




Introduction
Aging is associated with changes in various molecular and cellular processes that ultimately lead to physiological decline and a concomitant increased risk for developing diseases, such as cancer, inflammation, and diabetes [1]. While aging affects all tissues, the brain is especially susceptible to age-related impairments due to its high energy demands. Brain aging is associated with increased mitochondrial dysfunction and dysregulated energy metabolism, which may cause decline in neuronal function leading to neurodegeneration [2, 3].
The causes of age-related brain deterioration have been addressed using a variety of approaches including comparative analysis of gene expression in the brains of young and old organisms. These studies revealed shifts in transcriptional profiles that may underlie age-related alterations in brain function. For example, postmortem analyses of human brain cortex reported significant age-related reductions in genes involved in synaptic plasticity, mitochondrial function, and vesicular transport; and increased expression of genes involved in response to oxidative stress and DNA repair [4]. Similar groups of genes are affected by age in multiple model organisms including Drosophila due to the conservation of molecular pathways associated with aging [5]. Most studies examining age-dependent transcriptional changes measured gene expression in samples collected at a single unspecified time of day; however, expression of hundreds of genes change in a daily pattern due to their circadian regulation [6]. Measuring gene expression in samples collected at multiple times of day revealed age-related reprogramming of the diurnal transcriptome in various mammalian tissues [7–9]. Our recent RNA-seq study in Drosophila identified hundreds of genes with age-related changes in expression profiles in fly heads [10]. While many genes lost rhythmic expression with age, some genes gain a diurnal rhythmic expression pattern. The most prominent among the latter group was Ldh (also known as ImpL3) which encodes the metabolic enzyme lactate dehydrogenase. Expression of Ldh is low in the heads of young flies; however, it increases several-fold and becomes rhythmic in heads of old flies in light-dark (LD) conditions [10]. Here, we sought to determine whether elevated Ldh expression affects aging phenotypes and longevity in flies.
Lactate dehydrogenase (LDH) is an enzyme that catalyzes the conversion of pyruvate, the end product of glycolysis, into lactate (and vice versa) with concomitant interconversion of NADH and NAD+. Mammalian LDH proteins are tetramers of subunits encoded mainly by the LdhA and LdhB genes [11, 12]. The proportion of LDHA and LDHB isoforms composing the tetrameric enzyme complex influences its kinetic and catalytic properties [13, 14]. In Drosophila, only one Ldh gene is expressed in adults; thus, the LDH enzyme complex exists as a homotetramer of LDH subunits, which share 71 and 75% sequence similarity with the human LDHA and LDHB proteins, respectively [15].
Lactate produced by LDH has long been regarded as a glycolytic waste product; however, many reports suggest that lactate plays important roles in brain energetics. Lactate is considered a glucose sparing metabolite that can fuel neuronal energy production, modulate neuronal excitability, and facilitate memory formation [16, 17]. On the other hand, adverse effects associated with elevated lactate levels have been reported, for example, increased LDH activity and lactate levels have been associated with tumor malignancy [14, 18]. Elevated lactate levels have also been reported in the brains of aged mice [13, 19], Alzheimer’s disease (AD) patients [20], and in the heads of AD model flies [21]. The roles of LDH and lactate in brain functions remain a topic of debate [17, 22–24] and merit further molecular and organismal studies.
Although increased LDH activity and lactate are observed during aging and in AD-related pathologies, it is unknown whether lactate is causally involved in the aging process. To address this question, we studied the biological significance of the age-related increase in Ldh gene expression in Drosophila. Here, we present evidence that increased Ldh expression is associated with elevated LDH protein levels, enzyme activity, and lactate concentrations in the heads of Drosophila. Additionally, we show that invivo manipulation of Ldh gene expression alters lifespan, locomotor activity rhythms, and age-related neurodegeneration in a cell type-specific manner.
Results
Aging is associated with increased Ldh expression and elevated lactate levels
To determine the effect of age and time of day on Ldh mRNA expression, young (5-days old) and old (55-days old) white1118 (w1118) males reared in cycles of 12 hours of light and 12 hours of darkness (LD 12:12) were collected every six hours at Zeitgeber time (ZT) 0, 6, 12, and 18. The levels of Ldh mRNA were significantly (3-5 fold) higher in the heads of old flies than in young at each time point tested (Figure 1A). Additionally, Ldh mRNA levels in the heads of old flies showed a diurnal rhythm with a peak at ZT12 (Figure 1A), in agreement with rhythmicity measured by RNA-seq [10]. To investigate changes in LDH protein levels, we performed immunoblot analysis of head extracts from young and old flies collected at the same time points as samples for Ldh mRNA. Due to the lack of a specific antibody against fly LDH, we screened several commercially available LDH antibodies by immunoblotting protein extracts derived from flies with elevated or reduced Ldh gene expression. We determined that the antibody against human LDHA (PA5-26531) reacted with fly LDH (Supporting Information Supplementary Figure 1). LDH protein levels were considerably higher in the heads of old flies at each time point, albeit without evidence of significant rhythmic changes (Figure 1B). Since the function of LDH is to catalyze the interconversion of pyruvate and lactate with concurrent changes in the NADH/NAD+ redox couple, we next determined LDH enzymatic activity for both reactions by measuring the increase or decrease in NADH over time for each reaction. In both age groups, the activity of LDH was higher for the pyruvate to lactate reaction compared to the conversion of lactate to pyruvate. Notably, the conversion of pyruvate to lactate was significantly increased in the heads of old compared to young, while the conversion of lactate to pyruvate remained similar between age groups (Figure 1C).
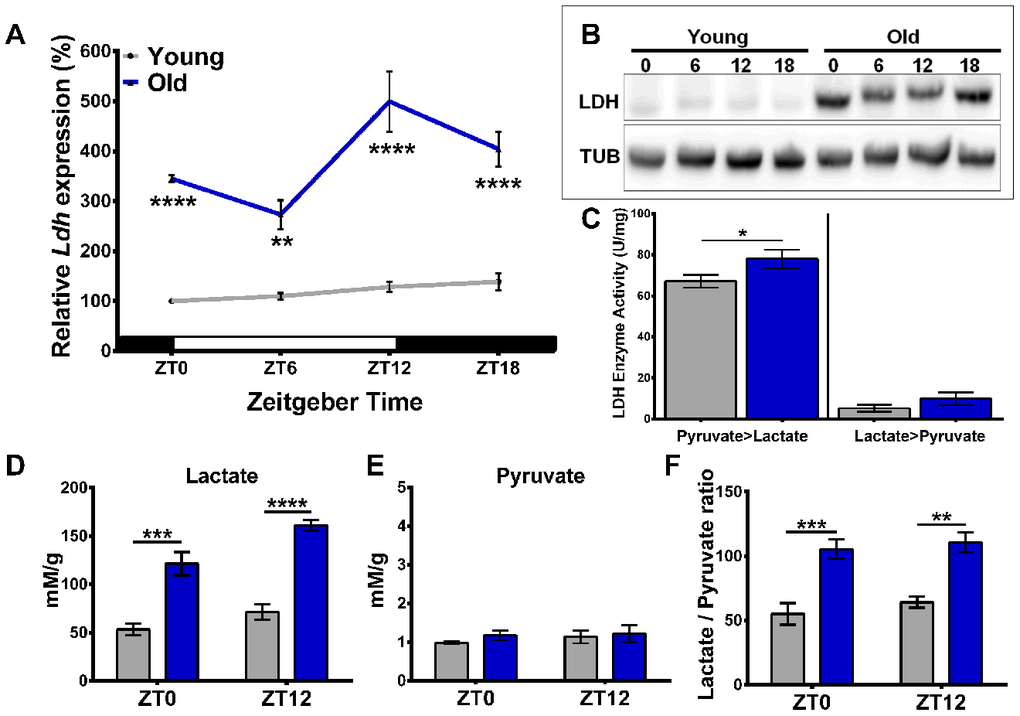
Figure 1. Age-related increase in Ldh expression is associated with elevated LDH protein levels, enzyme activity, and lactate concentration. (A) Profile of Ldh mRNA expression in the heads of young (5-days-old) and old (55-days-old) w1118 flies measured by qRT-PCR. Compared to young, old flies have increased Ldh mRNA levels at each time point with a peak at ZT12. Values are averages of 4 biorepeats reported as a percentage of expression relative to young at ZT0 set to 100%. (B) Representative western blot of LDH protein levels in young and old fly heads with tubulin as a loading control. (C) Graph showing LDH enzyme activity for both the pyruvate to lactate and lactate to pyruvate reactions in the heads of young (grey bars) and old (blue bars) flies. Enzymatic activity of LDH is higher in the heads of old flies compared to young for the pyruvate to lactate reaction (*p<0.05 by unpaired one-tailed t-test with Welch’s correction). (D) Lactate levels are significantly higher in the heads of old flies (blue bars) at both time points compared to young. In old flies, there is also a significant difference in lactate levels between ZT0 and ZT12 (n=4; p<0.001 by unpaired t-test with Welch’s correction). (E) Pyruvate levels do not significantly change between young (grey bars) and old (blue bars) (n=4). (F) Average lactate/pyruvate ratios increase in old flies (blue bars) (n=4). Error bars in A, C–F indicate standard error of the mean (SEM). Significance between age groups and time of day in each graph determined by 2-Way ANOVA with Bonferroni's correction (**p<0.01, ***p<0.001, and ****p<0.0001).
Given that LDH protein levels and enzymatic activity of the pyruvate to lactate reaction increase in the heads of old flies, we used 1D 1H nuclear magnetic resonance (NMR) to determine whether the concentration of lactate changes in fly heads as a function of age. Polar metabolites were extracted from heads of young and old flies collected at ZT0 and ZT12. Overall, we detected a significant age-dependent increase in lactate concentration at both time points examined (Figure 1D, Supporting Information Supplementary Figure 2). In addition, the level of lactate was significantly higher at ZT12 compared to ZT0 in old flies (Figure 1D) suggesting development of diurnal rhythm during aging, corresponding to the Ldh gene expression rhythm (Figure 1A). In contrast to lactate, pyruvate concentrations were not significantly different between the two age groups or time points (Figure 1E). Based on our NMR concentration data, we determined that the lactate to pyruvate ratios were significantly increased in the heads of old flies at both time points (Figure 1F).
Ldh overexpression shortens fly lifespan
To investigate whether the age-related increase in the expression of Ldh mRNA is protective or detrimental during Drosophila aging, we measured the lifespan of flies with genetically manipulated Ldh expression. Given the rhythmic expression of Ldh in old flies (Figure 1A), we first overexpressed Ldh in all circadian clock-containing cells using transgenic flies carrying the promoter region of the clock gene timeless fused to GAL4 (tim-GAL4); these flies were crossed to flies carrying UAS-Ldh construct (tim>Ldh) or to w1118 flies (tim>w) for control. Highly increased Ldh mRNA levels were detected in the heads of both 10-day and 55-day old tim>Ldh males relative to age-matched tim>w control flies (Figure 2A). We also detected significant increases in LDH enzyme activity in the heads of 55-day old tim>Ldh flies compared to tim>w for the pyruvate to lactate reaction (Figure 2B). Finally, NMR analysis revealed significantly increased lactate levels in the heads of 55-day old tim>Ldh flies (Figure 2C). Given that overexpression of Ldh increased lactate levels in fly heads compared to age-matched controls, we tested whether the rate of aging is affected in these flies. Overexpression of Ldh in all clock cells (a subset of neurons and glia) significantly reduced the median lifespan of tim>Ldh males by 17 days (20%) compared to tim>w control (Figure 2D).
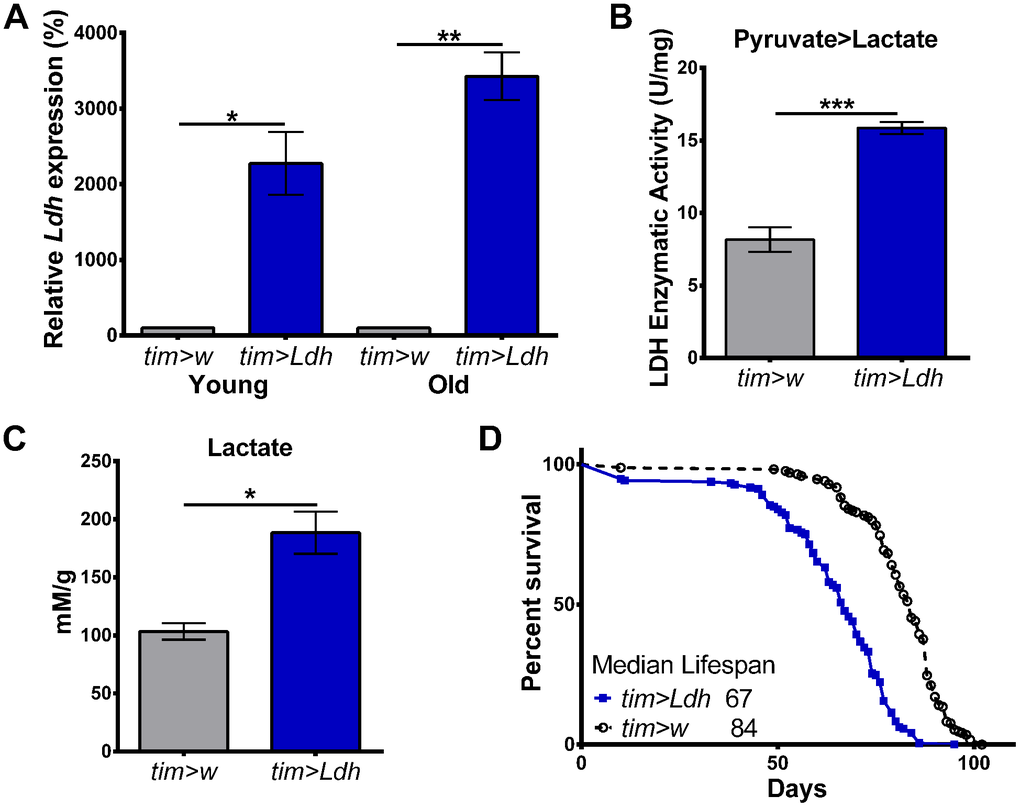
Figure 2. Flies overexpressing Ldh have increased mRNA levels, LDH activity, elevated lactate, and shortened lifespan. (A) Ldh mRNA levels were significantly increased in the heads of young and old tim>Ldh compared to age-matched tim>w control flies. Values are reported as a percentage of expression relative to age-matched tim>w set to 100% (n=4). (B) LDH enzyme activity was significantly increased for the pyruvate to lactate reaction in the heads of 55-day old tim>Ldh flies compared to tim>w controls (n=4). (C) Lactate concentrations were significantly higher in the heads of tim>Ldh flies compared to age-matched tim>w controls (n=4). Error bars in A-C indicate SEM. Statistical significance was determined by Unpaired t-test with Welch’s correction (***p<0.001; **p<0.01; *p<0.05). (D) Survival curves of tim>Ldh (n=193) and tim>w (n=170) males. Median lifespan was significantly reduced in tim>Ldh flies compared to control (Gehan-Breslow-Wilcoxon test; p<0.0001).
Since tim-GAL4 drives expression in a variety of cell types, including pacemaker neurons, photoreceptors, and most adult glial cells [25, 26], we also evaluated the effects of overexpressing Ldh selectively in neurons or glia. Ldh is expressed throughout fly development [27]; therefore, we employed the TARGET system [28] to manipulate expression only in adults. Cell-type specific GAL4 driver lines were combined with a tub-GAL80ts construct active in all cells, which blocks the action of GAL4 during development at 18°C. After eclosion, the flies were transferred to 27°C or 25°C, which inactivates GAL80ts to varying degrees, allowing GAL4 to induce the expression of the UAS-Ldh construct in all clock cells via tim-GAL4, all neurons via elav-GAL4, or all glia via repo-GAL4 (full genotypes are shown in Supporting Information Supplementary Table 1).
Flies overexpressing Ldh in all clock cells transferred from 18°C to 27°C after eclosion had significantly reduced median lifespan by 26% compared to timts>w control flies (Figure 3A). A less dramatic but significant lifespan reduction of 11% was observed in elavts>Ldh flies with pan-neuronal Ldh overexpression compared to elavts>w control flies (Figure 3B). Flies overexpressing Ldh in all adult glial cells had an 8.5% decrease in median lifespan compared to controls (Figure 3C). Given the abrupt mortality of timts>Ldh flies at 27°C (Figure 3A), we also recorded the lifespan of timts>Ldh, elavts>Ldh, repots>Ldh, and the respective control flies transferred to 25°C after eclosion. At 25°C, the lifespan of timts>Ldh flies was also significantly reduced relative to controls (Figure 3D) but less dramatically than in 27°C. Overexpression of Ldh in neurons reduced lifespan by 15% while glial overexpression reduced lifespan only by 8% when adult flies were maintained in 25°C (Figure 3E, 3F). The median and maximum lifespan data of all Ldh overexpressing flies and controls are provided in Supporting Information Supplementary Table 2.
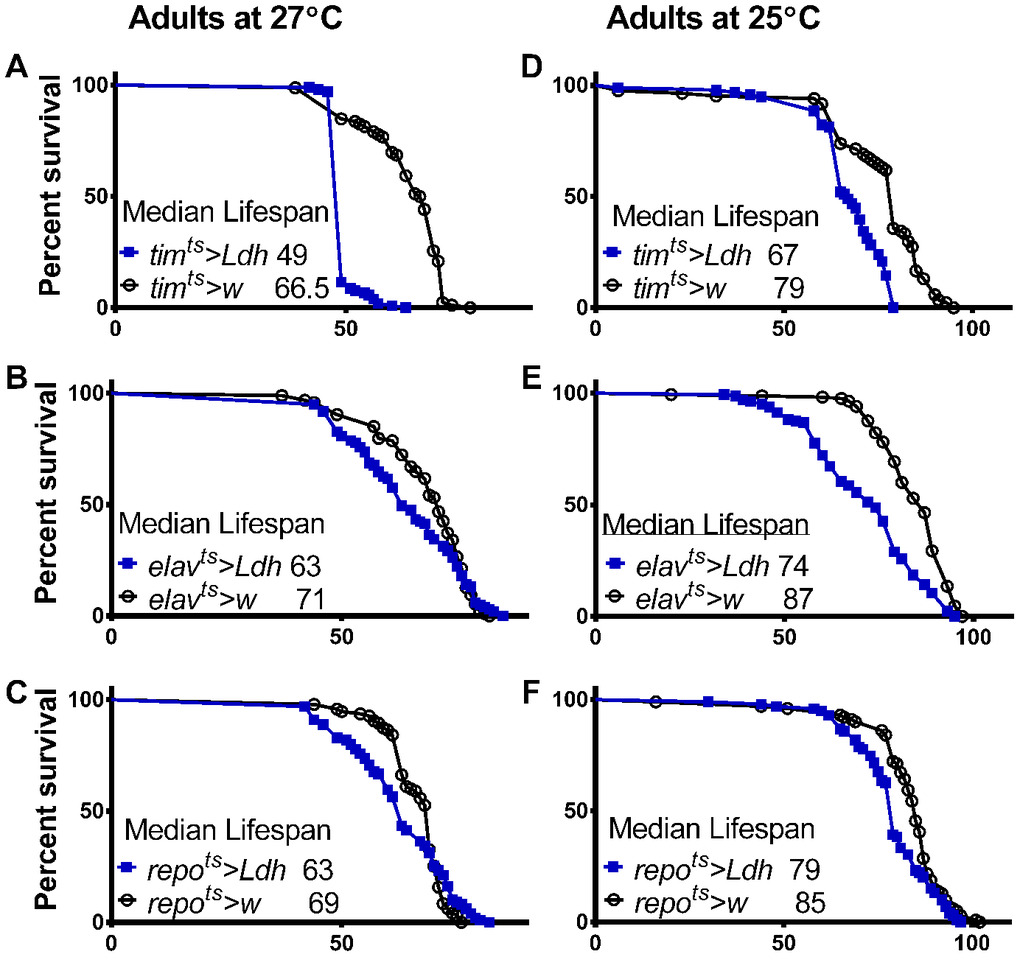
Figure 3. Adult specific overexpression of Ldh shortens fly lifespan. (A–F) Survival curves of adult males overexpressing Ldh in all clock cells via timts (A, D), in neurons via elavts (B, E), or in glia via repots (C, F) each graphed relative to their control group at 27°C and 25°C, respectively. Overexpression of Ldh via each driver resulted in decreased median lifespan compared to their respective controls in both 27°C and 25°C (Gehan-Breslow-Wilcoxon test; p<0.05 for graphs B and C and p<0.0001 for graphs A, D, E, and F). See Supporting Information Supplementary Table 2 for experimental details.
Overexpression of Ldh in neurons increases neurodegeneration and disrupts circadian locomotor activity rhythms
We hypothesized that reduced lifespan of flies with elevated Ldh gene expression could be linked to brain neurodegeneration. To test this, we evaluated the formation of vacuoles in the brains of elavts>Ldh, repots>Ldh, and their respective controls. Vacuoles are a reliable biological marker to measure the extent of neurodegeneration during normal aging or in fly models of neurodegenerative diseases [29–31]. At 55 days of age, the average number and area of vacuoles were significantly higher in elavts>Ldh brains than in age-matched elavts>w brains (Figure 4A–4D). In contrast to neuronal overexpression of Ldh, no significant differences were detected in the number or area of vacuoles in 55-day old repots>Ldh brains compared to repots>w brains (Figure 4E, 4F). Thus, neurodegenerative phenotypes seem to be specifically related to the elevated expression of Ldh in neurons, which also causes more dramatic lifespan reduction (see Figure 3E, 3F).
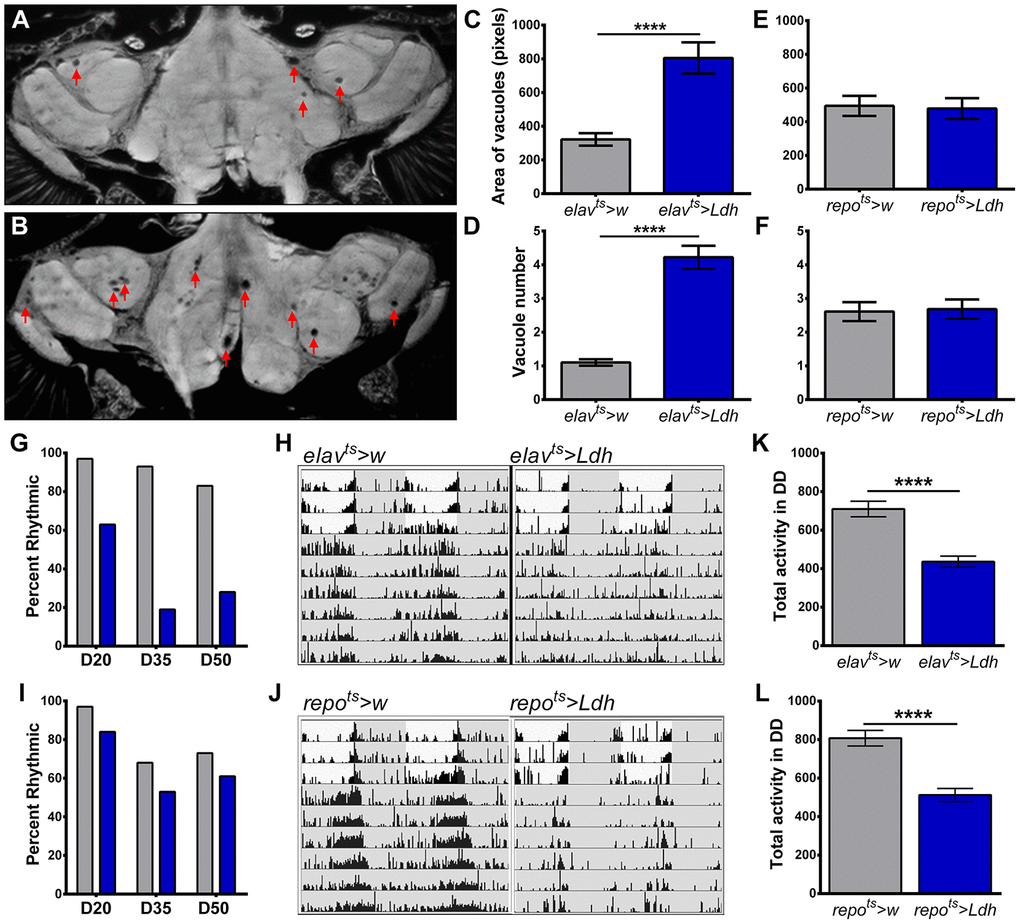
Figure 4. Flies with neuronal overexpression of Ldh show increased neurodegeneration and accelerated decline in locomotor activity rhythms. (A, B) Representative brain section images of 55-day old control elavts>w (A) and elavts>Ldh flies (B) in 25°C (arrows indicate vacuoles). (C, D) Graphs show the average area (C) and number (D) of vacuoles per brain in the brains of elavts>Ldh flies and age-matched controls. Both area and the number of vacuoles were significantly increased in elavts>Ldh (n=36) flies compared to elavts>w (n=38). ****p<0.0001; unpaired t-test with Welch’s correction. (E, F) There was no significant difference in the average area (E) and number (F) of vacuoles in 55-day old repots>Ldh (n=19) brains compared to age-matched repots>w control (n=18). Error bars indicate SEM. (G) Percent of rhythmic elavts>Ldh flies was markedly reduced with age compared to elavts>w controls. (H) Representative actrograms of individual 50-day old elavts>w (rhythmic) and elavts>Ldh (arrhythmic) flies. Gray areas indicate lights off. (I) Percentage of rhythmic repots>Ldh flies were similar to repots>w controls across lifespan. At least 30 flies were tested for each age group and each genotype. (J) Representative actograms of rhythmic 50-day old repots>w and repots>Ldh flies. (K) Total daily activity of 35-day old elavts>Ldh (n=31) flies averaged over six days in constant darkness was significantly reduced relative to control elavts>w (n=30) flies. (L) Total daily activity of 35 days-old repots>Ldh (n=30) flies was also significantly lower than in controls (n=31). Statistical significance by unpaired t-test with Welch’s correction (****p<0.0001). Error bars indicate SEM.
In addition to the increased risk of neurodegeneration, aging is associated with decline in locomotor activity rhythms [32, 33]. Therefore, we monitored the quality of circadian activity rhythms in elavts>Ldh, repots>Ldh, and their respective control at 20, 35, and 50 days of age. To assess endogenous circadian activity rhythms, we analyzed the robustness of rhythms over the course of 6 days in constant darkness (DD). Among 20-day old flies, 97% of elavts>w control flies and 63% of elavts>Ldh flies displayed rhythmic activity (Figure 4G). At 35 and 50 days of age, a high proportion of control flies remained rhythmic while less than 20% of elavts>Ldh flies displayed behavioral rhythmicity (Figure 4G, 4H). In contrast, repots>Ldh flies showed only modest decreases in the percentage of rhythmic flies in all age groups (Figure 4I, 4J). In addition to rest/activity rhythms, we tested the effects of Ldh overexpression on overall locomotor activity levels by recording the daily locomotor activity counts of 35-day old flies in DD. Compared to their respective controls, the average daily activity counts of both elavts>Ldh and repots>Ldh flies were significantly lower (Figure 4K, 4L). Taken together, these data suggest that accelerated loss of locomotor activity is a common feature of Ldh overexpression in both neurons and glia. However, neuronal Ldh overexpression also accelerates the loss of circadian activity rhythms while Ldh overexpression in glia had a modest effect on rhythmicity.
Reduced Ldh expression extends fly lifespan and delays neurodegeneration
Data showing that Ldh expression at the mRNA and protein level is elevated with age (Figure 1) and that overexpression of Ldh reduced lifespan (Figure 3) suggests that Ldh is involved in negative regulation of longevity. This hypothesis predicts that decreased Ldh expression during aging should enhance longevity. To test this, we used flies with reduced Ldh expression in clock cells, neurons, or glia, obtained by crossing the respective tub-GAL80ts containing GAL4 lines to flies carrying a UAS-LdhRNAi construct. For controls, the same tub-GAL80ts were crossed to w1118. Among flies held in 27°C as adults, lifespan was significantly extended in those with reduced Ldh expression in all clock cells, all neurons, or all glia compared to their respective controls (Figure 5A–5C). We also measured survivorship of flies housed at 25°C as adults and observed significant, but moderate increases in median lifespan in timts>LdhRNAi and repots>LdhRNAi flies compared to controls (Figure 5D and F, respectively), while considerable lifespan extension of 21% was recorded in elavts>LdhRNAi flies (Figure 5E). The median and maximum lifespan data of all LdhRNAi expressing flies and controls are provided in Supporting Information Supplementary Table 3.
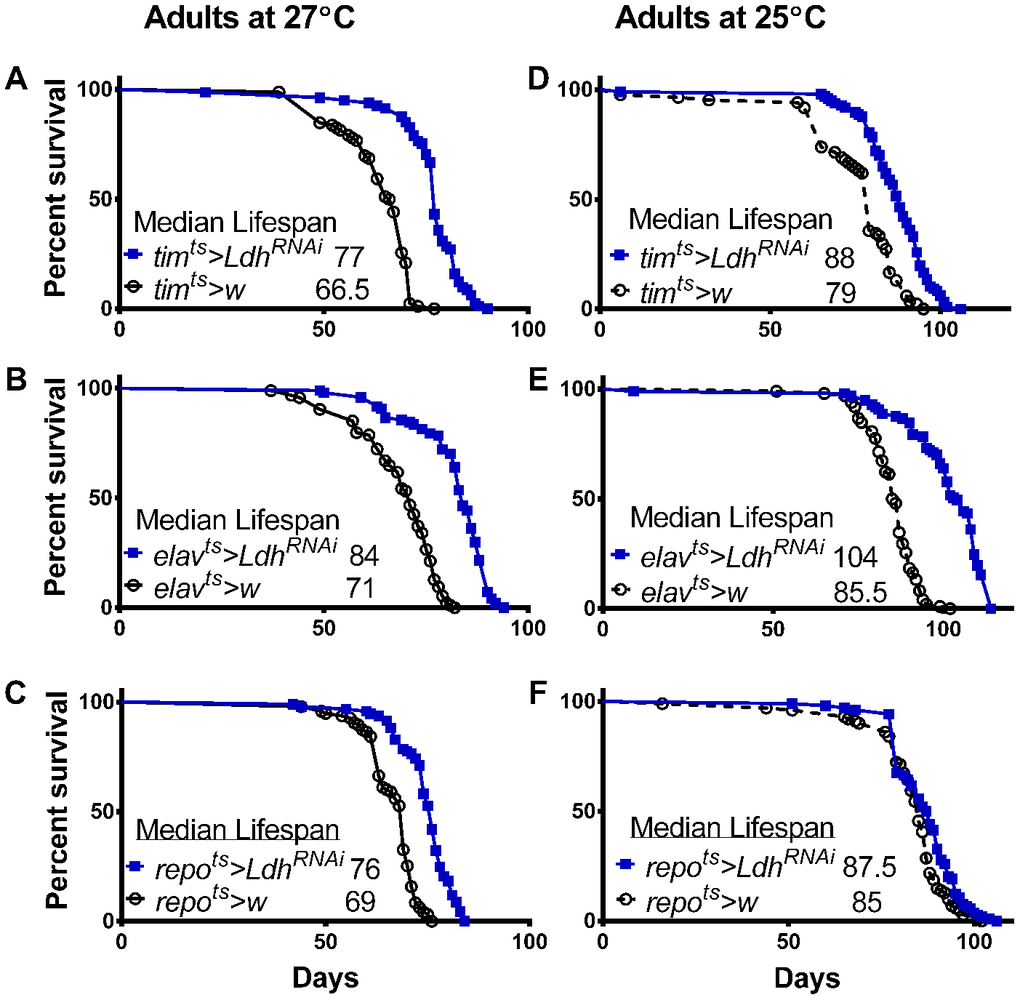
Figure 5. Decreased Ldh expression in adult brain extended lifespan. Survival curves of males with reduced Ldh expression via LdhRNAi combined with timts (A, D), elavts (B, E), or repots (C, F) and their controls at 27°C and 25°C, respectively. Ldh decrease significantly extends lifespan compared to controls (pairwise comparison by Gehan-Breslow-Wilcoxon test yielded p<0.0001 for all graphs except p<0.05 for repots> LdhRNAi versus control at 25°C). See Supporting Information Supplementary Table 3 for experimental details.
Given that flies with pan-neuronal knockdown of Ldh showed the most substantial lifespan extension, we asked whether reduced Ldh expression may have neuroprotective effects. To test this, we evaluated vacuolization in the brains of elavts>LdhRNAi flies using as controls progeny of elavts flies crossed to UAS-RNAicontrol flies, which have the same genetic background as LdhRNAi but carry no RNAi construct (Supporting Information Supplementary Table 1). First, we confirmed that the heads of 55-day old elavts>LdhRNAi flies had significantly reduced expression of Ldh mRNA compared to age-matched elavts>RNAicontrol (Figure 6A). Similar to the data shown in Figure 5E, the median lifespan of elavts>LdhRNAi flies was significantly increased compared to elavts>RNAicontrol flies (Figure 6B, Supporting Information Supplementary Table 3). To account for the extended lifespan of elavts>LdhRNAi flies and the fact that 55-day old elavts>w control flies show very few vacuoles per brain (Figure 4A); we tested brains of 75-day old flies that show more advanced neurodegeneration to facilitate observing differences between control and experimental flies. Compared to elavts>RNAicontrol, the brains of elavts>LdhRNAi flies had a significantly smaller number and area of vacuoles (Figure 6C–6F). Taken together, these data suggest that reducing Ldh expression in neurons may protect flies from age-associated neurodegeneration.
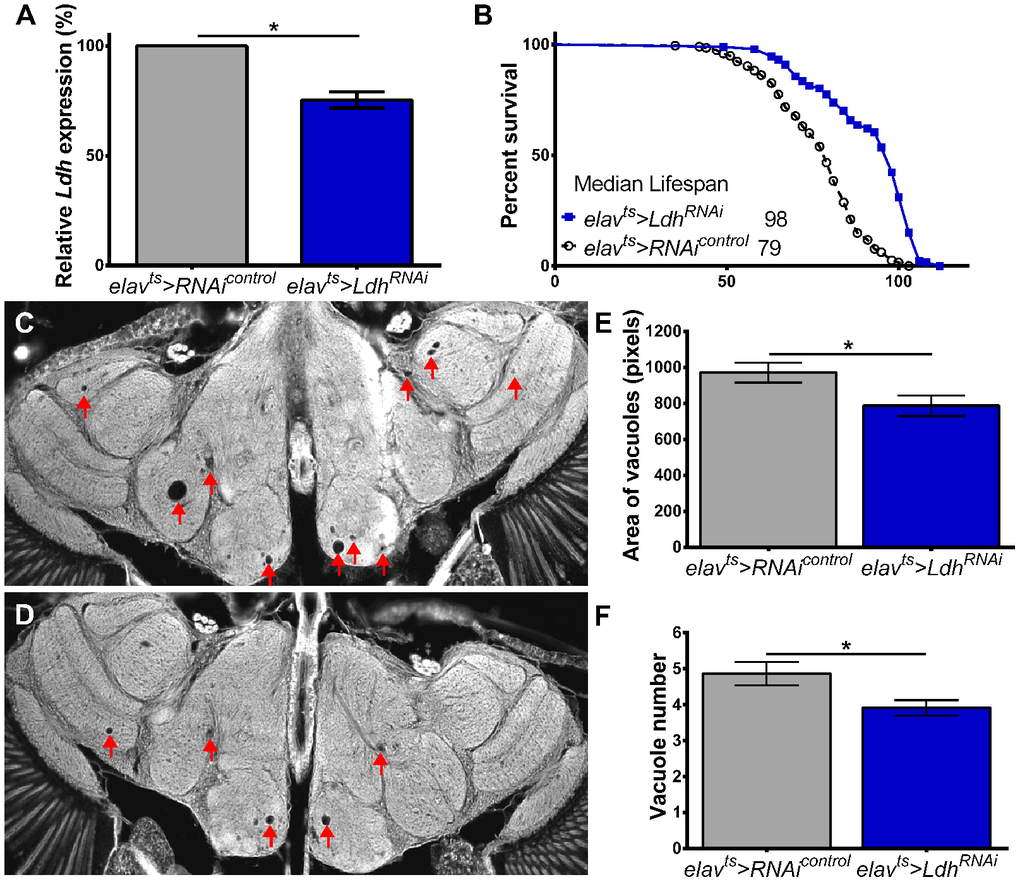
Figure 6. Neuronal knockdown of Ldh delays age-related neurodegeneration. (A) Ldh mRNA levels were significantly reduced in the heads of 55-day old elavts>LdhRNAi compared to age-matched elavts>RNAicontrol flies (n=3; Unpaired t-test with Welch’s correction *p<0.05). (B) Survival curves of elavts>LdhRNAi (n=187) and elavts>RNAicontrol (n=195). Median lifespan was significantly extended in elavts>LdhRNAi flies compared to control (Gehan-Breslow-Wilcoxon test; p<0.0001; see also Supplementary Table 3). (C, D) Representative brain section images of 75-day old (C) elavts>RNAicontrol and (D) elavts>LdhRNAi flies (arrowheads indicate vacuoles). (E, F) Graphs showing that the average area (E) and number (F) of vacuoles were significantly lower in the brains of 75-day old elavts>LdhRNAi flies (n=48) compared to controls (n=46). Statistical significance determined by unpaired t-test with Welch’s correction *p<0.05. Bars indicated SEM.
Discussion
Recent high-throughput techniques have demonstrated that aging correlates with many changes in gene expression and metabolite levels. However, it is often not clear whether these changes are protective to an aging organism or rather negatively affect healthspan and longevity. In this study, we first show that age-related increase in Ldh mRNA and protein levels are associated with elevated lactate levels in the heads of Drosophila melanogaster. We then demonstrate that adult-specific overexpression of Ldh significantly reduced fly lifespan and that neuronal overexpression increased brain neurodegeneration and accelerated loss of rest/activity rhythms compared to age-matched controls. In contrast, RNAi-mediated decrease in Ldh expression, especially in neurons, significantly extended lifespan and showed neuroprotective effects. Together, these data suggest that Ldh and lactate play causative roles in the modulation of aging and longevity in flies. We assume that these effects are related to late-life Ldh mRNA manipulations; however, because Ldh expression was altered through most of adult life, we cannot exclude that chronic changes in LDH enzyme activity could contribute to the observed phenotypes. Likewise, we cannot exclude the possibility that changes in ‘moonlighting’ functions of LDH upon UAS/GAL4 induced Ldh overexpression contribute to the phenotypes reported in these flies.
While adult-specific overexpression of Ldh in either neurons or glia significantly reduced fly longevity, only pan-neuronal Ldh overexpression caused a significant increase in brain neurodegeneration and decay of circadian rest/activity rhythms. This suggests that elevated Ldh levels in neurons have additional detrimental effects. This is supported by the fact that neuronal overexpression of Ldh shortened lifespan by 15% while glial overexpression shortened lifespan by only 8%.
We show that age-related increase in Ldh mRNA expression results in significantly elevated levels of LDH protein and lactate in fly heads. Given that Ldh mRNA is expressed in a diurnal rhythm in the heads of old flies, we measured lactate and pyruvate levels at ZT0 and ZT12. Indeed, in the heads of old flies, lactate levels were significantly higher at ZT12 than at ZT0, which is suggestive of diurnal rhythm. Diurnal rhythmicity in lactate levels have been reported in whole young flies [34], fly bodies [35], and human blood plasma [36]. Our data suggest that putative rhythm in lactate levels is age-dependent in fly heads, in agreement with rhythmic Ldh mRNA expression profile detected only in the heads of old flies.
It should be noted that a recent study of age-related metabolic changes in fly heads did not detect an increase in lactate [37]; however, lactate was measured in 35-day old flies and we reported that expression of Ldh is only moderately elevated at this age [10]. Our data showing increased lactate levels in the heads of old flies are consistent with the age-related increase in lactate reported in the brain of aging mice [13, 19]. Additionally, elevated lactate levels have been reported in the heads of AD model flies [21] and in an APP/PS1 AD model mice [38]. A significant increase in LDH enzyme activity or lactate was found in the brains of AD patients [20, 39, 40], and increased lactate levels have been suggested as a risk factor in amyloidogenesis related to AD [41]. In another study, elevated levels of lactate negatively correlated with memory performance in humans with mild cognitive impairments [42]. Aging is a well-known risk factor for AD, and it remains to be determined whether age-related increases in LDH and lactate could contribute to the development of AD-related pathologies.
While studies discussed above show links between increased lactate levels and neurodegenerative phenotypes, numerous experiments in young animals or in vitro suggest that lactate produced by astrocytes and transferred to neurons may be beneficial for processes that show decline during aging, such as memory formation, neuronal plasticity, and neurogenesis [16, 43]. In addition, a recent report showed that lactate produced by exercising muscles improves glucose tolerance by signaling to adipocytes [44]. Clearly, the reconciliation of reports showing positive and negative effects of lactate will require further investigations. Notably, a recent report suggested that the roles of lactate may change across lifespan as energy metabolism in neurons shifts toward glycolysis in an age-specific fashion [24]. We also note that caution should be used when comparing data obtained in mammals and flies as the ratio of glial cells to neurons increases approximately 6-fold between flies and humans suggesting an expanding role of glia as the complexity of the nervous system increases [45].
Lactate has been proposed as a biomarker of impaired mitochondrial function [42], and normal aging is associated with a decline in mitochondrial quality and activity [3]. Consistent with these metabolic trends, our recent RNA-seq data suggest that age-related increase in Ldh expression in fly heads is associated with decline in expression of pyruvate metabolism genes [10]. Following glycolysis, pyruvate is transferred into mitochondria via mitochondrial pyruvate carrier proteins (MPCs) and converted by pyruvate dehydrogenase complex (PDH) to acetyl-CoA, which enters the tricarboxylic acid (TCA) cycle. The activity of PDH is inhibited via phosphorylation by pyruvate dehydrogenase kinase (PDK). Importantly, our recent RNA-seq analysis revealed a significant age-related increase in the expression of the Pdk gene in the heads of old flies and a concomitant decrease in expression of the Mpc1 gene encoding mitochondrial pyruvate carrier [10]. Moreover, two recent transcriptomic studies of the aging fly brain revealed an overall decline in the expression of genes involved in mitochondrial oxidative phosphorylation [46, 47]. Taken together, these data provide evidence of decreased pyruvate uptake by age-damaged mitochondria and/or lowered conversion of pyruvate to acetyl-CoA in addition to decreased mitochondrial oxidative phosphorylation in the aging brain. These mitochondrial deficiencies, when combined with elevated Ldh expression, may contribute to increased lactate levels in cells now relying on glycolysis to meet their energetic needs.
Finally, it should be noted that LDH is highly upregulated in many cancers that undergo a metabolic shift towards aerobic glycolysis (Warburg effect) to reduce reactive oxygen species production [14]. This shift includes primary brain tumors; therefore, LDH inhibitors have already been suggested as a potential therapeutic target [14]. Our unbiased genetic approach demonstrates a causative relation between reduced neuronal Ldh gene expression and delayed neurodegeneration. Given the conservation of brain aging mechanisms from Drosophila to humans, our study highlights the possibility that interventions that reduce LDH activity and lactate levels could conceivably have neuroprotective effects in aging humans.
Materials and Methods
Fly rearing and genetics
Drosophila melanogaster were maintained on a diet containing yeast (35g/L), cornmeal (50g/L), and molasses (5%). Flies were maintained in a 12:12 hour light/dark cycles at 25°C. All experiments were performed on mated adult male flies. We used w1118 (BDSC #5905) flies to investigate changes in Ldh mRNA, LDH protein, enzyme activity, and polar metabolites in the heads of young (5 days) and old (55 days) flies. To manipulate the expression of Ldh in specific cell types, the Drosophila binary UAS/GAL4 system was used. tim>Ldh progeny were obtained by crossing tim-GAL4 [25] (gift from Jeff Hall) females to UAS-Ldh (FlyORF #F002924) males. Control flies were obtained by crossing tim-GAL4 females to w1118 males (tim>w). To increase spatiotemporal control and manipulate Ldh only in adults, we utilized the TARGET system [28]. Temperature-sensitive lines were used to drive expression in all circadian clock cells (tim-GAL4;tub-GAL80ts; gift from Patrick Emery), in all neurons (elav-GAL4;tub-GAL80ts), or all glia (tub-GAL80ts;repo-GAL4; gift from Rob Jackson); we refer to these lines as timts, elavts, and repots, respectively. Females carrying these temperature-sensitive drivers were crossed to UAS-Ldh males to obtain timts>Ldh, elavts>Ldh, and repots>Ldh flies (see Supporting Information Supplementary Table 1 for full genotypes). Flies with reduced Ldh expression in clock cells, neurons, or glia were obtained by crossing timts, elavts, and repots females to UAS-LdhRNAi (BDSC #33640) males. For controls, timts, elavts, and repots females were crossed to w1118 males. Additionally, elavts females were crossed to males with the genomic insertion of the LdhRNAi construct but lacking an RNAi coding sequence (BDSC #36303), which we call UAS-RNAicontrol (Supporting Information Supplementary Table 1). The progeny of these crosses were maintained in 18°C throughout development. Two to three days after eclosion, mated males were transferred to 25°C or 27°C as specified in results. Longevity was measured as described [48] for each genotype and temperature. Mortality curves were plotted using GraphPad Prism 6 (GraphPad Software Inc. San Diego, CA).
Quantitative reverse transcription polymerase chain reaction (qRT-PCR)
Young and old, 5 and 55 days of age respectively, w1118 flies were collected at 6-hour intervals over 24 hours in 12:12 LD. All other genotypes were collected at a single time point at the lights off transition (ZT12). Heads of 50 flies were separated using liquid nitrogen-cooled stainless steel sieves. Samples were homogenized in Trizol (Sigma-Aldrich, St Louis, Missouri) using a Kontes handheld pestle followed by rDNAse I (Takara, Japan) treatment, phenol/chloroform precipitation, and ethanol/sodium acetate precipitation as described [49]. Maxima First Strand cDNA synthesis kit (Thermo Scientific) was used to synthesize cDNA. Real-time PCR was performed with Power SYBR Green (Applied Biosystems) on a Step One Plus PCR system (Applied Biosystems). Primers had the following sequences: Dcp2 forward 5’ CCAAGGGCAAGATCAATGAG 3’, Dcp2 reverse 5’ GCATCGATTAGGTCCGTGAT 3’, Ldh forward 5’ CGTTTGGTCTGGAGTGAACA 3’, Ldh reverse 5’ GCAGCTCGTTCCACTTCTCT 3’. RNA levels were normalized to Dcp2 and analyzed by the 2-ΔΔCT method.
Analysis of neurodegeneration
Heads of 55-day old elavts>Ldh and elavts>w, or repots>Ldh and repots>w or 75-day old elavts>LdhRNAi and elavts>RNAicontrol mated males were randomly placed side by side in a collar, processed as one sample, embedded in the same paraffin block, and cut into 7μm sections. After removing the paraffin, the sections were embedded in Permount and analyzed by an experimenter blinded to the genotype. Due to the eye pigment, tissues are fluorescent, resulting in the vacuoles appearing in black. After examining all sections from a single head, an image was taken from the section that shows the most severe vacuolization phenotype. Vacuoles were identified, counted, and their surface area measured using the magic wand tool in Photoshop before the genotype was revealed. For a detailed description of the method and analysis including a video see [50].
Analysis of locomotor activity rhythms
Activity rhythms were measured as described [48, 49]. Briefly, 20-, 35-, and 50-day old males of the genotypes specified in the results section were placed in individual activity tubes and loaded into the TriKinetics Drosophila Activity Monitoring System (DAMS; TriKinetics, Waltham, MA). Data were collected with DAMS every 1 minute for three consecutive days in LD 12:12 followed by six days in constant darkness. Data were analyzed using Clocklab software (v.6.0.50, Actimetrics; Wilmette, IL) to generate actograms and periodograms. Chi-squared periodograms of individual flies were used to determine circadian rhythmic behavior in constant darkness. Flies with a periodogram amplitude peak near 24-hours breaking the 99% confidence line were deemed rhythmic. The average daily activity levels were calculated using the activity counts during the six consecutive days in constant darkness for each genotype.
Western blots
Groups of 20 whole heads were lysed, homogenized, and sonicated in an extraction buffer containing 50mM Tris, 2% SDS, 1mM EDTA, and protease inhibitors. Protein extracts were quantified using a colorimetric assay for protein concentration (DC Protein Assay; Bio-Rad), resolved in a 10% SDS-PAGE, and blotted onto a PVDF membrane (Bio-Rad). Membranes were probed with mouse monoclonal anti-β-tubulin (1:1000) (E7; Developmental Studies Hybridoma Bank) and polyclonal rabbit anti-LDHA (1:1000) (PA5-26531; ThermoFisher). This LDHA antibody was chosen based on its reactivity to Drosophila LDH protein using flies overexpressing Ldh (Supporting Information Supplementary Figure 1). Subsequently, membranes were probed with the appropriate horseradish peroxidase-conjugated secondary antibodies, goat anti-mouse AP130P (1:1000; Millipore) or goat anti-rabbit AP307P (1.5:1000; Millipore). Blots were imaged using Pierce ECL Western blotting substrate (ThermoFisher) in a Bio-Rad Molecular Imager (ChemiDoc XRS; Bio-Rad).
LDH enzymatic activity
LDH enzyme activity was determined by measuring the rate of interconversion between pyruvate and lactate and concurrent changes in the NAD+/NADH redox couple. Groups of 20-50 heads were lysed, homogenized, and sonicated in a buffer containing 100mM potassium phosphate, 2mM EDTA, and protease inhibitors. Protein concentration was determined using a colorimetric assay (DC Protein Assay), and 6μg of protein was loaded into 50μl reactions done in triplicate in a 96-well plate. The conversion of pyruvate to lactate results in the oxidation of NADH whereas the conversion of lactate to pyruvate results in the reduction of NAD+. Changes in NADH fluorescence were detected using a Tecan M1000 microplate reader with excitation/emission wavelengths of 340/460nm then plotted on a standard curve generated with known concentrations of NADH to determine NADH concentration at each time point. Pyruvate to lactate activity (decrease in NADH concentration over time) was measured in a reaction containing 1M sodium pyruvate (Sigma) and 6.4mM NADH (Sigma) in a 500mM potassium phosphate buffer. Lactate to pyruvate activity (increase in NADH concentration over time) was measured in a reaction containing 1M sodium lactate, 14.3mM NAD+, and 0.52mM hydrazine in a buffer containing 0.5M glycine and 2.5mM EDTA. Enzyme activity is reported in international units (U), as defined by the conversion of 1μmol of NADH/minute/mg protein.
Polar metabolite extraction and 1D 1H nuclear magnetic resonance (NMR)
Polar metabolites were extracted from the heads of 5- and 55-day old males using a modified 1:1:1 methanol/water/chloroform protocol [51, 52]. Groups of 200 flies were flash-frozen in liquid nitrogen; heads were separated in liquid nitrogen, lyophilized for 48 hours, and the dry weight of each sample was recorded. All reagents used in the following steps were chilled and kept on ice. Heads were pulverized and then homogenized in methanol/water mixture using a bead beater (Qiagen) with 5-7 ZR BashingBeads (Zymo Research). The remaining water and chloroform were added sequentially, followed by agitation and incubation on ice. Samples were centrifuged at 2000g for 5 minutes to separate the phases. The polar layer was transferred to new tubes, dried overnight in a speed-vac, and placed at -80°C. Pellets were rehydrated in 5mM sodium phosphate buffer with IS-2 Chenomx Internal Standard with DSS-d6 (Chenomx; Canada). Samples were loaded into 3mm NMR tubes (Bruker; Switzerland). NMR data were collected on an 800 MHz (18.8 T) Bruker Avance III HD NMR spectrometer (Bruker; Switzerland) equipped with a triple resonance cryogenic probe. 1D NOESY spectra with solvent pre-saturation were acquired at 25°C with a spectral window of 9615.385 Hz (12 ppm), the acquisition time of 4 seconds, recycle delay of 1 second, calibrated 90-degree pulse and 512 scans. The total acquisition time for each sample was ~45 minutes. Data were apodized with 0.5Hz exponential line broadening, zero-filled to twice the number of points, Fourier Transformed and phased using the Chenomx NMR Suite Processor program (Chenomx; Canada). Metabolites were identified and quantified using the Chenomx NMR Suite Profiler. Metabolite concentrations were normalized by the dry weight of the sample.
Author Contributions
D.M.L. and J.M.G. conceived and designed the project with input from DAH. D.M.L. conducted most of the experiments and analyzed the data. A.F. and R.C. performed and analyzed LDH Western blot and LDH enzymatic activity experiments. D.M.L. and P.N.R. performed and analyzed NMR experiments. D.K. performed and analyzed brain histology experiments. D.M.L. and J.M.G. wrote the manuscript with feedback from all authors.
Acknowledgments
We thank E. S. Chow for help with PCR and fly rearing, T. Nash, S. Fu, and J. Harwood for laboratory help, and M. Freitag for use of instruments in his lab. We thank J. Tennessen for performing pilot lactate measurements using GC-MS. We thank P. Emery, J. Hall, and R. Jackson for fly stocks. Other stocks were obtained from the Bloomington Drosophila Stock Center (NIH P40OD018537) and the TRiP at Harvard Medical School (NIH/NIGMS R01- GM084947). We acknowledge the support of the Oregon State University NMR Facility funded in part by the NIH, HEI Grant 1S10OD018518, and by the M. J. Murdock Charitable Trust grant #2014162.
Conflicts of Interest
The authors declare that there are no conflicts of interest.
Funding
This work was supported by the National Institute of Aging of NIH under award number R01 AG045830 and R56 AG062621 to J.M.G, R56 AG053460 and R01 AG061406 to DAH, and R21 AG052950 and AG055943 to DK.
References
- 1. Niccoli T, Partridge L. Ageing as a risk factor for disease. Curr Biol. 2012; 22:R741–52. https://doi.org/10.1016/j.cub.2012.07.024 [PubMed]
- 2. Grimm A, Eckert A. Brain aging and neurodegeneration: from a mitochondrial point of view. J Neurochem. 2017; 143:418–31. https://doi.org/10.1111/jnc.14037 [PubMed]
- 3. Mattson MP, Arumugam TV. Hallmarks of brain aging: adaptive and pathological modification by metabolic states. Cell Metab. 2018; 27:1176–99. https://doi.org/10.1016/j.cmet.2018.05.011 [PubMed]
- 4. Lu T, Pan Y, Kao SY, Li C, Kohane I, Chan J, Yankner BA. Gene regulation and DNA damage in the ageing human brain. Nature. 2004; 429:883–91. https://doi.org/10.1038/nature02661 [PubMed]
- 5. Bishop NA, Lu T, Yankner BA. Neural mechanisms of ageing and cognitive decline. Nature. 2010; 464:529–35. https://doi.org/10.1038/nature08983 [PubMed]
- 6. Zhang R, Lahens NF, Ballance HI, Hughes ME, Hogenesch JB. A circadian gene expression atlas in mammals: implications for biology and medicine. Proc Natl Acad Sci USA. 2014; 111:16219–24. https://doi.org/10.1073/pnas.1408886111 [PubMed]
- 7. Chen CY, Logan RW, Ma T, Lewis DA, Tseng GC, Sibille E, McClung CA. Effects of aging on circadian patterns of gene expression in the human prefrontal cortex. Proc Natl Acad Sci USA. 2016; 113:206–11. https://doi.org/10.1073/pnas.1508249112 [PubMed]
- 8. Solanas G, Peixoto FO, Perdiguero E, Jardí M, Ruiz-Bonilla V, Datta D, Symeonidi A, Castellanos A, Welz PS, Caballero JM, Sassone-Corsi P, Muñoz-Cánoves P, Benitah SA. Aged stem cells reprogram their daily rhythmic functions to adapt to stress. Cell. 2017; 170:678–92.e20. https://doi.org/10.1016/j.cell.2017.07.035 [PubMed]
- 9. Sato S, Solanas G, Peixoto FO, Bee L, Symeonidi A, Schmidt MS, Brenner C, Masri S, Benitah SA, Sassone-Corsi P. Circadian reprogramming in the liver identifies metabolic pathways of aging. Cell. 2017; 170:664–77.e11. https://doi.org/10.1016/j.cell.2017.07.042 [PubMed]
- 10. Kuintzle RC, Chow ES, Westby TN, Gvakharia BO, Giebultowicz JM, Hendrix DA. Circadian deep sequencing reveals stress-response genes that adopt robust rhythmic expression during aging. Nat Commun. 2017; 8:14529. https://doi.org/10.1038/ncomms14529 [PubMed]
- 11. Markert CL, Shaklee JB, Whitt GS. Evolution of a gene. Multiple genes for LDH isozymes provide a model of the evolution of gene structure, function and regulation. Science. 1975; 189:102–14. https://doi.org/10.1126/science.1138367 [PubMed]
- 12. Eventoff W, Rossmann MG, Taylor SS, Torff HJ, Meyer H, Keil W, Kiltz HH. Structural adaptations of lactate dehydrogenase isozymes. Proc Natl Acad Sci USA. 1977; 74:2677–81. https://doi.org/10.1073/pnas.74.7.2677 [PubMed]
- 13. Ross JM, Öberg J, Brené S, Coppotelli G, Terzioglu M, Pernold K, Goiny M, Sitnikov R, Kehr J, Trifunovic A, Larsson NG, Hoffer BJ, Olson L. High brain lactate is a hallmark of aging and caused by a shift in the lactate dehydrogenase A/B ratio. Proc Natl Acad Sci USA. 2010; 107:20087–92. https://doi.org/10.1073/pnas.1008189107 [PubMed]
- 14. Valvona CJ, Fillmore HL, Nunn PB, Pilkington GJ. The regulation and function of lactate dehydrogenase A: therapeutic potential in brain tumor. Brain Pathol. 2016; 26:3–17. https://doi.org/10.1111/bpa.12299 [PubMed]
- 15. Abu-Shumays RL, Fristrom JW. IMP-L3, a 20-hydroxyecdysone-responsive gene encodes Drosophila lactate dehydrogenase: structural characterization and developmental studies. Dev Genet. 1997; 20:11–22. https://doi.org/10.1002/(SICI)1520-6408(1997)20:1<11::AID-DVG2>3.0.CO;2-C [PubMed]
- 16. Suzuki A, Stern SA, Bozdagi O, Huntley GW, Walker RH, Magistretti PJ, Alberini CM. Astrocyte-neuron lactate transport is required for long-term memory formation. Cell. 2011; 144:810–23. https://doi.org/10.1016/j.cell.2011.02.018 [PubMed]
- 17. Magistretti PJ, Allaman I. Lactate in the brain: from metabolic end-product to signalling molecule. Nat Rev Neurosci. 2018; 19:235–49. https://doi.org/10.1038/nrn.2018.19 [PubMed]
- 18. Saraswathy S, Crawford FW, Lamborn KR, Pirzkall A, Chang S, Cha S, Nelson SJ. Evaluation of MR markers that predict survival in patients with newly diagnosed GBM prior to adjuvant therapy. J Neurooncol. 2009; 91:69–81. https://doi.org/10.1007/s11060-008-9685-3 [PubMed]
- 19. Datta S, Chakrabarti N. Age related rise in lactate and its correlation with lactate dehydrogenase (LDH) status in post-mitochondrial fractions isolated from different regions of brain in mice. Neurochem Int. 2018; 118:23–33. https://doi.org/10.1016/j.neuint.2018.04.007 [PubMed]
- 20. Mullins R, Reiter D, Kapogiannis D. Magnetic resonance spectroscopy reveals abnormalities of glucose metabolism in the Alzheimer’s brain. Ann Clin Transl Neurol. 2018; 5:262–72. https://doi.org/10.1002/acn3.530 [PubMed]
- 21. Ott S, Vishnivetskaya A, Malmendal A, Crowther DC. Metabolic changes may precede proteostatic dysfunction in a drosophila model of amyloid beta peptide toxicity. Neurobiol Aging. 2016; 41:39–52. https://doi.org/10.1016/j.neurobiolaging.2016.01.009 [PubMed]
- 22. Dienel GA. Brain lactate metabolism: the discoveries and the controversies. J Cereb Blood Flow Metab. 2012; 32:1107–38. https://doi.org/10.1038/jcbfm.2011.175 [PubMed]
- 23. Díaz-García CM, Mongeon R, Lahmann C, Koveal D, Zucker H, Yellen G. Neuronal stimulation triggers neuronal glycolysis and not lactate uptake. Cell Metab. 2017; 26:361–74.e4. https://doi.org/10.1016/j.cmet.2017.06.021 [PubMed]
- 24. Drulis-Fajdasz D, Gizak A, Wójtowicz T, Wiśniewski JR, Rakus D. Aging-associated changes in hippocampal glycogen metabolism in mice. Evidence for and against astrocyte-to-neuron lactate shuttle. Glia. 2018; 66:1481–95. https://doi.org/10.1002/glia.23319 [PubMed]
- 25. Kaneko M, Hall JC. Neuroanatomy of cells expressing clock genes in Drosophila: transgenic manipulation of the period and timeless genes to mark the perikarya of circadian pacemaker neurons and their projections. J Comp Neurol. 2000; 422:66–94. https://doi.org/10.1002/(sici)1096-9861(20000619)422:1<66::aid-cne5>3.0.co;2-2 [PubMed]
- 26. Long DM, Giebultowicz JM. Age-related changes in the expression of the circadian clock protein PERIOD in Drosophila glial cells. Front Physiol. 2018; 8:1131. https://doi.org/10.3389/fphys.2017.01131 [PubMed]
- 27. Li H, Chawla G, Hurlburt AJ, Sterrett MC, Zaslaver O, Cox J, Karty JA, Rosebrock AP, Caudy AA, Tennessen JM. Drosophila larvae synthesize the putative oncometabolite L-2-hydroxyglutarate during normal developmental growth. Proc Natl Acad Sci USA. 2017; 114:1353–58. https://doi.org/10.1073/pnas.1614102114 [PubMed]
- 28. McGuire SE, Le PT, Osborn AJ, Matsumoto K, Davis RL. Spatiotemporal rescue of memory dysfunction in Drosophila. Science. 2003; 302:1765–68. https://doi.org/10.1126/science.1089035 [PubMed]
- 29. Sakakibara Y, Sekiya M, Fujisaki N, Quan X, Iijima KM. Knockdown of wfs1, a fly homolog of wolfram syndrome 1, in the nervous system increases susceptibility to age- and stress-induced neuronal dysfunction and degeneration in Drosophila. PLoS Genet. 2018; 14:e1007196. https://doi.org/10.1371/journal.pgen.1007196 [PubMed]
- 30. Etchegaray JI, Elguero EJ, Tran JA, Sinatra V, Feany MB, McCall K. Defective phagocytic corpse processing results in neurodegeneration and can be rescued by TORC1 activation. J Neurosci. 2016; 36:3170–83. https://doi.org/10.1523/JNEUROSCI.1912-15.2016 [PubMed]
- 31. Burman JL, Itsara LS, Kayser EB, Suthammarak W, Wang AM, Kaeberlein M, Sedensky MM, Morgan PG, Pallanck LJ. A Drosophila model of mitochondrial disease caused by a complex I mutation that uncouples proton pumping from electron transfer. Dis Model Mech. 2014; 7:1165–74. https://doi.org/10.1242/dmm.015321 [PubMed]
- 32. Koh K, Evans JM, Hendricks JC, Sehgal A. A Drosophila model for age-associated changes in sleep:wake cycles. Proc Natl Acad Sci USA. 2006; 103:13843–47. https://doi.org/10.1073/pnas.0605903103 [PubMed]
- 33. Mattis J, Sehgal A. Circadian rhythms, sleep, and disorders of aging. Trends Endocrinol Metab. 2016; 27:192–203. https://doi.org/10.1016/j.tem.2016.02.003 [PubMed]
- 34. Gogna N, Singh VJ, Sheeba V, Dorai K. NMR-based investigation of the drosophila melanogaster metabolome under the influence of daily cycles of light and temperature. Mol Biosyst. 2015; 11:3305–15. https://doi.org/10.1039/c5mb00386e [PubMed]
- 35. Rhoades SD, Nayak K, Zhang SL, Sehgal A, Weljie AM. Circadian- and light-driven metabolic rhythms in Drosophila melanogaster. J Biol Rhythms. 2018; 33:126–36. https://doi.org/10.1177/0748730417753003 [PubMed]
- 36. Dallmann R, Viola AU, Tarokh L, Cajochen C, Brown SA. The human circadian metabolome. Proc Natl Acad Sci USA. 2012; 109:2625–29. https://doi.org/10.1073/pnas.1114410109 [PubMed]
- 37. Wang L, Davis SS, Borch Jensen M, Rodriguez-Fernandez IA, Apaydin C, Juhasz G, Gibson BW, Schilling B, Ramanathan A, Ghaemmaghami S, Jasper H. JNK modifies neuronal metabolism to promote proteostasis and longevity. Aging Cell. 2019; 18:e12849. https://doi.org/10.1111/acel.12849 [PubMed]
- 38. Harris RA, Tindale L, Lone A, Singh O, Macauley SL, Stanley M, Holtzman DM, Bartha R, Cumming RC. Aerobic glycolysis in the frontal cortex correlates with memory performance in wild-type mice but not the APP/PS1 mouse model of cerebral amyloidosis. J Neurosci. 2016; 36:1871–78. https://doi.org/10.1523/JNEUROSCI.3131-15.2016 [PubMed]
- 39. Bigl M, Brückner MK, Arendt T, Bigl V, Eschrich K. Activities of key glycolytic enzymes in the brains of patients with Alzheimer’s disease. J Neural Transm (Vienna). 1999; 106:499–511. https://doi.org/10.1007/s007020050174 [PubMed]
- 40. Xu J, Begley P, Church SJ, Patassini S, Hollywood KA, Jüllig M, Curtis MA, Waldvogel HJ, Faull RL, Unwin RD, Cooper GJ. Graded perturbations of metabolism in multiple regions of human brain in Alzheimer’s disease: snapshot of a pervasive metabolic disorder. Biochim Biophys Acta. 2016; 1862:1084–92. https://doi.org/10.1016/j.bbadis.2016.03.001 [PubMed]
- 41. Xiang Y, Xu G, Weigel-Van Aken KA. Lactic acid induces aberrant amyloid precursor protein processing by promoting its interaction with endoplasmic reticulum chaperone proteins. PLoS One. 2010; 5:e13820. https://doi.org/10.1371/journal.pone.0013820 [PubMed]
- 42. Weaver KE, Richards TL, Logsdon RG, McGough EL, Minoshima S, Aylward EH, Kleinhans NM, Grabowski TJ, McCurry SM, Teri L. Posterior cingulate lactate as a metabolic biomarker in amnestic mild cognitive impairment. Biomed Res Int. 2015; 2015:610605. https://doi.org/10.1155/2015/610605 [PubMed]
- 43. Steinman MQ, Gao V, Alberini CM. The role of lactate-mediated metabolic coupling between astrocytes and neurons in long-term memory formation. Front Integr Neurosci. 2016; 10:10. https://doi.org/10.3389/fnint.2016.00010 [PubMed]
- 44. Takahashi H, Alves CR, Stanford KI, Middelbeek RJ, Nigro P, Ryan RE, Xue R, Sakaguchi M, Lynes MD, So K, Mul JD, Lee MY, Balan E, et al. TGF-β2 is an exercise-induced adipokine that regulates glucose and fatty acid metabolism. Nat Metab. 2019; 1:291–303. https://doi.org/10.1038/s42255-018-0030-7 [PubMed]
- 45. Kremer MC, Jung C, Batelli S, Rubin GM, Gaul U. The glia of the adult Drosophila nervous system. Glia. 2017; 65:606–38. https://doi.org/10.1002/glia.23115 [PubMed]
- 46. Davie K, Janssens J, Koldere D, De Waegeneer M, Pech U, Kreft Ł, Aibar S, Makhzami S, Christiaens V, Bravo González-Blas C, Poovathingal S, Hulselmans G, Spanier KI, et al. A single-cell transcriptome atlas of the aging Drosophila brain. Cell. 2018; 174:982–98.e20. https://doi.org/10.1016/j.cell.2018.05.057 [PubMed]
- 47. Pacifico R, MacMullen CM, Walkinshaw E, Zhang X, Davis RL. Brain transcriptome changes in the aging Drosophila melanogaster accompany olfactory memory performance deficits. PLoS One. 2018; 13:e0209405. https://doi.org/10.1371/journal.pone.0209405 [PubMed]
- 48. Long DM, Blake MR, Dutta S, Holbrook SD, Kotwica-Rolinska J, Kretzschmar D, Giebultowicz JM. Relationships between the circadian system and alzheimer’s disease-like symptoms in Drosophila. PLoS One. 2014; 9:e106068. https://doi.org/10.1371/journal.pone.0106068 [PubMed]
- 49. Chow ES, Long DM, Giebultowicz JM. Circadian rhythm in mRNA expression of the glutathione synthesis gene Gclc is controlled by peripheral glial clocks in Drosophila melanogaster. Physiol Entomol. 2016; 41:369–77. https://doi.org/10.1111/phen.12164 [PubMed]
- 50. Sunderhaus ER, Kretzschmar D. Mass histology to quantify neurodegeneration in Drosophila. J Vis Exp. 2016; 54809. https://doi.org/10.3791/54809 [PubMed]
- 51. Jensen K, Sanchez-Garcia J, Williams C, Khare S, Mathur K, Graze RM, Hahn DA, McIntyre LM, Rincon-Limas DE, Fernandez-Funez P. Purification of transcripts and metabolites from Drosophila heads. J Vis Exp. 2013; e50245. https://doi.org/10.3791/50245 [PubMed]
- 52. Ha PT, Renslow RS, Atci E, Reardon PN, Lindemann SR, Fredrickson JK, Call DR, Beyenal H. Regulation of electron transfer processes affects phototrophic mat structure and activity. Front Microbiol. 2015; 6:909. https://doi.org/10.3389/fmicb.2015.00909 [PubMed]