



Introduction
Every living being undergoes aging. The process of aging affects the brain size, vasculature, cognition [1], and genes, through a constant change in structure from birth throughout the lifetime. In addition, loss of memory also occurs with aging, and brain activation becomes more two aspects for memory tasks [2].
In the present study, we focused on a few genes associated with the loss of memory, brain derived neurotrophic factor (Bdnf), interleukin 6 (IL-6), apolipoprotein E (ApoE), and intercellular adhesion molecule (Icam). ApoE is an important gene for normal brain lipid homeostasis [3, 4] and is involved in facilitating the availability of cholesterol for development, maintenance and repair of myelin sheaths, nerve membranes, and synaptic connections [5]. Bdnf is well known marker for play an important role in long-term potentiation (LTP), a key factor in synaptic plasticity [6, 7]. Cognitive and motivational systems may be affected by neurotoxicity from increased IL-6 expression in the elderly brain [8]. This can also cause Alzheimer’s disease [9]. In addition, increasing age is associated with memory impairment and memory-related dementia; levels of neurotransmitters such as dopamine, noradrenaline, and adrenaline are also altered [2]. For example, dopamine levels have been reported to decrease approximately 10% per decade from early adulthood and have been associated with decreased cognitive function and motor performance [10, 11].
Cuminaldehyde (CA) (Figure 1) is an aromatic monoterpenoid volatile compound. It is a natural p-isopropylbenzaldehyde, which is one of the activate compound of the essential oil obtained from eucalyptus, myrrh [12], caraway (Carumcarvi) [13, 14], Chinese cinnamon (Cinnamomum cassia), and others [13]. CA also has been reported as the major component of cumin (Cuminum cyminum), a medicinal plant whose seeds are commonly used as a spice in different cuisines. Structurally, CA is a benzaldehyde substituted at the 4th position with an isopropyl group. It is used commercially in perfumes and cosmetics, because it has a pleasant aroma. It has been reported that CA has antidiabetic [15], antitumor [16], anti-inflammatory [17], antimicrobial, and antifungal [18] effects. Recently, studies have shown CA to exert protective effects against neurodegenerative diseases, in particular Parkinson’s disease [19]. However, despite showing great therapeutic potential for the treatment of aging and memory loss in neurodegenerative diseases, no studies have been directed on these effects of CA so far. Thus, the aim of our study is to evaluate the effect of CA on aging, especially on spatial learning and memory using the human-derived neurotrophic SH-SY5Y cells and aged C57BL/6J mice. We also studied the change in expression levels of the associated genes, Bdnf, IL-6, ApoE, and Icam in mice brain.
Figure 1. Structure of cuminaldehyde (CA).
Results
Evaluation of the cytotoxicity of cuminaldehyde on human neurotypic SH-SY5Y cells
First of all, we assessed the effect of CA on SH-SY5Y cells viability using MTT assay. Cells were treated with 2.5, 5, 10, 25, or 50 μM of CA for 72 h (Figure 2). The exposure of the cells to CA for 72 h showed no significant changes in the percentage of the cell viability compared to the non-treated cells (control group) (P > 0.05). Therefore, CA showed no cytotoxicity on SH-SY5Y cells at varying concentrations.
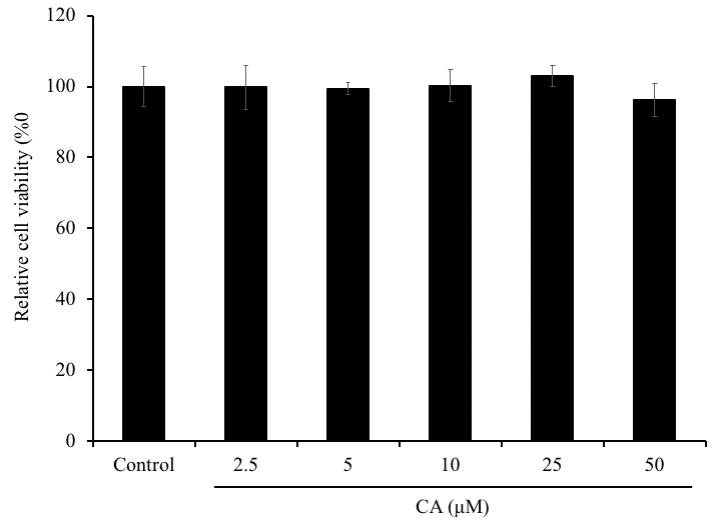
Figure 2. Effect of CA on SH-SY5Y cells viability. SH-SY5Y cells were seeded onto 96-well plate at a density of 2×105 cells/well. After overnight incubation, cells were treated with 5, 50 or 100 μM of CA for 72 h and the cell viability was evaluated using MTT assay. The experiment was repeated thrice. The bars signify relative viability as compared to the control. Data is represented as mean ± SD.
Evaluation of the neuroprotective effect of cuminaldehyde against dexamethasone-induced neurotoxicity on SH-SY5Y cells
SH-SY5Y cells were treated with several concentrations (5, 25, 50, 200, or 500 μM) of dexamethasone (Dexa), to determine the concentration that would be toxic for cells. Dexa-treated cells showed the significantly decrease in the cell viability in a dose-dependent manner, with a decrease of cell viability up to 55% at 500 μM comparing with the non-treated cells (P < 0.001) (Figure 3A). Therefore, the concentration 500 μM of Dexa was used to induce cell cytotoxicity in the further experiments. Then to estimate the effect of CA neurons, SH-SY5Y cells were treated with CA and/or co-treated with Dexa for 72h. The results showed that cell incubated with CA of 10, 25, or 50 μM reversed the Dexa-induced neuronal cell death (Figure 3B), and the cell viability was considerably increased by more than 20% as compared to only Dexa treated cells (P < 0.01).
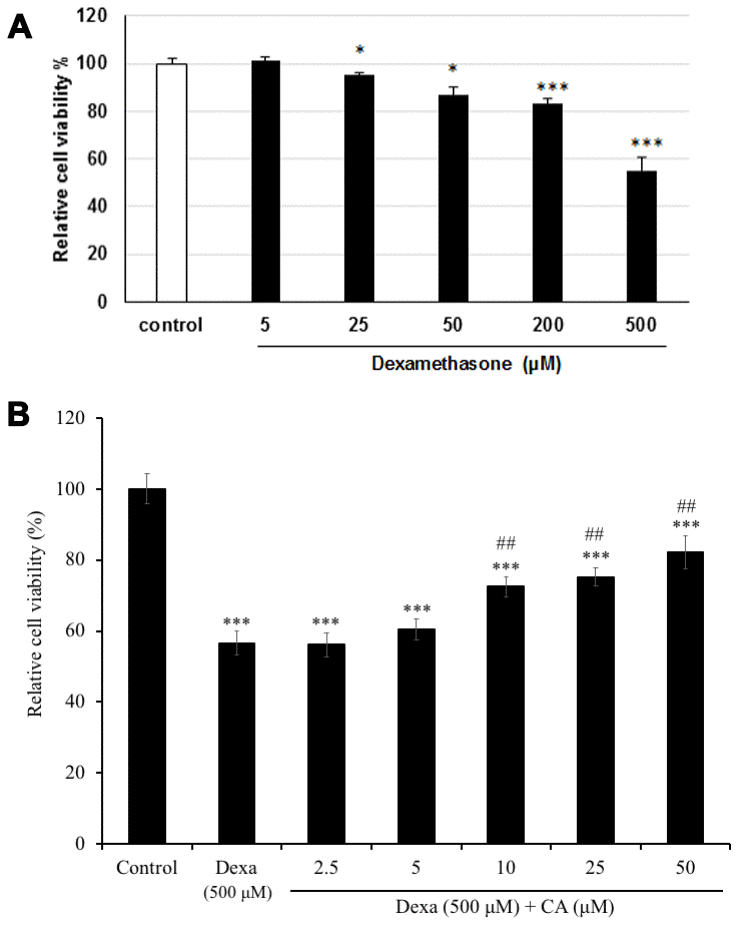
Figure 3. Effect on cell viability on SH-SY5Y cells after treatment with (A) dexamethasone (Dexa) and (B) Dexa co-treated with CA. SH-SY5Y cells were treated with CA or 500 μM Dexa for 72 h. Each bar represents the mean ± SD. ***: p < 0.001 as compared to the control; ##: p < 0.01; ###: p < 0.001 as compared to Dexa treatment.
Effects of sub-chromic administration of cuminaldehyde on body weight and brain weight of C57BL/6J mice
CA (25 mg/kg) was administered orally daily for 30 days to C57BL/6J mice, and the effect on body weight was studied. The results obtained showed that the body weight of CA treated mice did not differ from the young water-treated mice throughout the experiment (P > 0.05) (Figure 4A), no mortality and no changes in behavioral activity were recorded.
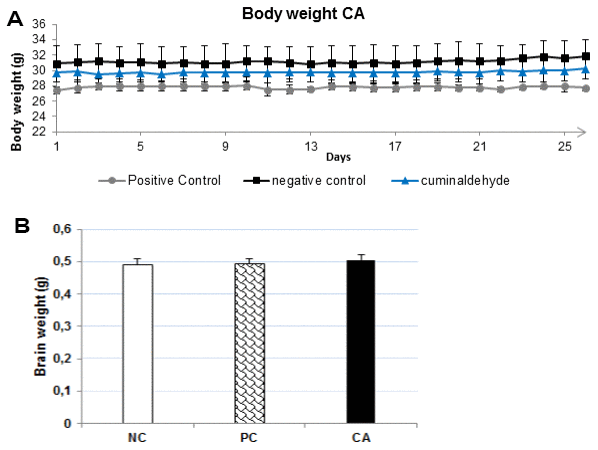
Figure 4. (A) Effects of CA (25 mg/kg) on body weight during the 30 days experimental period. All mice were fed a normal diet. The body weight was measured every day. Each point presents the values from that group for that day. The values are represented as mean ± standard deviation. (B) Brain weight after 30 days of treatment with CA. The brain was dissected out, washed with ice-cold PBS and weighed. All the values are expressed as mean ± SD. NC: negative control (aged water-treated group); PC: positive control (young water-treated group); CA: Cuminaldehyde treated group.
At the end of probe test of MWM, all mice were sacrificed by cervical dislocation and the brain was dissected and weighed. The results showed that CA-treated mice brain weight (Figure 4B) did not differ from those of young-water-treated mice (positive control) or aged-water-treated mice (negative control). CA did not produce any significant effect on the mice body and brain weights after daily administration for 30 days. This suggests that CA may not have any toxic effect in C57BL/6J mice.
Effect of cuminaldehyde on spatial learning and memory in C57BL/6J mice
To analyze effect CA on the ability of spatial learning and memory in aging C57BL/6J mice, the morris water maze (MWM) test was performed (Figure 5). The time (in seconds) taken by the animal to reach the platform indicates how animal can learn and memorize the platform location. In this experiment, we observed the decrease of escape latency compared to water-treated aging C57BL/6J mice indicating an improvement of spatial learning and memory ability. The escape latency time of both, aged C57BL/6J mice administered with CA (n = 8) and young water-treated C57BL/6J mice (positive control) was significantly decreased by 35% and 31% respectively (P < 0.05) as compared to the water-treated aging C57BL/6J mice (negative control) after 4 days of training session (Figures 4A, 5A). After 7 days of training, the time taken by aged-CA-treated mice (CA group) and young water-treated mice (positive control) significantly decreased by 20% compared to aged water-treated C57BL/6J mice (negative control). On the other hand, there was no significant difference between the escape latency time of aged-CA-treated mice (CA group) and young water-treated mice (positive control).
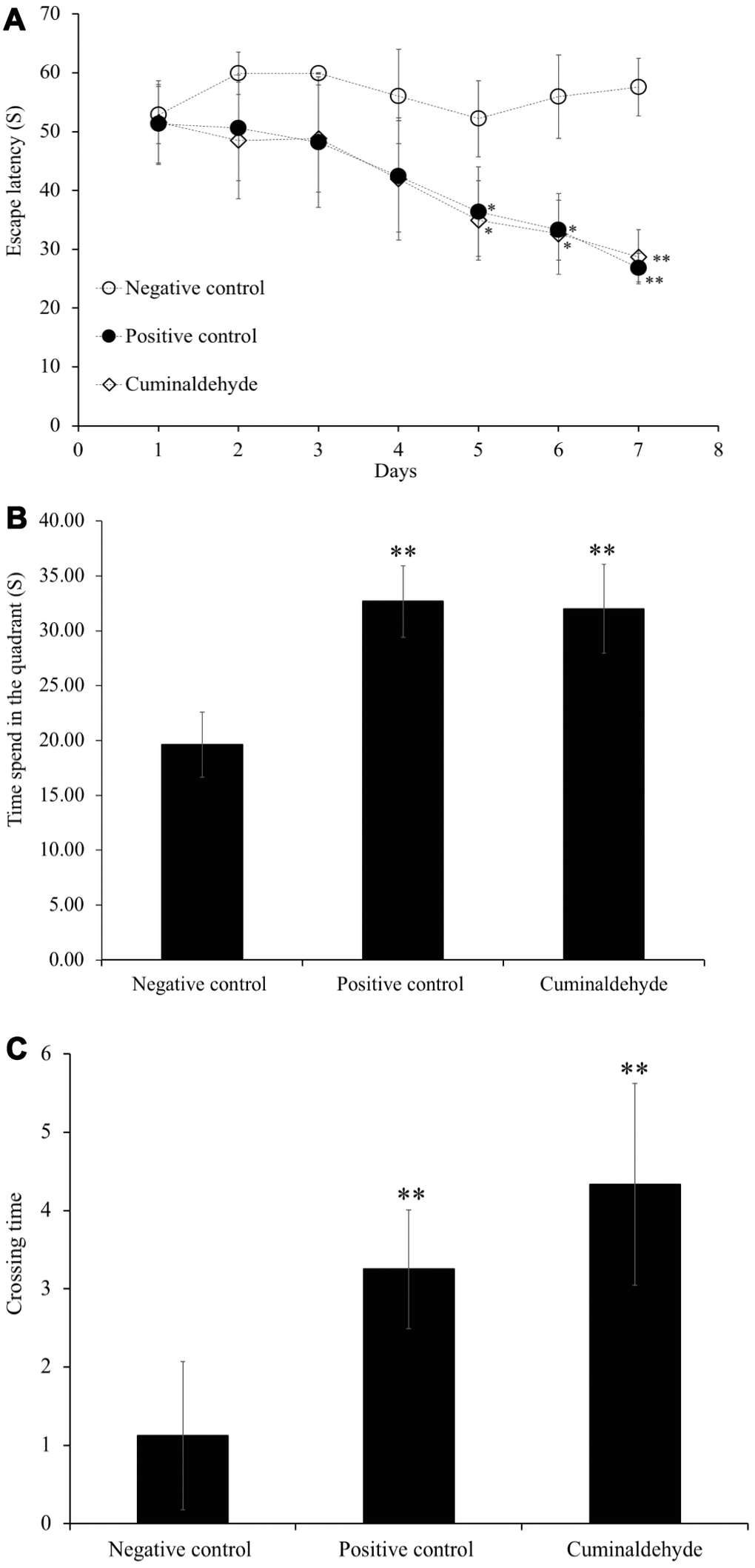
Figure 5. Effects of CA (25 mg/kg) on escape latency time during the morris water maze trial sessions, on (A) the time spent in the quadrant and (B) crossing time, (C) time taken during probe trial sessions on C57BL/6J mice. Mice were administered orally with water or CA (25 mg/kg/day) for 30 days, 60 min prior to trial sessions. The training trial and probe trial sessions were performed as described in the Materials and Methods. All the values are expressed as mean ± SD. NC: negative control (aged water-treated group); PC: positive control (young water-treated group); CA: Cuminaldehyde treated group. * P < 0.05; ** P < 0.01 as compared to the negative control.
After 7 days of training, a probe test was performed, in which the platform was removed and the time spent in the quadrant (Figure 5B) and time taken to cross the quadrant (Figure 5C) were evaluated. The results showed that aged-CA-treated mice showed a significant increase by 10% in the time spent in the quadrant (P < 0.01) and the crossing time (P < 0.01) as compared to the aged water-treated mice (negative control).
Effect of cuminaldehyde on locomotor activity in C57BL/6J mice
In the probe trial (day 8), we also measured immobility time of mice for evaluation of their motivation. Our results showed that there was a significant increase by 16% (P < 0.05) in the time spent swimming by aged-CA-treated group compared to the aged-water-treated group (negative control), however; no significance was observed between the aged-water-treated mice (negative control) and the young water-treated group (positive control) (P > 0.05) (Figure 6).
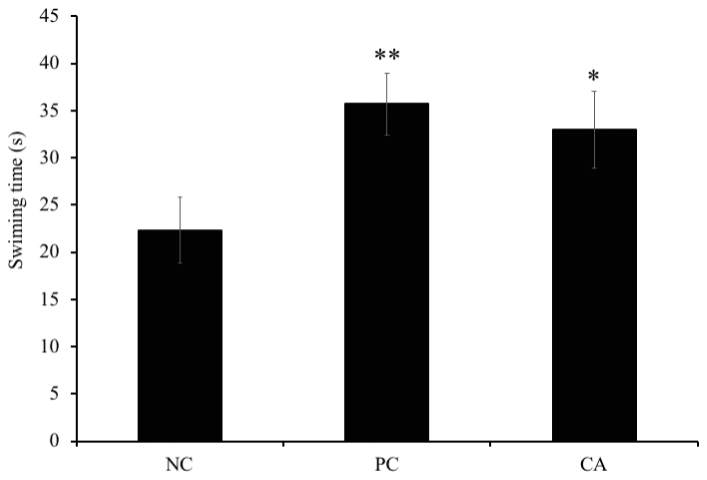
Figure 6. Effects of CA on swimming time during probe test of morris water maze trial sessions on C57BL/6J mice. Mice were administered orally with water or CA (25 mg/kg/day) for 30 days 60 min prior to trial sessions. Data is represented as means ± SEM (* P < 0.05 as compared to negative control).
Effect of cuminaldehyde on gene expression in C57BL/6J mice
To determine the molecular mechanism underlying the enhancement of spatial learning and memory of CA, we analyzed changes in gene expression of four genes (Bdnf, IL-6, Icam, and ApoE) in the mice brain using RT-PCR (Figure 6). Compared to the aged water-treated mice (negative control), a dose of 25 mg/kg/day of CA treatment in C57BL/6J mice, for 30 days, up-regulated the expression of Bdnf by 58% (Figure 7A), Icam by 34% (Figure 7B), and ApoE by 58% (Figure 7C), while it down-regulated the expression of IL-6 by 15% (P < 0.05) (Figure 7D).
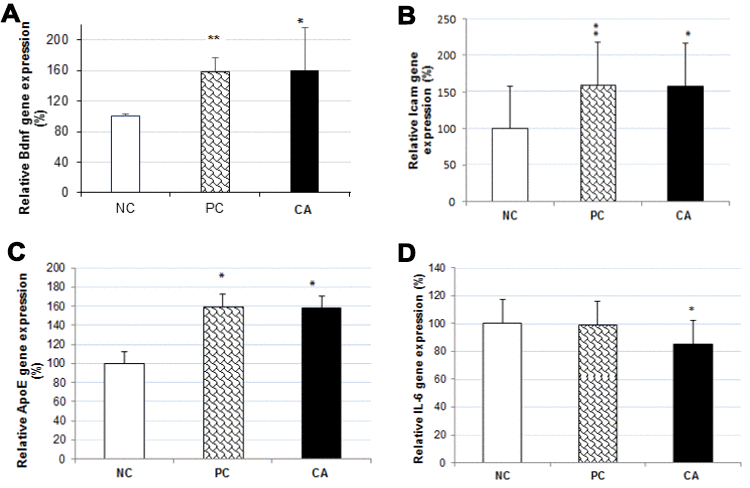
Figure 7. Effect of CA on mRNA expressions of Bdnf (A), Icam (B), ApoE (C) and IL-6 (D) in C57BL/6J mice brain. The mRNA expression of all genes were normalized to GAPDH mRNA expression and expressed with respect to the negative control (aged water-treated group). C57BL/6J mice were administrated with CA for 30 days. The control group was administered with distilled water. Each bar represents the mean ± SD. * P < 0.05, ** P < 0.01 as compared to negative control group. NC: negative control (aged water-treated group), PC: positive control (young water-treated group), CA: Cuminaldehyde treated group.
Effect of cuminaldehyde on neurotransmitters concentration in C57BL/6J mice brain
To evaluate the effect of CA on the levels of dopamine, adrenaline, and noradrenaline in the brain of C57BL/6J mice, their concentrations in the brain were measured using ELISA technique. The results showed no significant changes in the concentrations of dopamine (Figure 8A) and adrenaline (Figure 8B) in CA-treated mice brain compared to aged water-treated mice (negative control); whereas there was a significant increase (13%, P < 0.05,) in the concentration of noradrenaline (Figure 8C).
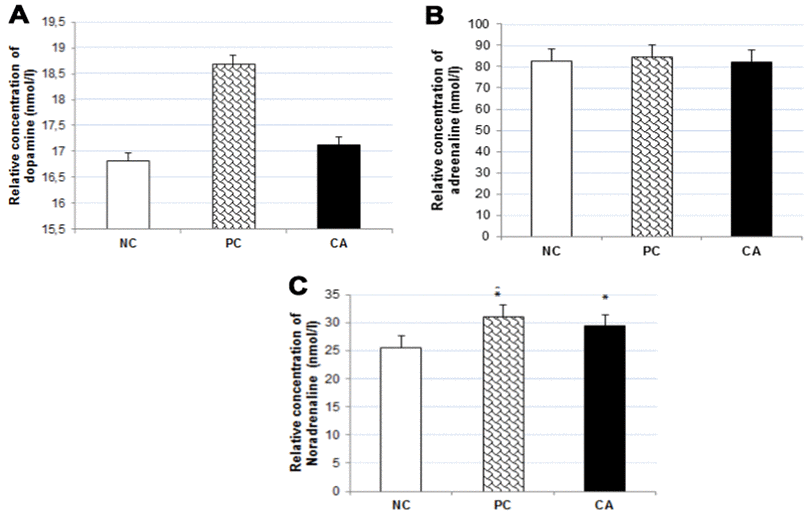
Figure 8. Effect of CA on catecholamine concentrations: Dopamine (A), Adrenaline (B), and Noradrenalin (C) in C57BL/6J mice brain. C57BL/6J mice were administrated with CA for 30 days. The control group was administered with distilled water. Each bar represents the mean ± SD. * P < 0.05, ** P < 0.01, *** P < 0.001 as compared to negative control group. NC: negative control (aged water-treated group), PC: positive control (young water-treated group), CA: Cuminaldehyde treated group.
Effect of cuminaldehyde on the levels of TNF-α and IL-6 in C57BL/6J mice brain
To evaluate the effect of CA on the levels of TNF-α and IL-6 in the brain of C57BL/6J mice, their concentrations were measured in the brain using ELISA technique. Our result showed that brain TNF-α was significantly (P < 0.05) increased in aged water-treated mice (negative control) (21.6 ± 2.3 pg/μg protein) compared with young water-treated group (positive control) (16.4 ± 1.7 pg/μg protein) (Figure 9A). However, CA-treated group showed a significant decrease (17.2 ± 1.2 pg/μg protein) of TNF-α levels in brain compared with aged water-treated group (negative control) (P < 0.05, Figure 9A).
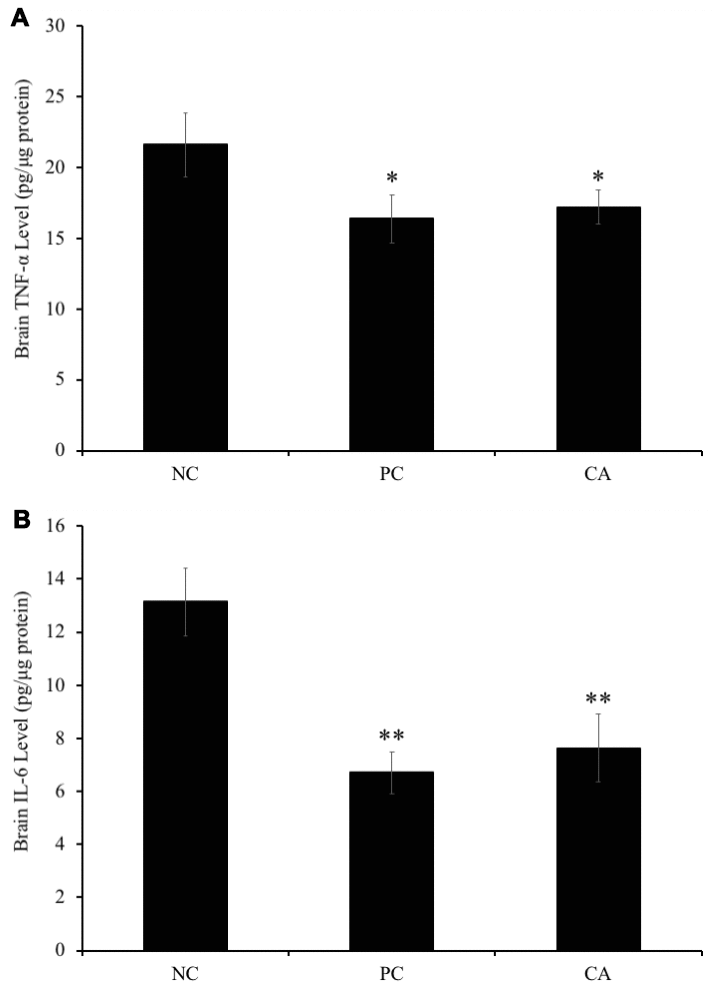
Figure 9. Effect of CA on levels of (A) tumor necrosis factor-α (TNF-α) and (B) interleukin 6 (IL-6) in C57BL/6J mice brain. C57BL/6J mice were administrated with CA for 30 days. The control group was administered with distilled water. Each bar represents the mean ± SD. * P < 0.05, ** P < 0.01 as compared to negative control group. NC: negative control (aged water-treated group), PC: positive control (young water-treated group), CA: Cuminaldehyde treated group.
Moreover, brain IL-6 level in aged water-treated group (negative control) was significantly (P < 0.01) increased compared with young water-treated group (positive control) (6.7 ± 0.8 pg/μg protein) (Figure 9B). Similarly, IL-6 level in the CA-administered group (7.6 ± 1.3 pg/μg protein) was significantly increased (P < 0.01) as compared to that of aged water-treated group (negative control) (13.1 ± 1.3 pg/μg protein).
Discussion
The activities of daily life of the majority of elderly people are affected by a form of memory loss. The incidence of dementia and memory impairment rise with the age. In addition, there are changes in the levels of neurotransmitters [2], for example, dopamine level is reported to decline with age and has been associated with decline in cognitive and motor performance [10, 11]. Therefore, any factor that may enhance the memory of elderly people will be of a great importance. CA is one of the major components of cumin, the second most popular spice in the world after black pepper. In the present study, CA was investigated for its neuroprotective effect and its potential in memory enhancement was evaluated.
To evaluate the neurotoxicity and the neuroprotective effect of CA, SH-SY5Y cells were treated with Dexa and/or CA. MTT results showed that the treatment of SH-SY5Y cells to CA for 72 h had no significant effect on the cell viability compared to the non-treated cells (control). All the concentrations of CA showed no cytotoxicity on SH-SY5Y cells; and the highest death rate occurred when Dexa was used alone at a concentration of 500 μM (Figure 3). However, when CA was co-incubated with Dexa, there was a significant decrease in cell death. Our results show that CA has a neuroprotective effect against Dexa-induced neurotoxicity. It has been shown that CA possesses antioxidant activity [20]; therefore, it can be speculated that the increased cell viability could be due to the antioxidant properties of CA.
In the present study, we also assessed the effect of CA on mice at the molecular level by real time PCR for analyzing the alterations in expression of some genes. We used the MWM to evaluate learning memory and locomotor activity. CA was administrated orally every day for 30 days at the concentration of 25 mg/kg, and the body weight was measured every day before oral administration. No mortality was observed and our results show that the body weight did not change throughout the experiment. This suggests that CA is not toxic at the tested concentration. However, unfortunately, there are currently no reports on the pharmacokinetics of CA. Therefore, in the future it is necessary to investigate the pharmacokinetics of CA, especially whether it can cross the blood-brain barrier.
The MWM test, tool for evaluation of spatial learning and memory tasks in rodents, was used in this experiment. Our results show that the escape latency time of aged C57BL/6J mice group administered with CA and young water-treated C57BL/6J mice group was significantly decreased as compared to the aged water-treated C57BL/6J mice from the 4th day of training. After 7 days of training, the escape latency time in aged CA-treated mice (CA group) and young water-treated mice significantly decreased with 20% compared to aged water-treated C57BL/6J mice. On the other hand, C57BL/6J mice administered with CA group show a decrease in the time spent in the quadrant and the crossing time compared to the aged water-treated C57BL/6J mice. Together, all the results obtained show that CA increases learning and memory. In the probe trial, time spent in the quadrant by mice swimming was recorded, and there was a significant increase in time spent swimming between aged CA-treated group and the aged water-treated group. Our findings suggest that CA also increases the locomotor activity in C57BL/6J mice.
Moreover, to understand the mechanism of action of CA and how it affects learning, memory, and locomotor activity, the expression of several genes involved in nerve cells growth and differentiation, neural circuitry, and synaptic plasticity was analyzed. Using RT-PCR, we analyzed the expression of Bdnf, IL-6, Icam, and ApoE genes. Our results showed that brain of 30 days CA treatment in C57BL/6J mice, up-regulated the gene expression of ApoE by 58% (P < 0.05) as compared to aged water-treated mice group. ApoE is a polymorphic protein synthesized by astrocytes and microglia but it can also be produced by neurons following injury [5]. ApoE is also strongly expressed in brain and liver [3, 4].
Various studies on learning and memory using animal models have identified many genes associated with these processes. Among them is the brain-derived neurotrophic factor (Bdnf), which is also a widely studied gene in brain research. Our results showed that brain of 30 days CA treatment in C57BL/6J mice up-regulated the gene expression of Bdnf by 58% as compared to aged water-treated group. The role of Bdnf in learning and memory has already been established by studies carried out in in vivo rodent models and in vitro cell models. Bdnf mRNA expression was reported to be increased in the hippocampus of rats following training with MWM [21], radial maze [22], passive avoidance [23], and contextual fear conditioning [24]. Increased Bdnf expression has been shown to play an important role in LTP, which contributes to synaptic plasticity, a form well-established as a cellular model for long-term memory (LTM) formation [6, 7]. In studies related to synaptic plasticity, LTP is the most studied form and is considered as a cellular correlate of learning and memory. The induction of LTP is associated with activation of multiple signaling cascades and is also activated by Bdnf [25]. In some in vitro studies, exogenous Bdnf induced the promotion of the LTP in young hippocampal slices [26], and rapidly increased the frequency of micro-excitatory postsynaptic currents in solitary neurons [27]. LTP and synaptic plasticity are known to be associated with ultrastructural changes in the dendritic spines at excitatory glutamatergic synapses [28–31]. LTP and synaptic plasticity are responsible for enormous ultrastructural changes [26–29] such as the formation of new spines [30–32] and the increase in the number of glutamatergic AMPA receptors at the level of dendritic spines [33–35]. These results suggest Bdnf plays a crucial role in learning and memory processes.
In our study, CA treatment down-regulated both, the gene and protein expression of IL-6 compared to the aged water-treated mice. The expression of IL-6 is under tight regulation in the leukocytes; however, with an increasing age this regulation is relaxed, so the circulating levels of IL-6 are reported to increase with an increase in the age [36, 37]. A recent study in old healthy people by Weaver (2002) reported a modest decline in cognitive ability in an age-dependent manner with increase in plasma IL-6. In addition, the results of gene expression levels showed an increase in IL-6 mRNA levels in the whole brain of aged mice compared to young adult mice [38]. Levels of IL-6 in the several brain parts such as hippocampus, cerebral cortex, cerebellum were increased in aged mice compared to young adult mice [39]. Therefore, the increase of gene expression levels of IL-6 in the aged mice brain is considered potentially relevant, as IL-6 may be neurotoxic and may affect cognitive and motivational systems [8]. This can also cause Alzheimer’s disease [9], in which senile plaques are formed around microglia and astrocytes, considering that the progression of the disease is partially caused by several cytokines and neurotoxic factors [40]. The expression of IL-6 in astrocytes, microglia, and neurons, which has been suggested to decrease the food intake, regulate the ability of memory and learning, cause neurodegeneration, and exacerbate illness induced by other cytokines in the brain [41]. Our results suggest that CA decreases the level of IL-6 and thereby, has a protective effect against IL-6 induced memory loss. Our results also showed an increase in the concentration of noradrenaline by 13% in CA-treated mice compared to the aged water-treated mice. During normal aging, noradrenaline level in old animals is significantly lower in the hypothalamus, as compared to the young rats [42]. Stimulation of noradrenergic receptors in the hippocampus results in changes in neuronal excitability and synaptic plasticity, considering an important role for noradrenaline in learning and memory. Consistent with this notion, noradrenaline enhances memory for various hippocampal-dependent tasks. For example, the effect of noradrenaline receptor activation on cellular plasticity may cause noradrenaline-dependent regulation of memory in the hippocampus. In addition, dysfunction of the noradrenergic neuromodulatory system causes many cognitive and mental disorders [43].
On the other hand, our results show no significant changes in the concentration of dopamine and adrenaline compared to aged water-treated mice, which contradicts the literature that reports that the levels of dopamine [44–46] and adrenaline in the rat and mice brain decrease with aging [47, 48]. Dopamine plays an essential role in the control of body movements. Age-related reductions in dopamine levels are considered to be responsible for many age-related neurological symptoms, such as decreased arm swing and increased rigidity [49]. In addition, fluctuations in dopamine levels can cause age-related changes in cognitive flexibility. In our study, we observed an increase in the swimming time in CA-treated mice, which corroborates with the finding that an increase in neurotransmitter level (dopamine) increases the locomotor activity in CA-treated group.
Conclusions
In conclusion, the treatment of SH-SY5Y cells with CA shows a neuroprotective effect against Dexa-induced cells toxicity. Moreover, oral administration of CA demonstrated learning and memory enhancement in the MWM test. This effect is co-related to a significant increase in the concentration of noradrenaline; and is also related to the up-regulation of Bdnf, Icam, and ApoE expression and down-regulation of IL-6 expression that is increased with aging. Our findings provide the first evidence that CA has a neuroprotective effect, as well as a spatial learning and memory enhancement potential through the modulation of genes coding for neurotrophic factors and/or those implicated in the imbalance of neural circuitry and impairments of synaptic plasticity. In the previous study, it should be pointed out that CA accounted for 48.77 % of the whole content of cumin essential oil [36]. Therefore, it may be considered that daily intake of cumin essential oil can contribute to reasonable improvement of cognitive function. Altogether, our results suggest that CA could be a promising drug for elderly against amnesia and memory deficits.
Materials and Methods
Chemicals and reagents
CA (Figure 1), Dulbecco’s Modified Eagle Medium (DMEM), Ham and Fetal bovine serum (FBS) were purchased from Sigma Aldrich Co., Ltd. (Irvine, United Kingdom). Penicillin and Streptomycin were purchased from Lonza Inc. (Walkersville, MD), while Eagle’s minimum essential medium (OPTI-MEM) was purchased from Gibco (Japan), and 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) from DOJINDO, (Japan). SDS was purchased from GE Healthcare (Sweden).
Cell culture
The human neuroblastoma SH-SY5Y cell line was obtained from American Type Culture Collection (ATCC). This cell line is widely used in the field of neuroscience for the research on Alzheimer's disease, neurotoxicity, ischemia, and amyotrophic lateral sclerosis [50]. According to our previous study [51], SH-SY5Y neuroblastoma cells were cultured in 100 mm petri dish or in 96-well plates with a 1:1 (v/v) mixture of Dulbecco’s Modified Eagle Medium (DMEM) and Ham’s F-12 nutrient mixture supplemented with 15% fetal bovine serum (FBS), and 1% penicillin (5000 μg/mL) streptomycin (5000 IU/mL) solution at 37° C in a 95% humidified air of 5% CO2 incubator. A serum-free Eagle’s minimum essential medium (Opti-MEM) was used to culture the cells for cell viability assay.
MTT assay
To assess the cell viability of SH-SY5Y cells, MTT assay was performed as previous our study [51]. SH-SY5Y cells were cultured at a density of 2 x 105 cells/well in a fibronectin coated 96-well micro-plate (BD, BioCoat). After a period of 24 h incubation at 37° C (5% CO), cells were treated with CA, at concentration of 5, 50 or 100 μM, dissolved in Opti-MEM. After 72 h of incubation, 10 μL of MTT (5 mg/mL) dissolved in 100 μL of Opti-MEM was added to each well and the plate was incubated for further 6 h. The colored formazan was dissolved in 100 μL of 10% SDS. The cell viability was determined using a multi-detection micro plate reader (Powerscan_1-42, HT, Dainippon Pharmaceutical, Japan) using absorbance at 570 nm, and the results were represented as percentage compared to control.
Neuroprotective assay
To investigate the neuroprotective effect of CA, Dexa, a synthetic glucocorticoid used as an inducer of apoptosis, was used according to our previous study [51]. SH-SY5Y cells were cultured in 96-well plate (fibronectin-coated plate) and treated with CA and co-treated with Dexa (500 μM) for 24 h and 72 h. After the treatment, 100 μL of Opti-MEM and 10 μL of MTT (5 mg/mL) were added, and cell viability assay was performed as described above.
Animals
C57BL/6J male mice aged 10 weeks and 9 months, weighting between 25 g and 35 g were used in the experiment. The animals were obtained from Japan (Japan SLC Company, Japan). Mice were housed individually, one mouse per cage and had free access to food and water, under a 12 h light/12 h dark cycle in a controlled environment of 56% humidity and 23° C temperature.
After 7 days of acclimatization to the laboratory conditions, mice were divided into 3 groups: a positive control group composed of 10 weeks-old mice (young water-treated group) (n = 7), a negative control group composed of 9 months-old mice (aged water-treated group) (n = 5), and CA-treated group composed of 9 months-old mice (n = 8) which were treated by oral administration of CA (25 mg/kg) for 30 days. The concentration dose (25 mg/kg) of CA for in vivo study was determined based on the previous study using cuminum cyminum extract [52]. The respective control groups (positive and negative groups) were administered an equivalent volume of distilled water. This animal experiment was approved by the Animal Care and Use Committee of the University of Tsukuba.
Morris water maze (MWM) test
The MWM test was carried out using a circular pool of 120 cm in diameter and 45 cm in height as reported previously [53]. On the inner surface of the circular pool four symbols (a square, a circle, a cross mark, and a triangle) were attached. The pool was divided into four quadrants (north, east, west, and south) and filled with water (23 ± 2° C). A Plexiglas platform (10 cm of diameter) was placed exactly in the northeast quadrant, submerged 1 cm below the water surface. The platform was fixed in the same place in the pool throughout the experiment. The mice were given a trial session (60 s) 4 times each day for 7 days, then each mice were placed on the platform for 30 s. CA was administered to the treatment group 30 min before the experiment.
After 7 days, each mouse performed one probe trial, in which the platform is removed from MWM pool. Probe trials are performed to validate the strategy followed by the animals when they notice an absence of the platform. The animals were released into the pool from a single point and the number of times the animal crosses the quadrant of the platform in 30 s was recorded.
Tissue preparation
Mice were sacrificed by cervical dislocation 24 h after the last trial of the MWM test and 30 min after the last oral administration of the pure compound. The brain samples were dissected and fixed in liquid nitrogen, and stored in -80° C until use.
RNA extraction from mouse brain
A total of 100 mg of the cerebral tissue was removed and washed with ice-cold phosphate-buffered solution (PBS). The total RNA was extracted from it using the ISOGEN kit (Nippon Gene Co. Ltd., Toyama, Japan), following the manufacturer’s instruction. Total RNA was quantified and assessed for its purity with the NanoDrop 2000 spectrophotometer (Thermo Scientific, Wilmington, DE, USA).
RT-PCR
The Superscript III reverse transcriptase kit (Invitrogen, Carlsbad, CA, USA) allowed us to carry out reverse transcription reactions [54]. Briefly, the RNA was denatured at 65° C for 5 min, incubated with 1 ml of oligo primer (dT) and cooled to 4° C. After adding the Superscript III reverse transcriptase (200 units), the reaction mixture was incubated at 42° C for 60 minutes, then for 10 minutes at 70° C. The cDNA was used to evaluate the gene expression of Bdnf, ApoE, Icam, and IL-6. This experiment was conducted using TaqMan Universal PCR mix and TaqMan Probes, and the amplification reaction was performed in a 7500 Fast Real-time PCR (Applied Biosystems, USA) with the following conditions: 50° C for 2 min, followed by 95° C for 10 min, and 40 cycles of 95° C for 15 s followed by 60° C for 1 min.
Dopamine, noradrenaline and adrenaline levels in mice brain
Dopamine, noradrenaline, and adrenaline levels in the mice brains were measured by Dopamine, Adrenaline, and Noradrenaline Research ELISA kits (ImmuSmol, Pessac, France) as reported in the previous study [54]. Briefly, the cerebral cortex (100 mg) was isolated from brain tissue of mice and homogenized in radioimmunoprecipitation assay (RIPA) buffer with protease inhibitor (Santa Cruz Biotechnology, Inc., Tokyo, Japan). Further, 50 μL of protein samples or 10 μL standards were used for catecholamine extraction, acylation, and determination according to manufacturer’s instructions. After optimal color development, the reaction was stopped and the absorbance was measured at 450 nm. Protein concentration was determined using the 2-D Quant kit (GE Healthcare Inc., Tokyo, Japan) and the data were expressed as ng/μg protein.
Brain tumor necrosis factor-α (TNF-α) and interleukin 6 (IL-6) quantification
Brain tumor necrosis factor-α (TNF-α) and interleukin 6 (IL-6) levels in the brain were measured using ELISA kit (TNF-α: Proteintech, Japan; IL-6: Invitrogen, Japan) in accordance with the manufacturer’s instructions. Briefly, 100 mg of the limbic area was isolated from the brain tissue and homogenized in RIPA buffer with protease inhibitor (Santa Cruz Biotechnology, inc., Tokyo, Japan). Following centrifugation (1000 × g, 20 min), the supernatants or standards were used to determine the TNF-α and IL-6 levels in brain. After the addition of TNF-α or IL-6 antibody, a second incubation was performed with streptavidin-horseradish peroxidase conjugate solution. After addition of substrate and stop solution, TNF-α and IL-6 levels were determined by measuring absorbance at 450 nm.
Statistical analysis
All results are expressed as mean ± standard deviation (SD), and statistical analyses were performed using a Student’s t-test using SigmaPlot 12.0 software. Results were considered statistically significant when P-value was less than 0.05.
Author Contributions
KS, ZO, and HI conceived and designed the experiments; KS and ZO performed the experiments; KS and ZO prepared the figures; KS and ZO analyzed and interpreted the results, and wrote the manuscript; HI edited and revised the manuscript.
Acknowledgments
The authors would like to thank the support provided by the Japan Science and Technology Agency (JST) and by the Technology Research Partnership for Sustainable Development (SATREPS).
Conflicts of Interest
The authors declare no conflicts of interest.
Funding
This study was partially funded by the project of Japan Science and Technology Agency (JST) Science and Technology Research Partnership for Sustainable Development (SATREPS).
References
- 1. Esler M, Kaye D, Thompson J, Jennings G, Cox H, Turner A, Lambert G, Seals D. Effects of aging on epinephrine secretion and regional release of epinephrine from the human heart. J Clin Endocrinol Metab. 1995; 80:435–42. https://doi.org/10.1210/jcem.80.2.7852502 [PubMed]
- 2. Martin SJ, Grimwood PD, Morris RG. Synaptic plasticity and memory: an evaluation of the hypothesis. Annu Rev Neurosci. 2000; 23:649–711. https://doi.org/10.1146/annurev.neuro.23.1.649 [PubMed]
- 3. Durand GM, Kovalchuk Y, Konnerth A. Long-term potentiation and functional synapse induction in developing hippocampus. Nature. 1996; 381:71–75. https://doi.org/10.1038/381071a0 [PubMed]
- 4. Wang E, Snyder S. Handbook of the aging brain. Academic Press. 1998. https://doi.org/10.1016/B978-0-12-734610-6.X5000-3
- 5. Bu G. Apolipoprotein E and its receptors in Alzheimer’s disease: pathways, pathogenesis and therapy. Nat Rev Neurosci. 2009; 10:333–44. https://doi.org/10.1038/nrn2620 [PubMed]
- 6. Ershler WB, Keller ET. Age-associated increased interleukin-6 gene expression, late-life diseases, and frailty. Annu Rev Med. 2000; 51:245–70. https://doi.org/10.1146/annurev.med.51.1.245 [PubMed]
- 7. Ye SM, Johnson RW. Increased interleukin-6 expression by microglia from brain of aged mice. J Neuroimmunol. 1999; 93:139–48. https://doi.org/10.1016/s0165-5728(98)00217-3 [PubMed]
- 8. Liao D, Hessler NA, Malinow R. Activation of postsynaptically silent synapses during pairing-induced LTP in CA1 region of hippocampal slice. Nature. 1995; 375:400–04. https://doi.org/10.1038/375400a0 [PubMed]
- 9. Venero JL, Machado A, Cano J. Changes in monoamines and their metabolite levels in substantia nigra of aged rats. Mech Ageing Dev. 1989; 49:227–33. https://doi.org/10.1016/0047-6374(89)90073-0 [PubMed]
- 10. Fjell AM, Walhovd KB. Structural brain changes in aging: courses, causes and cognitive consequences. Rev Neurosci. 2010; 21:187–221. https://doi.org/10.1515/revneuro.2010.21.3.187 [PubMed]
- 11. Kesslak JP, So V, Choi J, Cotman CW, Gomez-Pinilla F. Learning upregulates brain-derived neurotrophic factor messenger ribonucleic acid: a mechanism to facilitate encoding and circuit maintenance? Behav Neurosci. 1998; 112:1012–19. https://doi.org/10.1037//0735-7044.112.4.1012 [PubMed]
- 12. Nyberg L, Backman L. Cognitive ageing: a view from brain imaging. In: Dixon R, Backman L, Nilsson L, eds. New frontiers in cognitive ageing. Oxford University Press, 2004; 135–60. https://doi.org/10.1093/acprof:oso/9780198525691.003.0007
- 13. Ida Y, Tanaka M, Kohno Y, Nakagawa R, Iimori K, Tsuda A, Hoaki Y, Nagasaki N. Effects of age and stress on regional noradrenaline metabolism in the rat brain. Neurobiol Aging. 1982; 3:233–36. https://doi.org/10.1016/0197-4580(82)90044-6 [PubMed]
- 14. Taniguchi N, Takada N, Kimura F, Tsumoto T. Actions of brain-derived neurotrophic factor on evoked and spontaneous EPSCs dissociate with maturation of neurones cultured from rat visual cortex. J Physiol. 2000; 527:579–92. https://doi.org/10.1111/j.1469-7793.2000.t01-1-00579.x [PubMed]
- 15. Khan IA, Abourashed EA. Leung’s encyclopedia of common natural ingredients used in food, drugs and cosmetics, 3rd ed. John Wiley & Sons Inc: Hoboken, New Jersey. (2009).
- 16. Bliss TV, Collingridge GL. A synaptic model of memory: long-term potentiation in the hippocampus. Nature. 1993; 361:31–39. https://doi.org/10.1038/361031a0 [PubMed]
- 17. Godbout JP, Johnson RW. Interleukin-6 in the aging brain. J Neuroimmunol. 2004; 147:141–44. https://doi.org/10.1016/j.jneuroim.2003.10.031 [PubMed]
- 18. Lee HS. Cuminaldehyde: aldose reductase and alpha-glucosidase inhibitor derived from cuminum cyminum L. Seeds. J Agric Food Chem. 2005; 53:2446–50. https://doi.org/10.1021/jf048451g [PubMed]
- 19. Seals DR, Esler MD. Human ageing and the sympathoadrenal system. J Physiol. 2000; 528:407–17. https://doi.org/10.1111/j.1469-7793.2000.00407.x [PubMed]
- 20. Kannappan R, Gupta SC, Kim JH, Reuter S, Aggarwal BB. Neuroprotection by spice-derived nutraceuticals: you are what you eat!. Mol Neurobiol. 2011; 44:142–59. https://doi.org/10.1007/s12035-011-8168-2 [PubMed]
- 21. Isaac JT, Nicoll RA, Malenka RC. Evidence for silent synapses: implications for the expression of LTP. Neuron. 1995; 15:427–34. https://doi.org/10.1016/0896-6273(95)90046-2 [PubMed]
- 22. Mizuno M, Yamada K, Olariu A, Nawa H, Nabeshima T. Involvement of brain-derived neurotrophic factor in spatial memory formation and maintenance in a radial arm maze test in rats. J Neurosci. 2000; 20:7116–21. https://doi.org/10.1523/JNEUROSCI.20-18-07116.2000 [PubMed]
- 23. Karasawa N, Yamawaki Y, Nagatsu T, Kawase T, Nishiyama K, Watanabe K, Onozuka M, Nagatsu I. Age-associated changes in the dopamine synthesis as determined by GTP cyclohydrolase I inhibitor in the brain of senescence-accelerated mouse-prone inbred strains (SAMP8). Neurosci Res. 1999; 35:31–36. https://doi.org/10.1016/s0168-0102(99)00063-2 [PubMed]
- 24. Kálmán J, Juhász A, Laird G, Dickens P, Járdánházy T, Rimanóczy A, Boncz I, Parry-Jones WL, Janka Z. Serum interleukin-6 levels correlate with the severity of dementia in down syndrome and in Alzheimer’s disease. Acta Neurol Scand. 1997; 96:236–40. https://doi.org/10.1111/j.1600-0404.1997.tb00275.x [PubMed]
- 25. Chen L, Tracy T, Nam CI. Dynamics of postsynaptic glutamate receptor targeting. Curr Opin Neurobiol. 2007; 17:53–58. https://doi.org/10.1016/j.conb.2006.11.001 [PubMed]
- 26. Figurov A, Pozzo-Miller LD, Olafsson P, Wang T, Lu B. Regulation of synaptic responses to high-frequency stimulation and LTP by neurotrophins in the hippocampus. Nature. 1996; 381:706–09. https://doi.org/10.1038/381706a0 [PubMed]
- 27. Toni N, Buchs PA, Nikonenko I, Bron CR, Muller D. LTP promotes formation of multiple spine synapses between a single axon terminal and a dendrite. Nature. 1999; 402:421–25. https://doi.org/10.1038/46574 [PubMed]
- 28. Kafitz KW, Rose CR, Thoenen H, Konnerth A. Neurotrophin-evoked rapid excitation through TrkB receptors. Nature. 1999; 401:918–21. https://doi.org/10.1038/44847 [PubMed]
- 29. Pfrieger FW. Cholesterol homeostasis and function in neurons of the central nervous system. Cell Mol Life Sci. 2003; 60:1158–71. https://doi.org/10.1007/s00018-003-3018-7 [PubMed]
- 30. Ma YL, Wang HL, Wu HC, Wei CL, Lee EH. Brain-derived neurotrophic factor antisense oligonucleotide impairs memory retention and inhibits long-term potentiation in rats. Neuroscience. 1998; 82:957–67. https://doi.org/10.1016/S0306-4522(97)00325-4
- 31. Lee KS, Schottler F, Oliver M, Lynch G. Brief bursts of high-frequency stimulation produce two types of structural change in rat hippocampus. J Neurophysiol. 1980; 44:247–58. https://doi.org/10.1152/jn.1980.44.2.247 [PubMed]
- 32. Chang FL, Greenough WT. Transient and enduring morphological correlates of synaptic activity and efficacy change in the rat hippocampal slice. Brain Res. 1984; 309:35–46. https://doi.org/10.1016/0006-8993(84)91008-4 [PubMed]
- 33. Wei J, Xu H, Davies JL, Hemmings GP. Increase of plasma IL-6 concentration with age in healthy subjects. Life Sci. 1992; 51:1953–56. https://doi.org/10.1016/0024-3205(92)90112-3 [PubMed]
- 34. Morshedi D, Aliakbari F, Tayaranian-Marvian A, Fassihi A, Pan-Montojo F, Pérez-Sánchez H. Cuminaldehyde as the major component of cuminum cyminum, a natural aldehyde with inhibitory effect on alpha-synuclein fibrillation and cytotoxicity. J Food Sci. 2015; 80:H2336–45. https://doi.org/10.1111/1750-3841.13016 [PubMed]
- 35. Hajlaoui H, Mighri H, Noumi E, Snoussi M, Trabelsi N, Ksouri R, Bakhrouf A. Chemical composition and biological activities of Tunisian cuminum cyminum L. Essential oil: a high effectiveness against vibrio spp. Strains. Food Chem Toxicol. 2010; 48:2186–92. https://doi.org/10.1016/j.fct.2010.05.044 [PubMed]
- 36. Wei J, Zhang X, Bi Y, Miao R, Zhang Z, Su H. Anti-inflammatory effects of cumin essential oil by blocking JNK, ERK, and NF-κB signaling pathways in LPS-stimulated RAW 264.7 cells. Evid Based Complement Alternat Med. 2015; 2015:474509. https://doi.org/10.1155/2015/474509 [PubMed]
- 37. Geinisman Y, deToledo-Morrell L, Morrell F, Heller RE, Rossi M, Parshall RF. Structural synaptic correlate of long-term potentiation: formation of axospinous synapses with multiple, completely partitioned transmission zones. Hippocampus. 1993; 3:435–45. https://doi.org/10.1002/hipo.450030405 [PubMed]
- 38. Ye SM, Johnson RW. Regulation of interleukin-6 gene expression in brain of aged mice by nuclear factor kappaB. J Neuroimmunol. 2001; 117:87–96. https://doi.org/10.1016/s0165-5728(01)00316-2 [PubMed]
- 39. Allahghadri T, Rasooli I, Owlia P, Nadooshan MJ, Ghazanfari T, Taghizadeh M, Astaneh SD. Antimicrobial property, antioxidant capacity, and cytotoxicity of essential oil from cumin produced in Iran. J Food Sci. 2010; 75:H54–61. https://doi.org/10.1111/j.1750-3841.2009.01467.x [PubMed]
- 40. McRae A, Dahlström A, Ling EA. Microglial in neurodegenerative disorders: emphasis on Alzheimer’s disease. Gerontology. 1997; 43:95–108. https://doi.org/10.1159/000213838 [PubMed]
- 41. Harris KM, Fiala JC, Ostroff L. Structural changes at dendritic spine synapses during long-term potentiation. Philos Trans R Soc Lond B Biol Sci. 2003; 358:745–48. https://doi.org/10.1098/rstb.2002.1254 [PubMed]
- 42. Weaver JD, Huang MH, Albert M, Harris T, Rowe JW, Seeman TE. Interleukin-6 and risk of cognitive decline: MacArthur studies of successful aging. Neurology. 2002; 59:371–78. https://doi.org/10.1212/wnl.59.3.371 [PubMed]
- 43. Borodovitsyna O, Flamini M, Chandler D. Noradrenergic modulation of cognition in health and disease. Neural Plast. 2017; 2017:6031478. https://doi.org/10.1155/2017/6031478 [PubMed]
- 44. Mukherjee J, Christian BT, Dunigan KA, Shi B, Narayanan TK, Satter M, Mantil J. Brain imaging of 18F-fallypride in normal volunteers: blood analysis, distribution, test-retest studies, and preliminary assessment of sensitivity to aging effects on dopamine D-2/D-3 receptors. Synapse. 2002; 46:170–88. https://doi.org/10.1002/syn.10128 [PubMed]
- 45. Missale C, Nash SR, Robinson SW, Jaber M, Caron MG. Dopamine receptors: from structure to function. Physiol Rev. 1998; 78:189–225. https://doi.org/10.1152/physrev.1998.78.1.189 [PubMed]
- 46. Iacobellis NS, Lo Cantore P, Capasso F, Senatore F. Antibacterial activity of cuminum cyminum L. And carum carvi L. Essential oils. J Agric Food Chem. 2005; 53:57–61. https://doi.org/10.1021/jf0487351 [PubMed]
- 47. Holtzman DM, Herz J, Bu G. Apolipoprotein E and apolipoprotein E receptors: normal biology and roles in Alzheimer disease. Cold Spring Harb Perspect Med. 2012; 2:a006312. https://doi.org/10.1101/cshperspect.a006312 [PubMed]
- 48. Boelens MH. Spices and Condiments II. In: Volatile Compounds in Foods and Beverages. Maarse H, Ed.; Routledge: New York, 1991; 449–82. https://doi.org/10.1201/9780203734285-13
- 49. Engert F, Bonhoeffer T. Dendritic spine changes associated with hippocampal long-term synaptic plasticity. Nature. 1999; 399:66–70. https://doi.org/10.1038/19978 [PubMed]
- 50. Kovalevich J, Langford D. Considerations for the use of SH-SY5Y neuroblastoma cells in neurobiology. Methods Mol Biol. 2013; 1078:9–21. https://doi.org/10.1007/978-1-62703-640-5_2 [PubMed]
- 51. Sasaki K, El Omri A, Kondo S, Han J, Isoda H. Rosmarinus officinalis polyphenols produce anti-depressant like effect through monoaminergic and cholinergic functions modulation. Behav Brain Res. 2013; 238:86–94. https://doi.org/10.1016/j.bbr.2012.10.010 [PubMed]
- 52. Koppula S, Choi DK. Cuminum cyminum extract attenuates scopolamine-induced memory loss and stress-induced urinary biochemical changes in rats: a noninvasive biochemical approach. Pharm Biol. 2011; 49:702–08. https://doi.org/10.3109/13880209.2010.541923 [PubMed]
- 53. Sasaki K, Davies J, Doldán NG, Arao S, Ferdousi F, Szele FG, Isoda H. 3,4,5-tricaffeoylquinic acid induces adult neurogenesis and improves deficit of learning and memory in aging model senescence-accelerated prone 8 mice. Aging (Albany NY). 2019; 11:401–22. https://doi.org/10.18632/aging.101748 [PubMed]
- 54. Sasaki K, Iwata N, Ferdousi F, Isoda H. Antidepressant-like effect of ferulic acid via promotion of energy metabolism activity. Mol Nutr Food Res. 2019; 63:e1900327. https://doi.org/10.1002/mnfr.201900327 [PubMed]