krizhanovsky V, Yon M, Dickins RA, Hearn S, Simon J, Miething C, Yee H, Zender L, Lowe SW. Senescence of activated stellate cells limits liver fibrosis. Cell. 2008; 134:657–67. https://doi.org/10.1016/j.cell.2008.06.049 [PubMed]
- 82. Yu S, Li A, Liu Q, Li T, Yuan X, Han X, Wu K. Chimeric antigen receptor T cells: a novel therapy for solid tumors. J Hematol Oncol. 2017; 10:78. https://doi.org/10.1186/s13045-017-0444-9 [PubMed]
- 83. Vicente R, Mausset-Bonnefont AL, Jorgensen C, Louis-Plence P, Brondello JM. Cellular senescence impact on immune cell fate and function. Aging Cell. 2016; 15:400–06. https://doi.org/10.1111/acel.12455 [PubMed]
- 84. Burton DG, Stolzing A. Cellular senescence: immunosurveillance and future immunotherapy. Ageing Res Rev. 2018; 43:17–25. https://doi.org/10.1016/j.arr.2018.02.001 [PubMed]
- 85. Kim KM, Noh JH, Bodogai M, Martindale JL, Yang X, Indig FE, Basu SK, Ohnuma K, Morimoto C, Johnson PF, Biragyn A, Abdelmohsen K, Gorospe M. Identification of senescent cell surface targetable protein DPP4. Genes Dev. 2017; 31:1529–34. https://doi.org/10.1101/gad.302570.117 [PubMed]
- 86. Akbar AN. The convergence of senescence and nutrient sensing during lymphocyte ageing. Clin Exp Immunol. 2017; 187:4–5. https://doi.org/10.1111/cei.12876 [PubMed]
- 87. Di Mitri D, Azevedo RI, Henson SM, Libri V, Riddell NE, Macaulay R, Kipling D, Soares MV, Battistini L, Akbar AN. Reversible senescence in human CD4+CD45RA+CD27- memory T cells. J Immunol. 2011; 187:2093–100. https://doi.org/10.4049/jimmunol.1100978 [PubMed]
- 88. Martin Caballero J, Garzón A, González-Cintado L, Kowalczyk W, Jimenez Torres I, Calderita G, Rodriguez M, Gondar V, Bernal JJ, Ardavín C, Andreu D, Zürcher T, von Kobbe C. Chimeric infectious bursal disease virus-like particles as potent vaccines for eradication of established HPV-16 E7-dependent tumors. PLoS One. 2012; 7:e52976. https://doi.org/10.1371/journal.pone.0052976 [PubMed]
- 89. Dyer MR, Renner WA, Bachmann MF. A second vaccine revolution for the new epidemics of the 21st century. Drug Discov Today. 2006; 11:1028–33. https://doi.org/10.1016/j.drudis.2006.09.006 [PubMed]
- 90. Vetsika EK, Konsolakis G, Aggouraki D, Kotsakis A, Papadimitraki E, Christou S, Menez-Jamet J, Kosmatopoulos K, Georgoulias V, Mavroudis D. Immunological responses in cancer patients after vaccination with the therapeutic telomerase-specific vaccine Vx-001. Cancer Immunol Immunother. 2012; 61:157–68. https://doi.org/10.1007/s00262-011-1093-4 [PubMed]
- 91. Chou JP, Ramirez CM, Wu JE, Effros RB. Accelerated aging in HIV/AIDS: novel biomarkers of senescent human CD8+ T cells. PLoS One. 2013; 8:e64702. https://doi.org/10.1371/journal.pone.0064702 [PubMed]
- 92. Evangelou K, Lougiakis N, Rizou SV, Kotsinas A, Kletsas D, Muñoz-Espín D, Kastrinakis NG, Pouli N, Marakos P, Townsend P, Serrano M, Bartek J, Gorgoulis VG. Robust, universal biomarker assay to detect senescent cells in biological specimens. Aging Cell. 2017; 16:192–97. https://doi.org/10.1111/acel.12545 [PubMed]
- 93. Galbiati A, Beauséjour C, d’Adda di Fagagna F. A novel single-cell method provides direct evidence of persistent DNA damage in senescent cells and aged mammalian tissues. Aging Cell. 2017; 16:422–27. https://doi.org/10.1111/acel.12573 [PubMed]
- 94. Freitag J, Berod L, Kamradt T, Sparwasser T. Immunometabolism and autoimmunity. Immunol Cell Biol. 2016; 94:925–34. https://doi.org/10.1038/icb.2016.77 [PubMed]
- 95. Sun H, Wei S, Yang L. Dysfunction of immune system in the development of large granular lymphocyte leukemia. Hematology. 2019; 24:139–47. https://doi.org/10.1080/10245332.2018.1535294 [PubMed]
- 96. Gonzalez LC, Ghadaouia S, Martinez A, Rodier F. Premature aging/senescence in cancer cells facing therapy: good or bad? Biogerontology. 2016; 17:71–87. https://doi.org/10.1007/s10522-015-9593-9 [PubMed]
- 97. Ruscetti M, Leibold J, Bott MJ, Fennell M, Kulick A, Salgado NR, Chen CC, Ho YJ, Sanchez-Rivera FJ, Feucht J, Baslan T, Tian S, Chen HA, et al. NK cell-mediated cytotoxicity contributes to tumor control by a cytostatic drug combination. Science. 2018; 362:1416–22. https://doi.org/10.1126/science.aas9090 [PubMed]
- 98. Sieben CJ, Sturmlechner I, van de Sluis B, van Deursen JM. Two-Step Senescence-Focused Cancer Therapies. Trends Cell Biol. 2018; 28:723–37. https://doi.org/10.1016/j.tcb.2018.04.006 [PubMed]
- 99. Saleh T, Tyutyunyk-Massey L, Murray GF, Alotaibi MR, Kawale AS, Elsayed Z, Henderson SC, Yakovlev V, Elmore LW, Toor A, Harada H, Reed J, Landry JW, Gewirtz DA. Tumor cell escape from therapy-induced senescence. Biochem Pharmacol. 2019; 162:202–12. https://doi.org/10.1016/j.bcp.2018.12.013 [PubMed]
- 100. Saleh T, Tyutyunyk-Massey L, Gewirtz DA. Tumor Cell Escape from Therapy-Induced Senescence as a Model of Disease Recurrence after Dormancy. Cancer Res. 2019; 79:1044–46. https://doi.org/10.1158/0008-5472.CAN-18-3437 [PubMed]
- 101. Hecker L, Logsdon NJ, Kurundkar D, Kurundkar A, Bernard K, Hock T, Meldrum E, Sanders YY, Thannickal VJ. Reversal of persistent fibrosis in aging by targeting Nox4-Nrf2 redox imbalance. Sci Transl Med. 2014; 6:231ra47. https://doi.org/10.1126/scitranslmed.3008182 [PubMed]
- 102. Gurău F, Baldoni S, Prattichizzo F, Espinosa E, Amenta F, Procopio AD, Albertini MC, Bonafè M, Olivieri F. Anti-senescence compounds: A potential nutraceutical approach to healthy aging. Ageing Res Rev. 2018; 46:14–31. https://doi.org/10.1016/j.arr.2018.05.001 [PubMed]
- 103. Department of Economic and Social Affairs (DESA). World population ageing. New York: United Nations; 2015.
- 104. Schafer MJ, White TA, Iijima K, Haak AJ, Ligresti G, Atkinson EJ, Oberg AL, Birch J, Salmonowicz H, Zhu Y, Mazula DL, Brooks RW, Fuhrmann-Stroissnigg H, et al. Cellular senescence mediates fibrotic pulmonary disease. Nat Commun. 2017; 8:14532. https://doi.org/10.1038/ncomms14532 [PubMed]
- 105. Fu Q, Qin Z, Yu J, Yu Y, Tang Q, Lyu D, Zhang L, Chen Z, Yao K. Effects of senescent lens epithelial cells on the severity of age-related cortical cataract in humans: A case-control study. Medicine (Baltimore). 2016; 95:e3869. https://doi.org/10.1097/MD.0000000000003869 [PubMed]
- 106. Ogrodnik M, Miwa S, Tchkonia T, Tiniakos D, Wilson CL, Lahat A, Day CP, Burt A, Palmer A, Anstee QM, Grellscheid SN, Hoeijmakers JH, Barnhoorn S, et al. Cellular senescence drives age-dependent hepatic steatosis. Nat Commun. 2017; 8:15691. https://doi.org/10.1038/ncomms15691 [PubMed]
- 107. Yokoyama M, Okada S, Nakagomi A, Moriya J, Shimizu I, Nojima A, Yoshida Y, Ichimiya H, Kamimura N, Kobayashi Y, Ohta S, Fruttiger M, Lozano G, Minamino T. Inhibition of endothelial p53 improves metabolic abnormalities related to dietary obesity. Cell Rep. 2014; 7:1691–703. https://doi.org/10.1016/j.celrep.2014.04.046 [PubMed]
- 108. Ovadya Y, Krizhanovsky V. Strategies targeting cellular senescence. J Clin Invest. 2018; 128:1247–54. https://doi.org/10.1172/JCI95149 [PubMed]
- 109. Spector A. Oxidative stress-induced cataract: mechanism of action. FASEB J. 1995; 9:1173–82. https://doi.org/10.1096/fasebj.9.12.7672510 [PubMed]
- 110. Laberge RM, Sun Y, Orjalo AV, Patil CK, Freund A, Zhou L, Curran SC, Davalos AR, Wilson-Edell KA, Liu S, Limbad C, Demaria M, Li P, Hubbard GB, Ikeno Y, Javors M, Desprez PY, Benz CC, Kapahi P, Nelson PS, Campisi J. MTOR regulates the pro-tumorigenic senescence-associated secretory phenotype by promoting IL1A translation. Nat Cell Biol. 2015; 17:1049–61. https://doi.org/10.1038/ncb3195 [PubMed]