



Introduction
The functional decline of the immune system with age, or immunosenescence, is associated with different hematopoietic changes, including a decrease in the replication ability of hematopoietic stem cells and B lymphopoiesis, and lower efficiency in CD4 and CD8 T cells response [1]. It has been shown that with age, the immune cells’ composition shifts, with a reduction of B cells in the blood in old individuals; increased B1 cells, activated and memory B cells in old peripheral blood in mice; increased levels in the bone marrow in aged mice; increased proportion of NK cells in human blood; and decreased number of NK cells in mouse blood with age [2–6]. Some of these shifts have been proposed as markers for biological aging as they could predict longevity [7].
T cells can undergo replicative senescence in human aging after extensive antigen-driven proliferation. These cells can be identified by the Cd8+ Cd28- markers. Donor-specific replicative senescent T cells found in organ transplant patients are tolerable to rejection, suggesting a possible suppressive mechanism of these cells in reducing reactivity against allograft [8, 9]. Several studies have observed a significant correlation between the number of Cd8+ Cd28- T cells and the decline of antibody response to influenza vaccination in the elderly [9, 10].
Aging also has profound effects on hematopoietic stem cells (HSCs). The number of lymphoid HSCs decreases with age, whereas the number of myeloid HSCs increases. Besides the quantity, the quality of lymphoid HSCs also declines with age, resulting in a lower proliferation rate and a higher apoptosis rate in T and B lineage progenitor cells [11]. Beyond the decline in the immune function and the number of cells and declining with age, the blood also contains other factors that can modulate aging. Parabiosis of young and old mice has been shown to reverse cardiac hypertrophy in old mice and rejuvenate aged satellite cells [12, 13]. Aging has also been shown to increase cellular heterogeneity [14, 15]. For example, a recent study using single-cell RNA-seq (scRNA-seq) showed that transcriptional heterogeneity of CD4+ T cells increased upon stimulation in old mice compared to young mice, indicating that gene expression of immune cells is dysregulated with age [15].
Here, we applied scRNA-seq on young and old mice to dissect the transcriptional and cell composition changes of all cell types in the peripheral blood with age.
Results
scRNA-seq of young and old blood identifies different cell types
We performed scRNA-seq using 10x Chromium on peripheral blood obtained from 2 young and 2 old mice. We removed cells with more than 10% mitochondrial reads, less than 500 genes, or more than 4000 genes. 14588 cells that passed this filter (4642 cells from old mouse 1, 2187 cells from old mouse 2, 3902 cells from young mouse 1, and 3857 cells from young mouse 2) were subsequently processed using Seurat/2.3.0 and 17 clusters were obtained (Figure 1A, 1B). Cell-type specific marker genes identified the cell type of each cluster, including Cd3e for T cells, Cd79a and Ms4a2 for B cells, Nkg7 for NK cells, Ly6c2 and Cx3cr1 for monocytes or dendritic cells (DC), Ppbp, Pf4 and Gng11 for megakaryocytes, C1qa, C1qb and C1qc for macrophage, Fcer1a and Cd200r3 for basophil and Hba-a1 for erythrocytes (RBC) (Figure 1C). We identified 5 clusters of T cells, 4 clusters of B cells, 1 cluster of proliferating B or T cells, NK cells, monocytes, dendritic cells, megakaryocytes, macrophage, basophil, and (RBC), respectively (Figure 1C).
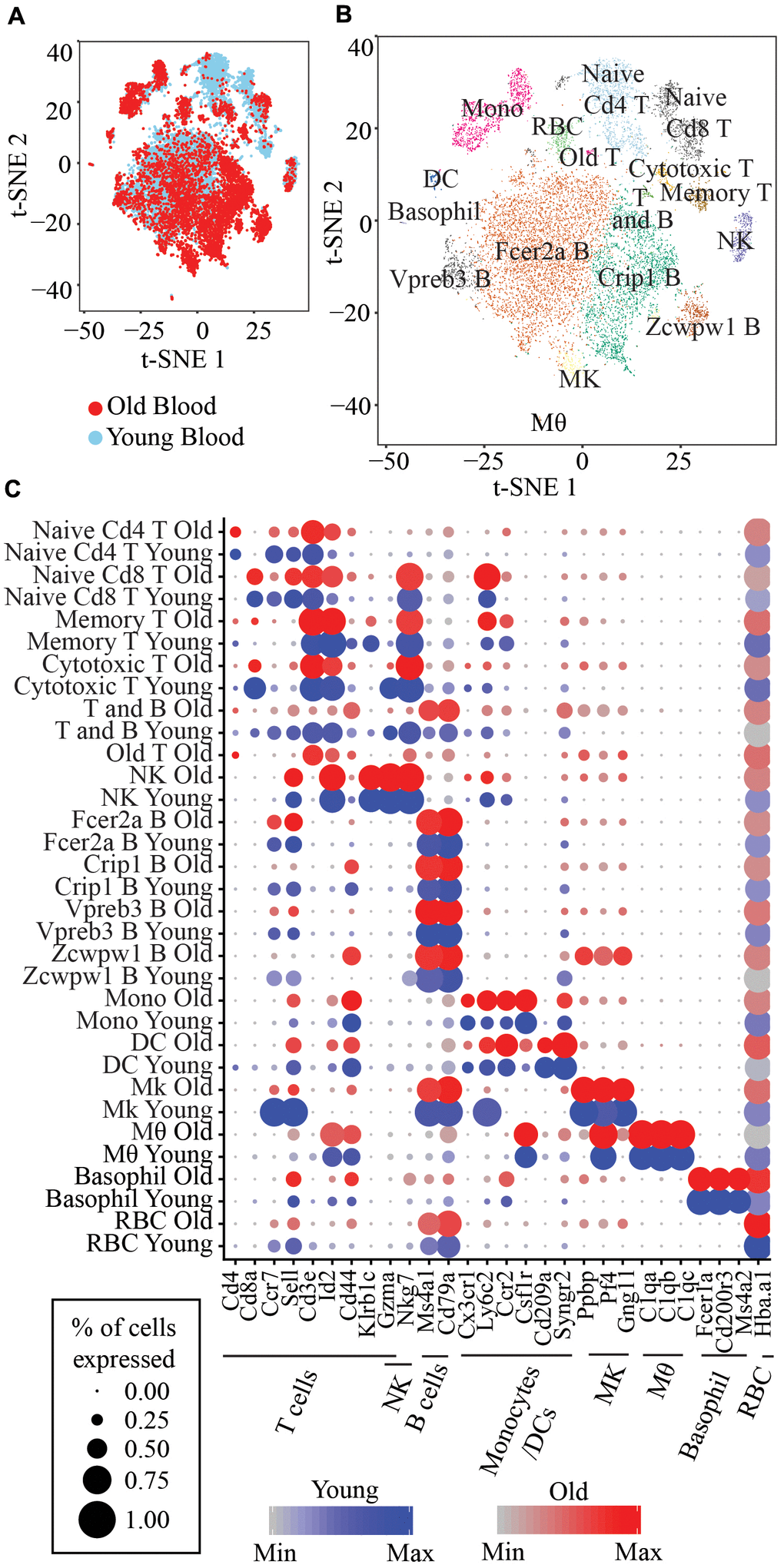
Figure 1. (A) t-SNE visualization of 14,588 old and young peripheral blood cells. (B) t-SNE visualization of the 17 clusters of peripheral blood cells. Memory T: Short-lived effector memory T cells; T and B: Proliferating T and B cells; NK: Natural killer cells; Fcer2a B: Fcer2a, Sell, Ccr7 B cells; Crip1 B: Crip1, S100a6 B cells; Vpreb3 B: Vpreb3, Spib B cells; Zcwpw1 B: Zcwpw1, S100a6 B cells; Mono: Classical monocytes; DC: Monocyte-derived DC; Mk: Megakaryocytes; Mθ: Macrophage; RBC: Red blood cells. (C) Marker genes for each immune cell type and the corresponding clusters.
The naive T cells cluster is identified by two markers, Sell+ and Cd44-, the memory T cells cluster is identified by Klrb1c+, Id2high, and Cd44+ and cytotoxic T cells are identified by high expression of Gzma (Figure 1C). We also identified four clusters of B cells, with one cluster expressing Fcer2a, Sell, and Ccr7 marker genes, which we herein refer to as Fcer2a B cells, and one cluster highly expressing Vpreb3 and Spib genes that we refer to as Vpreb3 B cells, one cluster with Crip1 and S100a6 high expression (Crip1 B cells) and lastly, one cluster of B cells highly expressing Zcwpw1, Lgals1 and Adm (Zcwpw1 B cells) (Supplementary Figure 1B). Classical monocytes are identified by a combination of markers, Ly6c2+, Ccr2+, Sell+ and Csf1r+ whereas monocyte-derived DC are Ly6c2+, Ccr2+, Cd209+ and Syngr2+ (Figure 1C).
Transcriptomic changes of the same cell type with age
We used Seurat to identify differential gene expression within clusters with age. Only genes expressed in at least 10% of cells in each cluster and age group were analyzed. We observed a general trend of the upregulation of genes involved in “Antigen processing and presentation” and “Chemokine signaling” pathways with age in naïve Cd4 and Cd8 T cells and Vpreb3 B cells. NK cells from old mice also exhibit increased expression of genes in the “Chemokine signaling” pathway (Figure 2A). In addition, we also observed an upregulation of genes in “Antigen processing and presentation” in macrophages, Crip1 B cells, and Fcer2aB cells. Crip1 B cells and monocytes also showed an upregulation of genes in the “Oxidative phosphorylation” pathway. In contrast, we observed a trend of downregulated genes involved in “Cytoplasmic Ribosomal Proteins” and “Ribosome” pathways in several cell types, naïve Cd4 and Cd8 T cells, Crip1 B cells, and monocytes, with age (Figure 2A).
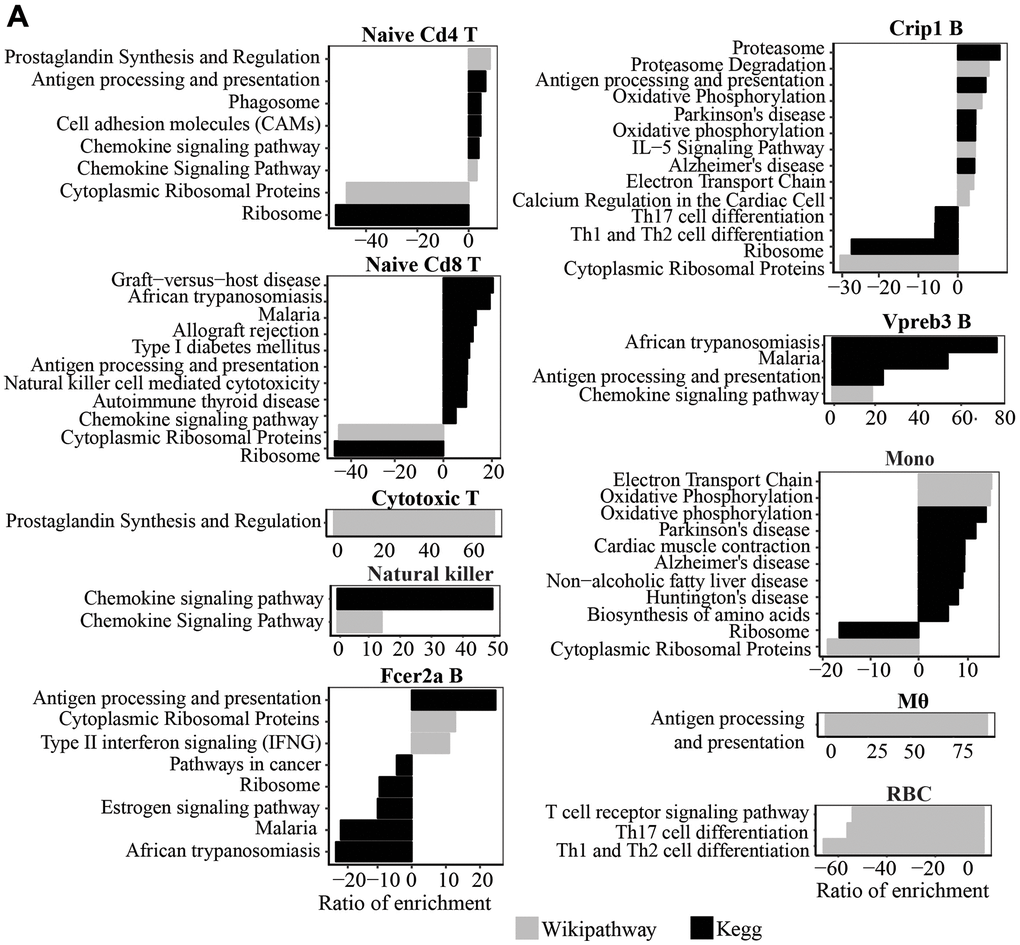
Figure 2. (A) Wikipathway and Kegg pathway analysis of upregulated and downregulated genes in old compared to young mice (P<0.05) in each cluster.
Cell type composition with age
B cells comprised the largest proportion of immune cells isolated from the peripheral blood (57.4% in young mice and 70.9% in old mice) (Figure 3A). In young mice, 30.38% of the isolated cells are T cells, 4.66% are monocytes, 4.16% are NK cells, 0.6% are DC, 0.39% are basophils, 0.27% are macrophages, and 0.01% are Mk cells. In old mice, 8.41% of the cells are T cells, 10.84% are monocytes, 4.03% are Mk cells, 1.34% are NK cells, 1.09% are DC, 0.44% are macrophages, and 0.26% are basophils. Although red blood cell lysis was performed on the samples, we still retrieved 2.06% of RBC from the young mice and 2.68% from old mice. Using the same technique in extracting the cells from young and old mice, we observed a few changes to the composition of cell types with age. First, we detected a significantly lower percentage of T cells and NK cells and higher percentage of B cells and Mk cells in old mice compared to young mice (Figure 3B). Second, within the subsets of B cells, we observed a decrease in the percentage of Fcer2a B cells and increased Crip1 B cells in old mice (Figure 3C). We did not detect any significant changes to the subsets of T cells (Figure 3D).
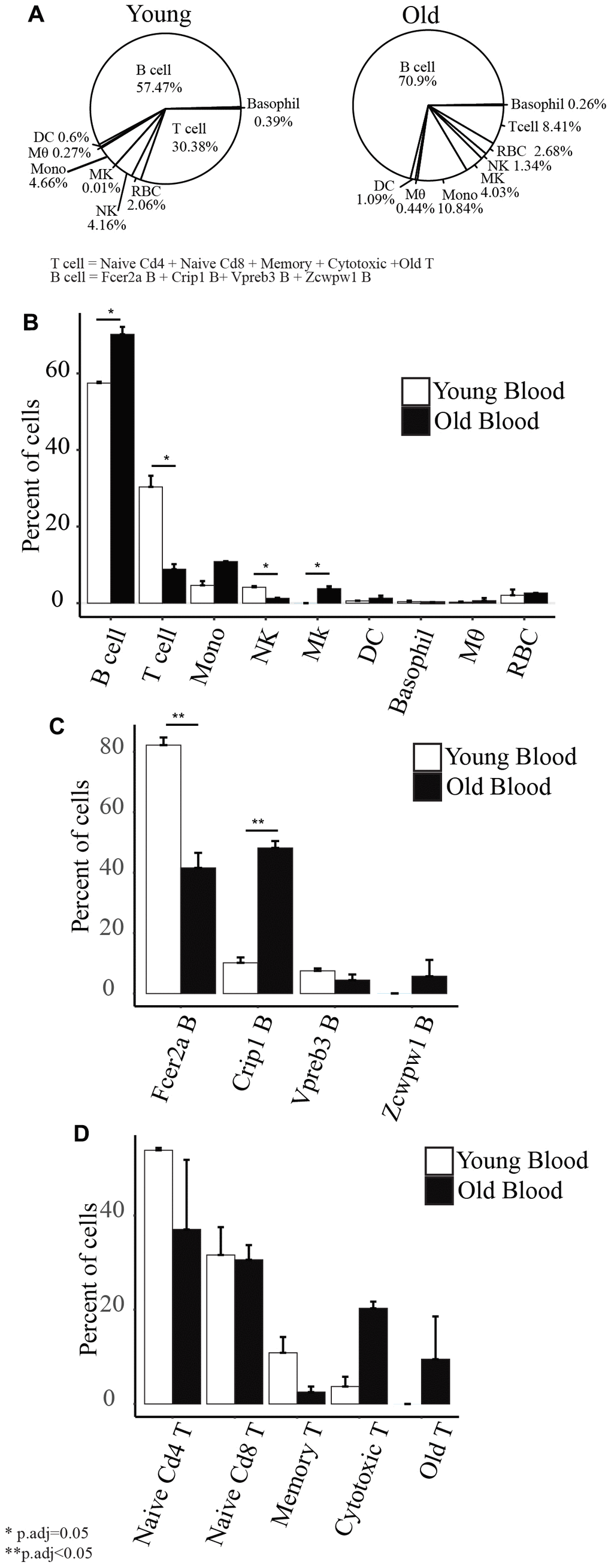
Figure 3. (A) Cell type composition of old and young peripheral blood. (B) Barplots showing the comparison of the percentage of each cell type with age. (C) Barplots showing the comparison of the subset of B cells’ percentage with age. (D) Barplots showing the comparison of the subsets of T cells’ percentage with age.
Immunosenescence
We identified one cluster that consists of only cells from old mice (labeled as “Old T” in Figure 1B), and these cells expressed Cd3e, the marker of T cells. In addition, this cluster also expressed Cd40lg and Tnfsf8 compared to other T cell clusters, suggesting that it consists of activated T cells (Supplementary Figure 2). Pathway analysis of genes significantly upregulated between this cluster and other T cells clusters showed enrichment of “Antigen processing and presentation” and “NF-kappa B signaling pathway” (Figure 4A). In addition, this cluster expressed significantly higher Bcl2 expression (p=3.9x10-15) (Figure 4B) than other T cells clusters. No other pro-apoptotic gene (Bax, Bak1, Bad, Bid, Bik) or anti-apoptotic gene (Bcl2, BclxL, Bcl2l1, Nfkb1, Cflar) was significantly differentially expressed. Notably, it also showed significantly higher expression of Cdkn2a and lower expression of Cd28 than all other clusters (p=5.38x10-40 and p=6.26x10-8, respectively) (Figure 4C and Supplementary Figure 2).
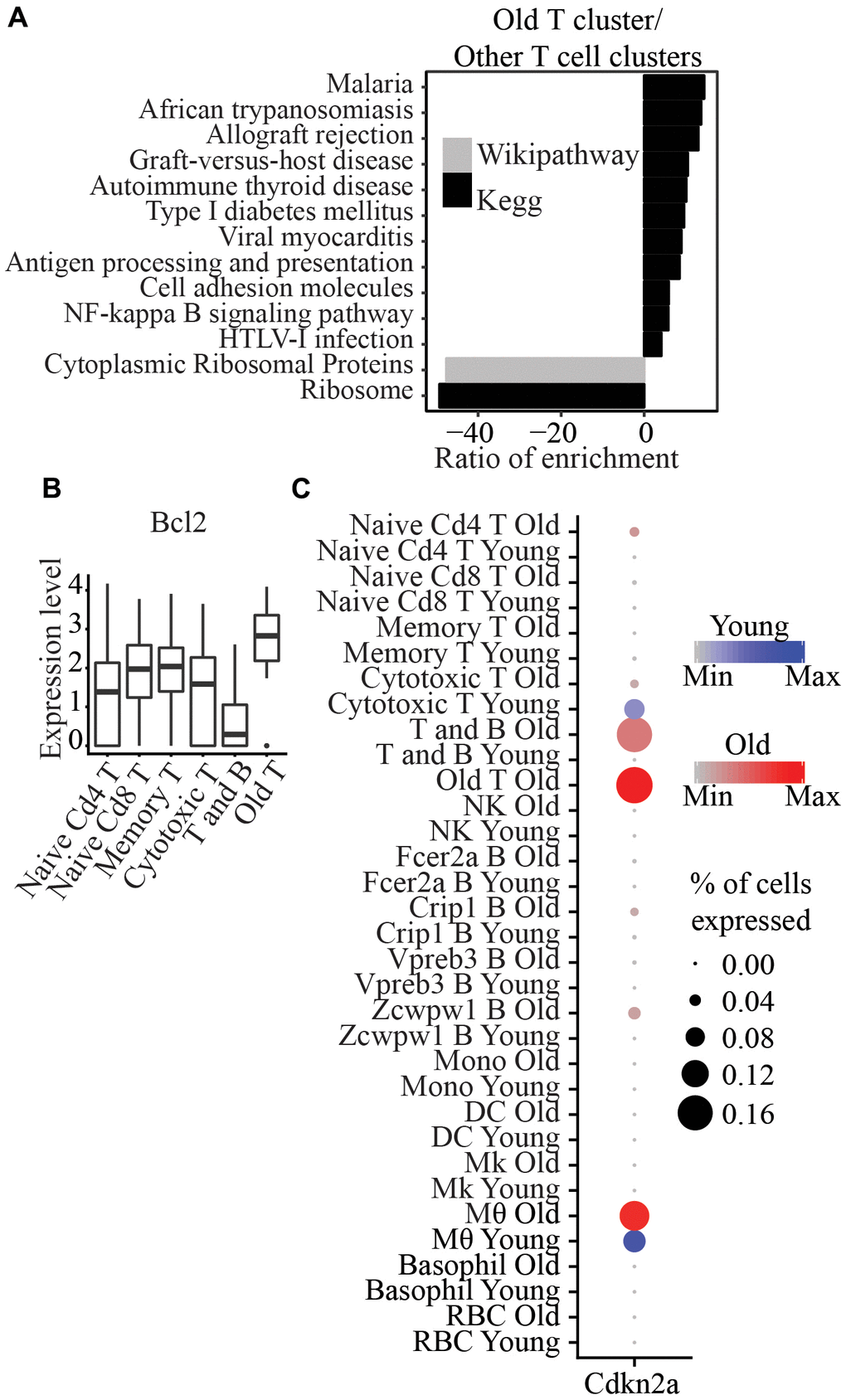
Figure 4. (A) Wikipathway and Kegg pathway analysis of differentially expressed genes between Old T cluster and other T cell clusters. (B) Bcl2 is significantly higher in Old T cluster compared to other T cell clusters. (C) Cdkn2a is significantly higher in Old T cluster compared to all other clusters.
Immunosenescence has been widely implicated in aging. Cellular senescence, an irreversible cell cycle arrest biological process, was first discovered in fibroblast, and senescent cells can be identified by a collection of markers, which include, for example, upregulation of Cdkn1a and Cdkn2a. We observed a significant increase in the percentage of old blood cells (6.3%) expressing Cdkn1a and/or Cdkn2a compared to young cells (Fisher’s Exact Test p<2.2x10-16) (Figure 5A). However, we did not detect any cell type with significant differences in the expression of these two genes between ages. Still, we did observe a moderate (p=0.066) increase in the number of Cdkn1a expressing cells in naïve Cd4 T cells (Figure 5B).
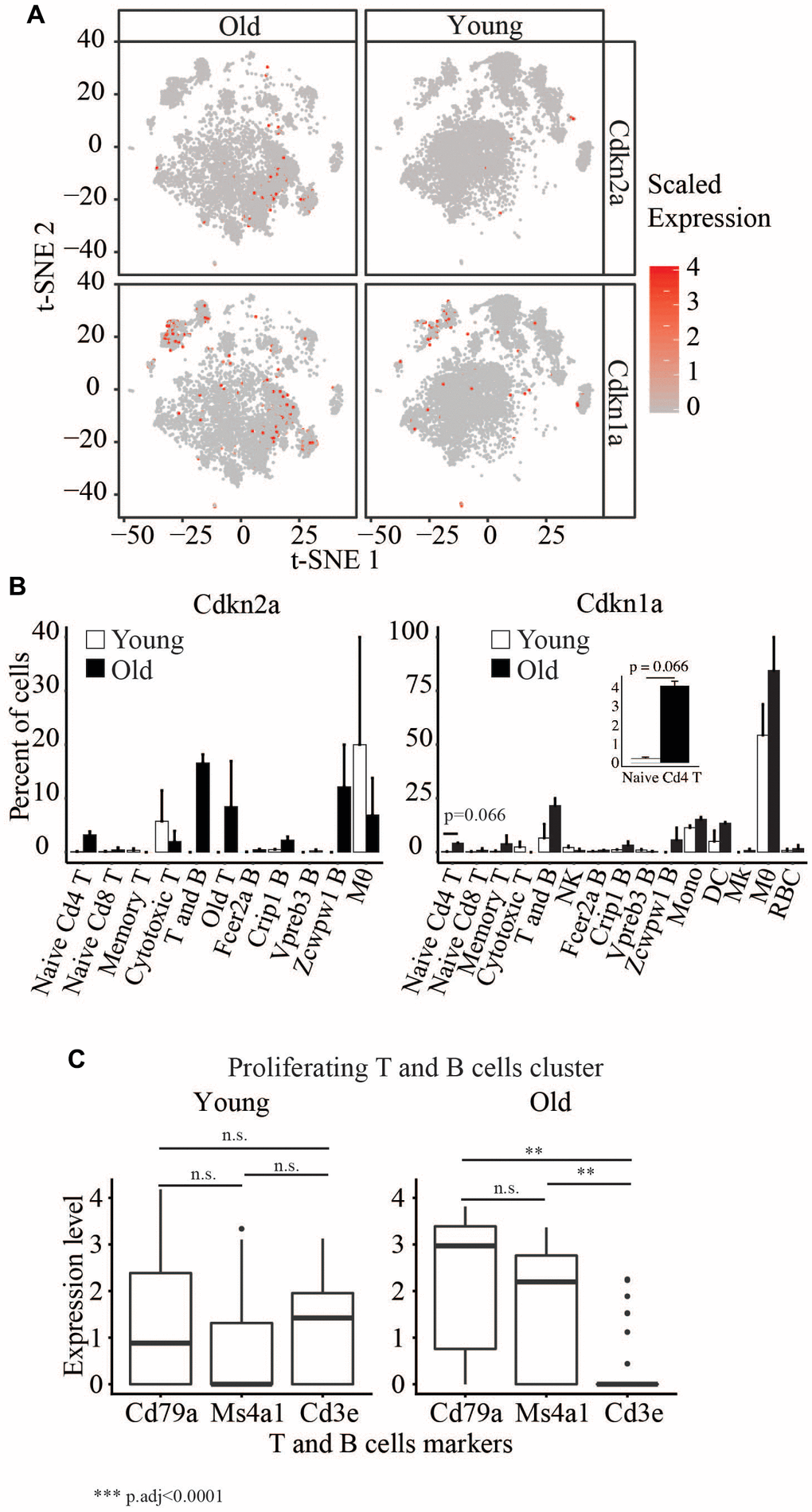
Figure 5. (A) t-SNE visualization of cells with different Cdkn2a and Cdkn1a expression levels in young and old. (B) Barplots showing the percent of cells in young and old expression Cdkn2a and Cdkn1a. (C) Boxplots showing the expression levels of Cd79a, Ms4a1 and Cd3e in the proliferating T and B cells cluster.
We also detected a cluster of old and young proliferating cells that consists of T and B cells (Figure 5C and Supplementary Table 1). The cells from old mice in this cluster are enriched in B cells but not T cell markers, whereas we saw a similar enrichment of T and B cells in young mice. This suggests a decreased number of proliferating T cells in the old mice.
Discussion
Transcriptional changes with age in specific immune cell types or lymphoblastoid cell lines in humans have been investigated in previous studies [16, 17]. scRNA-seq is a powerful technique that can dissect the transcription profiles of thousands or more cells from the same sample [18]. Other studies have used single-cell RNA-seq on the peripheral blood to identify and reclassify cell types and to investigate the changes in immune function of specific cell types upon stimulation with age in humans and mice, respectively [15, 19, 20]. Here, we profiled 14588 single cells from the mouse blood and assessed the differences between young and old mice.
We identified 17 clusters of cells that we further assigned to different cell types. Immune cells are typically identified using flow cytometry through cell surface markers [21]. We can assign various types of T cells but not B cells using markers identified from available studies. Further work is needed to link cell surface markers, immune function, and marker genes from the transcriptome of a subset of B cells in the peripheral blood.
We found a T cell cluster specific to only old mice; these cells have significantly higher Bcl2 expression than all other T cell clusters. Bcl2 is an anti-apoptotic factor that regulates cell death, and it has been used as a target for senolytic drugs to clear senescent cells. This cluster also exhibits a significantly higher expression of Cdkn1a, a senescent marker and an anti-proliferative marker found in T cells [22, 23]. Senescent T cells have been previously observed in aged samples and patients with cancer or autoimmune disease [24]. Previous studies showed that T cells that are antigen-induced to cell death in vitro could be rescued by the expression of p16 [25], that p16 reduces naïve T cell and memory T cell proliferation, and that its deletion in T cell lineage attenuates aging phenotypes associated with T cells [26]. Among other T cells, memory T cells exhibit higher expression of Bcl2 [27]. Loss of Cd28 has also been reported in replicative senescent cells [28]. Notably, we only detected this in old mice. It has been previously suggested that an accumulation of CD28- T cells in older individuals may be due to increased proliferation instead of the cells being more resistant to apoptosis with age [29, 30]. However, the higher expression of Cdkn1a and Bcl2 genes and a lower expression of Cd28 in the old T cluster in our dataset may indicate that these cells are senescent and more resistant to apoptosis than other T cells.
We generally observed enrichment of chemokine signaling and antigen-presenting pathways with age. This may indicate that the immune system works at a higher capacity in old mice. There is also an increased number of cells with Cdkn2a and Cdkn1a expression in old age, but the increase is not specific to any cell type, suggesting a general increase of possibly senescent cells.
The changes in the cell type composition that we observed are consistent with previous studies, such as increased B cells in old peripheral blood in mice and decreased number of NK cells and T cells in aged mouse blood [3, 5, 31]. In addition, we also observed an increase in Mk cells with age. To our knowledge, the change in the number of this cell type has not been documented before, but platelet counts were shown to be relatively stable until old age when it starts decreasing in humans. On the other hand, platelet count was shown to increase in 18 months old mice compared to young mice [32], but another study showed that the count does not change in 24-25 months mice compared to young [33].
One limitation of our study is that it only includes females. However, sex differences are well-known in aging and age-related diseases, and females have been historically underrepresented in mouse aging studies. Importantly, some longevity interventions work better in females.
Finally, targeting senescent cells using genetic approaches has been shown to ameliorate the aging phenotype [34, 35]. More recently, senolytics drugs are being identified or developed to target apoptotic pathways because senescent cells are known to be apoptosis-resistant [34]. Therefore, the Bcl2+ old T cells that we identified in old mice can potentially be targeted pharmacologically to ameliorate the phenotypes associated with the aging of the immune system.
Materials and Methods
Use of animals
C57BL/6 female 4- and 24 month mice were obtained from the National Institute of Aging (NIA). They were fed ad libitum and kept in standard housing conditions, and all procedures were approved by the Brown University Institutional Animal Care and Use Committees (IACUC) committee.
Isolation of immune cells from the peripheral blood
Blood samples were drawn from the heart of four young and four old mice. Samples from two mice within each age group were pooled, resulting in two pooled young and two pooled old samples. Pooling allowed us to increase the number of mice used in this study and simultaneously reduce the variance due to potential outliers. One limitation is that each individual sample does not represent an individual mouse. Subsequently, the pooled samples were diluted 1:1 with PBS+2% FBS and loaded into SepMate-15™ tubes with 4.5 mL of Lymphoprep™ with a density of 1.077g/mL (StemCell Technologies). Cells were centrifuged at 1200x g for 20min at 4° C. The top layer was collected and centrifuged at 300xg for 8 min. Supernatant was removed, and cells were resuspended in PBS + 2% FBS, and this step was repeated. Subsequently, red blood cell lysis was performed using the Red Blood Cell Lysis Solution (Miltenyi Biotec) following the manufacturer’s protocol, that includes the washing step.
Single-cell library construction
Single-cell RNA-seq protocol was performed using the Chromium™ Single Cell 3’ reagent kit v2 chemistry, and cells were loaded on a GemCode Single Cell Instrument (10x Genomics). Approximately 5000 single cells were targeted from each sample. Libraries were sequenced on Illumina HiSeq 2500 with the custom configuration of read 1 (26bp) and read 2 (98bp), i7 index (8bp), and i5 index (0bp).
Single-cell RNA-seq alignment, UMI counting, and analysis
The Cell Ranger Single Cell Software Suite 2.1.0 was used to perform single cells demultiplexing and UMI counting (https://support.10xgenomics.com/single-cell-gene-expression/software/overview/welcome). The transcriptome reference used was mm10. Subsequently, the duplicates from young and old mice, respectively, were aggregated using Cellranger aggr. Cells with more than 500 genes detected and less than 10% of mitochondrial reads were included in the downstream analysis using Seurat 2.3.0. Young and old UMI counts were merged using the MergeSeurat function. Default parameters of Seurat were used in the analysis unless otherwise stated. 1080 highly variable genes were used as an input for PCA. T-SNE projection and clustering analysis (dims=1:30 and resolution=0.4) were performed using Seurat. Markers genes for each cluster were found using the FindConservedMarkers function, and the changes with age within each cell type were identified using the FindMarkers function.
Number and percent of cells in each cell type cluster for each age: Basophil: N = (30 young, 18 old), percent = (0.4% young, 0.3% old); Crip1 B: N = (453 young, 2281 old), percent = (5.8% young, 33.4% old); Cytotoxic T: N = (82 young, 118 old), percent = (1.1% young, 1.7% old); DC: N = (46 young, 74 old), percent = (0.6% young, 1.1% old); Fcer2a B: N = (3647 young, 1914 old), percent = (47.0% young, 28.0% old); Macrophage: N = (21 young, 30 old), percent = (0.3% young, 0.4% old); Memory T: N = (247 young, 16 old), percent = (3.2% young, 0.2% old); Mk: N = (1 young, 274 old), percent = (0.0% young, 4.0% old); Mono: N = (360 young, 737 old), percent = (4.6% young, 10.8% old); NK: N = (321 young, 91 old), percent = (4.1% young, 1.3% old); Naive Cd4 T: N = (1261 young, 193 old), percent = (16.3% young, 2.8% old); Naive Cd8 T: N = (755 young, 179 old), percent = (9.7% young, 2.6% old); Old T: N = (0 young, 66 old), percent = (0.0% young, 1.0% old); RBC: N = (159 young, 182 old), percent = (2.0% young, 2.7% old); T and B: N = (39 young, 31 old), percent = (0.5% young, 0.5% old); Vpreb3 B: N = (335 young, 249 old), percent = (4.3% young, 3.6% old); Zcwpw1 B: N = (2 young, 376 old), percent = (0.0% young, 5.5% old).
Statistical analysis
All Student’s t-tests were performed in R, and p-values were adjusted using the Benjamini-Hochberg method. Barplots are represented as means with SEM. The top and bottom bounds of boxplots correspond to the 75 and 25th percentile, respectively. At the single-cell level, the statistical analyses were carried out with the cells as replicates. At the pooled samples level, two pooled samples are available for each age group; hence there are two replicates per group.
Data availability
Single-cell RNA-seq of old and young peripheral blood duplicates are accessible through GEO with the accession number GSE120505.
Author Contributions
Y.T. performed the sample collection, the single-cell RNA-seq assays, the data analysis, and the writing and editing of the manuscript. S.J.H. assisted with the data analysis and the editing of the manuscript. A.E.W. provided biological material and contributed to data interpretation. N.N. designed the experiments, supervised the data analysis and interpretation, and contributed to writing and editing the manuscript.
Acknowledgments
Part of this research was conducted using computational resources at the Center for Computation and Visualization, Brown University.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Ethical Statement
All experiments were approved by the Institutional Animal Care and Use Committee, and were performed in accordance with the current PHS Policy on Humane Care and Use of Laboratory Animals, USDA: Animal Welfare Act and Regulations, and “the Guide for the Care and Use of Laboratory Animals.”
Funding
This work was supported by the IDeA grant P20GM109035 (Center for Computational Biology of Human Disease) from NIH NIGMS, grant 1R01AG050582-01A1 from NIH NIA and grant 1 UG3CA268202 from NIH NCI (SenNet Consortium) to NN.
References
- 1. Linton PJ, Dorshkind K. Age-related changes in lymphocyte development and function. Nat Immunol. 2004; 5:133–9. https://doi.org/10.1038/ni1033 [PubMed]
- 2. Weiskopf D, Weinberger B, Grubeck-Loebenstein B. The aging of the immune system. Transpl Int. 2009; 22:1041–50. https://doi.org/10.1111/j.1432-2277.2009.00927.x [PubMed]
- 3. Beli E, Duriancik DM, Clinthorne JF, Lee T, Kim S, Gardner EM. Natural killer cell development and maturation in aged mice. Mech Ageing Dev. 2014; 135:33–40. https://doi.org/10.1016/j.mad.2013.11.007 [PubMed]
- 4. Borrego F, Alonso MC, Galiani MD, Carracedo J, Ramirez R, Ostos B, Peña J, Solana R. NK phenotypic markers and IL2 response in NK cells from elderly people. Exp Gerontol. 1999; 34:253–65. https://doi.org/10.1016/s0531-5565(98)00076-x [PubMed]
- 5. Frasca D, Landin AM, Riley RL, Blomberg BB. Mechanisms for decreased function of B cells in aged mice and humans. J Immunol. 2008; 180:2741–6. https://doi.org/10.4049/jimmunol.180.5.2741 [PubMed]
- 6. Scholz JL, Diaz A, Riley RL, Cancro MP, Frasca D. A comparative review of aging and B cell function in mice and humans. Curr Opin Immunol. 2013; 25:504–10. https://doi.org/10.1016/j.coi.2013.07.006 [PubMed]
- 7. Martínez de Toda I, Maté I, Vida C, Cruces J, De la Fuente M. Immune function parameters as markers of biological age and predictors of longevity. Aging (Albany NY). 2016; 8:3110–9. https://doi.org/10.18632/aging.101116 [PubMed]
- 8. Effros RB. The role of CD8 T cell replicative senescence in human aging. Discov Med. 2005; 5:293–7. https://doi.org/10.1111/j.0105-2896.2005.00259.x [PubMed]
- 9. Saurwein-Teissl M, Lung TL, Marx F, Gschösser C, Asch E, Blasko I, Parson W, Böck G, Schönitzer D, Trannoy E, Grubeck-Loebenstein B. Lack of antibody production following immunization in old age: association with CD8(+)CD28(-) T cell clonal expansions and an imbalance in the production of Th1 and Th2 cytokines. J Immunol. 2002; 168:5893–9. https://doi.org/10.4049/jimmunol.168.11.5893 [PubMed]
- 10. Goronzy JJ, Fulbright JW, Crowson CS, Poland GA, O’Fallon WM, Weyand CM. Value of immunological markers in predicting responsiveness to influenza vaccination in elderly individuals. J Virol. 2001; 75:12182–7. https://doi.org/10.1128/JVI.75.24.12182-12187.2001 [PubMed]
- 11. Abe E, Yamamoto M, Taguchi Y, Lecka-Czernik B, O’Brien CA, Economides AN, Stahl N, Jilka RL, Manolagas SC. Essential requirement of BMPs-2/4 for both osteoblast and osteoclast formation in murine bone marrow cultures from adult mice: antagonism by noggin. J Bone Miner Res. 2000; 15:663–73. https://doi.org/10.1359/jbmr.2000.15.4.663 [PubMed]
- 12. Conboy IM, Conboy MJ, Wagers AJ, Girma ER, Weissman IL, Rando TA. Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature. 2005; 433:760–4. https://doi.org/10.1038/nature03260 [PubMed]
- 13. Loffredo FS, Steinhauser ML, Jay SM, Gannon J, Pancoast JR, Yalamanchi P, Sinha M, Dall’Osso C, Khong D, Shadrach JL, Miller CM, Singer BS, Stewart A, et al. Growth differentiation factor 11 is a circulating factor that reverses age-related cardiac hypertrophy. Cell. 2013; 153:828–39. https://doi.org/10.1016/j.cell.2013.04.015 [PubMed]
- 14. Bahar R, Hartmann CH, Rodriguez KA, Denny AD, Busuttil RA, Dollé ME, Calder RB, Chisholm GB, Pollock BH, Klein CA, Vijg J. Increased cell-to-cell variation in gene expression in ageing mouse heart. Nature. 2006; 441:1011–4. https://doi.org/10.1038/nature04844 [PubMed]
- 15. Martinez-Jimenez CP, Eling N, Chen HC, Vallejos CA, Kolodziejczyk AA, Connor F, Stojic L, Rayner TF, Stubbington MJT, Teichmann SA, de la Roche M, Marioni JC, Odom DT. Aging increases cell-to-cell transcriptional variability upon immune stimulation. Science. 2017; 355:1433–6. https://doi.org/10.1126/science.aah4115 [PubMed]
- 16. Reynolds LM, Ding J, Taylor JR, Lohman K, Soranzo N, de la Fuente A, Liu TF, Johnson C, Barr RG, Register TC, Donohue KM, Talor MV, Cihakova D, et al. Transcriptomic profiles of aging in purified human immune cells. BMC Genomics. 2015; 16:333. https://doi.org/10.1186/s12864-015-1522-4 [PubMed]
- 17. Harris SE, Riggio V, Evenden L, Gilchrist T, McCafferty S, Murphy L, Wrobel N, Taylor AM, Corley J, Pattie A, Cox SR, Martin-Ruiz C, Prendergast J, et al. Age-related gene expression changes, and transcriptome wide association study of physical and cognitive aging traits, in the Lothian Birth Cohort 1936. Aging (Albany NY). 2017; 9:2489–503. https://doi.org/10.18632/aging.101333 [PubMed]
- 18. Haque A, Engel J, Teichmann SA, Lönnberg T. A practical guide to single-cell RNA-sequencing for biomedical research and clinical applications. Genome Med. 2017; 9:75. https://doi.org/10.1186/s13073-017-0467-4 [PubMed]
- 19. Villani AC, Satija R, Reynolds G, Sarkizova S, Shekhar K, Fletcher J, Griesbeck M, Butler A, Zheng S, Lazo S, Jardine L, Dixon D, Stephenson E, et al. Single-cell RNA-seq reveals new types of human blood dendritic cells, monocytes, and progenitors. Science. 2017; 356:eaah4573. https://doi.org/10.1126/science.aah4573 [PubMed]
- 20. Stubbington MJT, Rozenblatt-Rosen O, Regev A, Teichmann SA. Single-cell transcriptomics to explore the immune system in health and disease. Science. 2017; 358:58–63. https://doi.org/10.1126/science.aan6828 [PubMed]
- 21. Yu YR, O’Koren EG, Hotten DF, Kan MJ, Kopin D, Nelson ER, Que L, Gunn MD. A Protocol for the Comprehensive Flow Cytometric Analysis of Immune Cells in Normal and Inflamed Murine Non-Lymphoid Tissues. PLoS One. 2016; 11:e0150606. https://doi.org/10.1371/journal.pone.0150606 [PubMed]
- 22. Erickson S, Sangfelt O, Heyman M, Castro J, Einhorn S, Grandér D. Involvement of the Ink4 proteins p16 and p15 in T-lymphocyte senescence. Oncogene. 1998; 17:595–602. https://doi.org/10.1038/sj.onc.1201965 [PubMed]
- 23. Migliaccio M, Raj K, Menzel O, Rufer N. Mechanisms that limit the in vitro proliferative potential of human CD8+ T lymphocytes. J Immunol. 2005; 174:3335–43. https://doi.org/10.4049/jimmunol.174.6.3335 [PubMed]
- 24. Chou JP, Effros RB. T cell replicative senescence in human aging. Curr Pharm Des. 2013; 19:1680–98. https://doi.org/10.2174/138161213805219711 [PubMed]
- 25. Lissy NA, Van Dyk LF, Becker-Hapak M, Vocero-Akbani A, Mendler JH, Dowdy SF. TCR antigen-induced cell death occurs from a late G1 phase cell cycle check point. Immunity. 1998; 8:57–65. https://doi.org/10.1016/s1074-7613(00)80458-6 [PubMed]
- 26. Liu Y, Johnson SM, Fedoriw Y, Rogers AB, Yuan H, Krishnamurthy J, Sharpless NE. Expression of p16(INK4a) prevents cancer and promotes aging in lymphocytes. Blood. 2011; 117:3257–67. https://doi.org/10.1182/blood-2010-09-304402 [PubMed]
- 27. Grayson JM, Zajac AJ, Altman JD, Ahmed R. Cutting edge: increased expression of Bcl-2 in antigen-specific memory CD8+ T cells. J Immunol. 2000; 164:3950–4. https://doi.org/10.4049/jimmunol.164.8.3950 [PubMed]
- 28. Effros RB. Loss of CD28 expression on T lymphocytes: a marker of replicative senescence. Dev Comp Immunol. 1997; 21:471–8. https://doi.org/10.1016/s0145-305x(97)00027-x [PubMed]
- 29. Gupta S. Molecular mechanisms of apoptosis in the cells of the immune system in human aging. Immunol Rev. 2005; 205:114–29. https://doi.org/10.1111/j.0105-2896.2005.00261.x [PubMed]
- 30. Brzezińska A, Magalska A, Szybińska A, Sikora E. Proliferation and apoptosis of human CD8(+)CD28(+) and CD8(+)CD28(-) lymphocytes during aging. Exp Gerontol. 2004; 39:539–44. https://doi.org/10.1016/j.exger.2003.09.026 [PubMed]
- 31. Xie J, Zhang J, Wu H, Tang X, Liu J, Cheng G, Li P. The influences of age on T lymphocyte subsets in C57BL/6 mice. Saudi J Biol Sci. 2017; 24:108–13. https://doi.org/10.1016/j.sjbs.2016.09.002 [PubMed]
- 32. Jones CI. Platelet function and ageing. Mamm Genome. 2016; 27:358–66. https://doi.org/10.1007/s00335-016-9629-8 [PubMed]
- 33. Jayachandran M, Karnicki K, Miller RS, Owen WG, Korach KS, Miller VM. Platelet characteristics change with aging: role of estrogen receptor beta. J Gerontol A Biol Sci Med Sci. 2005; 60:815–9. https://doi.org/10.1093/gerona/60.7.815 [PubMed]
- 34. Kirkland JL, Tchkonia T, Zhu Y, Niedernhofer LJ, Robbins PD. The Clinical Potential of Senolytic Drugs. J Am Geriatr Soc. 2017; 65:2297–301. https://doi.org/10.1111/jgs.14969 [PubMed]
- 35. Baker DJ, Wijshake T, Tchkonia T, LeBrasseur NK, Childs BG, van de Sluis B, Kirkland JL, van Deursen JM. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature. 2011; 479:232–6. https://doi.org/10.1038/nature10600 [PubMed]