



Introduction
A number of stresses can cause cells to enter a non-dividing state called cellular senescence [1]. These stresses include repeated cell division, expression of activated oncogenes, oxidative stress, and irradiation. The cellular senescence pathway functions as an anti-tumor mechanism in mammals, and is regulated by the tumor-suppressor proteins p53 and Rb. Senescence of cells during aging may contribute to mammalian aging phenotypes by limiting the ability of stem cell populations to replenish tissues. Several Drosophila tissues are maintained by dividing stem cell populations, including the gonads [2], the gut [3,4] and the malpighian tubule (equivalent to mammalian kidney) [5], however it is currently unknown whether alterations in these stem cell populations during aging has an effect on Drosophila life span.
Apoptosis (programmed cell death) is also implicated in mammalian and Drosophila aging phenotypes.
Regulated apoptosis is required for normal homeostasis in dividing tissues such as the gut and hematopoetic system, and abnormal apoptotic events have been observed in muscle and other tissues during mammalian aging [6]. In addition, apoptosis is implicated in several human aging-related diseases, for example neurodegenerative diseases such as Alzheimer's disease and Parkinson's disease [7]. In aging Drosophila, abnormal apoptotic events have been observed in muscle and fat tissue [8], but the extent to which apoptosis (or cellular senescence) might modulate Drosophila life span remains largely unknown. Several genes that can affect apoptosis (and senescence) have been found to affect Drosophila life span, including DPOSH, MnSOD and p53 [9-12]. In mammals hyperactive p53 can produce an accelerated-aging-like phenotype [13], and in Drosophila a dominant-mutant p53 transgene can inhibit insulin-like signaling and cause increased life span [14]. However, the extent to which these effects on life span might be mediated by alterations in apoptosis and/or cellular senescence pathways is largely unknown. The potential importance of the cellular senescence and apoptosis pathways in modulating life span prompted a screen of additional genes implicated in these pathways for life span effects in the fly.
Conditional gene expression systems have several advantages for studies of aging: for example with the Tet-on system the expression of transgenes is triggered by feeding the flies the drug doxycycline, and with the Geneswitch system transgene expression is triggered using the drug RU486/Mifepristone [15-17]. These conditional systems allow for transgene expression to be limited to specific life cycle stages such as development or adulthood. Moreover, these systems provide powerful controls for genetic background effects on life span, since the control and gene-over-expressing animals have identical genetic backgrounds and differ only in the presence or absence of the drug. It is often desirable to over-express a gene in all the tissues of the fly, for example when screening genes for possible life span effects. We have recently reported the generation of a Geneswitch system driver (called "Act-GS-255B"), which contains multiple inserts of a construct in which the promoter of the cytoplasmic actin gene Actin5C is used to drive expression of the Geneswitch transcription factor [16]. Here the Act-GS-255B driver is further characterized using a UAS-GFP reporter, and we report that it is truly tissue-general in both the larval and adult stages. The tissue-general driver facilitated the screening of senescence and apoptosis regulatory genes for life span effects.
Results
Characterization of Geneswitch drivers in adult flies using the UAS-GFP reporter
To facilitate the screen of apoptosis and senescence-regulatory genes for life span effects, several Geneswitch system drivers were characterized for their tissue-specificity of transgene activation using a UAS-GFP reporter, both in adult flies and during larval development. The UAS-GFP reporter employed was "UAS-ultraGFP" which contains multiple copies of a UAS-eGFP construct, and yields particularly high levels of GFP expression [18]. Three Geneswitch system drivers were characterized: The Act-GS-255B driver strain contains multiple inserts of a construct in which the promoter from the cytoplasmic actin gene Actin 5C drives Geneswitch, and is expected to yield tissue-general expression [16]. The Elav-GS driver contains Geneswitch under control of the Elav gene promoter and produces nervous-system-specific expression [19]. Finally the whole-body fat-body Geneswitch driver strain ("WB-FB-GS") contains both a head fat-body driver (S1-32) and a body-fat-body driver (S1-106) [20-22], and is expected to yield expression in the fat-body tissue throughout the animal. The three driver strains were crossed to the UAS-ultraGFP reporter strain to produce adult progeny containing both the driver and reporter constructs, and the flies were cultured in the presence and absence of drug for two weeks. GFP expression was scored in live adult flies as well as in several dissected tissues (Figure 1). The Act-GS-255B driver was found to yield tissue-general expression of the UAS-ultraGFP reporter in adult flies. In whole adults, GFP expression was observed throughout the body of both males and females, with greater expression levels observed in females relative to males. Similarly with heads dissected in half and bodies dissected in half, expression was observed in all tissues, including abundant expression in nervous system, muscle (including flight muscle), and fat-body tissue. Note that flight muscle in male has lower expression than flight muscle in female, however inspection of the GFP-only image for male flight muscle (inset) reveals expression throughout this tissue. Abundant expression was also observed throughout dissected gut tissue, ovary and testes. The expression level was greater in some regions of the gut than others, however all regions of the gut exhibited staining, as revealed by inspection of the GFP-only images (inset). All tissues observed showed significant GFP expression, and therefore we conclude that Act-GS-255B yields truly tissue-general expression in adult flies. The WB-FB-GS driver produced GFP expression in the head-fat-body and body-fat-body tissues, as expected, as well as in the gut and testes, and very faint expression in ovary; there was no detectable expression in nervous, muscle, or other tissues. Notably, the expression in adult male head fat body was much reduced relative to female head fat body, consistent with recent characterization of the fat body drivers using a LacZ reporter [17]. Finally, the Elav-GS driver produced abundant expression in the brain and ventral nerve cord, as expected, and there was no detectable expression in any other tissues; for example, the muscle, gut and gonads were clearly negative. Note the GFP-only image for the gut (inset) shows a lack of expression. The Elav-GS driver was found to produce similar levels of UAS-GFP reporter expression in male versus female in our experiments.
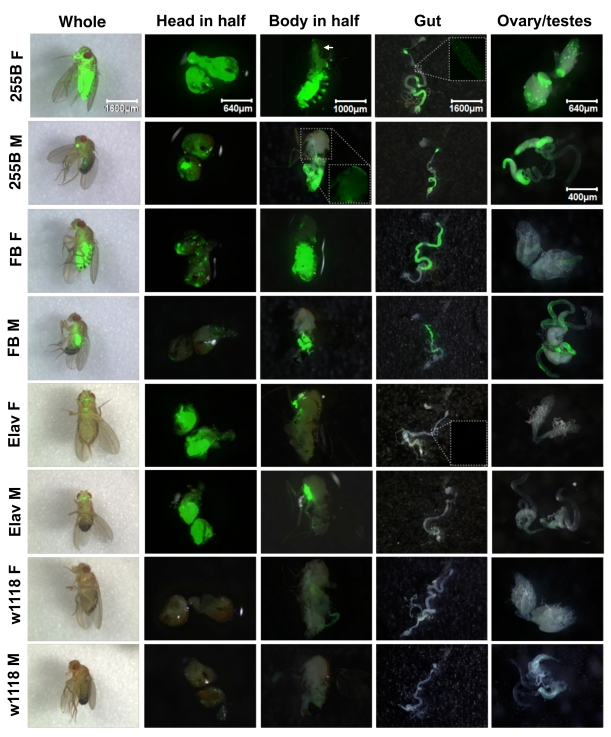
Figure 1. Expression pattern produced by Geneswitch drivers and UAS-GFP reporter in adult flies. The indicated GeneSwitch
drivers Act-GS-255B ("255B"), Elav-GS ("Elav") and WB-FB-GS ("FB") were
crossed to the UAS-ultraGFP reporter and adult progeny containing both
constructs were scored for GFP expression in various tissues. Control
flies were generated by crossing UAS-ultraGFP to white1118strain flies to produce progeny containing only UAS-ultraGFP. Age-synchronized
flies were cultured in the presence and absence
of the drug RU486 for two weeks prior to assay, and GFP expression was
scored in whole adult flies and dissected tissues, as indicated. Each
image is the overlay of the visible light and GFP images. Insets show
details of the regions boxed in white, GFP image only. M = male, F =
female. Pictures were taken at the magnification of 20X, 50X, 32X, 20X,
50X, and 80X, for whole fly, head in half, body in half, gut, ovary, and
testes, respectively. The white arrow indicates a region of 255B Female
flight muscle that is obscured by a fragment of cuticle.
Characterization of Geneswitch drivers in larvae using the UAS-GFP reporter
The Geneswitch driver strains were also scored for expression patterns in 3rd instar larvae and dissected tissues (Figure 2). The Act-GS-255B driver was found to yield tissue-general expression, including abundant expression throughout the body of whole 3rd instar larvae, as well as in dissected brain, gut, salivary gland, imaginal discs and fat-body tissues; all tissues observed showed abundant GFP expression (Figure 2A). The inset for the Act-GS-255B 3rd instar larval brain shows detail of the GFP-only image, and indicates that expression was present throughout the brain, with higher-level expression in a subset of cells. The WB-FB-GS driver was found to drive abundant expression in salivary gland and anterior midgut, but notably no expression in any other larval tissues including larval fat-body. Finally the Elav-GS driver produced abundant expression in larval nervous system and no detectable expression in any other larval tissues. The inset for the Elav-GS 3rd instar larval brain shows detail of the GFP-only image, and shows that expression was present throughout the brain, with higher-level expression in a subset of cells. Notably this subset of cells was different from that observed above with Act-GS-255B. Each of the three drivers was found to produce similar patterns of expression in 1st and 2nd instar larvae as well (Figure 1B). When the Act-GS-255B driver was induced using dilutions of RU486 drug in the culture media, it produced a dose-response of GFP expression in 3rd instar larvae (Figure 1D), as well as in adult flies (data not shown).
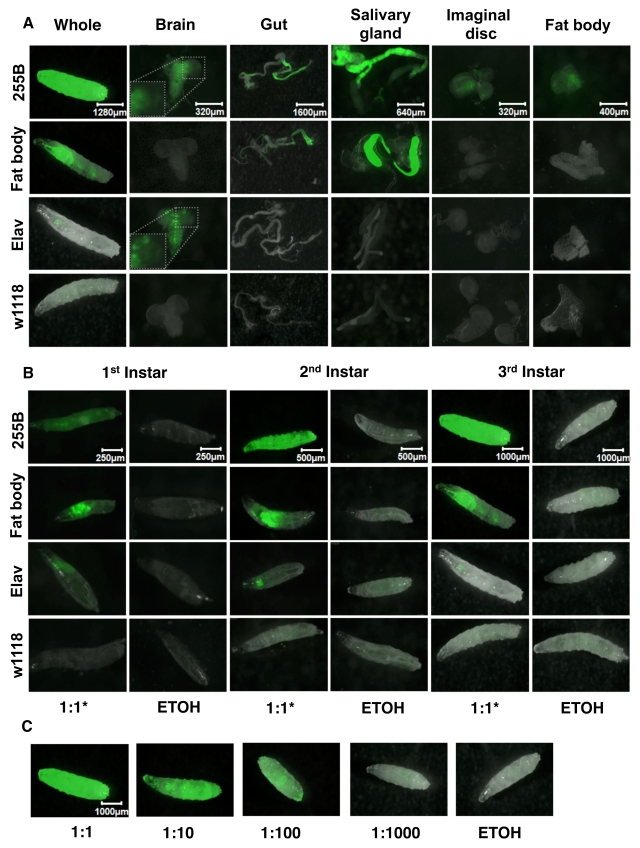
Figure 2. Expression pattern produced by GeneSwitch drivers and UAS-GFP reporter in larvae. The crosses are the same as
Figure 1, but larvae were cultured in the presence and absence of drug in
the food, from hatching to the indicated developmental stage. A.
Expression patterns in 3rd instar larvae and dissected tissues.
For the Elav-GS driver ("Elav") a 1:10 dilution of drug was used because of
the toxic effects of drug observed in larvae with this driver. Pictures
were taken at the magnification of 25X, 100X, 20X, 50X, 100X, 80X, for
whole larvae, brain, gut, salivary gland, imaginal discs, and fat body,
respectively. B. Expression patterns in the three larval stages. For Elav-GS
a 1:10 dilution of drug was used to avoid toxic effects. GFP pictures were
taken at the magnification of 100X, 50X, 25X, for 1st instar, 2nd
instar, and 3rd instar, respectively. C. Expression in 3rd
instar larvae using Act-GS-255B and titrations of drug. ETOH indicates the
ethanol solvent for the drug alone. Pictures were taken at the
magnification of 25X.
Effect of apoptosis and senescence-regulatory gene over-expression on life span
Fourteen apoptosis and senescence regulatory genes were chosen for analysis based on their relevance to human apoptosis and senescence pathways and the availability of reagents for Drosophila. Ras85D is a Drosophila homolog of the human oncogene Ras that encodes a GTPase involved in signal transduction.
Ras85D activated form contains an amino acid substitution that causes Ras to be constitutively active [23], and Ras85D dominant negative (DN) form contains an amino acid substitution that causes it to inhibit the endogenous Ras protein [23,24]. Wingless is a Drosophila homolog of the human Wnt signaling protein involved in development and tumorigenesis [25]. Pk61C is a serine/threonine protein kinase related to human PDK-1 and involved in growth signaling [26]. DIAP1 is a Drosophila member of the inhibitor of apoptosis protein (IAP) family [27]. Baculovirus p35 is a caspase inhibitor protein also related to the IAPs. Nemo (nmo) is the Drosophila homolog of a human protein kinase regulatory subunit involved in NF-kappaB signaling pathway [28]. Egfr is the Drosophila homolog of the human epidermal growth factor receptor [29]. The Drosophilapointed (pnt) gene encodes a transcription factor homologous to human Ets1 that is involved in the Ras signaling pathway. The DrosophilaMatrix metalloproteinase 2 gene (Mmp2) is involved in tissue remodeling and tumor progression and is related to a family of human matrix metalloproteinases [30]. The DrosophilaStat92E gene encodes a homolog of the human Stat transcription factor, which is a target of the Jak-Stat growth-regulatory pathway [31]. The Drosophilapuckered (puc) gene encodes a phosphatase homologous to the human VH-1 family that antagonizes JNK signaling, and heterozygous puc mutant flies have been reported to have increased stress resistance and life span [32,33]. The DrosophilaSphingosine kinase 2 (Sk2) gene encodes a lipid kinase involved in activation of protein kinase C-family signaling, and the human homolog Sphk2 is implicated in regulation of apoptosis [34]. Finally the CG14544 gene encodes a predicted methyltransferase, and the Drosophilabantam (ban) gene encodes a micro-RNA that inhibits expression of pro-apoptotic genes [35]. Each of these genes of interest was over-expressed in adult flies or during larval development, and assayed for effects on adult fly life span.
To control for any possible effects of the Geneswitch system and the RU486 drug itself, life span was assayed in flies that were the progeny of Act-GS-2555B driver crossed to either Oregon-R (Or-R) wild-type strain or to the w1118 control strain, to produce progeny containing only the driver. In these control flies, treatment with drug produced small, but statistically significant reductions in life span in both male and female adults: treatment during adulthood reduced mean life span by -4% to -10%, while treatment in larval stages reduced adult life span by -8% to -16% (Figure 3A, B; Figure 4A, B; Tables 2, 3). There were no significant increases in life span in control flies treated with RU486 in any of the replicate experiments. These data indicate that in these experiments, when the Act-GS-255B driver is present, the RU486 can cause small but significant reductions in adult life span, and this effect must be taken into account when interpreting the effects of transgene over-expression. Other studies [22], including ones from our own laboratory using the Act-GS-255B driver [36], found no negative effects of RU486 on adult fly life span. We conclude that the small negative effects observed here result from differences in the lot of RU486 drug, and/or small differences in effective concentrations due to specifics of media preparation. To confirm that the Act-GS-255B driver can produce increased life span, it was used to drive over-expression of the dominant p53 allele (p53-259H). Over-expression of p53-259H in adult flies using the ubiquitous Act-GS-255B driver produced increased median life span in females (+8%) but not males (-2.8%), and no life span increase when expressed in larvae (Table 3). These results are consistent with previous studies showing that expression of p53-259H in the adult nervous system with the Elav-GS driver can cause increased life span in females [14], and confirms that the Act-GS-255B driver can indeed produce increased life span when combined with an appropriate target gene.
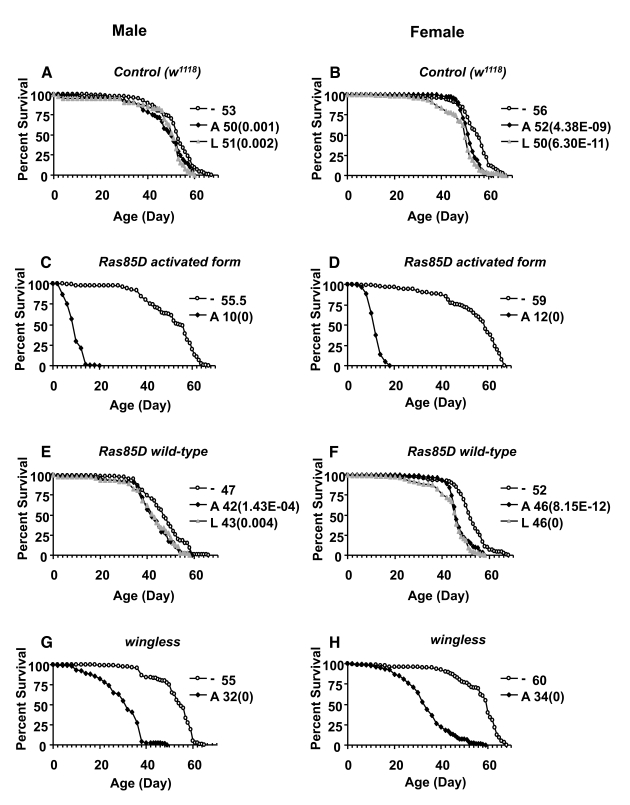
Figure 3. Effect of transgene over-expression on survival of adult flies. Apoptosis and
senescence-related genes wingless, Ras85D, and Ras85D
activated form were over-expressed during larval development or in
adults, and assayed for effects on adult life span in male and female
flies, as indicated. The life span assays were performed at 29°C. Open circles represent the no-drug control
("-"). Solid squares represent adults treated with drug ("A"). Grey
triangles represent larvae on drug ("L"). Survival curves are plotted as a
function of adult age in days. Median life span of each cohort is presented
along with p value for log rank test (in parentheses). (A, C,
E, G) male flies. (B, D, F, H) female flies. (A, B) Control
flies containing the driver and no target transgene. (C, D) Ras85D
activated form. (E, F) Ras85D wild-type. (G, H) wingless.
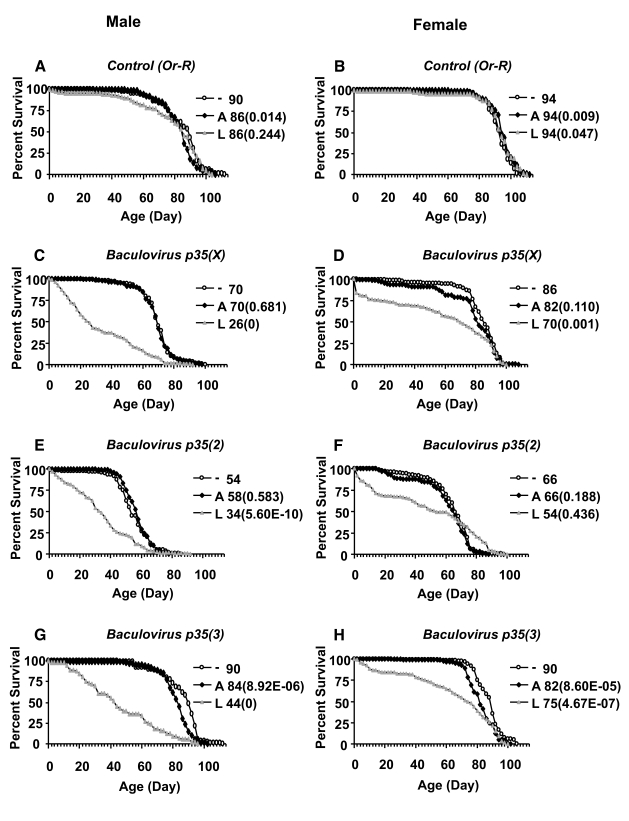
Figure 4. Effect of Baculovirus p35 over-expression on survival of adult flies. Baculovirus p35
transgenes inserted on the X chromosome, chromosome 2, and chromosome 3
were over-expressed during larval development or adult stage, as indicated.
The life span assays were performed at 25°C. Open circles represent the no-drug control ("-"). Solid squares
represent adults treated with drug ("A"). Grey triangles represent larvae
on drug ("L"). Survival curves are plotted as a function of adult age in
days. Median life span of each cohort is presented along with p
value for log rank tests (in parentheses). (A, C, E, G) male
flies. (B, D, F, H) female flies. (A,B) Control flies
containing the driver and no target transgene. (C, D) Baculovirus
p35 transgene on X chromosome. (E, F) Baculovirus p35 transgene
on second chromosome. (G, H) Baculovirus p35 transgene on
third chromosome.
Most of the genes tested by over-expression with the ubiquitous Act-GS-255B driver did not affect life span to an extent greater than the small changes observed with the control flies. However, Ras activated form transgene was lethal when expressed in larvae, and reduced both male and female life span by -80% when expressed in adults (Figure 3C, D; Table 2). Over-expression of wild-type Ras or a Ras dominant-negative allele was not lethal to larvae, and produced only small decreases (-4% to -12%) in both male and female adult life span (Figure 2 E, F; Table 2), thereby in the range of negative effects observed with control flies. Over-expression of the wingless gene was found to be lethal to male and female larvae, using two independent wingless transgenes (Table 2). Over-expression of wingless in adult flies produced significant reductions in both male and female life span: ~-42% with one wingless transgene (Figure 3 G, H) and ~-10% with the other transgene (Table 2).
Finally, the tissue-general Act-GS-255B driver was used to over-express three different transgenes encoding the caspase inhibitor Baculovirus p35, during larval development and in adult flies (Figure 4; Table 3). Over-expression of Baculovirusp35 in adult flies using the tissue-general Act-GS-255B driver produced only small decreases in life span that were within the range observed with control flies, suggesting there were no significant effects in adults. In contrast, when Baculovirus p35 was over-expressed during larval development using the tissue-general driver, it reduced the mean life span of male and female adults by -20% to -50%. Interestingly, over-expression of each of the three independent Baculovirus p35 transgenes during larval development produced an unusual biphasic-shaped survival curve in adult females (Figure 4 D, F, H), suggesting the presence of a subset of adult female flies with unchanged or even increased life span. A Gompertz-Makeham model was found to give the best fit to the life span data for females in which Baculovirus p35 was over-expressed during larval development (Figure 5; Table 4). This analysis revealed that the decrease in mean life span was due to increased age-independent mortality. When the age-independent mortality was removed and the data re-plotted, it revealed a subset of female flies with unchanged (Figure 5 B, F) or increased life span (Figure 5D).
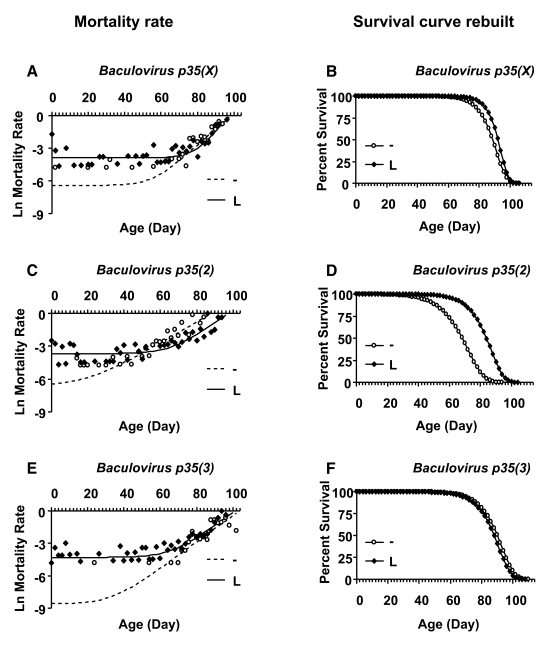
Figure 5. Mortality rate analysis of female larvae with and without Baculovirus p35 transgene expression. Open circles represent the
no-drug control ("-"). Solid squares represent larvae cultured with drug
("L"). (A, B) Baculovirus p35 transgene on X chromosome. (C,
D) Baculovirusp35 transgeneon second chromosome. (E, F) Baculovirus
p35 transgene on third chromosome. (A, C, E) Plots of
natural-log mortality rate vs. age in days. (B, D, F) The data
were fitted to the Gompertz-Makeham model, which best described the
mortality rate. The age-independent mortality was removed and the survival
curves were re-drawn using only the Gompertz components. Mortality rate
analysis showed that age-independent mortality was significantly higher for
female larvae on drug versus control for all three Baculovirus p35
lines (Table 4).
Two independent Baculovirus p35 transgenes were also over-expressed in adult flies using the head-fat-body driver S1-32, and the whole-body fat-body driver (S1-32 plus S1-106), and during larval development using the whole-body fat-body driver, however no consistent effects on life span were observed (Table 3).
The nervous system-specific Elav-GS driver was also used to over-express two baculovirus p35 transgenes. In adults the Elav-GS driver itself had little to no effect on life span, and over-expression of baculovirus p35 in adults using Elav-GS had no consistent effects on life span (Table 3). In contrast, when drug was administered to larvae, the Elav-GS driver itself was associated with significant decreases in life span in both males (~-30% to -40%) and females (~-25%), and significantly reduced the number of male adults, and no effects of the baculovirus p35 transgenes on life span could be identified in this background (Table 3). In an attempt to reduce this background toxicity and allow assay of baculovirus p35 transgenes with the Elav-GS driver in larvae, a 1:10 dilution of drug was used. Under these conditions the life span reductions caused by drug in males and females were smaller (~-2% to -12%), and the number of males obtained was approximately normal, however no increases in life span were observed upon over-expression of baculovirus p35 (Table 3).
The muscle-specific MHC-GS driver was used to drive over-expression of several transgenes in adult flies, however the MHC-GS driver itself was found to cause a significant RU486-dependent decrease in life span in both males and females (~-20% to -30%), and none of the target transgenes tested produced a significant life span increase in this background (Table 3).
Table 1. Starting Stocks
| St# | Genotype | Notes | Abbreviation |
| 1 | w; GS-Actin255-B:+ | Ubiquitous GeneSwitch 255B Driver | 255B |
| 2 | w; GS-Actin255-A;+ | Ubiquitous GeneSwitch 255A Driver | 255A |
| 3 | w; P{Switch}bun[Switch 1-32];+ | GeneSwitch Head Fat Body Driver | S32 |
| 4 | w; P{Switch}S1-106 P{Switch}bun[Switch 1-32];+ | GeneSwitch Head & Thorax-Abdomen Fat Body Driver | S106 S32 |
| 5 | yw; +; GS-Elav | GeneSwitch Elav Driver | Elav |
| 6 | yw; Sp/CyO,FLP.lacZ; MHC:GS | GeneSwitch Muscle Driver | Sp/CyO, MHC |
| 7 | Oregon R ( +; +; +) | wild type | |
| 8 | w1118; +; + | wild type | |
| 9 | P{UAS.p35.H}BH3,w*;+;+ | UAS-p35 on chromosome 1 | p35 |
| 10 | w*; P{UAS.p35.H}BH1;+ | UAS-p35 on chromosome 2 | p35 |
| 11 | w*; +; P{UAS.p35.H}BH2 | UAS-p35 on chromosome 3 | p35 |
| 12 | w1118; +; P{UAS-Ras85D.V12}TL1 | UAS-Ras85D activated form | Ras act |
| 13 | w*; P{UAS-Ras85D.K}5-1;+ | UAS-Ras85D WT form | Ras WT |
| 14 | P{UAS-Ras85D.N17}TL1, w1118; +; + | UAS-Ras85D DN form | Ras DN |
| 15 | w*; P{UAS-wg.H.T:HA1}3C;+ | UAS-wg on chromosome 2 | wga |
| 16 | w*; +; P{UAS-wg.H.T:HA1}6C | UAS-wg on chromosome 3 | wgb |
| 17 | y1 w67c23; +;P{EPgy2}EY04093 | EP-Pk61C | Pk61Ca |
| 18 | w; +; P{EP}Pk61CEP3644/TM6,Tb | EP-Pk61C | Pk61Cb |
| 19 | w*; +; P{UAS-DIAP1.H}3 | UAS-DIAP1 | DIAP1 |
| 20 | y1 w67c23; P{EPgy2}EY00935 | EP-nmo | nmo |
| 21 | y1 w*; +; P{UAS-Egfr.B}32-26-1 | UAS-Egfr | Egfr |
| 22 | y1 w67c23; +; P{EPgy2}pntEY03254 | EP-pnt | pnt |
| 23 | y1 w67c23; P{EPgy2}Mmp2EY08942/CyO; + | EP-Mmp2 | Mmp2 |
| 24 | y1 w67c23; +; P{EPgy2}Stat92EEY14209/TM3, Sb1 Ser1 | EP-Stat92E | Stat |
| 25 | w*; +; P{EPgy2}pucEY09772/TM6C | EP-puc | puc |
| 26 | y1 w67c23; +; P{EPgy2}scramb2EY01180 | EP-Sk2 | Sk2 |
| 27 | y1 w67c23; +; P{EPgy2}EY06207 | EP-ban | ban |
| 28 | w1118; +; PBac{WH}CG14544f01091/TM6B, Tb1 | XP-CG14544 | CG14544 |
| 29 | w1118; +; P{GUS-p53.259H}3.1 | UAS-p53 point mutation | p53.259H |
Table 2. Life span data of apoptosis-related gene experiments, with means, standard deviations, medians, percent change in mean and median, and log rank p value.
| CrossMxF | RU486 | Genotype | Sex | N | Meana | Median | %Change in Mean | %Change in Median | Log Rank p Value |
| Exp1 Life span assay using GS255B driver at 29C | |||||||||
| 8-1 | - | w/Y; 255B/+; + | M | 115 | 51.53±8.66 | 53 | --------- | --------- | --------- |
| A | w/Y; 255B/+; + | M | 120 | 47.29±11.06 | 50 | -8.23 | -5.66 | 0.001 | |
| L | w/Y; 255B/+; + | M | 40 | 47.28±12.15 | 51 | -8.26 | -3.77 | 0.002 | |
| - | w/w; 255B/+; + | F | 128 | 54.18±8.26 | 56 | --------- | --------- | --------- | |
| A | w/w; 255B/+; + | F | 120 | 51.74±3.89 | 52 | -4.5 | -7.14 | 4.38E-09 | |
| L | w/w; 255B/+; + | F | 120 | 48.18±8.38 | 50 | -11.07 | -10.71 | 6.30E-11 | |
| 12-1 | - | w/Y; 255B/+; Ras act/+ | M | 122 | 51.11±11.58 | 55.5 | --------- | --------- | --------- |
| A | w/Y; 255B/+; Ras act/+ | M | 128 | 9.45±3.42 | 10 | -81.5 | -81.98 | 0 | |
| L | w/Y; 255B/+; Ras act/+ | M | 0 | NA | NA | --------- | --------- | --------- | |
| - | w/w*; 255B/+; Ras act/+ | F | 123 | 54.19±13.26 | 59 | --------- | --------- | --------- | |
| A | w/w*; 255B/+; Ras act/+ | F | 123 | 12.11±2.8 | 12 | -77.64 | -79.66 | 0 | |
| L | w/w*; 255B/+; Ras act/+ | F | 0 | NA | NA | --------- | --------- | --------- | |
| 13-1 | - | w/Y; 255B/Ras WT;+ | M | 124 | 46.65±9.01 | 47 | --------- | --------- | --------- |
| A | w/Y; 255B/Ras WT;+ | M | 122 | 42.3±9.13 | 42 | -9.32 | -10.64 | 1.43E-04 | |
| L | w/Y; 255B/Ras WT;+ | M | 47 | 42.06±10.37 | 43 | -9.84 | -8.51 | 0.004 | |
| - | w/w*; 255B/Ras WT;+ | F | 126 | 51.31±8.64 | 52 | --------- | --------- | --------- | |
| A | w/w*; 255B/Ras WT;+ | F | 126 | 46.84±5.17 | 46 | -8.71 | -11.54 | 8.15E-12 | |
| L | w/w*; 255B/Ras WT;+ | F | 118 | 43.66±8.85 | 46 | -14.91 | -11.54 | 0 | |
| 1-14 | - | Ras DN, w/Y; 255B/+; + | M | 127 | 47.89±9.88 | 50 | --------- | --------- | --------- |
| A | Ras DN, w/Y; 255B/+; + | M | 123 | 43.64±6.87 | 44 | -8.87 | -12 | 5.78E-08 | |
| L | Ras DN, w/Y; 255B/+; + | M | 79 | 44.76±12.49 | 48 | -6.54 | -4 | 0.14 | |
| - | Ras DN, w/w; 255B/+; + | F | 121 | 51.65±14.25 | 57 | --------- | --------- | --------- | |
| A | Ras DN, w/w; 255B/+; + | F | 125 | 51.82±8.3 | 53 | 0.32 | -7.02 | 5.09E-04 | |
| L | Ras DN, w/w; 255B/+; + | F | 125 | 45.39±13.16 | 49 | -12.12 | -14.04 | 1.98E-09 | |
| 15-1 | - | w/Y; 255B/wga; + | M | 130 | 52.56±8.37 | 55 | --------- | --------- | --------- |
| A | w/Y; 255B/wga; + | M | 122 | 29.83±9.32 | 32 | -43.25 | -41.82 | 0 | |
| L | w/Y; 255B/wga; + | M | 0 | NA | NA | --------- | --------- | --------- | |
| - | w/w*; 255B/wga; + | F | 122 | 55.78±12.01 | 60 | --------- | --------- | --------- | |
| A | w/w*; 255B/wga; + | F | 125 | 33.61±10.55 | 34 | -39.75 | -43.33 | 0 | |
| L | w/w*; 255B/wga; + | F | 0 | NA | NA | --------- | --------- | --------- | |
| 16-1 | - | w/Y; 255B/+;wgb/+ | M | 124 | 52.31±8.81 | 56 | --------- | --------- | --------- |
| A | w/Y; 255B/+;wgb/+ | M | 131 | 45.02±8.04 | 47 | -13.94 | -16.07 | 0 | |
| L | w/Y; 255B/+;wgb/+ | M | 0 | NA | NA | --------- | --------- | --------- | |
| - | w/w*; 255B/+;wgb/+ | F | 120 | 51.29±10.38 | 53 | --------- | --------- | --------- | |
| A | w/w*; 255B/+;wgb/+ | F | 123 | 47.55±7.23 | 49 | -7.29 | -7.55 | 3.24E-10 | |
| L | w/w*; 255B/+;wgb/+ | F | 0 | NA | NA | --------- | --------- | --------- | |
| 17-1 | - | w/Y; 255B/+; Pk61Ca/+ | M | 122 | 47.9±9.67 | 47 | --------- | --------- | --------- |
| L | w/Y; 255B/+; Pk61Ca/+ | M | 21 | 42.81±13.89 | 47 | -10.63 | 0 | 0.224 | |
| - | w/yw; 255B/+; Pk61Ca/+ | F | 127 | 49.82±16.97 | 56 | --------- | --------- | --------- | |
| L | w/yw; 255B/+; Pk61Ca/+ | F | 121 | 50.24±11.55 | 52 | 0.84 | -7.14 | 8.57E-04 | |
| 18-1 | - | w/Y; 255B/+; Pk61Cb/+ | M | 126 | 57.29±8.23 | 59 | --------- | --------- | --------- |
| L | w/Y; 255B/+; Pk61Cb/+ | M | 24 | 48.08±10.99 | 51 | -16.06 | -13.56 | 4.11E-12 | |
| - | w; 255B/+; Pk61Cb/+ | F | 124 | 54.51±9.89 | 56.5 | --------- | --------- | --------- | |
| L | w; 255B/+; Pk61Cb/+ | F | 121 | 45.28±13.38 | 51 | -16.93 | -9.73 | 1.79E-14 | |
| 19-1 | - | w/Y; 255B/+; DIAP1/+ | M | 120 | 56.23±8.68 | 59 | --------- | --------- | --------- |
| L | w/Y; 255B/+; DIAP1/+ | M | 93 | 47.68±17.07 | 53 | -15.22 | -10.17 | 0.002 | |
| - | w/w*; 255B/+; DIAP1/+ | F | 120 | 54.64±11.2 | 57.5 | --------- | --------- | --------- | |
| L | w/w*; 255B/+; DIAP1/+ | F | 117 | 46.74±14.76 | 51 | -14.45 | -11.3 | 1.97E-07 | |
| 7-1 | - | w/Y; 255B/+; + | M | 124 | 52.97±7.68 | 56 | --------- | --------- | --------- |
| A | w/Y; 255B/+; + | M | 124 | 49.73±4.6 | 50 | -6.11 | -10.71 | 6.19E-13 | |
| L | w/Y; 255B/+; + | M | 22 | 44.5±16.86 | 52 | -15.99 | -7.14 | 2.94E-05 | |
| - | w/+; 255B/+; + | F | 118 | 58.39±5.25 | 59 | --------- | --------- | --------- | |
| A | w/+; 255B/+; + | F | 122 | 52.66±4.09 | 52 | -9.81 | -11.86 | 0 | |
| L | w/+; 255B/+; + | F | 122 | 50.45±9.78 | 53.5 | -13.6 | -9.32 | 1.18E-13 | |
| Ex 2 Life span assay using GS255B driver at 25C | |||||||||
| 7-1 | - | w/Y; 255B/+; + | M | 94 | 73.17±15.64 | 78 | --------- | --------- | --------- |
| A | w/Y; 255B/+; + | M | 93 | 69.97±12.33 | 72 | -4.38 | -7.69 | 5.22E-04 | |
| - | w/+; 255B/+; + | F | 92 | 87.2±18.44 | 92 | --------- | --------- | --------- | |
| A | w/+; 255B/+; + | F | 91 | 91.93±7.76 | 94 | 5.43 | 2.17 | 0.940 | |
| 20-1 | - | w/Y; 255B/+; nmo/+ | M | 95 | 66.74±16.11 | 68 | --------- | --------- | --------- |
| A | w/Y; 255B/+; nmo/+ | M | 90 | 64.66±14.3 | 66 | -3.12 | -2.94 | 0.102 | |
| - | w/yw; 255B/+; nmo/+ | F | 97 | 67.59±28.66 | 74 | --------- | --------- | --------- | |
| A | w/yw; 255B/+; nmo/+ | F | 95 | 68.79±30.17 | 80 | 1.78 | 8.11 | 0.878 | |
| 15-1 | - | w/Y; 255B/wga; + | M | 96 | 72.88±10.31 | 74 | --------- | --------- | --------- |
| A | w/Y; 255B/wga; + | M | 92 | 53.14±18.01 | 56 | -27.08 | -24.32 | 0 | |
| - | w/w*; 255B/wga; + | F | 97 | 78.89±19.38 | 84 | --------- | --------- | --------- | |
| A | w/w*; 255B/wga; + | F | 97 | 53.72±22.13 | 52 | -31.9 | -38.1 | 0 | |
| 17-1 | - | w/Y; 255B/+; Pk61Ca/+ | M | 91 | 64.11±13.4 | 64 | --------- | --------- | --------- |
| A | w/Y; 255B/+; Pk61Ca/+ | M | 94 | 62.85±13.08 | 66 | -1.96 | 3.13 | 0.555 | |
| - | w/yw; 255B/+; Pk61Ca/+ | F | 98 | 70.73±26.23 | 78 | --------- | --------- | --------- | |
| A | w/yw; 255B/+; Pk61Ca/+ | F | 94 | 79.81±23.77 | 90 | 12.83 | 15.38 | 0.149 | |
| 21-1 | - | w/Y; 255B/+; Egfr/+ | M | 89 | 62.38±11.19 | 66 | --------- | --------- | --------- |
| A | w/Y; 255B/+; Egfr/+ | M | 97 | 62.06±9.18 | 64 | -0.51 | -3.03 | 0.166 | |
| - | w/y w*; 255B/+; Egfr/+ | F | 95 | 65.71±21.16 | 68 | --------- | --------- | --------- | |
| A | w/y w*; 255B/+; Egfr/+ | F | 100 | 63.52±17.9 | 65 | -3.33 | -4.41 | 0.076 | |
| 19-1 | - | w/Y; 255B/+; DIAP1/+ | M | 102 | 76.57±13.04 | 78 | --------- | --------- | --------- |
| A | w/Y; 255B/+; DIAP1/+ | M | 94 | 73.4±9.53 | 74 | -4.13 | -5.13 | 0.002 | |
| - | w/w*; 255B/+; DIAP1/+ | F | 98 | 78.9±18.26 | 84 | --------- | --------- | --------- | |
| A | w/w*; 255B/+; DIAP1/+ | F | 95 | 81.39±19.17 | 88 | 3.16 | 4.76 | 0.011 | |
| 22-1 | - | w/Y; 255B/+; pnt/+ | M | 96 | 62.6±9.74 | 64 | --------- | --------- | --------- |
| A | w/Y; 255B/+; pnt/+ | M | 94 | 59.15±10.7 | 60 | -5.52 | -6.25 | 0.077 | |
| - | w/yw; 255B/+; pnt/+ | F | 92 | 74.32±27.19 | 85 | --------- | --------- | --------- | |
| A | w/yw; 255B/+; pnt/+ | F | 95 | 79.77±20.03 | 88 | 7.34 | 3.53 | 0.402 | |
| Exp3 Life span assay using GS255B driver at 25C | |||||||||
| 7-1 | - | w/Y; 255B/+; + | M | 100 | 81.01±15.38 | 86 | --------- | --------- | --------- |
| A | w/Y; 255B/+; + | M | 92 | 80.46±10.42 | 82 | -0.68 | -4.65 | 0.039 | |
| - | w/+; 255B/+; + | F | 85 | 92.49±11.86 | 94 | --------- | --------- | --------- | |
| A | w/+; 255B/+; + | F | 99 | 92.05±13.07 | 94 | -0.48 | 0 | 0.571 | |
| 13-1 | - | w/Y; 255B/Ras WT;+ | M | 95 | 75.87±12.26 | 78 | --------- | --------- | --------- |
| A | w/Y; 255B/Ras WT;+ | M | 98 | 67.92±14.13 | 70 | -10.48 | -10.26 | 1.62E-06 | |
| - | w/w*; 255B/Ras WT;+ | F | 96 | 83.43±12.26 | 86 | --------- | --------- | --------- | |
| A | w/w*; 255B/Ras WT;+ | F | 98 | 79.59±10.58 | 82 | -4.6 | -4.65 | 0.001 | |
| 23-1 | - | w/Y; 255B/Mmp2; + | M | 96 | 69.77±13.03 | 70 | --------- | --------- | --------- |
| A | w/Y; 255B/Mmp2; + | M | 96 | 68.81±10.06 | 70 | -1.37 | 0 | 0.117 | |
| - | w/yw; 255B/Mmp2; + | F | 98 | 85.94±18.45 | 91 | --------- | --------- | --------- | |
| A | w/yw; 255B/Mmp2; + | F | 101 | 84.85±18.67 | 90 | -1.27 | -1.1 | 0.109 | |
| 24-1 | - | w/Y; 255B/+; Stat/+ | M | 96 | 64.31±10.06 | 65 | --------- | --------- | --------- |
| A | w/Y; 255B/+; Stat/+ | M | 99 | 65.08±13.33 | 68 | 1.19 | 4.62 | 0.325 | |
| - | w/yw; 255B/+; Stat/+ | F | 99 | 70.48±21.48 | 78 | --------- | --------- | --------- | |
| A | w/yw; 255B/+; Stat/+ | F | 96 | 62.29±24.99 | 74 | -11.62 | -5.13 | 0.076 | |
| 25-1 | - | w/Y; 255B/+; puc/+ | M | 97 | 70.78±14.98 | 70 | --------- | --------- | --------- |
| A | w/Y; 255B/+; puc/+ | M | 96 | 68.96±13.62 | 68 | -2.58 | -2.86 | 0.269 | |
| - | w/w*; 255B/+; puc/+ | F | 84 | 94.07±15.00 | 98 | --------- | --------- | --------- | |
| A | w/w*; 255B/+; puc/+ | F | 97 | 98.1±7.86 | 100 | 4.29 | 2.04 | 0.135 | |
| Exp4 Life span assay using GS255B driver at 25C | |||||||||
| 7-1 | - | w/Y; 255B/+; + | M | 92 | 73.76±18.31 | 78 | --------- | --------- | --------- |
| A | w/Y; 255B/+; + | M | 86 | 71.84±10.87 | 74 | -2.61 | -5.13 | 1.43E-04 | |
| - | w/+; 255B/+; + | F | 86 | 86.28±15.46 | 90 | --------- | --------- | --------- | |
| A | w/+; 255B/+; + | F | 101 | 86.18±10.02 | 88 | -0.12 | -2.22 | 0.035 | |
| 26-1 | - | w/Y; 255B/+; Sk2/+ | M | 90 | 67.22±16.97 | 72 | --------- | --------- | --------- |
| A | w/Y; 255B/+; Sk2/+ | M | 95 | 69.85±12.26 | 72 | 3.91 | 0 | 0.953 | |
| - | w/yw; 255B/+; Sk2/+ | F | 101 | 73.29±24.6 | 84 | --------- | --------- | --------- | |
| A | w/yw; 255B/+; Sk2/+ | F | 106 | 78.08±19.61 | 86 | 6.53 | 2.38 | 0.84 | |
| 27-1 | - | w/Y; 255B/+; ban/+ | M | 98 | 66.59±21.91 | 70 | --------- | --------- | --------- |
| A | w/Y; 255B/+; ban/+ | M | 95 | 61.56±18.15 | 62 | -7.56 | -11.43 | 0.003 | |
| - | w/yw; 255B/+; ban/+ | F | 94 | 76.36±28.78 | 88 | --------- | --------- | --------- | |
| A | w/yw; 255B/+; ban/+ | F | 96 | 81.56±18.03 | 88 | 6.81 | 0 | 0.023 | |
| 28-1 | - | w/Y; 255B/+;CG14544/+ | M | 91 | 75.03±12.8 | 76 | --------- | --------- | --------- |
| A | w/Y; 255B/+;CG14544/+ | M | 97 | 77.01±9.82 | 78 | 2.64 | 2.63 | 0.844 | |
| - | w/w; 255B/+;CG14544/+ | F | 101 | 70.99±26.75 | 82 | --------- | --------- | --------- | |
| A | w/w; 255B/+;CG14544/+ | F | 96 | 69.33±20.57 | 79 | -2.33 | -3.66 | 2.28E-05 | |
| Exp5 Life span assay using GS255B driver, and MHC GS driver at 25C | |||||||||
| 7-1 | - | w/Y; 255B/+; + | M | 98 | 75.06±11.65 | 79 | --------- | --------- | --------- |
| A | w/Y; 255B/+; + | M | 97 | 73.96±12.27 | 78 | -1.47 | -1.27 | 0.161 | |
| - | w/+; 255B/+; + | F | 100 | 87.88±7.74 | 88 | --------- | --------- | --------- | |
| A | w/+; 255B/+; + | F | 101 | 85.33±12.78 | 88 | -2.91 | 0 | 0.014 | |
| 8-1 | - | w/Y; 255B/+; + | M | 99 | 66.55±11.82 | 68 | --------- | --------- | --------- |
| A | w/Y; 255B/+; + | M | 97 | 69.84±10.57 | 72 | 4.94 | 5.88 | 0.14 | |
| - | w/w; 255B/+; + | F | 100 | 79.6±14.08 | 84 | --------- | --------- | --------- | |
| A | w/w; 255B/+; + | F | 97 | 81.69±3.82 | 82 | 2.63 | -2.38 | 0.005 | |
| 17-1 | - | w/Y; 255B/+; Pk61Ca/+ | M | 99 | 64.87±12.41 | 64 | --------- | --------- | --------- |
| A | w/Y; 255B/+; Pk61Ca/+ | M | 98 | 62.16±12.33 | 64 | -4.17 | 0 | 0.113 | |
| - | w/yw; 255B/+; Pk61Ca/+ | F | 98 | 81.96±13.91 | 86 | --------- | --------- | --------- | |
| A | w/yw; 255B/+; Pk61Ca/+ | F | 99 | 80.46±11.15 | 82 | -1.82 | -4.65 | 0.001 | |
| 18-1 | - | w/Y; 255B/+; Pk61Cb/+ | M | 100 | 71.2±9.32 | 74 | --------- | --------- | --------- |
| A | w/Y; 255B/+; Pk61Cb/+ | M | 101 | 73.33±8.19 | 74 | 2.99 | 0 | 0.135 | |
| - | w; 255B/+; Pk61Cb/+ | F | 98 | 80.27±12.63 | 82 | --------- | --------- | --------- | |
| A | w; 255B/+; Pk61Cb/+ | F | 100 | 82.02±4.89 | 82 | 2.19 | 0 | 0.399 | |
| 19-1 | - | w/Y; 255B/+; DIAP1/+ | M | 101 | 72.97±11.13 | 76 | --------- | --------- | --------- |
| A | w/Y; 255B/+; DIAP1/+ | M | 101 | 71.15±11.37 | 74 | -2.5 | -2.63 | 0.425 | |
| - | w/w*; 255B/+; DIAP1/+ | F | 98 | 83.02±7.52 | 84 | --------- | --------- | --------- | |
| A | w/w*; 255B/+; DIAP1/+ | F | 106 | 79.47±12.78 | 82 | -4.27 | -2.38 | 0.011 | |
| 7-6 | - | yw/Y;+/CyO; MHC/+ | M | 96 | 71.31±13.18 | 76 | --------- | --------- | --------- |
| A | yw/Y;+/CyO; MHC/+ | M | 98 | 58.92±12.84 | 60 | -17.38 | -21.05 | 0 | |
| - | yw/+;+/CyO; MHC/+ | F | 96 | 78.04±15.76 | 84 | --------- | --------- | --------- | |
| A | yw/+;+/CyO; MHC/+ | F | 114 | 52.84±13.75 | 58 | -32.29 | -30.95 | 0 | |
| 8-6 | - | yw/Y;+/CyO; MHC/+ | M | 99 | 56.42±16.69 | 60 | --------- | --------- | --------- |
| A | yw/Y;+/CyO; MHC/+ | M | 92 | 43.26±15 | 48 | -23.33 | -20 | 1.61E-12 | |
| - | yw/w;+/CyO; MHC/+ | F | 93 | 60.73±18.82 | 68 | --------- | --------- | --------- | |
| A | yw/w;+/CyO; MHC/+ | F | 99 | 45.21±13.63 | 50 | -25.55 | -26.47 | 2.22E-16 | |
| 17-6 | - | yw/Y;+/CyO; MHC/Pk61Ca | M | 100 | 54.32±14.66 | 58 | --------- | --------- | --------- |
| A | yw/Y;+/CyO; MHC/Pk61Ca | M | 101 | 52.46±12.42 | 56 | -3.43 | -3.45 | 0.004 | |
| - | yw;+/CyO; MHC/Pk61Ca | F | 100 | 61.3±19.84 | 65 | --------- | --------- | --------- | |
| A | yw;+/CyO; MHC/Pk61Ca | F | 99 | 39.37±14.22 | 36 | -35.77 | -44.62 | 0 | |
| 18-6 | - | yw/Y;+/CyO; MHC/Pk61Cb | M | 96 | 56.44±18.89 | 62 | --------- | --------- | --------- |
| A | yw/Y;+/CyO; MHC/Pk61Cb | M | 108 | 53.56±10.21 | 56 | -5.11 | -9.68 | 1.75E-07 | |
| - | yw/w;+/CyO; MHC/Pk61Cb | F | 96 | 55.25±21.21 | 56 | --------- | --------- | --------- | |
| A | yw/w;+/CyO; MHC/Pk61Cb | F | 90 | 39.42±12.45 | 38 | -28.65 | -32.14 | 1.40E-11 | |
| 19-6 | - | yw/Y;+/CyO; MHC/DIAP1 | M | 98 | 66.71±12.27 | 68 | --------- | --------- | --------- |
| A | yw/Y;+/CyO; MHC/DIAP1 | M | 98 | 54.92±10.4 | 56 | -17.68 | -17.65 | 6.66E-15 | |
| - | yw/w*;+/CyO;MHC/DIAP1 | F | 108 | 64.89±18.19 | 71 | --------- | --------- | --------- | |
| A | yw/w*;+/CyO;MHC/DIAP1 | F | 104 | 57.04±14.85 | 58 | -12.1 | -18.31 | 6.72E-08 | |
| 7-6 | - | yw/Y;+/Sp; MHC/+ | M | 99 | 68.67±13.38 | 74 | --------- | --------- | --------- |
| A | yw/Y;+/Sp; MHC/+ | M | 98 | 61.76±12.62 | 64 | -10.07 | -13.51 | 5.34E-08 | |
| - | yw/+;+/Sp; MHC/+ | F | 94 | 73.66±13.99 | 78 | --------- | --------- | --------- | |
| A | yw/+;+/Sp; MHC/+ | F | 102 | 59.98±8.45 | 62 | -18.57 | -20.51 | 0 | |
| 8-6 | - | yw/Y;+/Sp; MHC/+ | M | 96 | 61.6±14.81 | 66 | --------- | --------- | --------- |
| A | yw/Y;+/Sp; MHC/+ | M | 94 | 54.55±15.5 | 56 | -11.45 | -15.15 | 2.47E-06 | |
| - | yw/w;+/Sp; MHC/+ | F | 93 | 58.41±17.34 | 62 | --------- | --------- | --------- | |
| A | yw/w;+/Sp; MHC/+ | F | 102 | 47.59±12.97 | 52 | -18.53 | -16.13 | 2.70E-14 | |
| 17-6 | - | yw/Y;+/Sp; MHC/Pk61Ca | M | 95 | 49.31±12.75 | 54 | --------- | --------- | --------- |
| A | yw/Y;+/Sp; MHC/Pk61Ca | M | 100 | 45.14±11.03 | 48 | -8.45 | -11.11 | 1.95E-04 | |
| - | yw;+/Sp; MHC/Pk61Ca | F | 99 | 42.99±20.17 | 40 | --------- | --------- | --------- | |
| A | yw;+/Sp; MHC/Pk61Ca | F | 100 | 35.78±16.13 | 30 | -16.77 | -25 | 0.003 | |
| 18-6 | - | yw/Y;+/Sp; MHC/Pk61Cb | M | 100 | 56.36±12.56 | 60 | --------- | --------- | --------- |
| A | yw/Y;+/Sp; MHC/Pk61Cb | M | 98 | 51.06±9.77 | 52 | -9.4 | -13.33 | 2.13E-06 | |
| - | yw/w;+/Sp; MHC/Pk61Cb | F | 97 | 56.6±16.25 | 60 | --------- | --------- | --------- | |
| A | yw/w;+/Sp; MHC/Pk61Cb | F | 94 | 41.79±11.81 | 42 | -26.17 | -30 | 1.67E-15 | |
| 19-6 | - | yw/Y;+/Sp; MHC/DIAP1 | M | 99 | 61.21±10.04 | 62 | --------- | --------- | --------- |
| A | yw/Y;+/Sp; MHC/DIAP1 | M | 97 | 55.51±9.17 | 58 | -9.32 | -6.45 | 5.02E-08 | |
| - | yw/w*;+/Sp; MHC/DIAP1 | F | 103 | 71.13±17.02 | 78 | --------- | --------- | --------- | |
| A | yw/w*;+/Sp; MHC/DIAP1 | F | 101 | 62.3±13.23 | 66 | -12.41 | -15.38 | 6.66E-16 | |
Table 3. Life span data for baculovirus p35 experiments, with means, standard deviations, medians, percent change in mean and median, and log rank p value.
| CrossMxF | RU486 | Genotype | Sex | N | Meana | Median | %Change in Mean | %Change in Median | Log Rank p Value |
| Exp1 Life span assay of three UAS-p35 lines and UAS-p53.259H with GS255B driver at 25C | |||||||||
| 7-1 | - | w/Y; 255B/+; + | M | 120 | 84.6±14.25 | 90 | --------- | --------- | --------- |
| A | w/Y; 255B/+; + | M | 119 | 83.08±10.94 | 86 | -1.8 | -4.44 | 0.014 | |
| L | w/Y; 255B/+; + | M | 123 | 78.44±22.48 | 86 | -7.28 | -4.44 | 0.244 | |
| - | w/+; 255B/+; + | F | 116 | 92.02±9.64 | 94 | --------- | --------- | --------- | |
| A | w/+; 255B/+; + | F | 121 | 94.69±8.61 | 94 | 2.9 | 0 | 0.009 | |
| L | w/+; 255B/+; + | F | 124 | 91.97±15.74 | 94 | -0.05 | 0 | 0.047 | |
| 1-9 | - | p35,w*/Y; 255B/+; + | M | 120 | 68.93±12.49 | 70 | --------- | --------- | --------- |
| A | p35,w*/Y; 255B/+; + | M | 123 | 68.62±11.76 | 70 | -0.45 | 0 | 0.681 | |
| L | p35,w*/Y; 255B/+; + | M | 98 | 33.1±22.91 | 26 | -51.98 | -62.86 | 0 | |
| - | p35,w*/+; 255B/+; + | F | 122 | 83.28±15.13 | 86 | --------- | --------- | --------- | |
| A | p35,w*/+; 255B/+; + | F | 130 | 77.15±20.92 | 82 | -7.36 | -4.65 | 0.11 | |
| L | p35,w*/+; 255B/+; + | F | 125 | 57.52±35.8 | 70 | -30.93 | -18.6 | 0.001 | |
| 10-1 | - | w/Y; 255B/p35; + | M | 117 | 54.48±13 | 54 | --------- | --------- | --------- |
| A | w/Y; 255B/p35; + | M | 121 | 57.26±8.45 | 58 | 5.1 | 7.41 | 0.583 | |
| L | w/Y; 255B/p35; + | M | 110 | 34.25±19.5 | 34 | -37.13 | -37.04 | 5.60E-10 | |
| - | w/w*; 255B/p35; + | F | 120 | 64.05±14.63 | 66 | --------- | --------- | --------- | |
| A | w/w*; 255B/p35; + | F | 126 | 60.79±16.68 | 66 | -5.09 | 0 | 0.188 | |
| L | w/w*; 255B/p35; + | F | 123 | 49.37±32.2 | 54 | -22.92 | -18.18 | 0.436 | |
| 11-1 | - | w/Y; 255B/+; p35/+ | M | 133 | 86.03±12.51 | 90 | --------- | --------- | --------- |
| A | w/Y; 255B/+; p35/+ | M | 122 | 81.31±13.87 | 84 | -5.49 | -6.67 | 8.92E-06 | |
| L | w/Y; 255B/+; p35/+ | M | 56 | 46.18±24.21 | 44 | -46.32 | -51.11 | 0 | |
| - | w/w*; 255B/+; p35/+ | F | 126 | 87.54±10.04 | 90 | --------- | --------- | --------- | |
| A | w/w*; 255B/+; p35/+ | F | 127 | 82.19±9.91 | 82 | -6.11 | -8.89 | 8.60E-05 | |
| L | w/w*; 255B/+; p35/+ | F | 126 | 64.63±29.62 | 75 | -26.17 | -16.67 | 4.67E-07 | |
| 29-1 | - | w/Y; 255B/+;p53.259H/+ | M | 118 | 71.54±13.86 | 72 | --------- | --------- | --------- |
| A | w/Y; 255B/+;p53.259H/+ | M | 125 | 68.90±10.41 | 70 | -3.70 | -2.78 | 0.002 | |
| L | w/Y; 255B/+;p53.259H/+ | M | 119 | 67.73±16.92 | 70 | -5.33 | -2.78 | 0.069 | |
| - | w; 255B/+; p53.259H/+ | F | 119 | 75.40±8.50 | 76 | --------- | --------- | --------- | |
| A | w; 255B/+; p53.259H/+ | F | 119 | 80.66±10.98 | 82 | 6.98 | 7.89 | 4.05E-08 | |
| L | w; 255B/+; p53.259H/+ | F | 125 | 70.24±22.02 | 76 | -6.84 | 0 | 0.202 | |
| Exp2 Life span assay of three UAS-p35 lines with head FB driver, whole body FB driver and GS255A driver at 25C | |||||||||
| 3-7 | - | +/Y; S32/+; + | M | 75 | 59.23±14.11 | 64 | --------- | --------- | --------- |
| A | +/Y; S32/+; + | M | 63 | 55.21±15.74 | 60 | -6.79 | -6.25 | 0.013 | |
| - | w/+; S32/+; + | F | 111 | 59.91±18.96 | 60 | --------- | --------- | --------- | |
| A | w/+; S32/+; + | F | 115 | 63.77±17.71 | 66 | 6.45 | 10 | 0.263 | |
| 3-9 | - | p35,w*/Y; S32/+; + | M | 122 | 62.69±10.62 | 64 | --------- | --------- | --------- |
| A | p35,w*/Y; S32/+; + | M | 105 | 58.3±13.45 | 60 | -6.99 | -6.25 | 0.022 | |
| - | p35,w*/w; S32/+; + | F | 112 | 59.95±25 | 72 | --------- | --------- | --------- | |
| A | p35,w*/w; S32/+; + | F | 108 | 59±24.86 | 68 | -1.58 | -5.56 | 0.974 | |
| 3-10 | - | w*/Y; S32/p35; + | M | 123 | 45.19±7.61 | 46 | --------- | --------- | --------- |
| A | w*/Y; S32/p35; + | M | 120 | 41.52±6.84 | 42 | -8.12 | -8.7 | 1.96E-04 | |
| - | w*/w; S32/p35; + | F | 121 | 61.62±8.71 | 62 | --------- | --------- | --------- | |
| A | w*/w; S32/p35; + | F | 105 | 63.28±10.6 | 66 | 2.69 | 6.45 | 0.036 | |
| 3-11 | - | w*/Y; S32/+; p35/+ | M | 125 | 62.67±12.41 | 64 | --------- | --------- | --------- |
| A | w*/Y; S32/+; p35/+ | M | 125 | 60.78±14.07 | 62 | -3.03 | -3.13 | 0.174 | |
| - | w*/w; S32/+; p35/+ | F | 109 | 68.44±17.09 | 74 | --------- | --------- | --------- | |
| A | w*/w; S32/+; p35/+ | F | 113 | 70.52±18.12 | 76 | 3.04 | 2.7 | 0.043 | |
| 4-7 | - | +/Y; S106 S32/+; + | M | 116 | 54.12±9.89 | 56 | --------- | --------- | --------- |
| A | +/Y; S106 S32/+; + | M | 118 | 52.68±9.77 | 52 | -2.67 | -7.14 | 0.208 | |
| - | w/+; S106 S32/+; + | F | 110 | 58.82±14.95 | 62 | --------- | --------- | --------- | |
| A | w/+; S106 S32/+; + | F | 120 | 58.07±16.45 | 63 | -1.28 | 1.61 | 0.569 | |
| 4-9 | - | p35,w*/Y; S106 S32/+; + | M | 121 | 47.21±8.48 | 46 | --------- | --------- | --------- |
| A | p35,w*/Y; S106 S32/+; + | M | 110 | 47.67±10.28 | 48 | 0.99 | 4.35 | 0.263 | |
| - | p35,w*/w; S106 S32/+; + | F | 119 | 55.18±22.95 | 66 | --------- | --------- | --------- | |
| A | p35,w*/w; S106 S32/+; + | F | 126 | 47.79±24.9 | 62 | -13.38 | -6.06 | 0.01 | |
| 4-10 | - | w*/Y; S106 S32/p35; + | M | 125 | 33.39±4.44 | 34 | --------- | --------- | --------- |
| A | w*/Y; S106 S32/p35; + | M | 125 | 32.3±6.16 | 32 | -3.26 | -5.88 | 0.475 | |
| - | w*/w; S106 S32/p35; + | F | 121 | 49.55±8.14 | 50 | --------- | --------- | --------- | |
| A | w*/w; S106 S32/p35; + | F | 121 | 50.84±8.85 | 50 | 2.6 | 0 | 0.107 | |
| 4-11 | - | w*/Y; S106 S32/+; p35/+ | M | 125 | 47.15±6.81 | 48 | --------- | --------- | --------- |
| A | w*/Y; S106 S32/+; p35/+ | M | 117 | 48.6±8.42 | 48 | 3.07 | 0 | 0.072 | |
| - | w*/w; S106 S32/+; p35/+ | F | 125 | 56.81±13.02 | 60 | --------- | --------- | --------- | |
| A | w*/w; S106 S32/+; p35/+ | F | 116 | 60.69±11.7 | 64 | 6.83 | 6.67 | 0.004 | |
| 2-7 | - | +/Y; 255A/+; + | M | 114 | 66.04±8.95 | 67 | --------- | --------- | --------- |
| A | +/Y; 255A/+; + | M | 117 | 58.97±15.36 | 62 | -10.69 | -7.46 | 1.48E-05 | |
| - | w/+; 255A/+; + | F | 114 | 72.65±13.95 | 78 | --------- | --------- | --------- | |
| A | w/+; 255A/+; + | F | 116 | 75.02±13.19 | 78 | 3.26 | 0 | 0.064 | |
| 2-9 | - | p35,w*/Y; 255A/+; + | M | 111 | 65.98±14.65 | 66 | --------- | --------- | --------- |
| A | p35,w*/Y; 255A/+; + | M | 115 | 59.82±13.31 | 60 | -9.34 | -9.09 | 3.78E-05 | |
| - | p35,w*/w; 255A/+; + | F | 113 | 58.95±20.26 | 64 | --------- | --------- | --------- | |
| A | p35,w*/w; 255A/+; + | F | 117 | 69.21±17.4 | 72 | 17.4 | 12.5 | 1.32E-06 | |
| 2-10 | - | w*/Y;255A/p35; + | M | 113 | 48.98±9.74 | 48 | --------- | --------- | --------- |
| A | w*/Y;255A/p35; + | M | 125 | 47.66±7.19 | 48 | -2.69 | 0 | 0.03 | |
| - | w*/w; 255A/p35; + | F | 115 | 60.57±16.71 | 66 | --------- | --------- | --------- | |
| A | w*/w; 255A/p35; + | F | 118 | 62±17.79 | 70 | 2.35 | 6.06 | 0.052 | |
| 2-11 | - | w*/Y; 255A/+; p35/+ | M | 115 | 63.66±11.4 | 64 | --------- | --------- | --------- |
| A | w*/Y; 255A/+; p35/+ | M | 114 | 64.92±9.41 | 64 | 1.98 | 0 | 0.776 | |
| - | w*/w; 255A/+; p35/+ | F | 120 | 67.05±11.58 | 70 | --------- | --------- | --------- | |
| A | w*/w; 255A/+; p35/+ | F | 120 | 68.75±9.08 | 70 | 2.54 | 0 | 0.41 | |
| Exp3 Life span assay of two UAS-p35 lines with whole body FB driver at 29C | |||||||||
| 7-4 | - | w/Y; S106 S32/+; + | M | 124 | 49.15±12.5 | 54 | --------- | --------- | --------- |
| L | w/Y; S106 S32/+; + | M | 121 | 49.11±10.75 | 52 | -0.08 | -3.7 | 0.655 | |
| - | w/+; S106 S32/+; + | F | 121 | 51.95±10.82 | 54 | --------- | --------- | --------- | |
| L | w/+; S106 S32/+; + | F | 118 | 55.29±10.06 | 60 | 6.42 | 11.11 | 0.029 | |
| 8-4 | - | w/Y; S106 S32/+; + | M | 121 | 47.16±10.27 | 48 | --------- | --------- | --------- |
| L | w/Y; S106 S32/+; + | M | 118 | 42.85±12.89 | 44 | -9.14 | -8.33 | 0.002 | |
| - | w/w; S106 S32/+; + | F | 124 | 50.48±11.91 | 56 | --------- | --------- | --------- | |
| L | w/w; S106 S32/+; + | F | 125 | 51.63±8.47 | 54 | 2.27 | -3.57 | 0.196 | |
| 10-4 | - | w/Y; S106 S32/p35; + | M | 121 | 50.43±6.73 | 52 | --------- | --------- | --------- |
| L | w/Y; S106 S32/p35; + | M | 121 | 46.5±8.28 | 48 | -7.8 | -7.69 | 4.23E-05 | |
| - | w*/w; S106 S32/p35; + | F | 120 | 50.4±12.84 | 56 | --------- | --------- | --------- | |
| L | w*/w; S106 S32/p35; + | F | 129 | 48.57±8.9 | 50 | -3.62 | -10.71 | 1.45E-05 | |
| 11-4 | - | w/Y; S106 S32/+; p35/+ | M | 126 | 44.03±6.5 | 46 | --------- | --------- | --------- |
| L | w/Y; S106 S32/+; p35/+ | M | 122 | 41.92±10.42 | 46 | -4.8 | 0 | 0.208 | |
| - | w*/w; S106 S32/+; p35/+ | F | 122 | 58.03±6.59 | 60 | --------- | --------- | --------- | |
| L | w*/w; S106 S32/+; p35/+ | F | 124 | 54.81±8.05 | 56 | -5.56 | -6.67 | 8.84E-07 | |
| Exp4 Life span assay of two UAS-p35 lines with Elav driver at 29C | |||||||||
| 7-5 | - | yw/Y; +/+; Elav/+ | M | 131 | 53.92±7.15 | 54 | --------- | --------- | --------- |
| A | yw/Y; +/+; Elav/+ | M | 129 | 52.33±8.14 | 53 | -2.95 | -1.85 | 0.083 | |
| L | yw/Y; +/+; Elav/+ | M | 59 | 35.85±10.58 | 38 | -33.51 | -29.63 | 0 | |
| - | yw/+; +/+; Elav/+ | F | 127 | 58.15±7.22 | 60 | --------- | --------- | --------- | |
| A | yw/+; +/+; Elav/+ | F | 129 | 57.11±5.19 | 58 | -1.79 | -3.33 | 0.013 | |
| L | yw/+; +/+; Elav/+ | F | 120 | 43.46±7.94 | 44 | -25.26 | -26.67 | 0 | |
| 8-5 | - | yw/Y; +/+; Elav/+ | M | 126 | 44.08±8.36 | 44.5 | --------- | --------- | --------- |
| A | yw/Y; +/+; Elav/+ | M | 120 | 43.32±8.76 | 45 | -1.73 | 1.12 | 0.186 | |
| L | yw/Y; +/+; Elav/+ | M | 102 | 26.24±8.51 | 26 | -40.48 | -41.57 | 0 | |
| - | yw/w; +/+; Elav/+ | F | 124 | 46.88±9.92 | 50 | --------- | --------- | --------- | |
| A | yw/w; +/+; Elav/+ | F | 124 | 48.13±7.5 | 49.5 | 2.67 | -1 | 0.406 | |
| L | yw/w; +/+; Elav/+ | F | 114 | 34.82±10.34 | 36 | -25.73 | -28 | 0 | |
| 10-5 | - | yw/Y; p35/+; Elav/+ | M | 125 | 42.34±6.38 | 44 | --------- | --------- | --------- |
| A | yw/Y; p35/+; Elav/+ | M | 122 | 43.34±10.34 | 46 | 2.38 | 4.55 | 0.007 | |
| L | yw/Y; p35/+; Elav/+ | M | 9 | 20.89±10.3 | 26 | -50.66 | -40.91 | 0 | |
| - | yw/w*; p35/+; Elav/+ | F | 121 | 49±10.63 | 52 | --------- | --------- | --------- | |
| A | yw/w*; p35/+; Elav/+ | F | 126 | 50.16±6.4 | 51 | 2.36 | -1.92 | 0.014 | |
| L | yw/w*; p35/+; Elav/+ | F | 9 | 28.22±10.27 | 32 | -42.4 | -38.46 | 1.60E-14 | |
| 11-5 | - | yw/Y; +; Elav/p35 | M | 120 | 51.24±10.46 | 54 | --------- | --------- | --------- |
| A | yw/Y; +; Elav/p35 | M | 121 | 48.62±10.1 | 52 | -5.12 | -3.7 | 1.22E-06 | |
| L | yw/Y; +; Elav/p35 | M | 1 | 10±NA | 10 | -80.48 | -81.48 | 5.60E-10 | |
| - | yw/w*; +; Elav/p35 | F | 118 | 56.77±3.89 | 58 | --------- | --------- | --------- | |
| A | yw/w*; +; Elav/p35 | F | 131 | 52.67±5.08 | 54 | -7.22 | -6.9 | 3.51E-13 | |
| L | yw/w*; +; Elav/p35 | F | 0 | NA | NA | --------- | --------- | --------- | |
| Exp5 Life span assay of two UAS-p35 lines with GS255B driver at 25C | |||||||||
| 8-1 | - | w/Y; 255B/+; + | M | 121 | 62.33±18.12 | 68 | --------- | --------- | --------- |
| L | w/Y; 255B/+; + | M | 119 | 62.57±16.22 | 68 | 0.39 | 0 | 0.478 | |
| L1-10 | w/Y; 255B/+; + | M | 120 | 66.02±19.38 | 72 | 5.91 | 5.88 | 7.82E-04 | |
| - | w/w; 255B/+; + | F | 123 | 75.95±9.37 | 78 | --------- | --------- | --------- | |
| L | w/w; 255B/+; + | F | 124 | 69.02±12.88 | 74 | -9.13 | -5.13 | 7.69E-07 | |
| L1-10 | w/w; 255B/+; + | F | 124 | 78.18±9.17 | 80 | 2.93 | 2.56 | 7.84E-04 | |
| 10-1 | - | w/Y; 255B/p35; + | M | 111 | 56.32±25.51 | 66 | --------- | --------- | --------- |
| L | w/Y; 255B/p35; + | M | 4 | 16±21.6 | 7 | -71.59 | -89.39 | 6.47E-05 | |
| L1-10 | w/Y; 255B/p35; + | M | 117 | 58.56±17.95 | 62 | 3.98 | -6.06 | 0.528 | |
| - | w/w*; 255B/p35; + | F | 119 | 68.47±13.26 | 72 | --------- | --------- | --------- | |
| L | w/w*; 255B/p35; + | F | 30 | 27.47±16.58 | 24 | -59.89 | -66.67 | 0 | |
| L1-10 | w/w*; 255B/p35; + | F | 124 | 64.5±16.45 | 70 | -5.8 | -2.78 | 0.757 | |
| 11-1 | - | w/Y; 255B/+; p35/+ | M | 117 | 66.15±9.97 | 68 | --------- | --------- | --------- |
| L | w/Y; 255B/+; p35/+ | M | 1 | 14±NA | 14 | -78.84 | -79.41 | 3.38E-14 | |
| L1-10 | w/Y; 255B/+; p35/+ | M | 123 | 64.98±15.73 | 70 | -1.78 | 2.94 | 0.099 | |
| - | w/w*; 255B/+; p35/+ | F | 123 | 74.41±5.98 | 76 | --------- | --------- | --------- | |
| L | w/w*; 255B/+; p35/+ | F | 0 | NA | NA | --------- | --------- | --------- | |
| L1-10 | w/w*; 255B/+; p35/+ | F | 123 | 74.37±11.95 | 78 | -0.04 | 2.63 | 0.003 | |
| Exp6 Life span assay of two UAS-p35 lines with Elav driver at 25C | |||||||||
| 8-5 | - | yw/Y; +/+; Elav/+ | M | 108 | 61.69±17.95 | 67 | --------- | --------- | --------- |
| L1-10 | yw/Y; +/+; Elav/+ | M | 115 | 54.17±15.55 | 58 | -12.18 | -13.43 | 1.80E-08 | |
| - | yw/w; +/+; Elav/+ | F | 120 | 57.42±14.79 | 64 | --------- | --------- | --------- | |
| L1-10 | yw/w; +/+; Elav/+ | F | 117 | 56.41±10.82 | 58 | -1.75 | -9.38 | 0.004 | |
| 10-5 | - | yw/Y; p35/+; Elav/+ | M | 121 | 46.6±7.2 | 46 | --------- | --------- | --------- |
| L1-10 | yw/Y; p35/+; Elav/+ | M | 115 | 37.81±9.05 | 38 | -18.86 | -17.39 | 7.06E-14 | |
| - | yw/w*; p35/+; Elav/+ | F | 123 | 50.63±15.32 | 54 | --------- | --------- | --------- | |
| L1-10 | yw/w*; p35/+; Elav/+ | F | 121 | 49.19±12.16 | 50 | -2.85 | -7.41 | 0.035 | |
| 11-5 | - | yw/Y; +; Elav/p35 | M | 120 | 54.32±13.04 | 56 | --------- | --------- | --------- |
| L1-10 | yw/Y; +; Elav/p35 | M | 111 | 52.31±10.98 | 52 | -3.7 | -7.14 | 0.037 | |
| - | yw/w*; +; Elav/p35 | F | 123 | 52.7±13.97 | 54 | --------- | --------- | --------- | |
| L1-10 | yw/w*; +; Elav/p35 | F | 118 | 56.63±10.73 | 58 | 7.45 | 7.41 | 0.091 | |
Table 4. Parameters for Gompertz-Makeham model and likelihood ratio test results.
| Parameters | L | - | chi2 | df | p Value | chi2 | df | p Value | ||
| Females | ||||||||||
| one parameter compared at each time | ||||||||||
| p35 (X) | Both a and b are constrained | |||||||||
| a | 5.39 x 10-9 | 3.96 x 10-7 | 1.789 | 1 | 0.181 | |||||
| b | 3.89 x 10-1 | 3.00 x 10-1 | 1.516 | 1 | 0.218 | |||||
| c | 2.08 x 10-2 | 1.63 x 10-3 | 59.967 | 1 | <0.001 | 57.983 | 1 | <0.001 | ||
| p35 (2) | b is constrained | |||||||||
| a | 7.71 x 10-6 | 2.10 x 10-4 | 5.234 | 1 | 0.022 | 50.203 | 1 | <0.001 | ||
| b | 2.41 x 10-1 | 1.92 x 10-1 | 1.700 | 1 | 0.192 | |||||
| c | 2.52 x 10-2 | 1.37 x 10-3 | 50.610 | 1 | <0.001 | 50.154 | 1 | <0.001 | ||
| p35 (3) | Both a and b are constrained | |||||||||
| a | 3.31 x 10-6 | 3.00 x 10-6 | 0.003 | 1 | 0.958 | |||||
| b | 2.50 x 10-1 | 2.46 x 10-1 | 0.009 | 1 | 0.923 | |||||
| c | 1.36 x 10-2 | 1.80 x 10-4 | 46.090 | 1 | <0.001 | 66.787 | <0.001 | |||
Discussion
The tissue and temporal specificity of transgene expression can have significant effects on Drosophila life span, and therefore the ability meaningfully to interpret results depends upon careful characterization of the expression patterns produced by the system chosen to drive transgene expression [17,37]. Here the Geneswitch system driver Act-GS-255B was found to yield tissue-general expression of target transgenes in both larvae and adults, including modulation of expression by titrating the concentration of drug in the food. Some sex-dependent effects on expression were observed with the Geneswitch drivers. For example, Act-GS-255B produced tissue-general expression in both males and females, however females consistently exhibited higher levels of expression than males. Poirier et al. have recently reported that the Geneswitch driver S1-106 (head fat body) is active in adult females but not males [17], and we found a similar result. Poirier et al. also reported that the Elav-GS (nervous system) driver had a female bias, but in our experiments the Elav-GS driver supported similar levels of UAS-GFP expression in males and females. It was particularly striking that while the S1-106 and S1-32 drivers produced abundant target gene expression in adult fat body, they did not support expression in the larval fat body.
For the Elav-GS driver, previous studies have reported pan-neuronal expression in larvae using a UAS-eGFP reporter [19], nervous system-specific expression in adults using a UAS-eGFP reporter [16], and expression in a subset of neurons in brain and ventral nerve cord in adults using a UAS-LacZ reporter [17]. Here, using the UAS-ultraGFP reporter, Elav-GS was found to produce pan-neuronal staining (i.e., expression in all nervous tissue), plus higher-level expression in a subset of neurons, in both larvae and adults, whereas no expression was observed in any tissues other than nervous system in either larvae or adults. In contrast, Poirier et al. reported that the Elav-GS driver produced staining in the digestive system (gut) when it was tested with the UAS-LacZ reporter, and that this signal in gut was not induced by drug [17]. One possible explanation for this difference in results is that the endogenous Drosophilaβ-galactosidase is expressed in subregions of the gut [38], and this could have resulted in a background signal when staining for transgenic LacZ activity. Alternatively, the expression pattern produced by the Elav-GS driver might be affected by culture conditions or genetic background differences.
When the Act-GS-255B ubiquitous driver was used to drive expression of the p53-259H transgene in adult flies, it produced life span extension in females, consistent with previous results using the Elav-GS driver [9], and therefore demonstrating that the Act-GS-255B driver can produce increased life span when combined with an appropriate target gene. Of the fourteen candidate genes tested by over-expression, only a subset caused significant and reproducible effects on life span: wingless and Ras activated form caused negative effects, while baculovirus p35 produced both positive and negative effects depending upon sex and developmental stage for over-expression. Care must be taken when interpreting negative effects on life span, since life span might be decreased due to a novel pathology unrelated to the normal mechanisms modulating life span. However, that said, it is interesting that these particular genes/pathways were identified from among the set of genes tested.
Over-expression of wingless using the tissue-general Act-GS-255B driver was lethal to male and female larvae, and when expressed in adult flies wingless dramatically decreased both male and female life span. In Drosophila, wingless signaling promotes maintenance of the gut stem cells [39,40] and somatic stem cells in the ovary [41]. Interestingly, the wingless homolog Wnt and the Wnt signaling pathway have been implicated in modulating aging-related cellular phenotypes in mammals [42]: Wnt signaling is implicated in tissue homeostasis and the maintenance of adult stem cell populations in younger mammals, while conversely Wnt signaling is implicated in promoting senescence of muscle stem cells in aging mammals [43] Moreover, the Klotho gene appears to function by inhibiting Wnt signaling, and Klotho mutation produces an accelerated aging-like phenotype in mice [44], consistent with a pro-aging effect of the Wnt pathway. Drosophila stem cell populations show defects in replicative homeostasis during aging in the gut [45,46] and gonads [47-50], however it is currently unknown to what extent alterations in stem cell function might limit adult Drosophila life span. It will be of interest to determine if wingless over-expression reduces adult fly life span by disrupting the function of one or more stem cell populations, and to further explore the role of wingless signaling in the maintenance of stem cell populations during Drosophila aging.
Over-expression of Ras activated form during Drosophila larval development was lethal to males and females, and when expressed in adult flies it dramatically decreased both male and female life span. Ras signaling has been found to shorten life span and promote cellular senescence in yeast and mammals [51-56], whereas in contrast Ras signaling is reported to promote longevity in long-lived C. elegans Daf-2 insulin-like receptor mutants [57]. It will be of interest in the future to test in what tissue Ras activated form acts to decrease adult fly life span and to determine if this might result from an induction of cellular senescence.
Over-expression of the caspase inhibitor baculovirus p35 in adult flies using the tissue-general Act-GS-255B driver had little to no effect on life span, using three independent baculovirus p35 transgenes. In addition, over-expression of the caspase inhibitor DIAP1 in adults had no consistent effects on life span. While caution must be exercised in interpreting a negative result, it would tend to suggest that adult fly life span is not limited by a canonical caspase-dependent apoptotic pathway. Relevant to this idea, the apoptotic events in aging rat skeletal muscle are reported to be relatively caspase-independent [6]. When baculovirus p35 was expressed during larval development using the tissue-general Act-GS-255B driver, it caused reduced mean life span in the resultant male and female adult flies, consistent with the requirement for regulated apoptosis in normal fly development. However, the female adults that resulted from tissue-general baculovirus p35 over-expression during development exhibited an unusual bi-phasic survival curve that included a subset of adult females with increased life span. This bi-phasic curve and subset of long-lived females was not observed with nervous-system expression of baculovirus p35 in larvae using the Elav-GS driver, suggesting that nervous-tissue may not be the critical tissue; however, these experiments were confounded by toxic effect of the Elav-GS driver itself in drug-treated larvae. It will be of interest in the future to determine what might be the mechanism by which baculovirus p35 over-expression in larvae produces a subset of females with increased life span, and if it might result from the inhibition of apoptosis in some critical tissue during female development.
Methods
Drosophila strains. All the target transgenes for over-expression (Table 1) were obtained from Bloomington Drosophila Stock Center. The ubiquitous Geneswitch driver lines Act-GS-255B and Act-GS-255A contain multiple copies of a P element construct in which expression of the Geneswitch cDNA is under the control of the tissue-general Actin5C promoter [16]. The UAS-ultraGFP strain contain multiple copies of a UAS-eGFP construct, and its construction and characterization have been recently described [18]. The Geneswitch system drivers Elav-GS, MHC-GS, S1-32 and S1-106 were generously provided by T. Osterwalder and R. Davis [19,20].
Drosophila culture. Drosophila culture and life span assays were performed as described previously [16]. GeneSwitch virgins were used in the crosses with males of other lines, with the exception of strains in which the target transgene for over-expression was on the X chromosome. Life span assays consisted of ~25 flies per vial, and a total 5 vials for each cohort. For survival assays performed at 25oC, flies were transferred to new vials ever other day. For survival assays preformed at 29oC flies were transferred to new vials every other day during the first 30-40 days, and then every day for the remainder of the life span. RU486 (Mifepristone, Sigma) was dissolved in ethanol (100%) to make a stock solution of 3.2mg/ml. For adult feeding, 50ul RU486 stock solution was added to the surface of each vial to produce a final concentration of ~160ug/ml; 50ul ethanol was added to the control vials. For larval feeding, 0.5ml of 3.2mg/ml RU486 stock solution (or the indicated diluted concentration) was added to the surface of each bottle to produce a final concentration of ~160ug/ml (or indicated diluted concentration); 0.5ml ethanol was added to control bottles.
GeneSwitch driver characterization . Adult flies were cultured in vials in the presence and absence of drug for two weeks prior to dissection. Adult male and female flies, head in half, body in half, midgut and hindgut, ovary and testes, were photographed. Larvae at 1st instar, 2nd instar and 3rd instar, as well as 3rd instar dissected tissues (brain, midgut and hindgut, salivary gland, imaginal discs, and fat body) were also photographed. The Leica MZ FLIII fluorescence stereomicroscope together with the SPOT software were used for photographs: The GFP pictures were taken under the fluorescent light with exposure time 4 sec and a gain of 2.
Statistical analysis. Mean, standard deviation, median, percent change in mean, percent change in median, and log rank p value were calculated using R 2.6.2 [58]. Analysis of mortality rate was performed with the WinModest statistical package [59]. In the Gompertz-Makeham model, the increaseof mortality (μx ) with age (x) is expressed as: μx= aebx+c, where the constant a is the initial mortalityrate, b is the rate of exponential increase in mortality, and c is the age-independent mortality. The age specific mortality rate (μx) was calculated using WinModest by binning the days over which deaths were counted (since fly deaths were recorded every other day) such that μx = (-ln(Nx + δx / Nx )) / δx(or Px = Nx + δx / Nx and μx = -1/δx ln(Px )), where Nxis the number of flies alive at day x and δx is the bin size [2]. Parameters (a, b, c) were also calculated based on a likelihood ratio test. The full model (aebx+c) was plotted, and the Gompertz-only component (aebx) was used to build the decomposed survival curves, using μx: μx= aebx, Px= e-μx. For the decomposed survival curves, any value below 0.5% survival was considered to be the final data point.
Acknowledgments
We thank Morris Waskar for constructing the "WB-FB-GS" driver strain, and thank Scott Pletcher for advice on use of WinModest, and Laurent Seroude for helpful comments. This work was supported by grants from the Department of Health and Human Services to JT (AG11833) and to ST (HG02790).
Conflicts of Interest
The authors in this manuscript have no conflict of interest to declare.
References
- 1. Campisi J Senescent cells, tumor suppression, and organismal aging: good citizens, bad neighbors. Cell. 2005; 120: 513 -22. [PubMed] .
- 2. Nystul TG and Spradling AC. Breaking out of the mold: diversity within adult stem cells and their niches. Curr Opin Genet Dev. 2006; 16: 463 -8. [PubMed] .
- 3. Ohlstein B and Spradling A. The adult Drosophila posterior midgut is maintained by pluripotent stem cells. Nature. 2006; 439: 470 -4. [PubMed] .
- 4. Micchelli CA and Perrimon N. Evidence that stem cells reside in the adult Drosophila midgut epithelium. Nature. 2006; 439: 475 -9. [PubMed] .
- 5. Singh SR , Liu W and Hou SX. The adult Drosophila malpighian tubules are maintained by multipotent stem cells. Cell Stem Cell. 2007; 1: 191 -203. [PubMed] .
- 6. Marzetti E , Wohlgemuth SE , Lees HA , Chung HY , Giovannini S and Leeuwenburgh C. Age-related activation of mitochondrial caspase-independent apoptotic signaling in rat gastrocnemius muscle. Mech Ageing Dev. 2008; 129: 542 -9. [PubMed] .
- 7. Bredesen DE Programmed cell death mechanisms in neurological disease. Curr Mol Med. 2008; 8: 173 -86. [PubMed] .
- 8. Zheng J , Edelman SW , Tharmarajah G , Walker DW , Pletcher SD and Seroude L. Differential patterns of apoptosis in response to aging in Drosophila. Proc Natl Acad Sci U S A. 2005; 102: 12083 -8. [PubMed] .
- 9. Bauer JH , Poon PC , Glatt-Deeley H , Abrams JM and Helfand SL. Neuronal expression of p53 dominant-negative proteins in adult Drosophila melanogaster extends life span. Curr Biol. 2005; 15: 2063 -8. [PubMed] .
- 10. Curtis C , Landis GN , Folk D , Wehr NB , Hoe N , Waskar M , Abdueva D , Skvortsov D , Ford D , Luu A , Badrinath A , Levine RL , Bradley TJ , Tavare S and Tower J. Transcriptional profiling of MnSOD-mediated lifespan extension in Drosophila reveals a species-general network of aging and metabolic genes. Genome Biol. 2007; 8: R262 [PubMed] .
- 11. Aigaki T , Seong KH and Matsuo T. Longevity determination genes in Drosophila melanogaster. Mech Ageing Dev. 2002; 123: 1531 -41. [PubMed] .
- 12. Tower J Sex-specific regulation of aging and apoptosis. Mech Ageing Dev. 2006; 127: 705 -18. [PubMed] .
- 13. Ungewitter E and Scrable H. Antagonistic pleiotropy and p53. Mech Ageing Dev. 2009; 130: 10 -7. [PubMed] .
- 14. Bauer JH , Chang C , Morris SN , Hozier S , Andersen S , Waitzman JS and Helfand SL. Expression of dominant-negative Dmp53 in the adult fly brain inhibits insulin signaling. Proc Natl Acad Sci U S A. 2007; 104: 13355 -60. [PubMed] .
- 15. Nicholson L , Singh GK , Osterwalder T , Roman GW , Davis RL and Keshishian H. Spatial and temporal control of gene expression in Drosophila using the inducible GeneSwitch GAL4 system. I. Screen for larval nervous system drivers. Genetics. 2008; 178: 215 -34. [PubMed] .
- 16. Ford D , Hoe N , Landis GN , Tozer K , Luu A , Bhole D , Badrinath A and Tower J. Alteration of Drosophila life span using conditional, tissue-specific expression of transgenes triggered by doxycycline or RU486/Mifepristone. Exp Gerontol. 2007; 42: 483 -97. [PubMed] .
- 17. Poirier L , Shane A , Zheng J and Seroude L. Characterization of the Drosophila gene-switch system in aging studies: a cautionary tale. Aging Cell. 2008; 7: 758 -70. [PubMed] .
- 18. Yang J and Tower J. Expression of hsp22 and hsp70 transgenes is partially predictive of Drosophila survival under normal and stress conditions. J Gerontol: Biol Sci. 2009; In press .
- 19. Osterwalder T , Yoon KS , White BH and Keshishian H. A conditional tissue-specific transgene expression system using inducible GAL4. Proc Natl Acad Sci U S A. 2001; 98: 12596 -601. [PubMed] .
- 20. Roman G , Endo K , Zong L and Davis RL. P[Switch], a system for spatial and temporal control of gene expression in Drosophila melanogaster. Proc Natl Acad Sci U S A. 2001; 98: 12602 -7. [PubMed] .
- 21. Giannakou ME , Goss M , Jacobson J , Vinti G , Leevers SJ and Partridge L. Dynamics of the action of dFOXO on adult mortality in Drosophila. Aging Cell. 2007; 6: 429 -38. [PubMed] .
- 22. Hwangbo DS , Gersham B , Tu MP , Palmer M and Tatar M. Drosophila dFOXO controls lifespan and regulates insulin signalling in brain and fat body. Nature. 2004; 429: 562 -6. [PubMed] .
- 23. Lee T , Feig L and Montell DJ. Two distinct roles for Ras in a developmentally regulated cell migration. Development. 1996; 122: 409 -18. [PubMed] .
- 24. Farnsworth CL and Feig LA. Dominant inhibitory mutations in the Mg(2+)-binding site of RasH prevent its activation by GTP. Mol Cell Biol. 1991; 11: 4822 -9. [PubMed] .
- 25. Clevers H Wnt/beta-catenin signaling in development and disease. Cell. 2006; 127: 469 -80. [PubMed] .
- 26. Osaki M , Oshimura M and Ito H. PI3K-Akt pathway: its functions and alterations in human cancer. Apoptosis. 2004; 9: 667 -76. [PubMed] .
- 27. Srinivasula SM and Ashwell JD. IAPs: what's in a name. Mol Cell. 2008; 30: 123 -35. [PubMed] .
- 28. Cordier F , Grubisha O , Traincard F , Veron M , Delepierre M and Agou F. The zinc finger of NEMO is a functional ubiquitin-binding domain. J Biol Chem. 2008; 284: 2902 -2907. [PubMed] .
- 29. Shilo BZ Regulating the dynamics of EGF receptor signaling in space and time. Development. 2005; 132: 4017 -27. [PubMed] .
- 30. Page-McCaw A Remodeling the model organism: matrix metalloproteinase functions in invertebrates. Semin Cell Dev Biol. 2008; 19: 14 -23. [PubMed] .
- 31. Li WX Canonical and non-canonical JAK-STAT signaling. Trends Cell Biol. 2008; 18: 545 -51. [PubMed] .
- 32. Wang MC , Bohmann D and Jasper H. JNK Signaling Confers Tolerance to Oxidative Stress and Extends Lifespan in Drosophila. Dev Cell. 2003; 5: 811 -6. [PubMed] .
- 33. McEwen DG and Peifer M. Puckered, a Drosophila MAPK phosphatase, ensures cell viability by antagonizing JNK-induced apoptosis. Development. 2005; 132: 3935 -46. [PubMed] .
- 34. Don AS , Martinez-Lamenca C , Webb WR , Proia RL , Roberts E and Rosen H. Essential requirement for sphingosine kinase 2 in a sphingolipid apoptosis pathway activated by FTY720 analogues. J Biol Chem. 2007; 282: 15833 -42. [PubMed] .
- 35. Nolo R , Morrison CM , Tao C , Zhang X and Halder G. The bantam microRNA is a target of the hippo tumor-suppressor pathway. Curr Biol. 2006; 16: 1895 -904. [PubMed] .
- 36. Ren C , Finkel SE and Tower J. Conditional inhibition of autophagy genes in adult Drosophila impairs immunity without compromising longevity. Exp Gerontol. 2009; In press .
- 37. Tower J Transgenic methods for increasing Drosophila life span. Mech Ageing Dev. 2000; 118: 1 -14. [PubMed] .
- 38. Schnetzer JW and Tyler MS. Endogenous beta-galactosidase activity in the larval, pupal, and adult stages of the fruit fly, Drosophila melanogaster, indicates need for caution in lacZ fusion-gene studies. Biol Bull. 1996; 190: 173 -87. [PubMed] .
- 39. Lin G , Xu N and Xi R. Paracrine Wingless signalling controls self-renewal of Drosophila intestinal stem cells. Nature. 2008; 455: 1119 -23. [PubMed] .
- 40. Takashima S , Mkrtchyan M , Younossi-Hartenstein A , Merriam JR and Hartenstein V. The behaviour of Drosophila adult hindgut stem cells is controlled by Wnt and Hh signalling. Nature. 2008; 454: 651 -5. [PubMed] .
- 41. Song X and Xie T. Wingless signaling regulates the maintenance of ovarian somatic stem cells in Drosophila. Development. 2003; 130: 3259 -68. [PubMed] .
- 42. DeCarolis NA , Wharton KA Jr and Eisch AJ. Which way does the Wnt blow? Exploring the duality of canonical Wnt signaling on cellular aging. Bioessays. 2008; 30: 102 -6. [PubMed] .
- 43. Brack AS and Rando TA. Intrinsic changes and extrinsic influences of myogenic stem cell function during aging. Stem Cell Rev. 2007; 3: 226 -37. [PubMed] .
- 44. Liu H , Fergusson MM , Castilho RM , Liu J , Cao L , Chen J , Malide D , Rovira II , Schimel D , Kuo CJ , Gutkind JS , Hwang PM and Finkel T. Augmented Wnt signaling in a mammalian model of accelerated aging. Science. 2007; 317: 803 -6. [PubMed] .
- 45. Biteau B , Hochmuth CE and Jasper H. JNK activity in somatic stem cells causes loss of tissue homeostasis in the aging Drosophila gut. Cell Stem Cell. 2008; 3: 442 -55. [PubMed] .
- 46. Choi NH , Kim JG , Yang DJ , Kim YS and Yoo MA. Age-related changes in Drosophila midgut are associated with PVF2, a PDGF/VEGF-like growth factor. Aging Cell. 2008; 7: 318 -34. [PubMed] .
- 47. Zhao R , Xuan Y , Li X and Xi R. Age-related changes of germline stem cell activity, niche signaling activity and egg production in Drosophila. Aging Cell. 2008; 7: 344 -54. [PubMed] .
- 48. Pan L , Chen S , Weng C , Call G , Zhu D , Tang H , Zhang N and Xie T. Stem cell aging is controlled both intrinsically and extrinsically in the Drosophila ovary. Cell Stem Cell. 2007; 1: 458 -69. [PubMed] .
- 49. Boyle M , Wong C , Rocha M and Jones DL. Decline in self-renewal factors contributes to aging of the stem cell niche in the Drosophila testis. Cell Stem Cell. 2007; 1: 470 -8. [PubMed] .
- 50. Wallenfang MR , Nayak R and DiNardo S. Dynamics of the male germline stem cell population during aging of Drosophila melanogaster. Aging Cell. 2006; 5: 297 -304. [PubMed] .
- 51. Fabrizio P , Gattazzo C , Battistella L , Wei M , Cheng C , McGrew K and Longo VD. Sir2 blocks extreme life-span extension. Cell. 2005; 123: 655 -67. [PubMed] .
- 52. Di Micco R , Fumagalli M , Cicalese A , Piccinin S , Gasparini P , Luise C , Schurra C , Garre M , Nuciforo PG , Bensimon A , Maestro R , Pelicci PG and d'Adda di Fagagna F. Oncogene-induced senescence is a DNA damage response triggered by DNA hyper-replication. Nature. 2006; 444: 638 -42. [PubMed] .
- 53. Bihani T , Chicas A , Lo CP and Lin AW. Dissecting the senescence-like program in tumor cells activated by Ras signaling. J Biol Chem. 2007; 282: 2666 -75. [PubMed] .
- 54. Courtois-Cox S , Genther Williams SM , Reczek EE , Johnson BW , McGillicuddy LT , Johannessen CM , Hollstein PE , MacCollin M and Cichowski K. A negative feedback signaling network underlies oncogene-induced senescence. Cancer Cell. 2006; 10: 459 -72. [PubMed] .
- 55. Hlavata L , Nachin L , Jezek P and Nystrom T. Elevated Ras/protein kinase A activity in Saccharomyces cerevisiae reduces proliferation rate and lifespan by two different reactive oxygen species-dependent routes. Aging Cell. 2008; 7: 148 -57. [PubMed] .
- 56. Longo VD Ras: the other pro-aging pathway. Sci Aging Knowledge Environ. 2004; 2004: pe36 [PubMed] .
- 57. Nanji M , Hopper NA and Gems D. LET-60 RAS modulates effects of insulin/IGF-1 signaling on development and aging in Caenorhabditis elegans. Aging Cell. 2005; 4: 235 -45. [PubMed] .
- 58. R Development Core Team Vienna, Austria R Foundation for Statistical Computing R: A language and environment for statistical computing. 2006; .
- 59. Pletcher SD Model fitting and hypothesis testing for age-specific mortality data. J Evol Biol. 1999; 12: 430 -9. .